

Океанология, 2021, T. 61, № 6, стр. 958-968
Пространственная изменчивость зоопланктона и гидрологических показателей вод в Южной и Центральной части Балтийского моря в позднелетний сезон 2016 г.
Ю. Ю. Полунина 1, *, В. А. Кречик 1, 2, В. Т. Пака 1
1 Институт океанологии им. П.П. Ширшова РАН
Москва, Россия
2 Балтийский федеральный университет им. И. Канта
Калининград, Россия
* E-mail: jul_polunina@mail.ru
Поступила в редакцию 13.09.2020
После доработки 25.02.2021
Принята к публикации 08.04.2021
Аннотация
Выявлены особенности структуры и распределения позднелетнего зоопланктона открытой части Южной и Центральной Балтики (глубины 55–215 м) в 2016 г. с учетом гидрологических условий. В центральной части Балтийского моря (поверхностная соленость ~6.5 епс) таксономическое разнообразие зоопланктона выше, а общие количественные показатели зоопланктона ниже, чем в Южной Балтике (поверхностная соленость ~7.5 епс) – соответственно 18 и 14 видов; 16 ± 11 и 36 ± ± 18 тыс. экз/м3 и 208 ± 166 и 474 ± 182 мг/м3. Эти показатели находились в пределах многолетних значений для этого сезона. Максимальная численность и биомасса зоопланктона отмечена в верхнем слое до термоклина. Вертикальное распределение отдельных видов обусловлено их экофизиологическими особенностями: тепловодные виды Eubosmina maritima и Acartia tonsa преобладали в прогретом поверхностном слое, в то время как холодноводные стеногалинные Pseudocalanus elongatus и Oithona similis – в слое ниже галоклина. При снижении солености вод встречены виды солоноватоводного комплекса: Keratella quadrata и Limnocalanus grimaldii grimaldii – и выявлен рост доли в зоопланктоне Eurytemora affinis.
ВВЕДЕНИЕ
Для экосистемы Балтийского моря типичны значительные вариации природных и антропогенных условий, что обусловлено его внутриконтинентальным расположением, большой водосборной площадью и слабым водообменом с Атлантическим океаном. В акватории регистрируются климатические изменения метеорологических, гидрологических и гидрохимических условий, выражающиеся в тенденции роста температуры воздуха и воды, изменении атмосферной циркуляции [17], а также гидрофизических параметров и содержания биогенных элементов [21]. В то же время одной из основных особенностей гидрологического режима моря является наличие перманентного пикноклина, препятствующего перемешиванию поверхностного слоя с глубинными водами и тем самым усиливающего роль нерегулярных затоков североморских вод в изменчивости абиотических факторов в глубинных и придонных слоях Балтики [22]. Адвекция значительных объемов воды из Северного моря изменяет термохалинные показатели и содержание кислорода в придонном слое, что приводит к структурным перестройкам биологических сообществ [16]. Эти природные процессы наблюдаются при мощном антропогенном прессе на экосистему Балтики – активном судоходстве, регулируемом рыболовстве, нефтедобыче, избытке биогенных элементов и химическом загрязнении [8, 20]. Специфика географического положения, изменчивость режимообразующих факторов и различная степень влияния каждого из них в разных районах моря в совокупности с антропогенным воздействием приводит к активным преобразованиям планктонных сообществ в Балтийском море.
В последние десятилетия отмечены существенные перестройки в зоопланктоне, которые выражаются в изменении доминантного комплекса и доли отдельных видов, появлении чужеродных видов и значительных межгодовых вариациях общей численности и биомассы [1, 5, 9, 10, 13, 15, 25, 27]. Эти трансформации могут сказаться на пищевой ценности этого кормового ресурса для промысловых рыб Балтики [18] и влиять на рыбный промысел. В разных гидрологических бассейнах Балтики могут происходить разнонаправленные изменения в зоопланктоне [27], поэтому исследования даже одного года оказываются полезными как для оценки текущего состояния, так и для анализа долговременных изменений зоопланктона разных районов моря.
Цель работы ‒ выявить особенности структуры и распределения зоопланктона в условиях изменчивости гидрологических показателей вод Южной и Центральной Балтики в позднелетний сезон 2016 г.
РАЙОН ИССЛЕДОВАНИЙ, МАТЕРИАЛ И МЕТОДЫ
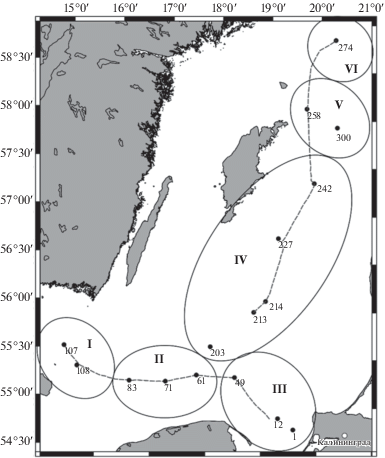
Исследование проводили в акватории Южной Балтики (ЮБ), включая Борнхольмский, Гданьский бассейны и Слупский желоб, а также Центральной Балтики (ЦБ), включая Готландскую, Северо-Балтийскую впадины и впадину Фарё. Материал отобран в 32-м рейсе НИС “Академик Николай Страхов” в период 02.08–16.09.2016 г. на глубинах 55–215 м.
Вертикальные профили распределения температуры (°С), солености (единицы практической солености – епс) и кислорода (мг/л) получены зондами CTD 90M (Sea&Sun Technology, Германия) и Ocean Seven 316 Plus (Idronaut, Италия) на отдельных участках маршрута судна и на 16 станциях, где были отобраны пробы зоопланктона (рис. 1). Для оценки гидрологической ситуации в районе работ по обработанным натурным данным был выполнен расчет характеристик основных элементов стратификации по методике [2]. Границы градиентных и однородных слоев вычислялись по значениям локального экстремума второй производной гидрофизических параметров. Для определения положения ядра в слоях скачка применялся экстремум первой производной температуры или солености для сезонного термоклина и галоклина соответственно. Такой метод показал хорошие результаты при исследовании параметров галоклина в бассейнах Балтийского моря [24]. Критерием выделения ядра холодного промежуточного слоя служило минимальное значение температуры на профиле [12].
Рис. 1.
Схема гидрологических разрезов (пунктирная линия) и расположения станций отбора зоопланктона в рейсе АНС-32 в позднелетний сезон 2016 г. Условные обозначения: Южная Балтика (I – Борнхольмский бассейн; II – Слупский желоб; III – Гданьский бассейн); Центральная Балтика (IV – Готландская впадина; V – впадина Фаре; VI – Северо-Балтийская впадина). Районы выделены по: [11].
Горизонты отбора проб зоопланктона на каждой станции выбирали на основании результатов измерений CTD-зондами с привязкой к положению сезонного термоклина и перманентного галоклина. Пробы отбирали в дневное время планктонной сетью WP-2 (Ø = 56 см, ячея 100 мкм) по слоям. Облавливали слои воды: от сезонного термоклина до поверхности (верхний квазиоднородный слой – ВКС); от начала галоклина до поверхности; весь столб воды от дна до поверхности (тотальный лов). Пробы фиксировали формалином до конечной концентрации 4%. Камеральную и статистическую обработку проб проводили по общепринятым методикам [7], биомассу рассчитывали по зависимости массы от длины тела организмов [3, 23].
РЕЗУЛЬТАТЫ
Гидрологические условия. В период наших исследований наблюдался “летний” тип термической стратификации, отличающийся от “зимнего” наличием теплого верхнего квазиоднородного слоя (ВКС) и сезонного термоклина (СТК), глубже которого располагался холодный промежуточный слой (ХПС). Ниже ХПС наблюдали галоклин (ГК), пространственно совпадающий с перманентным пикноклином, отделяющим поверхностную воду от глубинного слоя.
Термохалинная структура вод в южной части Балтийского моря представлена на рис. 2. Толщина ВКС здесь незначительно изменялась от бассейна к бассейну. Максимальные ее значения, как и значения температуры внутри данного слоя, наблюдались в Борнхольмской впадине (табл. 1). Толщина СТК снижалась в западном направлении (от Гданьской к Борнхольмской впадине). Аналогичным образом менялась мощность ХПС, при этом в западном направлении он становился теплее (табл. 1), а его ядро смещалось к нижней границе слоя. Наименьшая толщина ГК отмечалась в Слупском желобе. Градиенты в ядре менялись от 0.76 до 0.34 епс/м в восточном направлении (рис. 2а). Толщина глубинных слоев в каждой впадине была соизмеримой. Содержание кислорода в поверхностном слое и ХПС было схоже в разных бассейнах. Концентрация кислорода в поверхностном слое составляла: в Борнхольмском бассейне и Слупском желобе около 6, в Гданьском бассейне 6.1–6.7, в Готландской впадине 6.2–6.9, во впадинах Фарё и Северо-Балтийской 6.5–6.8 мл/л. Наименьшие значения кислорода отмечались в глубинном слое Борнхольмской и Гданьской впадин, где концентрация была менее 2 мл/л, в то время как в Слупском желобе глубинный слой был хорошо аэрирован (рис. 2a).
Рис. 2.
Расположение параметров стратификации вод в Южной (а) и Центральной Балтике (б), 2016 г. Условные обозначения: 1 – ВКС; 2 – ТК; 3 – ядро СТК; 4 – (ХПС); 5 – ядро ХПС; 6 – ГК; 7 – ядро ГК; 8 – изооксигена 2 мг/л; 9 – глубинный слой; 10 – дно.
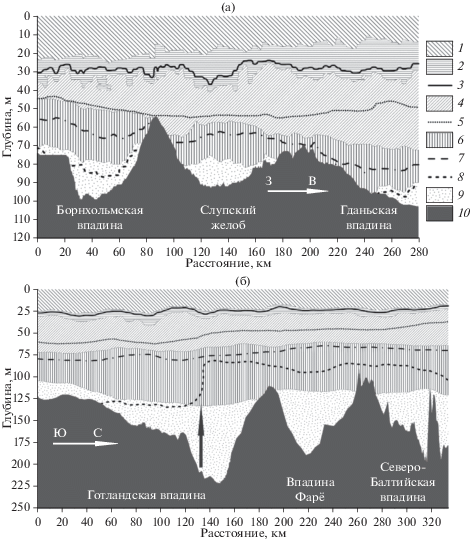
Таблица 1.
Основные количественные характеристики элементов термохалинной структуры в бассейнах Южной и Центральной Балтики в позднелетний сезон 2016 г.
| Элемент стратификации | Район | Толщина слоя, м | Температура, °С | Соленость, епс |
|---|---|---|---|---|
| ВКС | Борнхольмская впадина | 20–23 | 18.1–19.3 | 7.61–7.83 |
| Слупский желоб | 17–20 | 17.2–19.6 | 7.16–7.35 | |
| Гданьская впадина | 15–17 | 17.7–20.1 | 7.11–7.31 | |
| Готландская впадина | 20–27 | 16.8–17.4 | 6.21–7.22 | |
| Впадина Фарё | 17–23 | 16.1–16.6 | 5.99–6.34 | |
| Северо-Балтийская впадина | 17–20 | 15.8–16.3 | 5.95–6.16 | |
| СТК | Борнхольмская впадина | 10–12 | 18.1–7.2 | 7.63–7.88 |
| Слупский желоб | 12–15 | 17.5–6.8 | 7.19–7.56 | |
| Гданьская впадина | 12–15 | 17.1–6.8 | 7.21–7.52 | |
| Готландская впадина | 8–12 | 16.8–6.1 | 6.34–7.29 | |
| Впадина Фарё | 7–10 | 16.2–5.8 | 6.01–6.74 | |
| Северо-Балтийская впадина | 5–9 | 15.7–5.6 | 6.11–6.79 | |
| ХПС | Борнхольмская впадина | 13–20 | 5.5–6.8 | 7.67–8.83 |
| Слупский желоб | 15–24 | 4.8–5.7 | 7.54–8.59 | |
| Гданьская впадина | 27–36 | 4.8–5.4 | 7.51–8.63 | |
| Готландская впадина | 29–38 | 4.5–5.4 | 7.02–8.78 | |
| Впадина Фарё | 30–40 | 4.2–5.0 | 6.99–9.04 | |
| Северо-Балтийская впадина | 32–41 | 4.1–5.1 | 6.95–9.03 | |
| ГК | Борнхольмская впадина | 19–20 | 5.8–6.9 | 8.77–17.98 |
| Слупский желоб | 8–10 | 5.4–6.4 | 8.54–13.54 | |
| Гданьская впадина | 19–20 | 4.8–6.7 | 8.49–12.97 | |
| Готландская впадина | 34–71 | 4.9–6.5 | 8.67–12.46 | |
| Впадина Фарё | 50–56 | 4.8–6.4 | 8.83–12.23 | |
| Северо-Балтийская впадина | 48–57 | 4.9–6.1 | 8.89–11.59 | |
| Глубинный слой | Борнхольмская впадина | 6–17 | 6.4–6.9 | 17.99–18.88 |
| Слупский желоб | 5–16 | 5.9–6 | 13.55–14.99 | |
| Гданьская впадина | 3–12 | 6.7–7.1 | 12.98–13.37 | |
| Готландская впадина | 14–82 | 5.9–7.3 | 12.45–13.72 | |
| Впадина Фарё | 17–71 | 6.4–6.6 | 12.21–12.66 | |
| Северо-Балтийская впадина | 20–69 | 6.1–6.3 | 11.52–11.89 |
В центральной части моря элементы стратификации варьировали слабо (рис. 2б), однако величины основных гидрологических характеристик значительно изменялись от впадины к впадине (табл. 1). Наибольшая горизонтальная и вертикальная изменчивость значений температуры и солености наблюдалась в Готландской впадине, что связано как с ее большой протяженностью с юга на север, так и со значительными перепадами глубин внутри бассейна. Южный склон Готландской впадины был хорошо аэрирован до глубин около 130 м. Следует отметить резкий подъем изолинии концентрации кислорода (изооксигена) 2 мг/л на 50 м, который происходил над глубоководной частью впадины (отмечен стрелкой на рис. 2б). Этот подъем хорошо маркирует основную границу распространения обогащенных кислородом адвективных вод из юго-западных районов моря.
Зоопланктон был представлен 18 видами и таксономическими группами более высокого таксономического ранга, из которых Rotifera – 2, Cladocera – 6, Copepoda – 10, Tunicata – 1 вид (Fritillaria borealis Lohmann, 1896). В меропланктоне были многочисленны личинки Bivalvia, в незначительном количестве на некоторых станциях отмечены личинки Polychaeta, Gastropoda, Cirripedia, Decapoda и икра рыб.
Число видов зоопланктона на разных участках акватории изменялось от 10 до 20. Наиболее разнообразны были веслоногие ракообразные (5–10 видов на станции). Кладоцеры были представлены шестью видами: четыре вида, типичные обитатели Балтики (Eubosmina maritima P.E. Müller, 1867, Podon intermedius Lilljeborg, 1853, Pleopsis polyphemoides (Leuckart, 1859), Evadne nordmanni Lovén, 1836), отмечены на всех станциях, два вида понто-каспийских кладоцер – Cercopagis pengoi (Ostroumov, 1891) и Evadne anonyx G.O. Sars, 1897, встречены только в ЦБ, севернее о. Готланд. Популяции кладоцер-вселенцев активно размножались партеногенетически, у церкопагиса было отмечено половое поколение (самцы и гамогенетические самки с яйцами).
Наиболее разнообразен был зоопланктон в Центральной Балтике – здесь отмечено максимальное число видов (17–20). Только здесь зарегистрирован самый крупный вид калянид Балтики – Limnocalanus grimaldii grimaldii (Guerne, 1886), однако численность его была крайне низкой – 1 экз/м3. Вид встречался не повсеместно, а севернее о. Готланд в районе впадин Фарё и Северо-Балтийской, где в поверхностном слое воды соленость составляла 5.96–6.15 епс, а температура воды была +16°С.
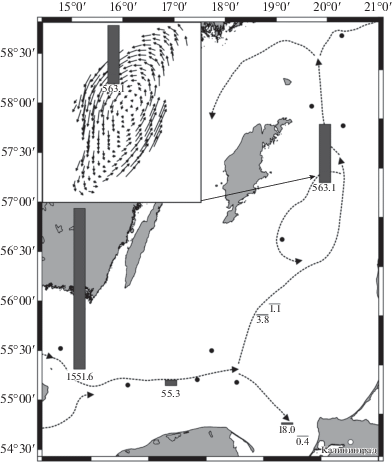
Циклоп Oithona similis Claus, 1866, который считается маркером передвижения каттегатских вод [4], был отмечен ниже галоклина во всех бассейнах Южной Балтики. Наиболее многочислен он был в Борнхольмском бассейне и Слупском желобе; в Гданьском бассейне его численность последовательно снижалась (рис. 3). Для Южной Балтики численность этого вида в среднем составила 203 экз/м3. В Центральной Балтике вид присутствовал на трех станциях, при этом наиболее многочисленный он был на ст. 242 (563.1 экз/м3), где глубина превышала 200 м (рис. 3), температура воды придонного слоя составляла 7.2°С, а соленость 13.6 епс. Севернее этот вид больше не встречался. Средняя численность циклопа в Центральной Балтике составила 81 экз/м3. Важно, что в районе ст. 242 по гидрофизическим данным выражен резкий подъем бескислородного слоя (рис. 2а). Это указывает на некую границу проникновения затоковых вод, что подтверждается и резким подъемом бескислородного слоя, и наличием здесь, и отсутствием севернее O. similis. Наличие значительного количества циклопа на ст. 242 может быть связано с динамическими особенностями слоя с затоковыми водами в Готландской впадине. Внутри глубоководной части впадины вода под пикноклином образует циклоническую циркуляцию [28]. Таким образом, гидродинамические условия в период нашего исследования могли способствовать образованию скопления данного вида в этом районе (рис. 3) с возможностью обитать в этих водах долгий период времени.
Рис. 3.
Распределение циклопа Oithona similis (экз/м3) в Южной и Центральной Балтике летом 2016 г. (столбики) и схема распространения затоковых вод (пунктирные стрелки) по: [16]. На врезке стрелками показана смоделированная и подтвержденная измерениями циклоническая схема циркуляции глубинных вод в Готландской впадине в предшествующий межзатоковый период [28], которая вероятна для периода наших наблюдений.
Основу зоопланктона по численности во всех слоях воды составляли мелкоразмерные организмы – кладоцера E. maritima (29–60%), науплиальные и копеподитные стадии калянид (22–38%).
Доля E. maritima была максимальной (до 60%) в поверхностном слое до термоклина. Доля Temora longicornis (Müller O.F., 1785) в общей численности зоопланктона изменялась в разных слоях незначительно (7.2–13.0%). Среди видов акарций наиболее многочисленной была Acartia longiremis (Lilljeborg, 1853), однако доля всех видов Acartia spp. в общей численности зоопланктона составляла от 3.4% в поверхностном слое до 10.4% в столбе воды. Доля Centropages hamatus (Lilljeborg, 1853) в столбе воды составила 5%, а в поверхностном слое всего 3%. Доля Pseudocalanus elongatus (Brady, 1865) была крайне низка и не превышала в среднем 1.5% от общей численности в столбе воды. В Центральной Балтике, при солености поверхностных вод 6.1–6.7 епс, существенно возросла доля Eurytemora affinis (Poppe, 1880) в общей численности зоопланктона – до 5.2%, в то время как в других районах Балтики, где соленость составляла 7.2–7.7 епс, доля это вида была менее 0.2%.
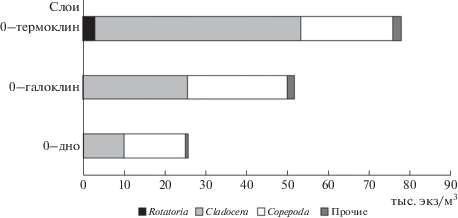
Количественное развитие массовых видов копепод T. longicornis, Ac. longiremis, C. hamatus в разных слоях отличалось. Наиболее высокие численности этих видов отмечены в слое, где облаливался термоклин, в котором температура воды ниже, чем в теплом ВКС. Холодноводный вид P. elongatus был, в основном, отмечен ниже галоклина. Выше галоклина вид встречался редко, преимущественно его копеподитные стадии, и доля этого вида составляла 0.3% от общей численности. В целом, вертикальное распределение отдельных видов зоопланктона обусловлено их экофизиологическими характеристиками. Тепловодные виды (кладоцеры, Acartia tonsa Dana, 1849) преобладали в прогретом ВКС, в то время как холодноводные стеногалинные P. elongatus; O. similis – в слое ниже галоклина. Максимальные количественные показатели зоопланктона были отмечены в слое ВКС. В этом слое отмечена максимальная доля кладоцер и заметна доля коловраток, чем в целом в столбе воды (рис. 4).
Рис. 4.
Структура зоопланктона (по численности) в разных слоях воды Балтике в позднелетний сезон 2016 г. Условные обозначения: 0–термоклин (диапазон глубин 17–27 м, в среднем 23 м); 0–галоклин (диапазон глубин 45–68, в среднем 58 м), 0–дно (диапазон глубин 50–205 м).
Основу численности зоопланктона составляли планктонные ракообразные. В Центральной Балтике на всех станциях более 50% всей численности зоопланктона принадлежало группе веслоногих ракообразных. В ЮБ на отдельных станциях более 50% составляли ветвистоусые ракообразные. Доля коловраток была незначительна, только в районе Северо-Балтийской впадины она возросла и составила почти 20% от общей численности зоопланктона (рис. 5).
Рис. 5.
Численность зоопланктона в столбе воды (тыс. экз/м3), Балтийское море, позднелетний сезон 2016 г. Условные обозначения: 1 – Copepoda, 2 – Cladocera, 3 – Rotifera, 4 – прочие. I–IV – районы Балтики.
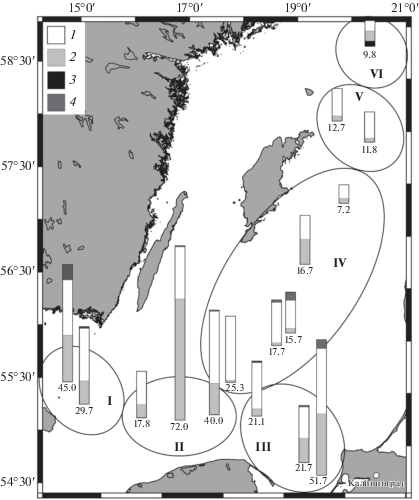
Высокие количественные показатели зоопланктона были характерны для ЮБ и превышали таковые в ЦБ (табл. 2, рис. 5). Хорошо заметна тенденция снижения общей численности и биомассы зоопланктона в направлении с юга на север. Встречаемость и распределение отдельных видов зоопланктона в разных районах Балтики обусловлены, главным образом, соленостными и температурными условиями. В более опресненной ЦБ, где поверхностная соленость составляла 6.2–6.7 епс, видовое разнообразие зоопланктона, включая меропланктон, достигало 17–20 видов, а общая численность зоопланктона в столбе воды составляла в среднем 16 ± 11 тыс. экз/м3. Минимальные количественные показатели планктона были отмечены севернее о. Готланд (районы V, VI) (табл. 2), но здесь же отмечено максимальное разнообразие зоопланктона, где появились коловратки, понто-каспийские кладоцеры и лимнокалянус.
Таблица 2.
Средние количественные показатели зоопланктона в столбе воды, температура воды и соленость в разных районах Балтики в позднелетний сезон 2016 г.
| Показатели/районы моря | Число видов голопланктона | N, тыс. экз/м3 | В, мг/м3 | Т воды, °С, поверхность/дно | S, епс, поверхность/дно | |
|---|---|---|---|---|---|---|
| Южная Балтика | I | 13 | 37.3 ± 7.7 | 468 ± 53 | 18.5/6.7 | 7.7/18.4 |
| II | 13 | 43.3 ± 19.2 | 536 ± 181 | 18.2/6.0 | 7.2/14.1 | |
| III | 14 | 31.5 ± 13.5 | 418 ± 154 | 19.2/6.8 | 7.2/13.4 | |
| Центральная Балтика | IV | 12 | 21.0 ± 9.3 | 318 ± 157 | 17.1/6.6 | 6.7/13.1 |
| V | 16 | 12.0 ± 0.5 | 167 ± 15 | 16.3/6.5 | 6.2/12.4 | |
| VI | 16 | 9.6 | 68 | 15.9/6.2 | 6.1/11.7 | |
Общая численность и биомасса зоопланктона в столбе воды изменялась в широких пределах: от 7 до 72 тыс. экз/м3 и от 96 до 808 мг/м3 соответственно. В Южной Балтике численность и биомасса зоопланктона в среднем была выше почти в два раза, чем в Центральной Балтике.
ОБСУЖДЕНИЕ
Видовой состав зоопланктона в позднелетний сезон 2016 г. был типичен для данного района Балтики и сравним с результатами предыдущих исследований [1, 10, 14, 27]. Доля меропланктона в общей численности и биомассе была несущественна, что согласуется с данными 2015 г. [27]. Разнообразие зоопланктона в ЦБ обусловлено значительной изменчивостью температуры и солености как на площади ее акватории, так и в толще воды. Это связано с ее большой субмеридиональной протяженностью, батиметрическими особенностями и спецификой водообмена как внутри впадин, так и с соседними районами моря. В районах, где было отмечено максимальное разнообразие зоопланктона (район впадин Фаре и Северо-Балтийской), отмечены самые низкие показатели температуры и солености как поверхностных, так и придонных вод (табл. 2).
Количественные показатели зоопланктона летом 2016 г. в столбе воды не превышали 80 тыс. экз/м3 и 808 мг/м3, составляя в среднем в разных районах 12–43 тыс. экз/м3 и 208–536 мг/м3. Полученные нами количественные показатели лежат в диапазоне значений, отмеченных в открытой части Юго-Восточной Балтики (ЮВБ) в летний период 2001–2015 гг. [10, 14] и в открытой части Южной и Центральной Балтики в 2015 г. [27].
Во многих районах Южной и Центральной Балтики наблюдаемое с 2000-го г. снижение численности зоопланктона продолжилось в 2015 г. [27]. В Борнхольмском и Готландском бассейнах численность массовых видов коловраток и кладоцер, как и всего зоопланктона, оставалась очень низкой в 2015 г. Максимальная численность коловраток была ниже 44 тыс. экз/м3, в то время как в 2000–2009 гг. регулярно превышала 100 тыс. экз/м3. Общее среднегодовое обилие зоопланктона 130 тыс. экз/м3 было самым низким с 2000 г. Также копеподы показали тенденцию к исторически низким количественным показателям: среди них особенно низкие были у видов P. elongatus, T. longicornis и C. hamatus, в то время как для Acartia spp. никакой тенденции не выявлено [27]. В целом, наши летние количественные показатели зоопланктона были ниже среднегодовых показателей 2015 г.
Особенности встречаемости и распределения отдельных видов обусловлены их экофизиологическими особенностями, прежде всего отношения к солености и температуре воды. Стеногалинные холодноводные виды O. similis, P. elongatus, F. borealis встречались только в слое воды ниже начала галоклина и не на всех станциях. Среди этих видов наиболее часто встречаемым и многочисленным был P. elongatus: численность взрослых особей и старших копеподитов в Борнхольмском бассейне и Слупском желобе была 12–331 экз/м3, а в Гданьском бассейне и Центральной Балтике, с учетом глубоководных впадин, размах численности варьировал от 45 до 1102 экз/м3. Численность этого вида в ЮВБ в летний период 1998–2007 гг. составляла 4368 экз/м3 [1], что многократно превышает современные показатели и косвенно указывает на возможное ухудшение соленостных или температурных условий для данного вида в этом районе. В придонном слое Гданьской впадина с 1990 по 2007 гг. температуры воды возросла почти на два градуса (от 5.3 до почти 7°С), в то же время соленость придонных вод также выросла почти на два епс и составляла 12.5 епс [1]. В нашем исследовании температура придонной воды в Гданьском бассейне также составила около 7°С. После большого затока 2014 г. температура в глубоководной части Гданьской впадины превышала среднемноголетний уровень. В частности, в период с августа 2014 г. по август 2016 г. наблюдаемые значения здесь варьировали в пределах 6.7–7.7°С [6]. Вероятно, повышение температуры воды придонного слоя стало неблагоприятным фактором для холодноводного P. elongatus.
Циклоп O. similis, предпочитающий холодные воды (1.6–15°С) с соленостью 7–34 епс, был малочисленным в летнем зоопланктоне 2016 г., самые высокие показатели отмечены в Борнхольмском бассейне – в среднем 775.8 экз/м3. Численность циклопа в Борнхольмском бассейне по летним батометрическим данным 1998 г. составляла 61–1226 тыс. экз/м3 [13], что превышает наши значения. По данным сетных сборов 1999 г., численность циклопа в этом районе в среднем варьировала от 400 (апрель) до 2100 экз/м3 (август) [19], что также выше показателей 2016 г. Статус популяции этого стенобионтного вида в изучаемом районе Балтики в настоящее время не ясен и требует более детальных исследований. По нашим данным, вид встречался не повсеместно, был малочисленен, и популяция была представлена половозрелыми самками и старшими копеподитными стадиями, но не отмечены самцы и науплии. Выявлено наличие положительной связи между показателями численности циклопа и соленостью глубинного слоя вод: R = 0.60; p = = 0.015, n = 16. Особенности пространственного распределения этого вида и выявленная положительная прямая зависимость между численностью этого вида и соленостью придонной воды дает основание предположить значительную вероятность пополнения популяции этого вида из Северного моря.
E. affinis обитает по данным разных авторов в широком диапазоне солености – от почти пресных вод (выше 0.2 епс) до распресненных морских (до 12–15 епс). В северной части Балтийского моря анализ многолетних данных (1973–1984 гг.) выявил, что E. ajfinis избегала солености выше 6.5 епс [26]. Вероятно, более низкая соленость вод ЦБ, в сравнении с ЮБ, более благоприятна для этого вида.
В целом, состав, распределение, динамика численности и биомассы отдельных групп или видов балтийского зоопланктона зависит как от сезонного цикла и естественного хода развития популяций, так и колебания теплового режима и солености вод. Поэтому сведения о состоянии зоопланктона и термохалинных условиях в конкретный период исследований даже одного года важны в контексте непрерывного мониторинга компонентов стрессовой экосистемы Балтийского моря.
ЗАКЛЮЧЕНИЕ
Встречаемость и распределение отдельных видов зоопланктона в разных районах Балтики обусловлены соленостными и температурными условиями. В более опресненной Центральной Балтике (где поверхностная соленость составляла в среднем 6.5 епс) видовое разнообразие зоопланктона выше (20 видов и таксонов голопланктона), в то же время общая количественные показатели зоопланктона невысоки (в среднем 16.1 тыс. экз/м3 и 208 мг/м3). В Борнхольмском и Гданьском бассейнах, при солености поверхностных вод 7.1–7.8 епс, разнообразие животного планктона было ниже (13–14 видов и таксонов голопланктона), а количественные показатели выше (в среднем 31.5–43.3 тыс. экз/м3 и 418–536 мг/м3). Вертикальное распределение летнего зоопланктона достаточно типично для морей умеренной зоны в целом и Балтики в частности – максимальная численность животного планктона отмечена в ВКС. Распределение отдельных видов по вертикали обусловлено их экофизиологическими особенностями: тепловодные виды преобладали в прогретом ВКС, в то время как холодноводные стеногалинные (P. elongatus; O. similis) – в слое ниже галоклина.
Источник финансирования. Работа выполнена в рамках госзадания ИО РАН, тема № 0128-2021-0012.
Список литературы
Александров С.В., Жигалова Н.Н., Зезера А.С. Многолетняя динамика зоопланктона в юго-восточном районе Балтийского моря // Биология моря. 2009. Т. 35. № 4. С. 241–248.
Белкин И.М. Морфолого-статистический анализ стратификации океана. Л.: Гидрометеоиздат, 1991. 130 с.
Виноградов М.Е., Шушкина Э.А. Функционирование планктонных сообществ эпипелагиали океана. М: Наука, 1987. 240 с.
Гидрометеорология и гидрохимия морей. Том III. Балтийское море. Выпуск 2: гидрохимические условия и океанологические основы формирования биологической продуктивности / Под. ред. Терзиева Ф.С., Рожкова В.А., Римша Е.Я., Шпаер И.С. Спб: Гидрометиздат, 1994. С. 7–8
Жигалова Н.Н., Пужакова Л.И. Состояние и межгодовая динамика развития зоопланктона в водах Южной Балтики в 1997-2001 годах / Промыслово-биологические исследования АтлантНИРО в 2000– 2001 годах. Т.2. Балтийское море. Калининград: АтлантНИРО, 2002. С. 12–21.
Кречик В.А., Капустина М.В., Бубнова Е.С. и др. Абиотические условия придонных вод Гданьской впадины Балтийского моря в 2016 году // Ученые записки РГГМУ. 2017. № 48. С. 186–194.
Методические рекомендации по сбору и обработке материалов при гидробиологических исследованиях на пресных водоемах. Зоопланктон и его продукция / Под ред. Салазкина А.А., Ивановой М.Б., Огородниковой В.А. Л.: ГосНИОРХ, 1984. 33 с.
Нефть и окружающая среда Калининградской области. Т. II.: Море / Под ред. Сивкова В.В. и др. Калининград: Терра Балтика, 2012. 576 с.
Полунина Ю.Ю. Стратегии размножения чужеродных планктонных ракообразных в Юго-восточной Балтике // Российский Журн. Биологических Инвазий № 4. 2017. С. 96–105.
Полунина Ю.Ю., Родионова Н.В. Характеристика зоопланктонного сообщества Балтийского моря / Система Балтийского моря. М.: Научный мир, 2017. С. 258–291.
Руденко М.В., Ражева Т.И. О новой батиметрической карте дна Балтийского моря // Океанология. 2004. Т. 44. № 3. С. 475–478.
Степанова Н.Б., Чубаренко И.П., Щука С.А. Структура и эволюция холодного промежуточного слоя в юго-восточной части Балтийского моря по данным натурных измерений в 2004–2008 гг. // Океанология. 2015. Т. 55. № 1. С. 32–32.
Щука Т.А. Характеристика современного состояния зоопланктона Балтийского моря. Автореф. дисс. канд. биол. наук. Москва, 2002. 28 с.
Щука Т.А. Зоопланктон. Биологические сообщества / Нефть и окружающая среда Калининградской области. Т. II: Море / Под ред. Сивкова В.В. и др. Калининград: Терра Балтика, 2012. С. 389–407.
Щука Т.А., Щука С.А. Динамика количественных характеристик чужеродных видов зоопланктона в юго-восточной части Балтийского моря в июле 2003–2015 гг. в связи с термохалинными условиями // Проблемы экологического мониторинга и моделирования экосистем. 2016. № 1. Т. 27. С. 86–108.
Bergen B., Naumann M., Herlemann D.P. et al. Impact of a Major Inflow Event on the Composition and Distribution of Bacterioplankton Communities in the Baltic Sea // Frontiers in Marine Science. 2018. 5. UNSP 383. https://doi.org/10.3389/fmars.2018.00383
Dailidienė I., Davulienė L., Kelpšaitė L. et al. Analysis of the climate change in Lithuanian coastal areas of the Baltic Sea // J. Coastal Res. 2012. V. 28. №. 3. P. 557–569.
Flinkman J., Aro E., Vuorinen I. et al. Changes in northern Baltic zooplankton and herring nutrition from 1980s to 1990s: top-down and bottom-up processes at work // Mar. Ecol.: Prog. Ser. 1998. V. 165. P. 127–136.
Hansen F.C., Möllmann C., Schütz U., Hinrichsen H. Spatio-temporal distribution of Oithona similis in the Bornholm Basin (Central Baltic Sea) // J. Plankton Res. 2004. V. 26. № 6. P. 659–668.
HELCOM, 2018. Input of nutrients by the seven biggest rivers in the Baltic Sea region // Baltic Sea Environment Proceedings. 2018. № 161. 31p.
Lainelaa S., Herkülb K., Leitoc I. et al. Contemporary trends in hydrophysical and hydrochemical parameters in the NE Baltic Sea // Estonian J. of Earth Sciences. 2020. V. 69. Is. 2. P. 91–10.
Mohrholz V., Naumann M., Nausch G., et al. Fresh oxygen for the Baltic Sea – An exceptional saline inflow after a decade of stagnation // J. of Marine Systems. 2015. V. 148. P. 152–166.
Recommendations on methods for marine biological studies in the Baltic Sea. Mesozooplankton biomass assessment. The Baltic marine biologists. Publication. Working Group 14. Hernroth L. (ed.). 1985. № 10. 26 p.
Reissmann J. On the representation of regional characteristics by hydrographic measurements at central stations in four deep basins of the Baltic Sea // Ocean Science Discussions. 2005. V. 2. P. 363–398.
Schulz J., Peck M.A., Barz K., Hirche H.-J. Spatial and temporal habitat partitioning by zooplankton in the Bornholm Basin (central Baltic Sea) // Prog. Oceanogr. 2012. № 107. P. 3–30.
Viitasalo M., Katajisto T., Vuorinen I. Seasonal dynamics of Acartia bifilosa and Eurytemora affinis (Copepoda: Calanoida) in relation to abiotic factors in the northern Baltic Sea // Hydrobiologia. 1994. 2921293: 415–422.
Wasmund N., Dutz J., Pollehne F. et al. Biological Assessment of the Baltic Sea 2015 / Meereswiss. Ber., Warnemünde, 2016. V. 102. 97 s.
Wieczorek G. Spatiotemporal Scales of the Deep Circulation in the Eastern Gotland Basin/Baltic Sea // Marine Science Reports. 2012. № 88. 146 p.
Дополнительные материалы отсутствуют.