

Океанология, 2021, T. 61, № 6, стр. 936-957
Первые данные о структуре фитопланктонных сообществ Восточно-Сибирского моря
И. Н. Суханова 1, М. В. Флинт 1, *, А. В. Федоров 2, Е. Г. Сахарова 3, П. Н. Маккавеев 1, А. А. Полухин 1, А. А. Недоспасов 1, А. С. Щука 1
1 Институт океанологии им. П.П. Ширшова РАН
Москва, Россия
2 Южное отделение Института океанологии им. П.П. Ширшова РАН
Геленджик, Россия
3 Институт биологии внутренних вод им. И.Д. Папанина РАН
Борок, Ярославская обл., Россия
* E-mail: m_flint@ocean.ru
Поступила в редакцию 12.01.2021
После доработки 18.05.2021
Принята к публикации 19.08.2021
Аннотация
Исследования структуры фитопланктонных сообществ Восточно-Сибирского моря выполнены в 69-м рейсе НИС “Академик Мстислав Келдыш” в рамках программы “Экосистемы морей Сибирской Арктики”. Материал получен с 5 по 9 сентября 2017 г. на двух квазимеридиональных разрезах от районов внутреннего шельфа, прилежащих к устьям Индигирки и Колымы, к внешнему шельфу. Приведен список доминирующих групп и видов водорослей, даны оценки численности, биомассы и описаны особенности вертикального распределения фитопланктона в биотопах с различными условиями пелагической среды. Установлена значительная количественная бедность фитопланктона Восточно-Сибирского моря по сравнению с морем Лаптевых в летне-осенний сезон, наиболее выраженная в восточной области бассейна. Численность водорослей на Индигирском разрезе менялась в пределах 11.9–66.3 × 103 кл/л, биомасса – 16.4–339.1 мг/м3, соответствующие цифры для Колымского разреза составили 7.7–90.3 × 103 кл/л и 9.4–42.1 мг/м3. Максимальные значения на обоих разрезах были отмечены в области внешней границы речного плюма при возрастании поверхностной солености до 21–25 PSU. Полученный материал позволяет констатировать существенные различия в структуре фитопланктонных сообществ западной и восточной областей Восточно-Сибирского моря, что является подтверждением его существенной зональной гетерогенности.
Несмотря на возрастающую активность исследований в эпиконтинентальных сибирских морях начиная с 2000-х гг., эта публикация традиционно начинается с констатации крайне низкой исследованности экосистемы Восточно-Сибирского моря по сравнению с другими районами арктического шельфа. Такое положение дел определяется, прежде всего, труднодоступностью бассейна и его высокой ледовитостью [1, 8, 26]. Наиболее масштабный сход сезонного льда здесь наблюдается в августе–самом начале сентября, когда открытую воду можно наблюдать на ~40% западной части акватории и на ~20% восточной [9]. И хотя в последнее время в бассейне наблюдается тенденция, связанная с общим снижением ледовитости в Арктике [24], она в малой степени проявляется во внутренней и срединной области огромного по площади (889 тыс. км2) и мелководного Восточно-Сибирского шельфа. Его средняя глубина менее 50 м, а глубины менее 30 м занимают половину района. Здесь скорее можно говорить об очень существенной межгодовой изменчивости площади акватории, освобождающейся летом от сезонного льда. Это наглядно иллюстрируют прекрасные арктические ледовые карты последнего десятилетия (http://www.aari.ru/clgmi/index.html; http://siows.solab.rshu.ru; https://nsidc.org). В частности, в августе 2018 г. в Восточно-Сибирском море была свободна ото льда лишь очень узкая прибрежная полоса шельфа, а в 2020 г. край постоянного ледяного поля в западной части бассейна наблюдался севернее континентального склона – севернее 77°–78° с.ш. Все это указывает на существенные короткопериодные флуктуации сигнала, несущего климатическую составляющую, в этом районе эпиконтинентальной Сибирской Арктики.
Восточно-Сибирское море является областью и границей проникновения на запад вод тихоокеанского происхождения, что хорошо видно по характеру зонального распределения океанологических характеристик [23, 28, 31]. Это явление хорошо прослеживается на шельфе и приводит к разделению его на две области – западную и восточную, существенно различающиеся по океанографическим условиям, важным для развития пелагической биоты, прежде всего солености, температуре и характеру вертикальной стратификации водной толщи. По имеющимся оценкам [26] граница проходит примерно по 160° в.д., но полученные нами данные (см. ниже) говорят о том, что ее положение на срединном шельфе, при относительно малом опресняющем влиянии речного стока на этот район, может быть восточнее. Проникновение вод тихоокеанского происхождения на шельф Восточно-Сибирского моря – важное явление, и его пространственно-временнáя изменчивость весьма вероятно может нести климатический сигнал, влияющий на параметры региональной экосистемы.
Немаловажное значение для многолетнего выпадения Восточно-Сибирского моря из числа ключевых объектов арктических исследований имело и малое (относительно других морей) воздействие континентального стока [22, 29], который рассматривается как мощнейший климатический сигнал в Арктику. Ежегодный речной сток в бассейн составляет около 10% всего стока в Сибирскую Арктику, он в три раза меньше, чем в море Лаптевых, и в шесть раз меньше, чем в Карском море [10, 22].
Имевшиеся сведения о низкой общей биологической продуктивности Восточно-Сибирского моря [5, 11, 12] также не стимулировали региональные исследования. Лишь в последние годы получены характеристики современного состава и биомассы основных компонентов пелагической экосистемы бассейна, прежде всего зоо- и ихтиопланктона [3, 4, 21].
Оценки состава, численности, биомассы фитопланктона и его продукции как наиболее динамичных характеристик состояния базовых трофических уровней экосистем являются ключевыми для оценки уровня и понимания механизмов климатической изменчивости арктических морей и их продуктивности. В этом отношении Восточно-Сибирское море следует считать критически недоисследованным. Оценки уровня первичной продукции в бассейне приведены в двух публикациях [6, 20]. При этом в [20] даны лишь данные измерений характеристик продуктивности автотрофного нанофитопланктона (0.5–7 мкм) на основании наблюдений в трех точках в северной части бассейна в области арктического континентального склона. [6] – единственное до настоящего времени исследование, основанное на массовых измерениях первичной продукции на шельфе Восточно-Сибирского моря с привлечением данных о концентрации хлорофилла и состоянии среды, включая подводную облученность. Эти работы выполнены параллельно с нашими наблюдениями, результаты которых представлены в настоящей статье.
В настоящее время данные о фитопланктоне Восточно-Сибирского моря содержатся только в трех доступных публикациях [2, 4, 27]. Материалы, представленные в [2, 4], имеют общее информационное значение, но при этом носят исключительно фрагментарный характер и не поддержаны необходимым для сегодняшних исследований описанием примененных методик, что лишает возможности использовать эти данные для сравнений. Даже наиболее полная на сегодня работа [27] содержит данные о фитоценозах лишь самой западной части Восточно-Сибирского моря и основана на материале, полученном на трех станциях на внутреннем шельфе южнее о. Новая Сибирь и пяти станциях севернее о. Котельный, т.е., по сути, на границе морей Восточно-Сибирского и Лаптевых. Пробы фитопланктона были собраны вертикальными ловами сетью Апштейна (ячея 20 мкм) и фиксированы 4% формальдегидом, что определяет недоучет важных размерных и таксономических компонентов фитоценозов и их ограниченную репрезентативность в части оценок численности и биомассы фитопланктона. Материалы по составу и количественным характеристикам фитопланктонных сообществ моря Лаптевых и восточной части Восточно-Сибирского моря рассмотрены авторами [27] в совокупности, что, в свою очередь, не позволяет говорить о специфике фитоценозов последнего района.
Настоящую работу можно считать первым исследованием фитопланктона шельфа Восточно-Сибирского моря, охватывающим западную и восточную части бассейна и основанным на современной методике, позволяющей максимально полно учесть все компоненты фитоценозов. Ее цель – оценить состав, количественные характеристики и особенности горизонтального и вертикального распределения фитопланктона, связать эти параметры с важнейшими условиями пелагической среды в разных по биотопическим условиям районах шельфа. Полученные данные сформируют базу для сравнительных оценок фитопланктонных сообществ с другими арктическими морями и дальнейших оценок возможной климатической изменчивости характеристик базового трофического компонента региональной экосистемы.
РАЙОН РАБОТ, МАТЕРИАЛ И МЕТОДЫ
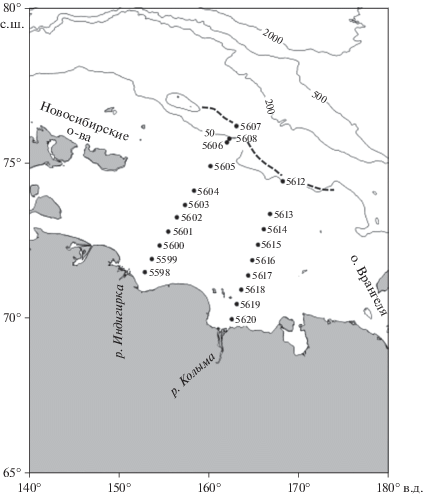
Материал получен в 69-м рейсе НИС “Академик Мстислав Келдыш” в рамках программы “Экосистемы морей Сибирской Арктики”. С 5 по 9 сентября 2017 г. на шельфе Восточно-Сибирского моря было сделано два квазимеридиональных разреза (рис. 1). Южные станции разрезов располагались в районах, находящихся под влиянием стока Индигирки и Колымы, северные – у кромки многолетнего льда. Расстояние между разрезами составило около 290 км. Индигирский разрез из 11 станций располагался между 71°28′ и 76°09′ с.ш. и имел протяженность 610 км. Фитопланктон на разрезе отобран на 8 станциях, на двух станциях 5602 и 5606 11 сентября были сделаны повторные наблюдения. Протяженность Колымского разреза составляла 550 км. На этом разрезе между 69°56′ и 74°23′ с.ш. было выполнено 9 станций, фитопланктон отобран на 6 станциях.
Рис. 1.
Схема расположения разрезов и станций в Восточно-Сибирском море. Пунктиром показано положение границы сезонного льда в период исследований.
Пробы воды для анализа фитопланктона объемом 2 л отбирали из 5-литровых батометров Нискина комплекса “Розетта”. Горизонты отбора были определены на основе данных предварительного зондирования температуры, солености и флуоресценции. Из верхнего перемешанного слоя получали 1–2 пробы, из слоя скачка плотности и максимума флюоресценции – 1–2 пробы и 1–2 пробы из слоя ниже пикноклина. На Индигирском разрезе было отобрано 38 проб, на Колымском разрезе – 22 пробы. Пробы для анализа фитопланктона и параллельных гидрохимических определений отбирали из одних и тех же батометров.
Концентрирование фитопланктона проводили с использованием лавсановых ядерных фильтров с размером пор 1 мкм на установках для мягкой обратной фильтрации. [13]. Объем полученного концентрата составлял 40–90 мл. Все пробы были обработаны без фиксации в живом состоянии сразу же после отбора проб или в течение одного-двух дней. Пробы хранили в холодильнике при температуре 2–3°С. Обработку проб вели в камерах Ножотта (объем 0.085 мл) при увеличении ×400 и Наумана (объем 1 мл) при увеличении ×200 на микроскопах Jena Lumar (Германия), Leica1000 (Швейцария) и МБИ-3 (Россия). Расчет сырой биомассы (объема) клеток делался на основе принципа геометрического подобия с использованием данных измерений линейных размеров. Содержание углерода в клетках рассчитывали, используя соответствующие коэффициенты для разных таксономических и размерных групп [25, 30].
РЕЗУЛЬТАТЫ
В целом, виды водорослей, встреченные в фитопланктоне Восточно-Сибирского моря (табл. 1), характерны для всех арктических морей. Как и в других морях арктического региона, наибольшим видовым разнообразием характеризовались диатомеи и динофлагелляты. Следует подчеркнуть, что часть клеток, в основном мелкоразмерных динофлагеллят и жгутиковых, а также клетки разных стадий развития динофлагеллят при полевой обработке материала определить более детально не удалось. Очевидно, что встреченное нами видовое разнообразие фитопланктона определялось сезоном максимально открытой ото льда поверхности моря в период проведения исследований.
Таблица 1.
Список основных видов фитопланктона на станциях Индигирского и Колымского разрезов
| BACILLARIOPHYCEAE | DINOPHYCEAE | DICTYOCHOPHYCEAE |
| Actinocyclus еhrenbergii | Alexandrium tamarense | Apedinella radians |
| Attheya septentrionalis | Alexandrium sp. | Dictyocha speculum |
| Chaetoceros comressus | Amphidinium cf. fusiforme | Pseudopedinella pyriforme |
| C. convolutus | A. longum | |
| C. debilis | A. sphaenoides | CHRYSOPHYCEAE |
| C. decipiens | Cochlodinium citron | Ollicola vangoorii (W. Conrad) |
| C. diadema | C. helix | Vørs, 1992 (Calycomonas |
| C. furcellatus | C. pupa | wulffii) |
| C. gracilis | Cochlodinium sp. | Dinobryon balticum |
| C. infolgianus | Dinophysis acuminata | D. faculiferum |
| C. socialis | D. acuta | Meringosphaera mediterranea |
| C. subtilis | D. arctica | M. tenerrima |
| C. tenuissimus | D. rotundata | |
| C. teres | Goniaulax scrippsae | PRASINOPHYCEAE |
| Cyclotella sp. | Goniaulax sp. | Nephroselmis sp. |
| Cylindrotheca closterium | Gymnodinium blax | Pyramimonas grossii |
| Fragilariopsis oceanica | G. abbreviatum | Pyramimonas sp. |
| Fragilariopsis sp. | G. gracile | |
| Gyrosigma cf. tenuirostrum | G. heterostriatum | EUGLENOPHYCEAE |
| Navicula directa | G. simplex | Eutreptiella sp. |
| N. distans | G. stellatum | |
| N. transitans | G. vitiligo | EBRIIDEA |
| Navicula sp. | G. wulffii | Ebria tripartite |
| Nitzschia frigida | Gymnodinium sp. | |
| N. longissima | Gyrodinium spirale | KINETOPLASTIDEA |
| Porosira glacialis | G. pingue | Leucocryptos marina |
| Pseudo-nitzschia delicatissima | Gyrodinium sp. | |
| P. seriata | Heterocapsa triquetra | |
| Rhizosolenia hebetata f. | Katodinium glaucum | |
| semispina | K. rotundatum | |
| Roperia tesselata | Lessardia elongata | |
| Skeletonema costatum | Nematodinium armatum | |
| Synedropsis hyperborea | Nematopsides vigilans | |
| Thalassiosira nordenskioeldii | Polykrikos schwarzii | |
| T. bioculata | Prorocentrum balticum | |
| Thalassiosira spp. | P. cordatum | |
| Protoceratium reticulatum | ||
| Protoperidinium bipes | ||
| P. brevipes | ||
| P. depressum | ||
| P. ovatum | ||
| P. pallidum | ||
| P. pellucidum | ||
| Torodinium robustum |
Несмотря на то, что южные станции разрезов были сделаны вблизи эстуариев Индигирки и Колымы, нами не были встречены пресноводные виды, характерные для аналогичных шельфовых районов Карского моря и моря Лаптевых, непосредственно прилежащих к эстуариям крупных рек Оби, Енисея и Лены [14, 16, 17]. Наиболее вероятно, что ограничивающим фактором была соленость. Соленость на южных станциях Индигирского и Колымского разрезов составляла 16–18 PSU (рис. 2), а как показали наши предыдущие исследования в других эпиконтинентальных районах Арктики, подверженных воздействию речного стока, при солености >15 PSU пресноводные виды практически исчезают из планктона [14, 16, 17].
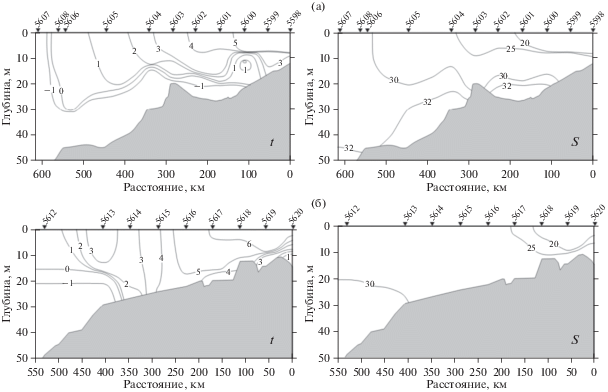
Индигирский разрез
Поверхностный опресненный слой с соленостью от 16 до 25 PSU, подстилаемый резким пикноклином, наблюдался на протяжении 330 км от дельты Индигирки вплоть до ст. 5604 (рис. 2а). На этом участке разреза толщина верхнего перемешенного слоя увеличивалась в мористом направлении от 5 до 10 м. Градиент солености в слое скачка при продвижении на север постепенно уменьшался от очень высоких значений 5.6–7.1 PSU/м на самых южных станциях разреза до 1.4 PSU/м в его центральной части. Температура поверхностного слоя вблизи эстуария составляла 6.2°С и снижалась до 3.2 °С на ст. 5603 в центральной части разреза.
Во внешней части разреза между станциями 5606 и 5608 по мере продвижения на север соленость верхнего перемешенного слоя быстро увеличивалась и превысила 30 PSU. Параллельно возрастала и толщина верхнего перемешенного слоя: его нижняя граница на ст. 5604 располагалась на глубине 12 м, а на станциях 5606–5608 опустилась на глубину 22–27 м (рис. 2а). Градиент солености в пикноклине колебался от 0.6 до 1.2 PSU/м. Температура перемешенного слоя во внешней части разреза опускалась до 0.2°С (ст. 5608), на ст. 5607 непосредственно у края ледяного поля составляла –1.4°С.
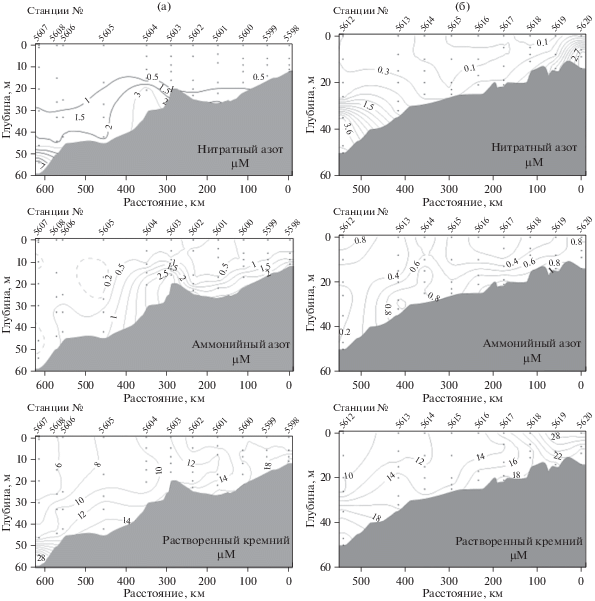
На всех станциях Индигирского разреза концентрация нитратов в верхнем 15–20 м слое была ниже 0.5 μМ. Величин 3.0 μМ содержание NO3 достигало только в придонном слое на станциях 5604 и 5607 (рис. 3а). Повышенное содержание аммонийного азота (1.02–1.28 μМ) наблюдалось в районе, прилежащем к эстуарию, и на ст. 5604. Максимальные концентрации кремния в поверхностном слое – 15–18 μМ – были зарегистрированы на ближайших к эстуарию станциях (рис. 3а). Севернее содержание SiO3 постепенно снижалось и вблизи кромки льда составляло 5–6 μМ.
Рис. 3.
Распределение нитратного азота, аммонийного азота и растворенного кремния на Индигирском (а) и Колымском (б) разрезах.
Численность фитопланктона на разрезе в толще воды колебалась от 11.9 × 103 кл/л в среднем для столба воды (1.3 × 108 кл/м2) на ст. 5598 до 66.3 × 103 кл/л (1.5 × 109 кл/м2) на ст. 5602 (рис. 4а, табл. 2). Ст. 5602 с максимальной численностью фитопланктона была сделана на внешней периферии области, существенно опресненной речным стоком, при солености верхнего перемешенного слоя 21.2 PSU (рис. 2а, 4а). 75% общей численности фитопланктона здесь приходилось на долю мелкоклеточного (объем клетки около 40 мкм3) миксотрофного вида Chrysophyceae Ollicola vangoorii (табл. 2). В поверхностном слое на ст. 5602, наряду с высоким обилием O. vangoorii, зарегистрирована высокая численность спор Chaetoceros socialis. Фитопланктон был в основном сконцентрирован в верхнем перемешенном слое (рис. 5). В придонном слое той же станции помимо спор С. socialis (58% от общего числа клеток Chaetoceros), 21% составляли споры С. debilis и 21% приходился на долю спор С. furcellatus, С. Compressus и C. diadema. O. vangoorii, иногда в сочетании с Katodinium rotundatum (объем клетки ~200 мкм3), определял численность фитопланктона на всех станциях южной части разреза (5598–5602).
Рис. 4.
Численность (N × 103 кл/л), биомасса (В, мг/м3) фитопланктона, поверхностная (Sпов) и придонная (Sпр) соленость (PSU), глубина верхнего перемешанного слоя (ВКС) на станциях Индигирского (а) и Колымского (б) разрезов.
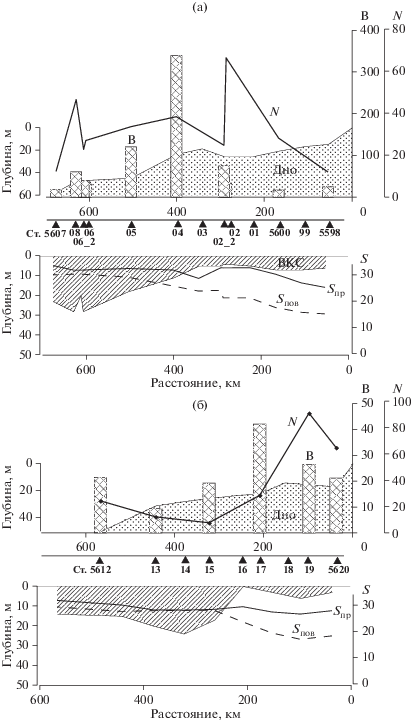
Таблица 2.
Индигирский разрез. Численность (N × 103кл/л, %), биомасса (B, мг/м3, %) и биомасса в углероде (Вс, мг/м3, %) основных видов/групп фитопланктона на горизонтах отбора проб. Численность (N × 103 кл/м2, %), биомасса (В, мг/м2, %), биомасса в углероде (Вс, мг/м2, %) в столбе воды
| Станция | Горизонт | N B |
Bacillariophyceae | Dynophyсеae | Ollicola vangoorii (Calycomonas wulffii) |
Rest | Общая | ||
|---|---|---|---|---|---|---|---|---|---|
| Chaetoceros (все) | Rhizosolenia hebetata | rest | |||||||
| 5598 | 0 | N | 1312 | _ | _ | 2820 | 1500 | 2634 | 8266 |
| 15.8% | 34.1% | 18.1% | 31.9% | ||||||
| B | 0.13 | _ | _ | 33.4 | 0.05 | 0.77 | 34.3 | ||
| 0.4% | 97.4% | 0.1% | 2.2% | ||||||
| 6 | N | 236 | _ | _ | 17 027 | 1707 | 1977 | 20 947 | |
| 1.1% | 81.4% | 8.1% | 9.4% | ||||||
| B | 1.35 | _ | _ | 34.34 | 0.06 | 0.42 | 36.17 | ||
| 3.7% | 94.9% | 0.2% | 1.2% | ||||||
| 9 | N | _ | _ | 756 | 3781 | _ | 3083 | 7620 | |
| 9.9% | 49.6% | 40.5% | |||||||
| B | _ | _ | 0.2 | 1.08 | _ | 0.83 | 2.12 | ||
| 9.9% | 50.9% | 39.1% | |||||||
| 11 | N | _ | _ | 50 | 50 | _ | _ | 100 | |
| 50% | 50% | ||||||||
| B | _ | _ | 0.02 | 0.11 | _ | _ | 0.13 | ||
| 5.4% | 84.6% | ||||||||
| В слое | N | 4998 | _ | 1940 | 94 584 | 12 181 | 17 570 | 131 273 | |
| 3.8% | 1.5% | 72.0% | 9.3% | 13.4% | |||||
| B | 6.5 | _ | 0.52 | 257.5 | 0.42 | 6.28 | 271.2 | ||
| 2.4% | 0.2% | 95.0% | 0.1% | 2.3% | |||||
| Bc | 0.43 | _ | 0.03 | 32.2 | 0.03 | 0.63 | 33.3 | ||
| 1.3% | 0.1% | 96.7% | + | 1.9% | |||||
| 5599 | 0 | N | 1378 | _ | _ | 11 850 | 10 914 | 642 | 24 784 |
| 5.6% | 47.8% | 44.0% | 2.6% | ||||||
| B | 5.9 | _ | _ | 43.7 | 0.38 | 0.07 | 50.02 | ||
| 11.7% | 87.3% | 0.8% | 0.1% | ||||||
| 5600 | 0 | N | 336 | _ | _ | 504 | 20 856 | 13 272 | 34 968 |
| 1.0% | 1.4% | 59.6% | 37.9% | ||||||
| B | 0.3 | _ | _ | 15.7 | 0.8 | 0.95 | 17.8 | ||
| 1.7% | 88.4% | 4.5% | 5.3% | ||||||
| 6 | N | 688 | _ | _ | 2552 | 36 646 | 1133 | 41 019 | |
| 1.7% | 6.2% | 89.3% | 2.8% | ||||||
| B | 2.4 | _ | _ | 21.85 | 1.5 | 0.15 | 25.7 | ||
| 9.3% | 85.0% | 5.0% | 0.6% | ||||||
| 9 | N | 96 | _ | 48 | 1671 | 12 254 | 605 | 14 674 | |
| 0.6% | 0.3% | 11.4% | 83.5% | 4.1% | |||||
| B | 0.9 | _ | 0.05 | 0.31 | 0.49 | 0.07 | 1.82 | ||
| 49.5% | 2.7% | 17.0% | 26.9% | 3.8% | |||||
| 12 | N | 116 | _ | _ | 231 | _ | 1778 | 2125 | |
| 5.5% | 10.9% | 83.7% | |||||||
| B | 0.16 | _ | _ | 11.7 | _ | 0.67 | 12.5 | ||
| 1.3% | 93.4% | 5.4% | |||||||
| В слое | N | 4566 | _ | 144 | 18 355 | 264 237 | 49 396 | 336 698 | |
| 1.4% | + | 5.4% | 78.4% | 14.8% | |||||
| B | 14.7 | _ | 0.15 | 163.9 | 13.1 | 4.7 | 196.55 | ||
| 7.5% | + | 83.4% | 6.7% | 2.4% | |||||
| Bc | 1.0 | _ | 0.01 | 20.5 | 1.1 | 0.5 | 23.1 | ||
| 4.3% | + | 88.7% | 4.8% | 2.2% | |||||
| 5601 | 0 | N | 6320 | 45 | _ | 8667 | 99 757 | 21 486 | 136 275 |
| 4.6% | + | 6.4% | 73.2% | 15.8% | |||||
| B | 1.8 | 0.8 | _ | 26.13 | 2.8 | 2.7 | 34.2 | ||
| 5.2% | 2.3% | 76.4% | 8.2% | 7.9% | |||||
| 5602 | 0 | N | 52 256 | 850 | 25 | 8102 | 93 720 | 9088 | 164 016 |
| 31.9% | 0.5% | + | 4.9% | 57.1% | 5.5% | ||||
| B | 3.5 | 46.4 | + | 19.5 | 6.6 | 0.9 | 77.0 | ||
| 4.5% | 60.2% | + | 25.4% | 8.6% | 1.2% | ||||
| 5 | N | _ | 1652 | _ | 4766 | 112 800 | 2950 | 122 168 | |
| 1.4% | 3.9% | 92.3% | 2.4% | ||||||
| B | _ | 29.6 | _ | 13.4 | 4.0 | 1.42 | 48.4 | ||
| 61.2% | 27.7% | 8.3% | 2.9% | ||||||
| 16 | N | 1430 | 228 | 325 | 845 | _ | _ | 2828 | |
| 50.6% | 8.1% | 11.5% | 29.8% | ||||||
| B | 3.04 | 5.23 | 0.05 | 0.54 | _ | _ | 8.86 | ||
| 34.3% | 59.0% | 0.6% | 6.1% | ||||||
| 20 | N | _ | 73 | _ | 1570 | 2372 | 949 | 4964 | |
| 1.5% | 31.6% | 47.8% | 19.1% | ||||||
| B | _ | 1.42 | _ | 2.85 | 0.06 | 1.02 | 5.35 | ||
| 26.5% | 53.4% | 1.1% | 19.0% | ||||||
| 23 | N | 59 229 | 160 | 6320 | _ | _ | _ | 65 709 | |
| 90.2% | 0.2% | 9.6% | |||||||
| B | 16.57 | 2.02 | 55.0 | _ | _ | _ | 73.6 | ||
| 22.6% | 2.7% | 74.7% | |||||||
| В слое | N | 230 209 | 17 547 | 12 280 | 70 216 | 1 145 002 | 49 642 | 1 524 896 | |
| 15.1% | 1.1% | 0.8% | 4.6% | 75.2% | 3.2% | ||||
| B | 56.5 | 400.0 | 83.0 | 170.0 | 48.7 | 17.2 | 775.4 | ||
| 7.3% | 51.6% | 10.7% | 21.9% | 6.3% | 2.2% | ||||
| Bc | 3.8 | 25.0 | 5.5 | 21.2 | 4.0 | 1.7 | 61.2 | ||
| 6.2% | 40.9% | 9.0% | 34.6% | 6.5% | 2.8% | ||||
| 5603 | 0 | N | 121 875 | 8250 | 6000 | 5250 | 60 000 | 7500 | 208 875 |
| 58.3% | 3.9% | 2.9% | 2.5% | 28.7% | 3.6% | ||||
| B | 14.7 | 206.25 | 1.5 | 13.9 | 4.2 | 1.8 | 242.4 | ||
| 6.1% | 85.2% | 0.6% | 5.7% | 1.7% | 0.7% | ||||
| 5604 | 0 | N | 4640 | 14 500 | _ | 2926 | 5220 | 12 760 | 40 046 |
| 11.5% | 36.3% | 7.3% | 13.0% | 31.9% | |||||
| B | 0.34 | 394.4 | _ | 10.8 | 0.36 | 3.2 | 409.1 | ||
| + | 96.4% | 2.6% | + | 0.8% | |||||
| 11 | N | 1050 | 22 660 | 859 | 3988 | 6592 | 5768 | 40 917 | |
| 2.6% | 55.4% | 2.1% | 9.7% | 16.1% | 14.1% | ||||
| B | 1.59 | 354.9 | 1.0 | 8.94 | 0.23 | 1.03 | 367.7 | ||
| 0.4% | 96.5% | 0.3% | 2.4% | + | 0.3% | ||||
| 15 | N | 21 648 | 1185 | 3317 | 2290 | _ | _ | 28 440 | |
| 76.1% | 4.2% | 11.7% | 8.0% | ||||||
| B | 4.7 | 29.2 | 0.28 | 5.26 | _ | _ | 39.44 | ||
| 11.9% | 4.1% | 0.7% | 13.3% | ||||||
| В слое | N | 76 690 | 252 070 | 13 078 | 50 583 | 78 150 | 113 440 | 584 011 | |
| 13.1% | 43.2% | 2.2% | 8.7% | 13.4% | 19.4% | ||||
| B | 23.2 | 4889.4 | 8.1 | 137.0 | 3.7 | 25.3 | 5086.7 | ||
| 0.5% | 96.1% | 0.2% | 2.7% | + | 0.5% | ||||
| Bc | 1.6 | 326.0 | 0.5 | 17.1 | 0.3 | 2.5 | 348.0 | ||
| 0.5% | 93.8% | + | 4.9% | + | 0.7% | ||||
| 5605 | 0 | N | 5120 | 4076 | 330 | 1436 | 3960 | 1708 | 16 630 |
| 30.8% | 24.5% | 2.0% | 8.6% | 23.8% | 10.3% | ||||
| B | 1.0 | 102.1 | 0.25 | 5.03 | 0.28 | 0.83 | 109.5 | ||
| 0.9% | 93.2% | 0.2% | 4.6% | 0.2% | 0.8% | ||||
| 17 | N | 6765 | 8820 | 55 | 1485 | _ | _ | 17 125 | |
| 39.5% | 51.5% | 0.3% | 8.7% | ||||||
| B | 11.45 | 150.4 | 0.05 | 7.68 | _ | _ | 169.6 | ||
| 6.7% | 88.7% | + | 4.5% | ||||||
| 33 | N | 83 205 | 237 | 631 | 39 | _ | _ | 84 112 | |
| 98.9% | 0.3% | 0.7% | + | ||||||
| B | 17.4 | 9.4 | 3.6 | 0.02 | _ | _ | 30.42 | ||
| 57.2% | 30.9% | 11.8% | + | ||||||
| В слое | N | 820 782 | 182 072 | 8760 | 37 020 | 33 660 | 14 518 | 1 096 812 | |
| 75.0% | 16.6% | 0.8% | 3.3% | 3.0% | 1.3% | ||||
| B | 336.6 | 3424.6 | 31.7 | 169.6 | 2.4 | 7.0 | 3971.9 | ||
| 8.5% | 86.2% | 0.8% | 4.3% | + | 0.2% | ||||
| Bc | 22.4 | 228.3 | 2.1 | 21.2 | 0.2 | 0.7 | 274.9 | ||
| 8.1% | 83.1% | 0.8% | 7.7% | + | 0.2% | ||||
| 5606 | 0 | N | 27118 | 1107 | 65 | 1302 | _ | 8130 | 37 722 |
| 71.9% | 2.9% | 0.2% | 3.4% | 21.6% | |||||
| B | 20.9 | 32.1 | 0.05 | 5.1 | _ | 0.92 | 59.1 | ||
| 35.4% | 54.3% | + | 8.6% | 1.6% | |||||
| 15 | N | 14 470 | 376 | 965 | 750 | 697 | 6968 | 24 226 | |
| 59.6% | 1.6% | 4.0% | 3.1% | 2.9% | 28.8% | ||||
| B | 14.6 | 11.1 | 0.6 | 1.74 | 0.03 | 0.1 | 28.2 | ||
| 51.8% | 39.5% | 2.1% | 6.2% | 0.1% | 0.3% | ||||
| 33 | N | 18 468 | 108 | 270 | 881 | 638 | 54 | 20 419 | |
| 90.4% | 0.5% | 1.3% | 4.3% | 3.1% | 0.3% | ||||
| B | 4.4 | 2.2 | 0.5 | 4.5 | 0.02 | 0.03 | 11.65 | ||
| 37.8% | 18.9% | 4.3% | 38.5% | 0.2% | 0.3% | ||||
| В слое | N | 608 352 | 15 478 | 18 840 | 52 209 | 17 243 | 176 433 | 888 555 | |
| 68.5% | 1.7% | 2.1% | 5.9% | 1.9% | 19.9% | ||||
| B | 437.3 | 443.7 | 14.8 | 107.5 | 0.7 | 8.8 | |||
| 43.2% | 43.8% | 1.5% | 10.6% | + | 0.9% | 1012.8 | |||
| Bc | 29.2 | 29.6 | 1.0 | 13.4 | 0.06 | 0.9 | |||
| 39.4% | 39.9% | 1.3% | 18.1% | + | 1.2% | 74.2 | |||
| 5608 | 0 | N | 20 119 | 1023 | 1364 | 2728 | _ | 2869 | 28 103 |
| 71.6% | 3.6% | 4.8% | 9.7% | 10.2% | |||||
| B | 29.1 | 22.1 | 1.0 | 27.7 | _ | 0.71 | 80.6 | ||
| 36.1% | 27.4% | 1.2% | 34.4% | 0.9% | |||||
| 8 | N | 43 770 | 390 | 2114 | 754 | _ | _ | 47 028 | |
| 93.1% | 0.8% | 4.5% | 1.6% | ||||||
| B | 72.8 | 12.4 | 1.8 | 6.5 | _ | _ | 93.5 | ||
| 77.9% | 3.2% | 1.9% | 7.0% | ||||||
| 33 | N | 49 658 | _ | 248 | 1302 | _ | 403 | 51 611 | |
| 96.2% | 0.5% | 2.5% | 0.8% | ||||||
| B | 7.9 | _ | 0.4 | 1.25 | _ | 0.05 | 9.6 | ||
| 82.3% | 4.2% | 13.0% | 0.5% | ||||||
| В слое | N | 1 423 406 | 10 527 | 4343 | 38 428 | _ | 16 514 | 1 493 218 | |
| 95.3% | 0.7% | 0.3% | 2.6% | 1.1% | |||||
| B | 1416.4 | 293.0 | 38.7 | 233.6 | _ | 3.5 | 1985.2 | ||
| 71.4% | 14.7% | 1.9% | 11.8% | 0.2% | |||||
| Bc | 94.4 | 19.5 | 2.6 | 29.2 | _ | 0.3 | 146.0 | ||
| 64.8% | 13.4% | 1.8% | 20.0% | 0.2% | |||||
| 5607 | 0 | N | 14 314 | _ | 33 | 3478 | 3081 | 15 570 | 36 476 |
| 39.3% | + | 9.5% | 8.4% | 42.7% | |||||
| B | 11.6 | _ | 0.02 | 19.0 | 0.07 | 1.9 | 32.6 | ||
| 35.6% | + | 58.3% | 0.2% | 5.8% | |||||
| 10 | N | 3822 | 260 | _ | 2472 | _ | 1618 | 8172 | |
| 46.8% | 3.2% | 30.2% | 19.8% | ||||||
| B | 6.5 | 5.6 | _ | 10.8 | _ | 0.5 | 23.4 | ||
| 27.9% | 23.9% | 46.1% | 2.1% | ||||||
| 20 | N | 1705 | _ | 341 | 2728 | 341 | 4092 | 9207 | |
| 18.5% | 3.7% | 29.7% | 3.7% | 44.4% | |||||
| B | 0.2 | _ | 0.1 | 6.5 | 0.02 | 4.3 | 11.1 | ||
| 1.8% | 0.9% | 58.5% | 0.2% | 38.6% | |||||
| 30 | N | 448 | 56 | _ | 744 | _ | 1018 | 2266 | |
| 19.8% | 2.5% | 32.8% | 44.9% | ||||||
| B | 1.05 | 0.75 | _ | 2.6 | _ | 0.4 | 4.8 | ||
| 21.9% | 15.6% | 54.2% | 8.3% | ||||||
| В слое | N | 129 080 | 2880 | 3575 | 73 110 | _ | 140 040 | 367 500 | |
| 35.1% | 0.8% | 1.0% | 9.9% | 8.1% | |||||
| B | 130.3 | 63.5 | 1.1 | 281.0 | _ | 59.5 | |||
| 24.3% | 11.9% | 0.2% | 52.5% | 11.0% | 535.4 | ||||
| Bc | 8.6 | 4.2 | 0.07 | 31.2 | _ | 6.0 | |||
| 17.2% | 8.4% | 0.1% | 62.2% | 12.0% | 50.1 | ||||
| 5602-2 | 0 | N | 1204 | 10 014 | _ | 11 160 | 42 140 | 15 077 | 79 595 |
| 1.5% | 12.7% | 14.0% | 52.9% | 18.9% | |||||
| B | 0.12 | 200.3 | _ | 44.0 | 2.9 | 6.4 | 253.7 | ||
| + | 79.0% | 17.3% | 1.1% | 2.5% | |||||
| 10 | N | _ | 924 | 2600 | 1882 | 1032 | 2164 | 8602 | |
| 10.8% | 30.2% | 21.8% | 12.0% | 25.2% | |||||
| B | _ | 18.5 | 1.3 | 1.2 | 0.05 | 0.15 | 21.2 | ||
| 87.3% | 6.1% | 5.7% | 0.2% | 0.7% | |||||
| 20 | N | 350 | 100 | 25 | 25 | _ | 1325 | 1824 | |
| 19.2% | 5.5% | 1.4% | 1.4% | 72.5% | |||||
| B | 0.21 | 2.0 | 0.01 | 0.015 | _ | 0.16 | 2.4 | ||
| 8.8% | 83.4% | 0.4% | 0.6% | 6.7% | |||||
| В слое | N | 7770 | 59 810 | 26 125 | 74 745 | 221 020 | 103 650 | 493120 | |
| 1.6% | 12.1% | 5.3% | 15.2% | 44.8% | 21.0% | ||||
| B | 1.65 | 1226 | 13.5 | 232.1 | 15.0 | 34.3 | 1522.6 | ||
| 0.1% | 80.5% | 0.9% | 15.2% | 1.0% | 2.2% | ||||
| Bc | 0.1 | 81.7 | 0.9 | 29.0 | 1.25 | 3.4 | 116.5 | ||
| + | 70.3% | 0.8% | 24.9% | 1.1% | 2.9% | ||||
| 5606-2 | 0 | N | 9860 | 2926 | 1414 | 870 | 1160 | 2056 | 18 322 |
| 53.9% | 16.0% | 7.7% | 4.9% | 6.3% | 11.2% | ||||
| B | 30.0 | 58.5 | 1.0 | 1.5 | 0.08 | 1.3 | 92.4 | ||
| 32.5% | 3.3% | 1.1% | 1.6% | + | 1.4% | ||||
| 10 | N | 7816 | 1196 | 3901 | 1880 | 1625 | 997 | 17 415 | |
| 44.9% | 6.9% | 2.4% | 10.8% | 9.3% | 5.7% | ||||
| B | 6.8 | 23.9 | 0.74 | 10.7 | 0.08 | 0.34 | 42.6 | ||
| 16.0% | 56.1% | 1.7% | 25.1% | 0.2% | 0.8% | ||||
| 33 | N | 33 200 | 240 | _ | 825 | _ | 35 | 34 300 | |
| 96.8% | 0.7% | 2.4% | 0.1% | ||||||
| B | 7.4 | 4.8 | _ | 1.2 | _ | 0.09 | 13.5 | ||
| 54.8% | 35.6% | 8.9% | 0.7% | ||||||
| В слое | N | 560 064 | 37 124 | 71 437 | 17 257 | 36 612 | 27 133 | 749 627 | |
| 74.8% | 4.9% | 9.5% | 2.3% | 4.9% | 3.6% | ||||
| B | 347.3 | 742.1 | 25.7 | 197.8 | 1.3 | 16.0 | 1330.2 | ||
| 26.1% | 55.8% | 1.9% | 14.9% | 0.1% | 1.2% | ||||
| Bc | 23.2 | 49.5 | 1.7 | 24.7 | 0.1 | 1.5 | 100.7 | ||
| 23.0% | 49.2% | 1.7% | 24.5% | 0.1% | 1.5% | ||||
Рис. 5.
Вертикальное распределение солености (S, PSU), нитратного азота (NO3, μМ) численности (N × 103 кл/л) и биомассы (B, мг/м3) фитопланктона на станциях Индигирского (ст. 5602, 5608) и Колымского (ст. 5612, 5617, 5619) разрезов.
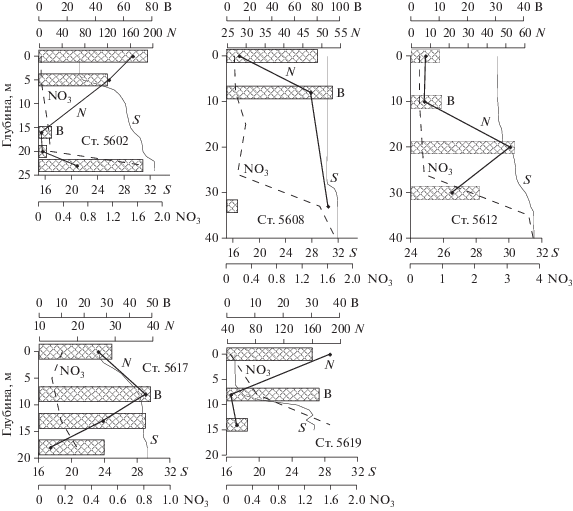
К северу от ст. 5602 во внешней части шельфа вплоть до кромки льда численность фитопланктона уменьшалась. В этом районе по численности доминировали разные виды Chaetoceros: C. diadema (объем клетки от 1000 до 6000 мкм3), С. debilis (от 300 до 2500 мкм3), C. compressus (1000–2000 мкм3), С. socialis (100–200 мкм3) и др. (табл. 2). Все виды Chaetoceros были представлены вегетативными клетками, клетками на разных стадиях спорообразования и спорами. Клетки, готовые к спорообразованию, и споры составляли от 40 до 100% численности и концентрировались, как правило, в нижних слоях водной толщи (рис. 5, табл. 2).
Исключением из общей картины была ст. 5604 в центральной части разреза, где соленость в верхнем слое возрастала до значений >25 PSU, что можно рассматривать как внешнюю границу области речного плюма. Здесь доминировала Rhizosolenia hebetatа f. semispina cо средним объемом клеток ~19 000 мкм3 (табл. 2), ее вклад в общую численность фитопланктона достигал 44%.
Изменения биомассы фитопланктона на разрезе были более резко выражены, чем колебания численности и составляли от 16.4 до 339.1 мг/м3. Низкая биомасса (16–20 мг/м3, ~550 мг/м2) была связана с областью сильно выраженного воздействия речного стока и доминирования мелкоклеточного фитопланктона (южнее ст. 5602), а также отмечена на самой северной станции разреза 5607, сделанной непосредственно у границы многолетнего льда (рис. 4а, табл. 2).
Основной вклад в биомассу на южных станциях вблизи эстуария Индигирки вносили малочисленные крупноразмерные динофлагелляты: Dinophysis acuminatа, D. arctica (средний объем клеток 24 000 и 14 000 мкм3 соответственно), Prorocentrum baltica (2000–2500 мкм3).
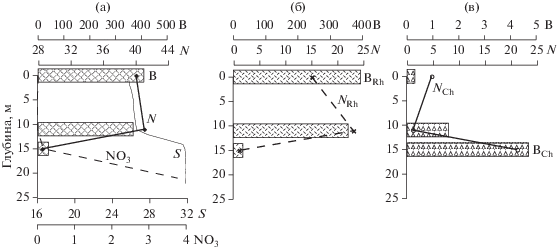
Самая высокая биомасса фитопланктона, 339.1 мг/м3 (5086 мг/м2), зарегистрирована в центральной области шельфа (ст. 5604) и определялась массовым развитием Rhizosolenia hebetatа f. semispina. Вклад вида в общую биомассу составлял 96.1%. Этот же вид вносил наибольший вклад в биомассу на соседних станциях 5602, 5603, 5605 южнее и севернее (табл. 2). Основная часть его популяции была сконцентрирована в верхнем перемешенном слое и в верхней части пикноклина (рис. 6). На станциях 5604 и 5606 равнозначный вклад в биомассу наряду с R. hebetatа f. semispina вносили несколько видов Chaetoceros (рис. 6). На ст. 5608 виды рода Chaetoceros доминировали в биомассе, также как и в численности. На ст. 5607 у кромки льда увеличилась численность крупных гетеротрофных динофлагеллят родов Protoperidinium и Gyrodinium, вклад которых в общую биомассу фитопланктона в столбе воды составлял 52.5% (табл. 2).
Рис. 6.
Вертикальное распределение солености (S, PSU), нитратного азота (NO3, μМ) общей численности (N × 103 кл/л) и биомассы (B, мг/м3) фитопланктона (а), численности (NRh × 103 кл/л) и биомассы (BRh, мг/м3) видов рода Rhizosolenia (б), численности (NCh × 103 кл/л) и биомассы (BCh, мг/м3) видов рода Chaetoceros (в) на ст. 5604 Индигирского разреза.
Между станциями 5607 и 5608 в воде, свободной ото льда, была отобрана проба из поверхностного слоя. Здесь, так же как на ст. 5608, в фитоцене и по числу клеток, и по биомассе доминировали виды рода Chaetoceros, при этом 87% общей численности и 90.2% общей биомассы фитопланктона приходилось на долю С. diadema. По состоянию клеток C. diadema было очевидно, что популяция находилась в начале перехода в покоящуюся стадию: более 40% клеток вида находились в состоянии спорообразования и в виде спор. В пробах в большом количестве встречены мертвые клетки С. diadema. Подобное соотношение вегетативных клеток и спор также наблюдалось в популяции С. debilis.
На двух станциях Индигирского разреза (5602 и 5606) отбор проб был проведен дважды с интервалом в 5 сут, что позволило оценить возможные изменения структурных параметров фитоцена в этих точках.
На ст. 5602_2 общая численность фитопланктона по сравнению с первыми наблюдениями уменьшилась в три раза в основном за счет пятикратного сокращения численности Ollicola vangoorii. При этом вид сохранял доминирование по численности и составлял 44.8% общего числа клеток водорослей (табл. 2) На фоне снижения численности водорослей биомасса фитопланктона увеличилась в два раза, что определялось более чем трехкратным возрастанием обилия крупноразмерного вида Rhizosolenia hebetatа f. semispina. Максимум численности вида был приурочен к верхнему перемешенному слою и верхней части пикноклина.
Повторные наблюдения на ст. 5606 (ст. 5606_2) выявили незначительное (в 1.2 раза) уменьшение общей численности фитопланктона, в основном связанное с сокращением численности динофлагеллят и жгутиковых. Численность Chaetoceros практически не изменилась, и они составляли 74.7% общей численности фитопланктона. Доминирующие виды Chaetoceros diadema, С. debilis и С. socialis были представлены главным образом в виде спор. Основная часть популяций этих видов концентрировалась в нижнем слое водной толщи. На горизонте 33 м виды рода Chaetoceros составляли 96.8% общей численности и 55% общей биомассы фитопланктона.
Параллельно со снижением численности водорослей на ст. 5606_2 по сравнению со ст. 5606 было зарегистрировано увеличение их биомассы в 1.3 раза. Это определялось ростом обилия крупноклеточного вида Rhizosolenia hebetatа f. emispina. Вид занял доминирующее положение в биомассе фитопланктона (55.8% общей биомассы фитоцена, табл. 2) и был сконцентрирован, как и в первоначальных наблюдениях, в верхнем перемешенном слое и в верхней части пикноклина.
Наблюдаемые изменения фитопланктонных сообществ на станциях 5602 и 5606 скорее следует рассматривать как следствие пространственной неравномерности структурных характеристик фитоценоза и адвекции, чем как показатели возможных временных перестроек.
Колымский разрез
В районе, прилежащем к эстуарию Колымы, при поверхностной солености 17–19 PSU и температуре 6.0–6.8°С наблюдалась четко выраженная стратификация водной толщи (рис. 2б). Верхняя граница скачка плотности лежала на глубине 3–6 м. Градиент солености составлял от 1 до 1.6 PSU на метр. Область, занятая речным плюмом (за его внешнюю границу принята поверхностная соленость 25 PSU), на Колымском разрезе имела в два раза меньшую широтную протяженность, чем на Индигирском – 150 и 300 км, соответственно. Между станциями 5617 и 5618 поверхностная соленоcть возрастала на 4 PSU. Далее по разрезу гидрофизические условия становились специфичными, существенно отличными от других районов сибирского арктического шельфа [14–17]. На ст. 5617 верхний однородный слой отсутствовал, слой скачка начинался непосредственно у поверхности и занимал верхние 10 м. Градиент солености в слое скачка не превышал 0.5 PSU на метр. Севернее ст. 5617 на участке акватории широтной протяженностью почти 200 км (станции 5614–5616) вся толща воды от поверхности до дна представляла собой однородный слой с соленостью около 28 PSU (рис. 2б). Практически однородным в этой области было и вертикальное распределение температуры, которая снижалась от 6°С на юге до 3°С на севере. Только на ст. 5613 на глубине 15 м появлялся слой скачка солености и температуры, в котором соленость повышалась на 2.4 PSU, температура понижалась с +3.2 до ‒0.5°С. На станции 5612, ближайшей к кромке льда, слой скачка солености был выражен очень слабо, температура в верхнем перемешанном слое снижалась до <1°C (рис. 2б).
Зарегистрированная гидрофизическая структура на Колымском разрезе позволяет говорить о том, что он пересекал характерную для Восточно-Сибирского шельфа специфическую восточную область, находящуюся под существенным воздействием адвекции вод с востока.
Содержание нитратного азота в верхнем двадцатиметровом слое на Колымском разрезе было значительно ниже, чем на Индигирском разрезе, и варьировало, в основном, от 0.1 до 0.3 μМ. (рис. 3б). Концентрация кремния, напротив, была повсеместно выше и изменялась в пределах от 10 до 28 μМ (рис. 3б). Максимальные концентрации были отмечены в самой южной части разреза в зоне воздействия стока р. Колымы. Концентрации аммонийного азота в верхнем 20-метровом слое варьировали от 0.2 до 0.8 μМ.
Изменения численности фитопланктона на Колымском разрезе превышали порядок величин – от 7.7 × 103 кл/л на ст. 5615 до 90.9 × 103 кл/л на ст. 5619. Величины биомассы варьировали в меньших пределах – от 9.4 мг/м3 на ст. 5613 до 42.1 мг/м3 на ст. 5617 (рис. 4б).
Высокими численностью и биомассой водорослей характеризовался район шельфа, прилежащий к эстуарию Колымы, с соленостью от 17.0 до 19.3 PSU (станции 5618–5620, рис. 2б, 4б). Основу численности здесь составляли три мелкоклеточных вида: Ollicola vangoorii, Skeletonema costatum и Cylindroteca closterium. Наибольший вклад в биомассу вносили несколько видов динофлагеллят родов Gymnodinium и Dynophisis.
Резкие изменения видового состава и биомассы фитопланктона наблюдались сразу же за внешней границей области, в наибольшей степени опресненной стоком Колымы, где соленость возрастала на 4 PSU и практически исчезала вертикальная стратификация водной толщи. На самой внешней границе опресненной области (ст. 5617) зарегистрирована наиболее высокая на разрезе биомасса водорослей – от 42.1 мг/м3 в среднем для столба воды (рис. 4б, табл. 3). По численности и биомассе на ст. 5617 доминировала Dyctiocha speculum – вид-космополит, характерный и для прибрежных, и для открытых вод, редко образующий высокие по плотности скопления. Максимальная численность вида (5.9 × 103–9.1 × 103 кл/л) зарегистрирована на горизонтах 8 и 13 м при солености 26–28 PSU и температуре 5.5–6.0°С. (рис. 5, табл. 3). Далее к северу в области шельфа, где отсутствовала или была слабо выражена стратификация водной толщи, численность и биомасса фитопланктона были существенно ниже. Биомассу определяли малочисленные крупные гетеротрофные виды динофлагеллят. Во всех пробах наблюдалось большое количество мелких гетеротрофных жгутиковых, часто концентрирующихся на отмирающих клетках водорослей. В нижнем слое были обнаружены солоноватоводные виды Actinocyclus оctonarius, Navicula distans, N. directa, Gyrosigma macrum.
Таблица 3.
Колымский разрез. Численность (N, кл/л, %), биомасса (В, мг/м3, %), биомасса в углероде (Вс, мг/м3, %) основных видов/групп фитопланктона на горизонтах отбора проб. Численность (N × 103 кл/м2, %), биомасса (В, мг/м2, %) и биомасса в углероде Вс (мг/м2, %) в столбе воды. Ollicola vangoorii (W. Conrad) Vørs, 1992 (Calycomonas wulffii)
| Станции | Горизонт | Bacillariophyceae | Dyno-phyceae | Silico- phyceae |
Ollicola (Calyco- monas) |
Rest | Общая | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Chaetoceros | Sкеleto-nema | Cylindro-teca | rest | ||||||||
| 5612 | 0 | N | 1072 | _ | 1650 | _ | 1594 | _ | _ | 3107 | 7423 |
| 14.4% | 22.2% | 21.4% | 41.9% | ||||||||
| В | 2.0 | _ | 0.6 | _ | 7.3 | _ | _ | 0.17 | 10.1 | ||
| 19.9% | 5.9% | 72.5% | 1.7% | ||||||||
| 10 | N | 2448 | _ | 2240 | 48 | 592 | _ | 560 | 840 | 6728 | |
| 36.3% | 33.4% | 0.7% | 8.6% | 8.2% | 12.8% | ||||||
| В | 4.0 | _ | 0.9 | 0.53 | 5.14 | _ | 0.02 | 0.17 | 10.8 | ||
| 37.0% | 8.3% | 43.9% | 47.6% | 0.6% | 1.6% | ||||||
| 20 | N | 31 898 | _ | 13 974 | _ | 1700 | _ | _ | 4550 | 52 122 | |
| 61.2% | 26.8% | 3.3% | 8.7% | ||||||||
| В | 27.2 | _ | 5.6 | _ | 3.0 | _ | _ | 0.78 | 36.6 | ||
| 74.3% | 51.3% | 8.2% | 2.2% | ||||||||
| 30 | N | 17 236 | _ | 1428 | _ | 2297 | _ | _ | 357 | 21 318 | |
| 80.8% | 6.7% | 10.8% | 1.7% | ||||||||
| В | 17.9 | _ | 0.6 | _ | 5.7 | _ | _ | 0.1 | 24.3 | ||
| 73.6% | 2.4% | 23.4% | 0.4% | ||||||||
| В слое | N | 435 000 | _ | 177 530 | 480 | 42 375 | _ | 5600 | 71 220 | 732 505 | |
| 59.4% | 24.2% | 0.1% | 5.8% | 0.8% | 9.7% | ||||||
| B | 411.5 | _ | 71.0 | 5.3 | 146.3 | _ | 0.2 | 10.85 | 645.2 | ||
| 63.8% | 11.0% | 0.8% | 22.7% | + | 1.7% | ||||||
| Вс | 27.4 | _ | 4.5 | 0.3 | 20.8 | _ | 0.02 | 1.4 | 54.4 | ||
| 50.3% | 8.2% | 1.8% | 37.2% | + | 2.5% | ||||||
| 5613 | 0 | N | 3877 | _ | _ | 788 | 2768 | _ | _ | 1734 | 9167 |
| 42.3% | 8.6% | 30.2% | 18.9% | ||||||||
| B | 7.3 | _ | _ | 1.83 | 5.5 | _ | _ | 0.5 | 15.13 | ||
| 48.2% | 12.1% | 36.4% | 3.3% | ||||||||
| 15 | N | 5141 | _ | 477 | _ | 1457 | _ | _ | 8611 | 15 686 | |
| 32.8% | 3.0% | 9.3% | 54.9% | ||||||||
| B | 4.7 | _ | 0.2 | _ | 1.6 | _ | _ | 0.43 | 6.9 | ||
| 68.1% | 2.5% | 23.5% | 6.2% | ||||||||
| 25 | N | 2926 | _ | 1672 | 108 | 1290 | _ | _ | 2090 | 8086 | |
| 36.2% | 20.7% | 1.4% | 15.9% | 25.8% | |||||||
| B | 0.2 | _ | 0.7 | 5.13 | 0.35 | _ | _ | 0.47 | 6.85 | ||
| 3.0% | 10.2% | 74.9% | 5.1% | 6.8% | |||||||
| В слое | N | 107 970 | _ | 14 322 | 6450 | 45 422 | _ | _ | 131 092 | 305 256 | |
| 35.4% | 4.7% | 2.1% | 14.9% | 42.9% | |||||||
| B | 114.5 | _ | 5.7 | 39.4 | 63.0 | _ | _ | 11.5 | 234.0 | ||
| 48.9% | 2.4% | 16.8% | 26.9% | 4.9% | |||||||
| Bc | 7.6 | _ | 0.4 | 2.2 | 9.3 | _ | _ | 1.15 | 20.65 | ||
| 36.8% | 1.9% | 10.6% | 45.0% | 5.6% | |||||||
| 5614 | 0 | N | 31 | _ | 750 | _ | 31 | _ | _ | 694 | 1506 |
| 2.0% | 49.8% | 2.5% | 46.2% | ||||||||
| B | 0.08 | _ | 0.3 | _ | 0.3 | _ | _ | 15.9 | 16.6 | ||
| 0.7% | 1.8% | 1.8% | 95.7% | ||||||||
| Bc | 0.06 | _ | 0.02 | _ | 0.04 | _ | _ | 1.4 | 1.52 | ||
| 3.9% | 1.4% | 2.6% | 98.1% | ||||||||
| 5615 | 0 | N | _ | _ | 335 | _ | 2404 | 118 | 670 | 4020 | 7547 |
| 4.4% | 31.8% | 1.6% | 8.9% | 53.3% | |||||||
| В | _ | 0.13 | _ | 16.8 | 0.5 | 0.05 | 2.3 | 19.8 | |||
| 0.6% | 84.8% | 2.5% | 0.5% | 11.6% | |||||||
| 10 | N | 114 | 1378 | 285 | _ | 971 | 256 | 689 | 2015 | 5708 | |
| 2.1% | 24.1% | 5.0% | 17.0% | 4.5% | 12.0% | 35.3% | |||||
| B | 0.05 | 0.15 | 0.1 | _ | 11.8 | 1.02 | 0.01 | 0.14 | 13.3 | ||
| 0.4% | 1.1% | 0.1% | 88.8% | 7.9% | 0.07% | 1.0% | |||||
| 15 | N | _ | _ | 12 210 | 644 | 414 | 140 | 330 | 330 | 14 068 | |
| 86.8% | 4.6% | 2.9% | 1.1% | 2.3% | 2.3% | ||||||
| B | _ | _ | 4.9 | 26.6 | 1.0 | 0.56 | 0.01 | 0.38 | 33.45 | ||
| 14.6% | 79.5% | 3.0% | 1.7% | 0.1% | 1.1% | ||||||
| В слое | N | 855 | 10 335 | 34 337 | 1610 | 20 338 | 2860 | 9343 | 36 037 | 115 715 | |
| 1.0% | 8.9% | 29.6% | 1.4% | 17.6% | 2.4% | 8.0% | 31.1% | ||||
| B | 0.38 | 1.13 | 13.65 | 66.5 | 184.1 | 11.4 | 0.35 | 13.5 | 291.0 | ||
| 0.2% | 0.4% | 4.7% | 22.8% | 63.2% | 3.9% | 0.2% | 4.6% | ||||
| Bc | 0.3 | 0.07 | 0.9 | 4.2 | 23.0 | 1.1 | 0.03 | 1.4 | 31.0 | ||
| 1.0% | 0.3% | 2.9% | 13.5% | 74.2% | 3.5% | 0.2% | 4.5% | ||||
| 5616 | 0 | N | 990 | 87 | _ | 29 | 1911 | 174 | _ | 3144 | 6335 |
| 15.6% | 1.4% | 0.6% | 30.1% | 2.7% | 49.6% | ||||||
| B | 0.3 | 0.01 | _ | 1.0 | 3.2 | 0.7 | _ | 3.17 | 8.38 | ||
| 3.6% | 0.2% | 11.9% | 38.2% | 8.3% | 37.8% | ||||||
| Bc | 0.02 | + | _ | 0.07 | 0.4 | 0.06 | _ | 0.2 | 0.75 | ||
| 2.8% | 9.3% | 53.3% | 8.0% | 26.6% | |||||||
| 5617 | 0 | N | 341 | 5115 | _ | _ | 1624 | 3410 | 7161 | 7843 | 25 494 |
| 1.5% | 20.0% | 6.4% | 13.3% | 28.1% | 30.7% | ||||||
| B | 0.13 | 0.61 | _ | _ | 15.16 | 13.64 | 0.5 | 2.28 | 32.32 | ||
| 0.5% | 1.9% | 46.9% | 42.2% | 1.5% | 7.0% | ||||||
| 8 | N | _ | 7916 | 754 | 29 | 2465 | 9105 | 754 | 16 964 | 37 987 | |
| 20.8% | 2.0% | 0.1% | 6.5% | 24.0% | 2.0% | 44.6% | |||||
| B | _ | 1.0 | 0.3 | 0.15 | 8.17 | 36.4 | 0.03 | 3.44 | 49.5 | ||
| 2.0% | 0.6% | 0.3% | 16.5% | 73.5% | 0.1% | 6.9% | |||||
| 13 | N | 455 | 1365 | 8189 | 35 | 7104 | 5914 | _ | 3640 | 26 702 | |
| 1.78% | 5.1% | 30.7% | 0.2% | 26.6% | 22.1% | 13.6% | |||||
| B | 0.02 | 0.15 | 3.3 | 0.35 | 17.9 | 23.66 | _ | 1.74 | 47.1 | ||
| 0.1% | 0.3% | 7.0% | 0.7% | 38.0% | 50.2% | 3.7% | |||||
| 18 | N | 968 | 1452 | 7744 | 1107 | 525 | 968 | _ | _ | 12 764 | |
| 7.6% | 11.4% | 60.7% | 8.6% | 4.1% | 7.6% | ||||||
| B | 0.05 | 0.06 | 3.1 | 21.45 | 0.43 | 3.8 | _ | _ | 28.9 | ||
| 0.2% | 0.3% | 10.7% | 74.2% | 1.5% | 13.1% | ||||||
| В слое | N | 6059 | 79 069 | 65 206 | 3130 | 59 351 | 104 812 | 33 545 | 159 838 | 514 311 | |
| 1.3% | 15.5% | 12.7% | 0.6% | 11.6% | 20.5% | 6.5% | 31.3% | ||||
| B | 0.74 | 9.6 | 26.2 | 56.3 | 204.3 | 419.0 | 2.2 | 40.2 | 758.5 | ||
| 0.1% | 1.3% | 3.4% | 7.4% | 26.9% | 55.2% | 0.3% | 5.3% | ||||
| Bc | 0.05 | 1.9 | 1.6 | 3.5 | 25.5 | 41.8 | 0.2 | 4.0 | 78.6 | ||
| 0.1% | 2.4% | 2.0% | 4.4% | 32.4% | 53.3% | 0.3% | 5.1% | ||||
| 5618 | 0 | N | _ | 10 346 | _ | _ | 3154 | 726 | 23 648 | 16 997 | 54 871 |
| 18.8% | 5.7% | 1.4% | 43.1% | 31.0% | |||||||
| B | _ | 1.24 | _ | _ | 12.1 | 2.9 | 1.0 | 17.45 | 34.7 | ||
| 3.5% | 34.9% | 8.3% | 3.0% | 50.3% | |||||||
| Bc | _ | 0.08 | _ | _ | 1.5 | 0.24 | 0.08 | 1.6 | 3.5 | ||
| 2.8% | 42.9% | 6.8% | 2.3% | 45.7% | |||||||
| 5619 | 0 | N | _ | 14 553 | _ | 5666 | 1882 | _ | 159 390 | 3111 | 184 602 |
| 7.9% | 3.1% | 1.0% | 86.3% | 1.7% | |||||||
| B | _ | 1.6 | _ | 4.07 | 6.5 | _ | 6.5 | 11.65 | 30.32 | ||
| 5.3% | 13.3% | 21.4% | 21.4% | 38.4% | |||||||
| 8 | N | 1132 | 5712 | 357 | 31 | 4482 | 155 | 29 274 | 3667 | 44 810 | |
| 2.5% | 12.7% | 0.8% | + | 10.0% | 0.3% | 65.5% | 8.2% | ||||
| B | 2.8 | 0.36 | 0.14 | 2.2 | 21.5 | 0.6 | 1.2 | 3.9 | 32.7 | ||
| 8.5% | 1.1% | 0.4% | 6.7% | 65.7% | 1.9% | 3.7% | 11.9% | ||||
| 14 | N | 1040 | 46 276 | 4160 | 80 | _ | _ | 520 | 1039 | 53 115 | |
| 2.0% | 87.6% | 7.8% | 0.15% | 1.0% | 1.9% | ||||||
| B | 0.18 | 3.17 | 1.7 | 1.82 | _ | _ | 0.02 | 0.38 | 7.3 | ||
| 2.5% | 43.5% | 23.4% | 24.9% | 0.3% | 5.3% | ||||||
| В слое | N | 11 044 | 295 236 | 14 979 | 23 121 | 42 334 | 1085 | 844 038 | 41 230 | 1 273 067 | |
| 0.9% | 23.2% | 1.2% | 1.8% | 3.3% | + | 66.3% | 3.2% | ||||
| B | 20.14 | 18.4 | 6.0 | 37.1 | 176.3 | 4.3 | 34.5 | 75.0 | 371.8 | ||
| 5.4% | 4.9% | 1.6% | 10.0% | 47.5% | 1.1% | 9.3% | 20.2% | ||||
| Bc | 1.4 | 1.2 | 0.4 | 2.3 | 22.8 | 0.4 | 2.9 | 7.5 | 38.9 | ||
| 3.6% | 3.1% | 1.0% | 5.9% | 58.7% | 1.0% | 7.4% | 19.3% | ||||
| 5620 | 0 | N | _ | 45 436 | _ | 2537 | 3259 | 614 | 54 488 | 4298 | 110 632 |
| 41.0% | 2.7% | 2.8% | 0.5% | 49.1% | 3.9% | ||||||
| B | _ | 5.68 | _ | 5.74 | 17.6 | 2.45 | 2.2 | 1.2 | 34.9 | ||
| 16.3% | 16.4% | 50.5% | 7.0% | 6.3% | 3.5% | ||||||
| 9 | N | 849 | 9622 | 1698 | 945 | 849 | 568 | 1420 | 2840 | 18 791 | |
| 4.5% | 51.2% | 9.0% | 5.0% | 4.5% | 3.0% | 7.5% | 15.1% | ||||
| B | 0.08 | 0.94 | 0.7 | 1.0 | 1.12 | 2.27 | 0.05 | 1.4 | 7.56 | ||
| 1.0% | 12.4% | 9.2% | 13.2% | 14.8% | 30.1% | 0.8% | 18.5% | ||||
| В слое | N | 3820 | 247 761 | 7641 | 15 669 | 18 486 | 5319 | 251 586 | 32 121 | 582 403 | |
| 0.7% | 42.5% | 1.2% | 2.7% | 3.1% | 0.9% | 43.2% | 5.5% | ||||
| B | 0.36 | 29.8 | 3.15 | 30.3 | 84.24 | 21.2 | 10.1 | 11.7 | 190.8 | ||
| 0.5% | 15.6% | 1.6% | 15.9% | 44.0% | 11.0% | 5.3% | 6.0% | ||||
| Bc | 0.02 | 2.0 | 0.2 | 1.9 | 10.5 | 2.1 | 0.8 | 1.2 | 18.7 | ||
| 0.1% | 10.7% | 1.0% | 10.2% | 43.9% | 11.2% | 4.2% | 6.4% | ||||
Область с относительно высокими количественными характеристиками фитопланктона была отмечена вблизи кромки льда (ст. 5612). (рис. 4б, табл. 3). Здесь основным компонентом фитопланктона были несколько видов рода Chaetoceros (С. diadema, С. debilis, C. gracilis, С. socialis). Также как на Индигирском разрезе в популяциях всех видов Chaetoceros (за исключением С. gracilis) наблюдалось большое число спор и мертвых клеток (до 48% от общего числа живых и мертвых клеток). Максимальная численность и биомасса водорослей рода Chaetoceros зарегистрированы в нижних слоях водной толщи. Появление в фитоцене значительного числа C. gracilis – один из показателей отмирания более крупноразмерных видов, на которых часто поселяются одиночные клетки C. gracilis.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Анализ состава фитопланктона западной и восточной частей Восточно-Сибирского моря в первой декаде сентября показал, что он типичен для Сибирских Арктических морей в летне-осенний сезон [14–19, 27]. Наиболее разнообразными по таксономическом составу группами были динофлагелляты и диатомеи (табл. 1). В [27] отмечено очевидное преобладание диатомовых водорослей в фитоценозах западной части Восточно-Сибирского моря, что, вероятнее всего, связано с использованными методами отбора и фиксации материала.
По численности и биомассе практически повсеместно, кроме области внутреннего шельфа, доминировали диатомеи (табл. 2, 3). Характерным для двух квазимеридональных разрезов, пересекающих области внутреннего и внешнего шельфа в западной и восточной частях бассейна, было доминирование на мелководье вблизи эстуариев Индигирки и Колымы мелкоклеточного фитопланктона, основу которого составляли Ollicola vangoorii (класс Chrysophyceae), Katodinium rotundata (класс Dinophyceae) и мелкая диатомовая водоросль Sceletonema costatum. Эта структурная особенность фитоценоза наблюдалась на фоне солености от 16–18 до 23–24 PSU, резко выраженного слоя скачка плотности/солености на глубине 3–6 м, низкого содержания биогенных элементов и высокой концентрации взвеси. Пресноводная флора на ближайших к эстуариям станциях обоих разрезов не была обнаружена, что, вероятнее всего, связано с тем, что соленость в исследованных районах внутреннего шельфа превышала значения 15 PSU, при которых могут существовать пресноводные виды водорослей [14, 16, 17, 19]. Отсутствие пресноводных видов на внутреннем шельфе юго-западной части Восточно-Сибирского моря при солености >15 PSU явствует и из данных [27].
Численность и биомасса фитопланктона в первой декаде сентября на всей исследованной акватории были крайне низкими, существенно ниже, чем на шельфе Карского моря и моря Лаптевых в аналогичный сезон [14–17]. Об этом же свидетельствуют данные измерений интегральной первичной продукции и концентрации хлорофилла [6, 7].
Биомасса фитопланктона в западной части Восточно-Сибирского моря на Индигирском разрезе была существенно выше, чем восточной части на Колымском разрезе, при сравнимом уровне численности (рис. 4, табл. 2, 3). Соответствующие цифры составляют 16.4–339.1 против 9.4–42.1 мг/м3 и 11.9–66.3 × 106 против 7.7–90.3 × 106 кл/м3. При этом вклад динофлагеллят в биомассу фитоцена на Индигирском разрезе был выше (табл. 2, 3). Высокие концентрации фитопланктона на Индигирском разрезе в основном тяготели к верхнему 5–10 м слою водной толщи, на Колымском разрезе такой тенденции не прослеживалось (рис. 5). Следует отметить очень высокую пространственную изменчивость количественных характеристик фитопланктона на обоих разрезах (рис. 4), что для восточной части Восточно-Сибирского моря подтверждается данными [27].
Через неделю после наших наблюдений в Восточно-Сибирском море (17–20 сентября 2017 г.) нами были проведены работы в западной части моря Лаптевых на разрезе от внутреннего района Хатанского залива до континентального склона. На станциях разреза, выполненных в области с поверхностной соленостью 15–23 PSU, основным компонентом фитопланктона была мелкоклеточная диатомея Sceletonema costatum, входившая в число доминантов в аналогичном биотопе на внутреннем шельфе Восточно-Сибирского моря. Обилие вида достигало 1 × 106 кл/л, а его вклад в общую численность водорослей составлял от 78 до 90% [19]. Вероятно, в осенний период в районах шельфа вблизи эстуарных зон арктических рек с увеличением солености до значений, препятствующих развитию пресноводного фитопланктона, в условиях крайне низкого содержания нитратов преимущество получают мелкоклеточные водоросли разных систематических групп. Возможно, эти виды способны к смешенному питанию, и их развитию способствует высокое содержание аллохтонного растворенного органического вещества, выносимого сибирскими реками.
Полученные нами ранее оценки [19] говорят о том, что в западной части моря Лаптевых в районе смешения морских вод и вод, выносимых Хатангой, при солености 17–19 PSU численность и биомасса фитопланктона были на порядок выше – 0.6–1 × 106 кл/л и 90–160 мг/м3 соответственно, чем в аналогичных районах, прилежащих к устьям Индигирки и Колымы. Это вместе с другими материалами, полученными нами ранее в море Лаптевых [17], свидетельствует о том, что фитопланктон внутреннего шельфа Восточно-Сибирского моря в сезон, свободный ото льда, характеризуется крайней бедностью. Здесь уместно привести количественные характеристики фитопланктона, полученные в тот же сезон (сентябрь) в восточной части Восточно-Сибирского моря на основе тотальных сетных проб [27]. Максимальные значения составляли 0.2–0.4 × 103 кл/л и 0.2–0.3 мг C/м3, что на 1–2 порядка ниже полученных нами значений (рис. 4, табл. 2, 3). Эти различия, очевидно, следствие использованной в [27] методики, приводящей к недоучету значительной части фитопланктона, и приведенные цифры не могут обсуждаться в сравнении с нашими данными.
Полученные материалы показывают, что на внутреннем шельфе Восточно-Сибирского моря на внешней границе области, существенно опресненных речным стоком вод, при поверхностной солености 21–25 PSU формируются локальные благоприятные условия для развития фитопланктона. На Индигирском разрезе (ст. 5602) при солености 21.2 PSU наблюдалось максимальная для разреза численность мелкоклеточного фитопланктона 1.5 × 109 кл/м2 (рис. 2а, 4а, табл. 2). На Колымском разрезе с фронтальной зоной на периферии речного плюма (ст. 5617) при солености 23.3 PSU было ассоциировано самое высокое значение биомассы фитопланктона – 42.1 мг/м3 (рис. 2б, 4б, табл. 3). Оно определялось высокой концентрацией морского вида Dictyocha speculum, вклад которого в общую биомассу фитоцена составлял 55.2%. На этой станции также зарегистрированы максимальное для разреза содержание хлорофилла и высокая первичная продукция [6]. Максимальная биомасса на внешней границе речного плюма (24.3 PSU) отмечена и на Индигирском разрезе: 339 мг/м3 (рис. 4а).
Средняя часть Колымского разреза пересекала область шельфа со специфическими особенностями гидрофизической структуры. В этой области практически отсутствовал скачок плотности, и водная толща была перемешена от поверхности до дна (рис. 2б). Концентрация нитратного азота во всей толще воды была крайне низкой (рис. 3б). Численность и биомасса фитопланктона в этой области снижались до минимальных для всего исследованного района величин: 7.7 × 106 кл/м3 и 9.4 мг/м3 соответственно (рис. 4б, табл. 3).
Биомасса фитопланктона на Индигирском разрезе была существенно выше, чем на Колымском (рис. 4, табл. 2, 3). Максимальные значения биомассы для этих разрезов составляли 339 и 42.1 мг/м3 соответственно. Это было связано с различием в комплексах видов, определявших обилие фитопланктона. В средней части Индигирского разреза на станциях 5602–5606 основу биомассы, а на станции 5604 и основу численности формировала крупноклеточная диатомея Rhizosolenia hebetatа f. semispina (рис. 6, табл. 2). В области срединного шельфа, которую пересекал Колымский разрез, наибольший вклад в численность вносили диатомея Сylindrotheca closterium, динофлагелляты, главным образом рода Gymnodinium, cf. dicrateria (класс Рrimnesiophyceae), и жгутиковые с небольшим размером клеток. Биомассу формировали немногочисленные крупные виды динофлагеллят, такие как Dinophysis acuminata, D. rotundata, несколько видов гетеротрофных Protoperidinium. В нижних слоях в небольшом количестве были встречены солоноватоводные неритические диатомеи Navicula distans, N. directa и Gyrosigma macrum.
На обоих разрезах увеличение численности и/или биомассы фитопланктона наблюдалось в районах вблизи кромки льда. Основным компонентом фитопланктона был комплекс видов рода Chaetoceros. Доминирование видов рода Chaetoceros в приледной области северо-западной части Восточно-Сибирского моря отмечено и в [27]. В нашем материале по биомассе доминировали С. diadema и С. debilis, по численности – С. diademа и С. socialis. Численность и биомасса Chaetoceros в приледной области на Индигирском разрезе была в несколько раз выше, чем на Колымском. Наблюдалось высокое содержание клеток на разных стадиях спорообразования и спор – от 30–40% численности (у С. socialis до 100%), а также до 40% мертвых клеток. Такое физиологическое состояние популяций всех видов Chaetoceros свидетельствовало о переходе их в покоящуюся стадию. При этом клетки Chaetoceros концентрировались в нижних слоях водной толщи.
В целом, полученный материал позволяет говорить о крайней бедности фитопланктона Восточно-Сибирского моря в исследованный безледный сезон по сравнению с другими эпиконтинентальными морями Сибирской Арктики, а также о существенных различиях в структуре фитопланктонных сообществ западной и восточной областей бассейна, что является дополнительным подтверждением его существенной зональной гетерогенности.
Работа выполнена в рамках темы государственного задания № 0128-2021-007, проекта РФФИ “Арктика” № 18-05-60069 и проекта РНФ № 19-17-00196. Экспедиционные исследования проведены при финансовой поддержке Министерства науки и высшего образования РФ (целевое финансирование на проведение морских экспедиционных исследований).
Список литературы
Атлас Арктики. Трешников А. Ф. (ред). М.: Гл. управление геодезии и картографии при Совете Министров СССР, 1985. 204 с.
Георгиев А.А., Георгиева М.Л. Новые данные о фитопланктоне Восточно-Сибирского моря // Труды VI Международной научно-практической конференции “Морские исследования и образование: MARESEDU – 2017”. 2017. Тверь: ПолиПРЕСС. С. 443–446.
Глебов И.И., Надточий В.А., Савин А.Б. и др. Результаты комплексных исследований в Восточно-Сибирском море в августе 2015 г. // Известия ТИНРО. 2016. Т. 186. С. 81–92.
Горбатенко К. М., Кияшко С. И. Состав зоопланктона и трофический статус гидробионтов моря Лаптевых и Восточно-Сибирского моря // Океанология. 2019. Т. 59. № 6, С. 987–997.
Гуков А.Ю., Дударев О.В., Семилетов И.П. и др. Особенности распределения биомассы макробентоса и донных биоценозов в южной части Восточно-Сибирского моря // Океанология. 2005. Т. 45. № 6. С. 889–896.
Демидов А.Б., Гагарин В.И. Первичная продукция и условия ее формирования в Восточно-Сибирском море в осенний период // Докл. АН. 2019. Т. 487. № 6. С. 696–700.
Демидов А.Б., Гагарин. В И., Арашкевич Е.Г. и др. Пространственная изменчивость первичной продукции и хлорофилла в море Лаптевых в августе-сентябре // Океанология. 2019. Т. 59. № 5. С. 755–770.
Добровольский А.Д., Залогин Б.С. Моря СССР. М.: Изд-во МГУ, 1982. 192 с.
Карклин В.П., Карелин И.Д. Сезонная и многолетняя изменчивость характеристик ледового режима морей Лаптевых и Восточно-Сибирского // Система моря Лаптевых и прилегающих морей Арктики. М.: Изд-во МГУ, 2009. С. 187–201.
Никоноров А.М., Иванов В.В., Брызгало В.А. Реки Российской Арктики в современных условиях антропогенного воздействия. Ростов-на-Дону: Изд-во “НОК”. 2007. 280 с.
Павштикс Е.А. Обзор состава и количественного распределения зоопланктона в Восточно-Сибирском море // Исследования фауны морей. Санкт-Петербург, 1994. Т. 48. С. 17–16.
Пинчук А.И. О зоопланктоне Чаунской губы. Экосистемы, флора и фауна Чаунской губы, Восточно-Сибирское море // Исследования фауны морей. Санкт-Петербург, 1994. Т. 47(55). С. 121–127.
Суханова И.Н. Концентрирование фитопланктона в пробе // Современные методы количественной оценки распределения морского планктона. М.: Наука, 1983. С. 97–105.
Суханова И. Н., Флинт М.В., Мошаров С.А. и др. Структура сообществ фитопланктона и первичная продукция в Обском эстуарии и на прилежащем Карском шельфе // Океанология. 2010. Т. 50. № 5. С. 785–800.
Суханова И.Н., Флинт М.Ф., Дружкова Е.И., и др. Фитопланктон северо-западной части Карского моря // Океанология. 2015. Т. 55. № 4. С. 605–619.
Суханова И.Н., Флинт М.В., Сергеева В.М. и др. Структура сообществ фитопланктона Енисейского эстуария и прилежащего Карского шельфа // Океанология. 2015. Т. 55. № 6. С. 935–949.
Суханова И.Н., Флинт М.В., Георгиева Е.Ю. и др. Структура сообществ фитопланктона в восточной части моря Лаптевых // Океанология. 2017. Т. 57. № 1. С. 86–102.
Суханова И.Н., Флинт М.В., Сахарова Е.Г. и др. Фитоценозы Обского эстуария и Карского шельфа в поздневесенний сезон // Океанология. 2018. Т. 58. № 6. С. 882–898.
Суханова И.Н., Флинт М.В., Фёдоров А.В. и др. Фитопланктон Хатангского залива, шельфа и континентального склона западной части моря Лаптевых // Океанология. 2019. Т. 59. № 5. С. 724–733.
Bhavya P.S., Lee J.H., Lee H.W. et al. First in situ estimations of small phytoplankton carbon and nitrogen uptake rates in the Kara, Laptev, and East Siberian seas // Biogeosciences. 2018. V. 15. P. 5503–5517.
Ershova E.A., Kosobokova K.N. Cross‑shelf structure and distribution of mesozooplankton communities in the East‑Siberian Sea and the adjacent Arctic Ocean // Polar Biol. 2019. V. 42. P. 1353–1367. https://doi.org/10.1007/s00300-019-02523-2
Gordeev V.V., Martin J.M., Sidorov J.S. et al. A reassessment of the Eurasian river input of water, sediment, major elements, and nutrients to the Arctic Ocean // American J. Sci. 1996. V. 296. P. 664–691.
Jones, E. P., Anderson L.G., Swift J.H. Distribution of Atlantic and Pacific waters in the upper Arctic Ocean: Implications for circulation // Geophys. Res. Lett. 1998. V. 25. № 6. P. 765–768.
Maslanik JA, Fowler C, Stroeve J. et al. A younger, thinner Arctic ice cover: increased potential for rapid, extensive sea-ice loss // Geophys. Res. Lett. 2007. V. 34. https ://doi.org/ L0320 43https://doi.org/10.1029/2007G
Menden-Deuer S., Lessard E.J. Carbon to volume relationships for dinoflagellates, diatoms and other protist plankton // Limnol. Oceanogr. 2000. V. 45. № 3. P. 569–579.
Nghiem S.V., Chao Y., Neumann G. et al. Depletion of perennial sea ice in the East Arctic Ocean // Geophys. Res. Lett. 2006. V. 33. https://doi.org/10.1029/2006G L0271 98
Polyakova Ye.I., Kryukova I.M., Martynov F.M. et al. Community structure and spatial distribution of phytoplankton in relation to hydrography in the Laptev Sea and the East Siberian Sea (autumn 2008) // Polar Biol. 2021. V. 44. P. 1229–1250.
Semiletov I., Dudarev O., Luchin V. et al. The East Siberian Sea as a transition zone between Pacific-derived waters and Arctic shelf waters. // Geophys Res Lett. 2005.https://doi.org/10.1029/2005GL0224 90
Stein R. Circum Arctic river discharge and its geological record // Int. J. Earth Science. 2000. V. 89. P. 447–449.
Strathmann R.R. Estimating the organic carbon content of phytoplankton from cell volume, cell area or plasma volume // Limnol. Oceanogr. 1967. V. 12. № 3. P. 411–418.
Weingartner T.J., Danielson S., Sasaki Y. et al. The Siberian Coastal Current: a wind- and buoyancy-forced Arctic coastal current. // J Geophys Res Ocean. 1999. V. 104. P. 29 697–29 713. https://doi.org/10.1029/1999J C9001 61
Дополнительные материалы отсутствуют.