

Океанология, 2021, T. 61, № 2, стр. 220-232
Метод модельных оценок экологического риска для экосистем Арктического шельфа различной продуктивности
Н. В. Соловьева *
Институт океанологии им. П.П. Ширшова РАН
Москва, Россия
* E-mail: soloceanic@yandex.ru
Поступила в редакцию 30.06.2020
После доработки 10.08.2020
Принята к публикации 15.08.2020
Аннотация
В работе предложен метод оценки экологического риска, изменяющегося в течение года в соответствие с естественным функционированием морских экосистем и воздействием различных стрессоров. Метод был применен для контрастных по продуктивности экосистем Арктического шельфа. Были проанализированы материалы экспедиционных исследований в Баренцевом, Белом, Чукотском, Карском, Лаптевых, Восточно-Сибирском морях за период 1996–2018 гг. Рассчитана вероятность допустимой интенсивности антропогенного воздействия для трех сценариев антропогенных воздействий с учетом допустимых рисков, принятых при освоении нефтегазовых ресурсов на шельфах. Показано, что определяющим фактором риска в рамках предложенной постановки задачи выступает отклонение значений средней биомассы фитопланктона от средних и критических значений.
ВВЕДЕНИЕ
Необходимость оценок экологического риска для экосистем Арктического шельфа обусловлена перспективой его интенсивного освоения. Происходящие климатические изменения способствуют возрастанию объема доступных биологических ресурсов и упрощению добычи минеральных ресурсов. Вместе с тем, шельфовые экосистемы Арктики значительно отличаются по своим физико-химическим, биологическим и географическим характеристикам, а, следовательно, отклик на климатические и антропогенные воздействия будет различным. Для оценки последствий интенсивного воздействия на морские экосистемы Арктики необходимы количественные методы оценки экологического риска. В настоящей работе представлен метод экоскрининговой оценки риска, основанный на вероятностном подходе и реализованный для арктических шельфовых экосистем различной степени продуктивности. В ряду существующих методов оценки экологического риска предложенный подход имеет ряд существенных преимуществ, связанных с возможностью расчета внутригодовых вариаций риска при различных сочетаниях воздействующих факторов.
Существующие методы анализа и оценки рисков основаны на сетевом анализе ENA (Ecological Network Analysis) [25, 26, 32] и его развитии – методы ERA (Ecological Risk Assessment) [20, 22]. Количественные методы оценки экологического риска (ERA) являются инструментами управления природными ресурсами [21]. Но, как отмечается при практическом применении, методы ERA для морских систем часто дают лишь общие указания о том, как можно решать проблемы в форме стратегий. В региональной оценке экологического риска широко используется относительная оценка риска RRA (Relative Risk Assessment), основанная на весе фактических данных (WOE) [27, 28]. Перечисленные подходы широко варьируются от качественных до количественных и включают качественное сочетание воздействующих факторов, экспертное ранжирование, полуколичественное ранжирование, индексацию уровня риска, составление карт и матриц риска, матриц полезности сети. В качестве источников воздействия и стрессоров учитывают изменения климата, антропогенные воздействия, в том числе разливы нефти, сброс буровых растворов, сточные воды, инвазии. Эти подходы могут быть использованы в ретроспективной оценке, оценке причинно-следственных связей и во всем процессе оценки экологического риска. Методы региональной оценки экологического риска обладают высоким потенциалом для предоставления практической информации лицам, принимающим решения в области окружающей среды, и имеют потенциал стать количественным инструментом принятия управленческих решений, если такая оценка интегрирована с экономикой [21, 25].
К сожалению, широкий спектр методов и усилий, предпринимаемых для создания эффективного инструмента оценки экологического риска, оказывается мало востребованным из-за весьма общих рекомендаций. Модели ограничиваются мгновенным вычислением причинно-следственных связей отдельных факторов и часто игнорируют системные эффекты, поэтому не могут реализовать целостную оценку на экосистемном уровне [21].
Методология оценки экологического риска наиболее активно разрабатывается в отношении аварий с разливами нефти [11]. При этом используется построение матриц экологического риска, отражающих вероятность возникновения опасного события и тяжесть возможных последствий с учетом известных технологий на шельфовых объектах нефтегазодобычи [11]. Использование таких матриц позволяет оценить степень ущерба природным ресурсам шельфа и время восстановления после опасного события, например разлива нефти, сброса бурового раствора и т.д. Этот метод учитывает пространственно-временнóй диапазон процессов, влияющих на риск, но при этом динамика экологического риска в течение года в зависимости от смены сезонов и внутригодовых вариаций параметров сообществ экосистемы остается неучтенной. По сути все имеющиеся к настоящему времени методы, в том числе перечисленные выше, не учитывают внутригодовые изменения экологического риска. Очевидно, что недостаточно рассматривать только кумулятивный эффект стрессоров в условиях статичной экосистемы. Необходимо сочетать динамику стрессоров с динамикой функционирования экосистемы. Последнее приобретает значение при совместном действии климатических, природных неблагоприятных факторов и антропогенного воздействия. Исходя из принципов предосторожности [11], особое внимание следует уделять комбинированному воздействию наиболее неблагоприятных условий на экосистему и прогнозированию последствий их кумулятивного воздействия.
Процессы естественного функционирования различных экосистем Арктического шельфа изучаются на основе данных комплексных экспедиций ИО РАН [17–19] и модельных количественных построений связей между их основными компонентами [2–4, 23]. Результаты таких натурных исследований были использованы в предложенном подходе для оценки экологического риска и вероятности допустимого воздействия для экосистем Арктического шельфа различной продуктивности.
Метод, представленный в настоящей работе, позволяет рассматривать воздействие комбинаций стрессоров различной природы на процесс естественного функционирования морских экосистем, выявлять интервалы повышенного экологического риска и рассчитывать допустимую вероятность антропогенного воздействия на основе вероятностных моделей. Метод основан на применении экокрининговых оценок риска [12, 16, 29, 30] и использовании данных наблюдений. Структура метода предполагает следующую последовательность действий. Оценка экологического риска рассчитывается на основе натурных наблюдений и/или моделирования состояния экосистем. Входные данные модели оценки риска составляются на основе данных натурных наблюдений биомассы организмов или как результаты моделирования с использованием динамических моделей – экосистемный подход [1, 12, 30]. В обоих случаях наблюдаются пики и спады биомассы и численности популяций, которые используются в качестве исходных данных для расчета вероятности экологического риска, значения которого также претерпевают изменения в течение года.
МАТЕРИАЛЫ И МЕТОДЫ. ВЕРОЯТНОСТНАЯ МОДЕЛЬ РИСКА
Предложенный подход к оценке экологического риска, основанный на экоскрининге [16], учитывает допустимый риск. В этом принципиальное отличие от концепций нормирования, стратегия которых состоит в назначении пороговых значений для конкретных переменных: предельно-допустимых концентраций, выбросов и нагрузок. Основное требование в концепции нормирования – не оказывать прямого или косвенного влияния на функционирование организмов и экосистем при значениях пагубных факторов меньше допустимых. Например, при соблюдении концентраций загрязнителей ниже ПДК, состояние экосистемы по критерию загрязнений оценивается как норма. Другими словами, полагается, что экологический риск равен нулю. В отличие от принципов нормирования, учет допустимого риска в предложенном подходе обусловлен тем обстоятельством, что полное устранение риска при освоении Арктического шельфа практически невозможно или экономически неоправданно. Исследования текущего состояния, структурно-функциональных особенностей и прогноз возможных реакций экосистем на хозяйственное освоение Арктического шельфа, а также климатический отклик являются основой для оценки экологического риска с последующей оценкой допустимого воздействия. Важнейшим в экологическом смысле при получении таких оценок является первичность определения величины допустимого риска экосистемы, а не наоборот – допустимой интенсивности антропогенного воздействия.
Критерий экологической безопасности на основе риска имеет вид: $y{}_{ > }^{ \leqslant }{{y}_{p}}$, где y – экологический риск, yp – допустимый риск. Значение допустимого экологического риска лежит в интервале между верхней yh и нижней yl оценками риска: yl ≤ yp≤ yh. Расчеты риска в теории экоскрининга основываются на верхних оценках: ${{y}_{h}}~{}_{ > }^{ \leqslant }{{y}_{p}}$ [16].
Шельфовая экосистема в нормальном режиме внутригодового функционирования может находиться в m состояниях, где $m = \overline {1,M} $. На экосистему действует L внешних негативных воздействий занумерованных индексами $i = \overline {1,L} $. Антропогенное воздействие для упрощения будем далее связывать с состоянием технической системы (например, системы добычи биологических, минеральных и нефтяных ресурсов на шельфе). Учтем для нее k состояний, где k = $\overline {1,K} $. Тогда, согласно неравенствам Буля, имеем:
Функционирование любой арктической шельфовой экосистемы предполагает внутригодовую динамику биомасс и концентраций составляющих ее компонент. Периоды подъемов и спадов биомассы (численности) популяций не совпадают с календарными сезонами. Максимальные значения биомассы для каждого m-го подъема Nmax и m-го спада $N_{{\max }}^{{\text{'}}}$; m = $\overline {1,M} $, где М – количество периодов “подъем–спад” в течение года, а также средние значения, полученные по данным натурных наблюдений, являются входными данными для вероятностной модели риска. Значение М может меняться от года к году под воздействием естественных и антропогенных факторов. Под подъемами и спадами биомассы фитопланктона подразумеваются естественные сезонные максимумы и минимумы, не связанные с аномальным ростом биомассы.
В общем случае для 2М внутригодовых состояний популяции “подъем–спад” при К внутригодовых состояниях технической системы (технологического режима) получена оценка внутригодового риска в следующем виде [16, 29, 30]:
(1)
$\begin{gathered} {{y}_{k}} \leqslant \mathop \sum \limits_{k = 1}^K {{q}_{k}}\mathop \sum \limits_{m = 1}^M {{p}_{{km}}}{{y}_{{km}}} = \\ = \mathop \sum \limits_{k = 1}^K {{q}_{k}}\mathop \sum \limits_{m = 1}^M {{({{p}_{{am}}}{{y}_{{am}}} + p_{{am}}^{{\text{'}}}y_{{am}}^{{\text{'}}})}_{k}} = \mathop \sum \limits_{k = 1}^K {{q}_{k}}\overline {{{y}_{a}}} ,~ \\ \end{gathered} $(2)
$\begin{gathered} \overline {{{y}_{a}}} {\kern 1pt} {\kern 1pt} ~ = \sum\limits_{m = 1}^M {{{{({{p}_{{am}}}{{y}_{{am}}} + p_{{am}}^{{\text{'}}}y_{{am}}^{{\text{'}}})}}_{k}}} , \\ \sum\limits_{m = 1}^M {({{p}_{{am}}} + p_{{am}}^{{\text{'}}}) = 1,\,\,\,\,\sum\limits_{k = 1}^K {{{q}_{k}} = 1,} } \\ \end{gathered} $(3)
${{p}_{{am}}} = \frac{{{{t}_{m}}}}{t},\,\,\,\,p_{{am}}^{{\text{'}}} = \frac{{t_{m}^{{\text{'}}}}}{t},\,\,\,\,\sum\limits_{m = 1}^M {({{t}_{m}} + t_{m}^{{\text{'}}})} = t,$Обобщение соотношений экоскрининга на случай зависимости риска от времени дает следующее [16, 30]:
(4)
${{y}_{a}} \leqslant \left\{ \begin{gathered} \frac{{1 - {{\overline {E\nu } } \mathord{\left/ {\vphantom {{\overline {E\nu } } {{{N}_{{{\text{max}}}}}}}} \right. \kern-0em} {{{N}_{{{\text{max}}}}}}}}}{{{{{\left( {1 - {{{{N}_{{{\text{cr}}}}}} \mathord{\left/ {\vphantom {{{{N}_{{{\text{cr}}}}}} {{{N}_{{{\text{max}}}}}}}} \right. \kern-0em} {{{N}_{{{\text{max}}}}}}}} \right)}}^{2}}}} \hfill \\ {\text{exp}}\left( { - \frac{{{{{\left( {1 - {{{{N}_{{{\text{cr}}}}}} \mathord{\left/ {\vphantom {{{{N}_{{{\text{cr}}}}}} {{{N}_{{{\text{max}}}}}}}} \right. \kern-0em} {{{N}_{{{\text{max}}}}}}}} \right)}}^{2}}}}{{2\left( {1 - {{\overline {E\nu } } \mathord{\left/ {\vphantom {{\overline {E\nu } } {{{N}_{{{\text{max}}}}}}}} \right. \kern-0em} {{{N}_{{{\text{max}}}}}}}} \right)}}} \right). \hfill \\ \end{gathered} \right.$(5)
$\overline {E\nu } = \mathop \sum \limits_1^M {{({{p}_{{am}}}{{E}_{m}}\nu + p_{{am}}^{{\text{'}}}E_{m}^{{\text{'}}}\nu )}_{k}},$Используя соотношения экоскрининга для определения предельно допустимого воздействия на экосистему, получим ее вероятность в следующем виде [16, 30]:
(6)
$Q = \left\{ \begin{gathered} 1,~\,\,\,\,\,{\text{при}}\,\,~{{y}_{k}} \leqslant {{y}_{d}} \hfill \\ \frac{{{{y}_{d}}}}{{{{y}_{k}}}},\,\,{\text{при}}\,\,{{y}_{d}} < {{y}_{k}} < 1 \hfill \\ {{y}_{d}},\,\,\,{\text{при}}\,\,~{{y}_{k}} = 1, \hfill \\ \end{gathered} \right.$Для расчета ya и Q используются значения Nmax, Ncr, pam, $p_{{am}}^{{\text{'}}}$, ${{E}_{m}}\nu $ и $E_{m}^{{\text{'}}}\nu $, полученные на основе данных натурных наблюдений.
ИСПОЛЬЗОВАНИЕ ДАННЫХ НАТУРНЫХ НАБЛЮДЕНИЙ
Освоение ресурсов Арктического шельфа с экосистемами, принципиально различающимися по свойствам гидрофизических, гидрологических, гидрохимических и гидробиологических процессов, требует методов оценки риска, учитывающих эти особенности. Особый интерес представляют оценки риска для осистем с контрастной продуктивностью, интенсивное освоение ресурсов которых может иметь различные последствия. Экспедиционные исследования на шельфе Арктики, проведенные ИО РАН, выявили уникальные особенности как в распределении биомасс и численностей основных компонент различных экосистем, так и в их функционировании. Большая часть Арктического шельфа с первичной продукцией менее 50 мгС/м2 в день относится к ультраолиготрофным районам [17], но в отдельных локальных областях продукция в сотни раз выше. Если в среднем показатели крайне низкие – 8 мгС/м3 в сутки, то диапазон вариаций значений первичной продукции на порядок выше, от 1.12 до 66.31 мгС/м3 в сутки [17]. В результате проведения натурных наблюдений на трансарктическом разрезе протяженность около 6 тыс. км выявлена изменчивость значений биомассы фитопланктона в интервале от 40 до 700 мг/м3, хлорофилла “а” в поверхностном слое – от 0.04 до 2.32 мг/м3 [17]. Установлено различие в механизмах функционирования высокопродуктивных локальных экосистем и преобладающих сильно обедненных в продукционном отношении акваториях Арктического шельфа. В частности, выявлено наличие физического барьера – сильного распреснения поверхностного 5–12-метрового слоя за счет речного стока в краевые арктические моря от Карского до Чукотского [18]. Устойчивость сформированной плотностной стратификации не преодолевается вертикальной сезонной конвекцией, и процесс обогащения фотического слоя биогенными элементами, участвующими в процессе фотосинтеза, не происходит [17]. Этот естественный барьер не ослабляют современные климатические изменения – увеличение безледного периода и прогрев поверхностного слоя воды [17]. Учет таких особенностей функционирования морских экосистем Арктики будет определять разные уровни экологического риска, а следовательно, разные уровни допустимости антропогенного воздействия, что связано с экономическим обеспечением безопасности освоения ресурсов Арктического шельфа. Другими словами, открывается возможность оптимального согласования экологических и экономических требований, которые зачастую на практике вступают в противоречия. Оценка риска по предлагаемой методике дает возможность получить ответ на вопрос о принципиальной допустимости аварий и штатных воздействий определенных масштабов в различных по продуктивности акваториях Арктического шельфа.
Будем рассчитывать оценку экологического риска для экосистем двух контрастных для Арктики типов: высокопродуктивных и низкопродуктивных. Характер формирования биопродуктивности определяется интенсивностью фотосинтетически активной радиации, стратифицированностью водного слоя, притоком биогенных элементов, топографическими параметрами, присутствием льда и другими. Различные географические особенности и гидротермодинамические режимы функционирования экосистем арктического шельфа формируют благоприятные условия (апвеллинг, фронтальные зоны) для высокой биопродуктивности районов Баренцева и Чукотского морей (каньон Барроу) [6, 17]. Низкопродуктивными считаются моря Карское, Лаптевых, Восточно-Сибирское. Оценку риска будем проводить с учетом внутригодовых изменений биомассы фитопланктона как начального звена трофической цепи. Более высокие трофические уровни зависят от состояния и функционирования фитопланктона. Все звенья пищевой цепи связаны друг с другом и имеют непосредственное или опосредованное взаимовлияние. Существенные различия в сезонной динамике биомассы фитопланктона выявлены в ходе экспедиционных наблюдений для высокопродуктивных и низкопродуктивных экосистем шельфа (рис. 1 ) [5, 6, 9]. Для южной акватории Баренцева моря весенний период активности фитопланктона длится с середины марта до начала июня при наибольших значениях численности и биомассы 1–8 млн кл/л и 1–1.7 мг/л соответственно [7]. Истощение запасов биогенных элементов и трофическая активность зоопланктона приводит к снижению биомассы фитопланктона [5]. Летом, с конца июня до конца августа, максимальная численность пелагических водорослей не превышает 20 тыс. кл/л [7]. Осенью с середины сентября (осенний максимум биомассы фитопланктона) до начала ноября численность не превышает 2 тыс. кл/л, к началу декабря – 1 тыс. кл/л и биомасса менее 5 мкг/л [7]. При продвижении в сторону высоких широт осенний максимум практически не присутствует в годовой динамике фитопланктона, которая приобретает однопиковый характер [7]. Стадия покоя для фитопланктонного сообщества этой экосистемы длится с середины ноября до середины марта [7].
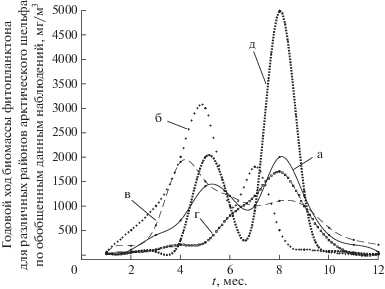
В субарктических прибрежных экосистемах наблюдаются три максимума – ранневесенний, поздневесенний и осенний [7]. Первый весенний максимум приходится на середину апреля с численностью до 2 млн кл/л и биомассой от 1 до 3 мг/л. Второй весенний максимум с конца мая до начала июня обусловлен континентальным стоком. При этом максимально зарегистрированная численность фитопланктона составила 8 млн кл/л, биомасса – 1.7 мг/л. Осенний максимум формируют диатомовые, биомасса которых не превышает 5 мкг/л [7].
Для Белого моря весенний пик фитопланктона приурочен к началу разрушения ледового покрова. В начале мая численность его достигает 2.3–3.8 млн кл/л при биомассе 1.8–1.5 г/м3. К июню численность снижается до сотен клеток на 1 л, а в июле по мере повышения температуры нарастает до 1–2 млн кл/л. Второй летний пик обусловлен сменой доминирующих видов. В отдельные годы для Белого моря были отмечены три максимума биомассы фитопланктона [7].
Интенсивность функционирования фитопланктона Карского моря в отличие от Баренцева сокращается в среднем в два раза и составляет примерно четыре месяца в году [7, 13, 18]. Анализ сезонного хода биомассы фитопланктона [9] и интегральной первичной продукции [4, 10, 23, 24] показали следующее. Весенняя вегетация в прибрежных районах эстуариев Оби и Енисея начинается в конце февраля задолго до таяния ледового покрова [19]. Значения биомассы диатомовых водорослей составляют от 35 до 103 мкг/л и численности – от 4 до 40 тыс. кл/л [9]. Начало массового цветения начинается в конце апреля при смене доминирующих видов [14, 15]. Нарастание биомассы сообщества весной до 1.5 мг/л имеет резкий характер, как и последующий стремительный спад значений [9]. В июле при увеличении температуры, образовании стратифицированного слоя и усилении распреснения достигаются максимальные значения численности 103 тыс. кл/л и биомассы 5 мг/л за весь сезон вегетации [7, 9]. Быстрое понижение температуры в конце сентября–начале октября способствует постепенному убыванию численности фитопланктона до 8 тыс. кл/л при биомассе 50 мкг/л [7, 9]. Годовой минимум активности сообщества приурочен к зимнему периоду с биомассами 5–20 мкг/л.
Из всего многообразия сезонных изменений биомассы фитопланктона в Баренцевом, Белом, Чукотском, Карском, Лаптевых, Восточно-Сибирском морях за период 1996–2018 гг. [2–7, 9, 10, 13–19, 23, 24, 31] (рис. 1) были выбраны два характерных варианта годового цикла: для продуктивных экосистем с двумя максимумами биомассы (рис. 1б) и для низкопродуктивных с одним максимумом (рис. 1г). Для этих контрастных случаев годовых вариаций значений биомассы фитопланктона были проведены расчеты вероятности экологического риска и соответствующие им вероятности допустимого воздействия на экосистему для нескольких сценариев, описанных ниже.
РЕЗУЛЬТАТЫ РАСЧЕТА
Необходимые входные данные для модели риска – значения Nmax, Ncr, pam, $p_{{am}}^{{\text{'}}}$, ${{E}_{m}}\nu $ и $E_{m}^{{\text{'}}}\nu $ могут быть получены как на основе данных натурных наблюдений, так и при расчете по динамическим экосистемным моделям [1, 8, 29, 30]. В нашем случае использовались данные изменения биомассы фитопланктона (рис. 1б, г).
Значения yd в соотношениях (6) выбраны на основе данных экологического состояния, обобщенных оценок техногенных воздействий и экологических последствий на этапах освоения нефтяных ресурсов шельфа [11]. Согласно этим данным, интервал значений вероятности допустимого экологического риска при различных видах и этапах техногенных воздействий на шельфе составляет от 10–7 до 10–1 (табл. 1) [11]. По сути, это вероятности нарушения состояния популяции по показателям отклонения средних значений биомасс за пределы порога нарушений – 10% отклонения [11]. На основании этих данных, для расчета Q выбрано значение yd = 10–5, 10–4, 10–3. Другими словами, это возможная допустимая вероятность сохранения качества экосистемы, сообщества, популяции. Вероятность воздействия на экосистему со стороны технических систем в процессе работ на шельфе, заложенная в технологические процессы, qk выбиралась с учетом зарубежных и отечественных [11] данных. Из диапазона изменения qk от 10–4 до 10–1 [11] были выбраны значения вероятности редкого события q1 = 10–3, события средней частоты q2 = 10–2 и часто происходящего q3 = 10–1. Для аварийных событий при нефтяных разработках на шельфе можно выделить два типа воздействий: 1) кратковременные выбросы загрязняющих веществ в больших объемах (острый стресс для экосистемы) и 2) регулярные, штатные выбросы небольших объемов, осуществляемые в течение длительного времени (хронический стресс). Можно выделить также третью группу неблагоприятных сочетаний воздействий, наложение на регулярные выбросы кратковременных воздействий высокой интенсивности. Используя приведенные выше условия и данные наблюдений, приведенные выше по соотношениям (1)–(6), были проведены оценочные расчеты допустимой вероятности Q воздействия на экосистему для следующих сценариев.
Таблица 1.
Обобщенные оценки антропогенных (техногенных) воздействий и последствий для шельфовых экосистем на основных этапах освоения нефтегазовых ресурсов [11]
| Виды антропогенного воздействия на экосистемы | Масштаб воздействия | Допустимые последствия воздействия | ||
|---|---|---|---|---|
| пространственный | временнóй | обратимость нарушений | ориентировочный допустимый риск | |
| Сейсмическая разведка | Местный | Временный | Обратимые | 10–1 |
| Бурение разведочных скважин | Локальный | Кратковременный | Обратимые | 10–7 |
| Промысловые работы с одиночных платформ | Местный | Временный | Слабообратимые | 10–5 |
| Промысловые региональные работы | Региональный | Долговрененный | Необратимые | 10–2 |
| Строительство платформ, трубопроводов и др. | Локальный | Временный | Обратимые | 10–5–10–7 |
| Эксплуатация трубопроводов в безаварийном режиме | Региональный | Долговременный | Обратимые | 10–5 |
| Танкерные перевозки в безаврийном режиме | Субрегиональный | Временный | Обратимые | 10–7 |
Сценарий А1. Для продуктивной экосистемы арктического шельфа с двумя максимумами биомассы фитопланктона задавались значения Nmax = = 3 × 103 мг/м3, Ncr = 0.5 мг/м3, pam = tm/t = 1/12, значения Emν взяты из данных натурных наблюдений (рис. 1б, 2а). Расчеты внутригодовых вариаций экологического риска и допустимого воздействия были проведены при значениях q1 = = 10–3, q2 = 10–2, q3 = 10–1 и yd = 10–5 (рис. 2 б, в). Результаты показали снижение экологического риска при высоких значениях биомассы планктона и высокие вероятности риска при минимальных значениях биомассы (рис. 2а, б). Соответствующие расчеты для вероятности допустимых воздействий выявили интервалы безопасного воздействия и интервалы с низкими значениями допустимых воздействий (рис. 2в). Анализ полученных результатов показывает, что только при редких воздействиях на экосистему q1 = 10–3 достигается относительно безопасное состояние экосистемы и соответственно высокие значения допустимых воздействий Q1 от 70 до 100% (рис. 2в). Вероятность такого воздействия (при соответствующем риске, рис. 2б) возможна только на короткий промежуток времени, на пике биомассы фитопланктона (рис. 2а, в). Этот участок выделен на рис. 3 и отражает рассчитанную зависимость вероятности допустимого воздействия от экологического риска. В течение остальной части года допустимая вероятность воздействия крайне низкая, менее 30% (рис. 2в, 3). Это обуславливается низкими значениями допустимого экологического риска yd =10–5 (согласно табл. 1).
Рис. 2.
Годовые изменения: (а) – значений биомассы фитопланктона для продуктивных экосистем с двумя максимумами биомассы в мг/м3 по данным наблюдений [6]; (б) – рассчитанных значений вероятности экологического риска и (в) – рассчитанных значений вероятности допустимых воздействий на экосистему при редкой q1, средней q2 и высокой q3 вероятности их осуществления и допустимом значении yd = 10–5.
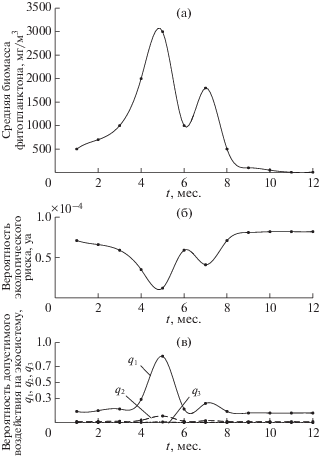
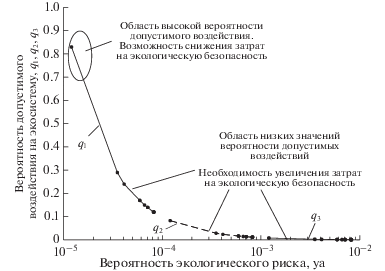
Рис. 3.
Зависимость вероятности допустимого воздействия на высокопродуктивную экосистему от значений экологического риска при допустимом значении yd = 10–5 и редкой q1, средней q2 и высокой q3 вероятности их осуществления.
Сценарий А2. Если для экосистемы (в нашем случае для популяции) возможно понизить жесткость требований по безопасности, т.е. увеличить значения допустимого риска до yd =10–4, то полученные результаты расчетов показывают практически стопроцентную допустимость редких воздействий Q1 и достаточно высокую допустимую вероятность воздействий средней частоты Q2, но только при максимуме биомассы фитопланктона (рис. 4 ). Для воздействий высокой частоты, q3 = = 10–1, вероятность допустимого осуществления воздействия Q3 < 0.1 (меньше 10%). Другими словами, ослабление требований к качеству экосистемы существенно расширяет диапазон допустимых на нее воздействий. На рис. 5 расчет зависимости вероятности допустимого воздействия от экологического риска выявляет области безопасного воздействия на экосистему и области значительного риска, что требует соответствующего увеличения затрат на поддержание безопасности. Для сценария А2 значения биомасс задавались такими же, как в сценарии А1.
Рис. 4.
Годовые изменения рассчитанных значений вероятности допустимых воздействий на высокопродуктивную экосистему при редкой q1, средней q2 и высокой q3 вероятности их осуществления и допустимом значении yd =10–4.
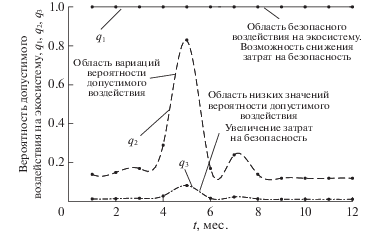
Рис. 5.
Зависимость вероятности допустимого воздействия на высокопродуктивную экосистему от значений экологического риска при допустимом значении yd = 10–4 и редкой q1, средней q2 и высокой q3 вероятности их осуществления.

Сценарий Б. Для низкопродуктивной экосистемы арктического шельфа с одним максимумом биомассы фитопланктона задавались значения Nmax = 1.7 × 103 мг/м3, Ncr = 0.5 мг/м3, pam = = tm/t = 1/12, значения Emν взяты из данных натурных наблюдений (рис. 1г, 6 а). Значение допустимого экологического риска yd = 10–4 принималось таким же, как в сценарии А2 для дальнейшего сравнения полученных результатов. На рис. 6б, в представлены результаты расчета внутригодовых вариаций экологического риска и допустимого воздействия при значениях q1 = 10–3 , q2 = 10–2 , q3 = 10–1. Анализ полученных результатов показывает, что для низкопродуктивной экосистемы шельфа существует возможность безопасного воздействия (Q1 = 100% и Q2, достигающего 100% на коротком интервале внутригодовых вариаций – при максимуме биомассы и минимальном риске, рис. 6б, в). Соответствующие области безопасности и повышенного риска (рис. 7 ) получены при расчете зависимости вероятности допустимого воздействия от значений вероятности экологического риска.
Рис. 6.
Годовые изменения (а) – значений биомассы фитопланктона для низкопродуктивных экосистем с одним максимумом биомассы в мг/м3 по данным наблюдений [6]; (б) – рассчитанных значений вероятности экологического риска и (в) – рассчитанных значений вероятности допустимых воздействий на экосистему при редкой q1, средней q2 и высокой q3 вероятности их осуществления и допустимом значении yd = 10–4.
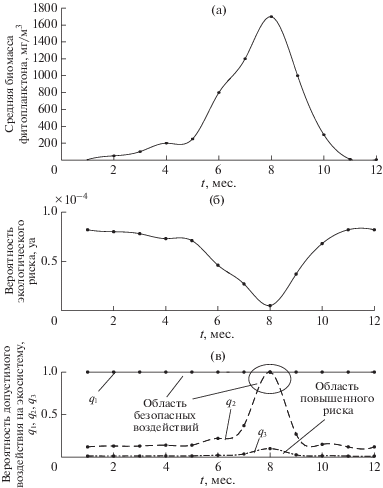
Рис. 7.
Зависимость вероятности допустимого воздействия на низкопродуктивную экосистему от значений экологического риска при допустимом значении yd = 10–4 и редкой q1, средней q2 и высокой q3 вероятности их осуществления.
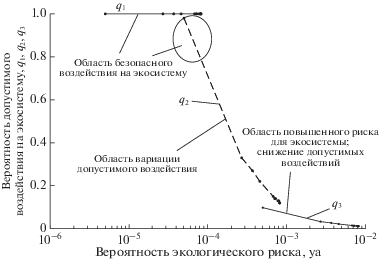
Весьма интересным оказывается результат сравнения расчетов для сценария А2 высокопродуктивной и Б низкопродуктивной экосистем Арктического шельфа. Из сравнения рис. 5 и 7 видно, что при одинаковом допустимом значении экологического риска yd = 10–4 значения вероятности возможного воздействия на экосистемы очень близки, и при редких воздействиях q1 = = 10–3 в обоих случаях достигается практически стопроцентная безопасность, а при воздействиях высокой частоты q3 = 10–1 вероятность допустимости таких воздействий ниже 10% (рис. 5, 7). Это нетривиальный результат: в силу нелинейности процессов в экосистеме и наложения стрессоров сам отклик на воздействия носит существенно нелинейный характер. При ужесточении требований к безопасности (уменьшении значений yd) для редких (q1 = 10–3) и частых (q3 = 10–1) воздействий на экосистемы с различной степенью продуктивности полученные результаты выявили эти нелинейные эффекты (рис. 5, 7).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Эффективное практическое применение указанного подхода требует достаточно полной базы данных натурных наблюдений за компонентами и процессами в исследуемых экосистемах Арктического шельфа, а также построение экосистемных моделей и проведение численных экспериментов с ними при вариации внешних воздействий. Понижение оценки условного риска для групп живых организмов по мере понижения неопределенности ситуации (при проведении необходимых исследований и пополнении баз данных) ведет к уточнению значения допустимой вероятности антропогенных воздействий. Группы живых организмов занимают в экосистеме определенные ступени иерархии и при анализе ее состояния выделяют так называемые ключевые виды, функционирование которых имеет существенное значение для благополучия других [7] и безопасности всей экосистемы в целом. Поэтому предварительные расчеты риска были проведены для сообщества фитопланктона высокопродуктивных и низкопродуктивных экосистем Арктического шельфа.
В настоящей работе не ставилась задача расчета конкретной ситуации для конкретной экосистемы Арктического шельфа, хотя использовались данные конкретных натурных наблюдений значений биомассы фитопланктона для акваторий Арктики различной продуктивности (рис. 1). Целью исследований было показать возможности метода, его реализуемость для ситуаций, возникающих в процессе освоения морских ресурсов Арктики. В проведенных предварительных расчетах для сообщества фитопланктона при описанных сценариях воздействия выявлены сезоны с минимальными и максимальными значениями вероятности экологического риска (рис. 2б, 3–5, 6б, 7). Это позволило рассчитать допустимую вероятность воздействия и выделить интервалы, близкие как к безопасным, так и противоположные – с особо повышенной опасностью воздействия (рис. 2в, 3–5, 6в, 7). При этом результаты расчетов, полученные для высокопродуктивных и низкопродуктивных экосистем Арктического шельфа, выявили весьма близкие значения риска и вероятности допустимого воздействия (ср. рис. 4 и 6в; рис. 5 и 7) при одном и том же типе и продолжительности воздействий (сценарий 2А и Б). Это свидетельствует о том, что определяющим фактором риска в рамках предложенной постановки задачи выступает отклонение значений средней биомассы фитопланктона от значений Nmax и Ncr, с характерными не совпадающими для высокопродуктивных и низкопродуктивных экосистем Арктического шельфа значениями. Другими словами, при приближении к границам устойчивости риск возрастает. Сама величина критического состояния экосистемы не является постоянной. Она нелинейно зависит от внешних природных условий, климатических изменений, антропогенного воздействия и состояния самой экосистемы в текущий и предыдущий моменты времени.
В перспективе рассмотренного подхода представляет интерес рассмотрение риска, обусловленного не только сезонными вариациями состояния экосистем, но и положением продуктивных экосистем, приуроченных к фронтальным зонам. В этом случае преимущественным фактором продуктивности на протяжении всего вегетационного периода являются восходящие движения вод фронтальной области, и можно говорить о пространственно-временнóй неоднородности экологического риска.
Вывести какую-либо закономерность последствий всего многообразия сценариев хозяйственной деятельности человека применительно к шельфовым экосистемам Арктики весьма затруднительно. В то же время необходимо количественно оценивать допустимый уровень конкретных воздействий на шельф с учетом как антропогенной, так и климатической составляющей всего спектра стрессоров. Перечисленные во введении методы оценки риска (ENA, ERA, RRA, WOE) в основном являются качественными или полуколичественными методами. Эти методы не учитывают внутригодовые вариации экологического риска, связанные с естественной внутригодовой динамикой основных компонент экосистемы. Учет этого фактора в предложенном методе приобретает особую важность в связи с тем, что область устойчивости динамических систем, к которым относится экосистема шельфа, не является статически фиксированной, а охватывает изменения в пределах вариаций ее основных параметров. Поэтому в предложенном методе первичным является расчет экологического риска, а затем уже – допустимого воздействия на экосистему.
Допустимая вероятность воздействия на экосистему шельфа предполагает сохранение устойчивости природной системы, ее функционирования, трофической структуры и компонентного состава. Действие стрессоров накладывается на динамику естественного функционирования экосистемы шельфа. Поэтому при оценке экологического риска необходимо учитывать не только совокупность стрессоров различного генезиса, их кумулятивный эффект и режимы воздействия, но и сочетать эти воздействия с динамикой естественного функционирования экологических систем. Внутригодовые колебания основных компонентов экосистем Арктического шельфа в контексте действия стрессоров вызывают колебания степени опасности, а, следовательно, и годовые колебания экологического риска. Предложенный метод позволяет рассматривать воздействие комбинаций стрессоров различной природы на процесс естественного функционирования шельфовых экосистем, выявлять интервалы повышенного экологического риска и рассчитывать приемлемую вероятность антропогенного воздействия на основе вероятностных моделей.
Отметим еще одно из преимуществ предложенного подхода. Его следует рассматривать в рамках вероятности допустимого риска, когда полное устранение риска либо практически невозможно, либо экономически неоправданно. В процессе реализации крупномасштабных проектов освоения морских ресурсов Арктического шельфа (например, разработка нефтяных и газовых месторождений) ошибки в оценке рисков могут привести к серьезным экологическим и экономическим потерям. Так, завышение значения риска, например при принятии невероятно высокой вероятности последствий аварийного случая, может привести к расходованию больших финансовых ресурсов на предотвращение последствий, вероятность которых незначительна. В то же время недооценка риска (опасная ситуация скрыта за внешним благополучием) может привести к катастрофическим последствиям. Наиболее важным вопросом при получении оценок риска, как было показано в проведенных исследованиях, является первичность определения допустимой величины риска для экологических систем, а не наоборот, определение допустимой интенсивности антропогенного воздействия [16]. Это позволяет учесть специфику задач экологической безопасности, заключающуюся в асимметрии требований по устранению ошибок 1-го и 2-го рода (понятия взяты из статистической теории испытаний). Если имеется ошибка 1-го рода, т.е. под безопасной ситуацией понимается ситуация, представляющая угрозу безопасности экосистемы, тогда чрезмерная перестраховка, связанная с ложной тревогой, хотя и приводит к неоправданным затратам, не так опасна, как ошибка 2-го рода, когда опасная ситуация скрыта внешним благополучием.
ВЫВОДЫ
Возможность рассчитывать внутригодовую динамику экологического риска параллельно с внутригодовой динамикой компонент экосистемы шельфа является новым подходом по сравнению с ранжированием и матричными методами оценки риска. Вместе с тем синтез всех имеющихся методов может стать весьма эффективным инструментом в управлении риском. Предложенный метод модельных оценок учитывает перебор возможных сочетаний стрессоров и регламентов воздействий. Метод применим для морских экосистем различных географических условий, контрастных биопродуктивностей и учета широких спектров воздействий. Модельные расчеты позволяют выделять области с практически несущественными последствиями воздействий и критические области с высоким экологическим риском и допустимой вероятностью воздействий менее 5%. Знание таких областей и периодов вариаций риска открывает новые возможности согласования экологических и экономических требований к безопасному освоению ресурсов шельфа. Другими словами, нет необходимости постоянно поддерживать постоянный уровень финансирования экономических мероприятий по обеспечению экологической безопасности. Открывается возможность перераспределения затрат в течение года, позволяющая минимизировать как риски, так и затраты, что сводится к возможности управления риском на основе количественного подхода к оценкам допустимых воздействий.
Благодарности. Автор благодарит сотрудников ИО РАН: акад. РАН Л.И. Лобковского за обсуждение возможных подходов к оценке риска, акад. РАН М.В. Флинта за обсуждение вопросов риска для специфики шельфа Арктики и доктора биологических наук В.А. Силкина за обсуждения по вопросам моделирования.
Источник финансирования. Работа выполнена по Государственному заданию, тема № 00149-2020-0005.
Список литературы
Беляев В.И., Кондуфорова Н.В. Математическое моделирование экологических систем шельфа. Киев: Наук. Думка. 1990. 240 с.
Демидов А.Б., Мошаров С.А. Вертикальное распределение первичной продукции и хлорофилла “а” в Карском море // Океанология. 2015. Т. 55. № 4. С. 577– 591.
Демидов А.Б., Мошаров С.А., Артемьев В.А. и др. Интегрированные и разрешающие по глубине модели первичной продукции Карского моря // Океанология. 2016. Т. 56. № 4. С. 563–576.
Демидов А.Б., Шеберстов С.В., Гагарин В.И. и др. Сезонная изменчивость первичной продукции фитопланктона Карского моря по спутниковым данным // Океанология. 2017. Т. 57. № 14. С. 103– 117.
Дружкова Е.И., Макаревич П.Р. Годовой цикл нанофитопланктона прибрежных вод Баренцева моря. Изв. РАН. Сер. Биологическая. 2008. № 4. С. 497–506.
Ильяш Л.В., Ратькова Т.Н., Радченко И.Г., Житина Л.С. Фитопланктон Белого моря // Система Белого моря. Т. II. Водная толща и взаимодействующие с ней атмосфера, криосфера, речной сток и биосфера. М.: Научный мир. 2012. С. 605–639.
Комплексные исследования больших морских экосистем России / Отв. ред. Матишов Г.Г. Апатиты: Изд-во КНЦ РАН, 2011. 516 с.
Леонов А.В., Колтовская Е.В.,Чичерина О.В. Биогидрохимический портрет Белого моря // Компьютерные исследования и моделирование. 2018. Т. 10. № 1. С. 125–160.
Макаревич П.Р., Ларионов В.В. Годовой цикл развития планктонного фитоценоза Обь-Енисейского мелководья Карского моря // Океанология. 2011. Т. 37. № 1. С. 3–8.
Мошаров С.А., Демидов А.Б., Симакова У.В. Особенности процессов первичного продуцирования в Карском море в конце вегетационного периода // Океанология. 2016. Т. 56. № 1. С. 90–100.
Патин С. А. Нефть и экология континентального шельфа: в 2-х т. 2-е изд. т. 1: Морской нефтегазовый комплекс: состояние, перспективы, факторы воздействия. М.: Изд-во ВНИРО, 2017. 326 с.
Соловьева Н.В., Лобковский Л.И. Моделирование эколого-экономического риска при освоении шельфа // ДАН. 2015. Т. 464. № 3. С. 356–360.
Суханова И.Н., Флинт М.В., Дружкова Е.И. и др. Фитопланктон северо-западной части Карского моря // Океанология. 2015a. Т. 55. № 4. С. 605–619.
Суханова И.Н., Флинт М.В., Мошаров С.А., Сергеева В.М. Структура сообществ фитопланктона и первичная продукция в Обском эстуарии и на прилежащем Карском шельфе // Океанология. 2010. Т. 50. № 5. С. 785–800.
Суханова И.Н., Флинт М.В., Сергеева В.М. и др. Структура сообществ фитопланктона Енисейского эстуария и прилежащего Карского шельфа // Океанология. 2015б. Т. 55. № 6. С. 935–948.
Флейшман Б.С. Основы системологии. Изд-во: Радио и связь. М. 1982. 368 с.
Флинт М.В. Биоресурсы Арктических морей России: изменения под воздействием климата и факторов антропогенной природы, экосистемные основы охраны // Научно-технические проблемы освоения Арктики. РАН. М.: Наука, 2015. С. 55–71.
Флинт М.В. 54-й экспедиционный рейс научно-исследовательского судна “Академик Мстислав Келдыш” в Карское море // Океанология. 2010. Т. 50. № 5. С. 677–682.
Флинт М.В., Семенова Т.Н., Арашкевич Е.Г. и др. Структура зоопланктонных сообществ в области эстуарной фронтальной зоны реки Обь // Океанология. 2010. Т. 50. № 5. С. 809–822.
Chen S.Q., Chen B., Fath B.D. Ecological risk assessment on the system scale: A review of state-of-the-art models future perspectives // Ecol. Modell. 2013. V. 250. P. 25–33.
Chen S.Q., Fath B.D., Chen B. Information-based Network Environ Analysis: a system perspective for ecological risk assessment // Ecol. Indic. 2011. V. 11. P. 1664–1672.
Dale V.H., Biddinger G.R., Newman M.C. et al. Enhancing the ecological risk assessment process // Integrated Environmental Assessment and Management. 2008. V. 4(3). P. 306–313.
Demidov A.B., Kopelevich O.V., Mosharov S.A. et al. Modelling Kara Sea phytoplankton primary production: development and skill assessment of regional algorithms // J. Sea Res. 2017. V. 125. P. 1–17.
Demidov A.B., Mosharov S.A., Makkaveev P.N. Patterns of the Kara Sea primary production in autumn: Biotic and abiotic forcing of subsurface layer // J. Mar. Sys. 2014. V. 132. P. 130–149.
Fath B.D., Asmus H., Asmus R., Baird D., Borrett S.R., Jonge V., Ludovisi A., Niquil N., Scharler U.M., Schücke U., Wolff M. Ecological network analysis metrics: The need for an entire ecosystem approach in management and policy // Ocean and Coastal Management. 2019. V. 174. P. 1–14.
Fath B.D., Borrett S.R. A Matlab® Function for Network Environ Analysis // Env. Model. Soft. 2006. V. 21. P. 375–405.
Landis W.G. Ecological risk assessment conceptual model formulation for nonindigenous species // Risk Anal. 2004. V. 24. P. 847–858.
Landis W.G. Why has Ecological Risk Assessment found such limited application // Human and Ecological Risk Assessment. 2009. V. 15. P. 849–857.
Solovjova N.V. Synthesis of ecosystemic and ecoscreening modelling in solving problems of ecological safety // Ecol. Modell. 1999. V. 124. P. 1–10.
Solovjova N.V. Ecological risk modelling in developing resources of ecosystems characterized by varying vulnerability levels // Ecol. Modell. 2019. V. 406. P. 60–72.
Sukhanova I.N., Flint M.V., Pautova L.A. et al. Phytoplankton of the western Arctic in the spring and summer of 2002: Structure and seasonal changes // Deep-Sea Res., Part II. 2009. V. 56. P. 1223–1236.
Ulanowicz R.E. Quantitative methods for ecological network analysis // Comput. Biol. Chem. 2004. V. 28. P. 321–339.
Дополнительные материалы отсутствуют.