

Онтогенез, 2019, T. 50, № 4, стр. 228-236
Сравнительный анализ звукового предупреждающего об опасности сигнала разных возрастных и половых групп рыжеватого суслика (Spermophilus major Pallas 1778)
О. В. Брандлер a, А. Р. Тухбатуллин a, *, А. А. Никольский b
a Федеральное государственное бюджетное учреждение науки Институт биологии развития
им. Н.К. Кольцова РАН
119334 Москва, ул. Вавилова, д. 26, Россия
b Российский университет дружбы народов, экологический факультет
113093 Москва, Подольское ш., д. 8/5, Россия
* E-mail: rusmarmot@yandex.ru
Поступила в редакцию 08.01.2019
После доработки 08.01.2019
Принята к публикации 15.02.2019
Аннотация
На примере большого суслика (S. major) изучалось формирование частотно-временных характеристик звукового сигнала на разных стадиях индивидуального развития. Проверялась гипотеза о вокализации молодых особей наземных беличьих на более низких частотах, чем у взрослых. Результаты исследования свидетельствуют о дифференцированном формировании звукового сигнала у разных полов большого суслика в постнатальном онтогенезе. Звуковой сигнал у самок формируется в течение первого года жизни в период сезона активности, а у самцов на следующий год – после первой зимней спячки. По совокупности исследованных параметров сигнала голос взрослых самок выше, чем у молодых сусликов обоих полов, а взрослых самцов – ниже. Обсуждаются возможные причины и значение выявленных половых и возрастных особенностей звукового сигнала большого суслика.
ВВЕДЕНИЕ
Звуковая коммуникация является одним из важнейших приспособительных механизмов, как позвоночных, так и беспозвоночных животных. Формирование голосового аппарата у млекопитающих происходит на ранних этапах постнатального онтогенеза и тесно связано с периодом полового созревания, что более подробно изучено у человека (Орлова и др., 2013).
Процесс формирования физиологических механизмов звукоизвлечения в ходе индивидуального развития может проявляться в различиях частотных характеристик сигнала у разных возрастных групп, что на эмпирическом уровне воспринимается как разница тональности криков молодых и взрослых особей. На основе многочисленных наблюдений и специальных исследований сформировалось широко распространенное мнение о том, что мелкие молодые животные издают более высокие звуки, чем крупные взрослые особи (Colvin, 1973; Morton, 1977; Tembrock, 1987; Blumstein, Munos, 2005; Никольский, 2007; Grimsley et al., 2011).
На этом фоне неожиданным было сообщение об отсутствии различий в тональности звукового сигнала молодых и взрослых животных у крапчатого суслика Spermophilus suslicus и более высокая частота крика у взрослых животных по сравнению с молодыми у желтого суслика S. fulvus (Matrosova et al., 2007), что противоречит принципу обратной зависимости частоты сигнала от размеров тела источника звука, соответствующему второму закону Ньютона. В проведенном с целью проверки данного феномена исследовании частотных характеристик звуковых сигналов двух возрастных групп у четырех видов грызунов, для которых данный сигнал является характерной формой поведения, малого суслика (S. pygmaeus), степного сурка (Marmota bobak), большой песчанки (Rhombomys opimus) и полевки Брандта (Lasiopodomys brandtii) были получены противоречащие этим данным результаты (Никольский, 2007). У представителей двух семейств грызунов частота предупреждающего об опасности сигнала особей младших возрастных групп оказалась выше частоты сигнала более крупных взрослых особей.
Звуковой предупреждающий об опасности сигнал (далее – сигнал) свойствен многим видам млекопитающих открытых пространств с дневной активностью и высокой плотностью населения. Основной функцией сигнала является предупреждение соседей об опасности. Но также он может передавать информацию о половой принадлежности источника сигнала, его возрасте и положении в пространстве. Данный сигнал является характерной формой адаптивного поведения наземных беличьих – сусликов и сурков. Видовая специфика структуры сигнала генетически детерминирована и остается постоянной в разные возрастные периоды, что позволяет использовать его в качестве диагностического признака (Никольский, 1969, 1984).
Учитывая высокую адаптивную значимость звукового предупреждающего об опасности сигнала для наземных беличьих, вопрос об основах формирования его частотно-временных характеристик в онтогенезе представляет существенный интерес для понимания биологии данной группы. Полученные к настоящему времени противоречивые результаты требуют проведения дополнительных исследований на других близкородственных видах, имеющих свою независимую эволюционную историю. С этой целью нами было проведено изучение частотно-временных характеристик сигнала большого, или рыжеватого, суслика S. major Pallas, 1778 в качестве модельного объекта для исследования формирования звуковой коммуникации в онтогенезе.
МАТЕРИАЛЫ И МЕТОДЫ
Материалом для исследования послужили записи сигнала 107 особей рыжеватого суслика, собранные в 2016–2018 гг. в период со второй декады июня по третью декаду июля. Отлов сусликов и запись звуковых сигналов проведены в 39 популяциях на большей части ареала S. major, в том числе на территории России в Курганской области – в 12 локалитетах, Оренбургской обл. – 41, Самарской обл. – 7, Челябинской обл. – 16, Республике Башкортостан – 22, Республике Татарстан – 3, а также в Казахстане в Актюбинской обл. – 2, Западно-Казахстанской обл. – 2, Кустанайской обл. – 2.
Животных отлавливали во время дневной активности с помощью сетчатых живоловок 60 × × 27.5 × 40 см с падающей дверцей, устанавливаемых на выходных норовых отверстиях или выливанием водой из кормовых нор. Суслики после отлова помещались в сетчатые клетки размером 40 × 17.5 × 17.5 см с подстилкой из сена или (на короткое время) оставались в живоловках. Звуковой предупреждающий об опасности сигнал сусликов записывали непосредственно после поимки или спустя 0.5–24 ч. Запись криков особей, отловленных при помощи воды, производили после того, как суслики высохнут. Суслики издавали крики самопроизвольно или в ответ на движения оператора. Запись осуществляли с помощью цифрового магнитофона “Marantz-PMD 660” (Япония) и конденсаторного микрофона “Audio-Technica AT897” (Япония) в несжатом цифровом формате Wave (WAV) с частотой дискредитации 48 кГц. Расстояние от объекта до микрофона составляло 1–2 м. Животных взвешивали на электронных портативных весах WeiHeng (Китай) с точностью до 10 г, производили стандартные линейные измерения с помощью штангенциркуля, определяли пол и возраст.
Животные были разделены на три возрастные группы: взрослые (старше одного года, adultus, ad), полувзрослые (subadultus, sad) и молодые (juvenilis, juv) в соответствии с состояниянием шерстного покрова, относительной длиной и массой тела, состоянием наружных половых признаков (Клевезаль, 2007). Выборка состояла из 107 особей, включая 7 ♂♂ и 33 ♀♀ ad, 17 ♂♂ и 26 ♀♀ sad, 8 ♂♂ и 12 ♀♀ juv, а также 1 ♂ и 3 ♀♀ переходного между juvenilis и subadultus возраста.
Прежде чем приступить к анализу звуков, все фонограммы прослушивали и визуализировали в программе SpectraPLUS 5.0 (Pioneer Hill Software LLC, USA). В анализ отбирались сигналы без заметных искажений, издаваемые животными в устойчивом ритме. Для 87 особей проанализированы серии по 10 криков. В анализ также включены серии, содержащие менее 10 пригодных для анализа криков: по 9 криков от 5 особей, по 8 от 5, по 7 от 2, по 6 от 4, по 5 от 3, по 4 от 1. Всего в статистическую обработку включены 1012 криков. Спектральный анализ проведен в программе SpectraPLUS 5.0, с помощью которой определялись максимум основной (Fo) и доминантная (пиковая) (Fd) частоты звукового сигнала (Никольский, 2007). В программе Sonic Visualiser 2.5 (Chris Cannam and Queen Mary, UK) были определены значения основной частоты в начале сигнала (стартовой частоты, Fs) и длительность от начала сигнала до достижения максимума основной частоты (начальная длительность, t1) (рис. 1). Начальная длительность соответствует суммарной длительности 1-й и 2-й из четырех фаз частотной модуляции сигнала большого суслика (Никольский, Румянцев, 2004).
Рис. 1.
Сонограмма предупреждающего об опасности звукового сигнала большого суслика. Стрелками показано положение стартовой (Fs), доминантной (Fd), максимума основной (Fo) частот и начальная длительность (t1) сигнала.
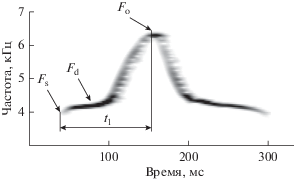
Для анализа зависимости параметров звукового сигнала от возраста в качестве размерной характеристики, связанной с возрастом, использовалась не только масса тела (М), как в предыдущих исследованиях (Matrosova et al., 2007; Volodina et al., 2010), но и длина (L) животных, которая является более стабильной характеристикой, чем масса, зависимая от множества факторов метаболизма животных. Связь длины и массы тела с параметрами звукового сигнала определяли с помощью коэффициента детерминации R2.
Все полученные распределения частотных и временных характеристик звукового сигнала отличались от нормального (тест Колмогорова–Смирнова, p < 0.01), поэтому для оценки влияния пола и возраста на исследованные параметры сигнала мы использовали факторный дисперсионный анализ (ANOVA). В качестве категориальных факторов использованы пол и возраст, задавался сигма-ограниченный тип параметризации модели при VI типе сумм квадратов (SS). Различия считали достоверными при p < 0.05. Статистическая обработка данных выполнена в программе STATISTICA 8.0. (StatSoft, Inc. Tulsa, OK, USA).
РЕЗУЛЬТАТЫ
Длина и масса тела сусликов увеличиваются с возрастом и значимо различаются в разных возрастных группах (ANOVA, p = 0.00, табл. 1). Это позволило нам использовать данные признаки в качестве размерных характеристик при анализе зависимости параметров звукового сигнала от возраста. Попарный корреляционный анализ показал, что ни один исследуемый параметр звукового сигнала большого суслика не находится в зависимости от длины или массы тела в значительной степени (значения R2 находятся в интервале от 0.0 до 0.2). Низкие значения коэффициента детерминации могут указывать на неоднородность выборки и свидетельствовать о влиянии других факторов на частотно-временную структуру сигнала.
Таблица 1.
Средние значения ($\bar {x}$ ± s.e.; минимум–максимум) длины тела L, массы тела М, максимума основной Fo, доминантной Fd, стартовой Fs частот, начальной длительности t1 предупреждающего об опасности звукового сигнала трех возрастных групп S. major
| juv | sad | ad | ||||
|---|---|---|---|---|---|---|
| самцы | самки | самцы | самки | самцы | самки | |
| L, мм | 194 ± 7.2 172–218 |
195 ± 5.5 160–219 |
233 ± 5.7 206–262 |
236 ± 3 213–249 |
281 ± 7.1 254–300 |
274 ± 2.4 251–300 |
| М, г | 263 ± 13 155–485 |
271 ± 8.3 125–435 |
395 ± 5.4 285–520 |
439 ± 5.4 300–660 |
783 ± 20.4 650–1015 |
654 ± 4.8 455–865 |
| Fo, Гц | 5999 ± 69.4 4746–7460 |
5984 ± 52.3 4597–7394 |
6000 ± 38.4 5175–6961 |
6278 ± 28.9 5168–7187 |
5872 ± 100.7 4345–6898 |
6283 ± 33.1 4514–7586 |
| Fd, Гц | 4117 ± 69.2 3468–7265 |
4203 ± 29.5 3750–5531 |
4057 ± 25 3421–6188 |
4299 ± 32.5 3469–6750 |
4208 ± 97.3 3563–6468 |
4342 ± 36.6 3281–7219 |
| Fs, Гц | 3886 ± 48.2 3309–4905 |
3996 ± 19 3456–4835 |
3797 ± 24.2 3107–4423 |
4004 ± 23.2 3192–5098 |
3704 ± 40.3 2930–4229 |
3978 ± 20.4 3142–5030 |
| t1, мс | 101 ± 1.6 71–130 |
97 ± 1.2 68–133 |
122 ± 2.1 80–190 |
115 ± 1.2 80–181 |
125 ± 2.1 97–170 |
120 ± 1.3 46–217 |
| Fd/Fo | 0.687 ± 0.007 0.556–0.996 |
0.709 ± 0.008 0.590–0.997 |
0.678 ± 0.003 0.599–0.987 |
0.686 ± 0.005 0.547–0.999 |
0.724 ± 0.02 0.583–0.997 |
0.693 ± 0.005 0.522–0.999 |
| Fo–Fs, Гц | 2113 ± 49.9 1362–2880 |
1988 ± 49.3 501–3123 |
2203 ± 23.3 1630–3005 |
2244 ± 19.6 1401–3056 |
2168 ± 81.9 664–3005 |
2306 ± 27.5 941–3330 |
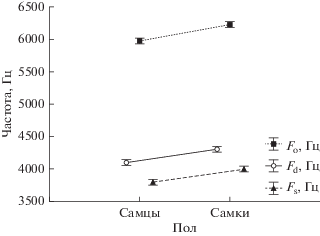
Известно, что как частотные, так и временные параметры звукового сигнала у млекопитающих часто зависят от пола (Lapshina et al., 2012), поэтому нами был проведен анализ влияния пола на значения начальной, доминантной и максимума основной частот (рис. 2). Результаты однофакторного дисперсионного анализа указывают на то, что все три частотные характеристики сигнала у самцов достоверно ниже (p < 0.05) по сравнению с самками.
Рис. 2.
Влияние возраста на значения начальной (Fs), доминантной (Fd) и максимума основной (Fo) частот (однофакторный анализ ANOVA), вертикальные штрихи – значения стандартной ошибки (s.e.).
Для оценки влияния обоих факторов (пола и возраста) на структуру сигнала большого суслика проведен многофакторный дисперсионный анализ. Поскольку оценка зависимости исследуемых характеристик сигнала показала низкую связь данных признаков между собой (табл. 2), оценка пола и возраста проведена для каждой характеристики сигнала независимо (рис. 3). Было обнаружено влияние обоих факторов на показатели Fo, Fs и t1 (p < 0.05). Значения максимума основной частоты (Fo) у молодых особей обоих полов совпадают, у полувзрослых самцов остаются на том же уровне, а у самок становятся выше на 300 Гц и остаются такими у взрослых животных. У взрослых же самцов это значение понижается на 150 Гц (рис. 3б). Значения начальной частоты сигнала (Fs) имеют половые различия во всех возрастных группах (рис. 3в). При этом у самок всех возрастов эти значения находятся в одном диапазоне, а у самцов линейно снижаются. Начальная длительность сигнала меньше у самок по сравнению с самцами во всех возрастных группах (рис. 3г). У обоих полов она достоверно короче у молодых особей по сравнению с полувзрослыми и взрослыми.
Таблица 2.
Коэффициенты корреляции r, p < 0.05, n = = 1012 (над диагональю) и детерминации R2 (под диагональю) частотных и временных характеристик звукового сигнала большого суслика (обозначения как в табл. 1)
| Fd | Fo | Fs | t1 | |
|---|---|---|---|---|
| Fd | 0.46 | 0.56 | –0.18 | |
| Fo | 0.21 | 0.63 | –0.09 | |
| Fs | 0.32 | 0.39 | –0.35 | |
| t1 | 0.03 | 0.01 | 0.12 |
Рис. 3.
Зависимость характеристик звукового предупреждающего об опасности сигнала большого суслика от пола и возраста (ANOVA): а – доминантной частоты; б – максимума основной частоты; в – стартовой частоты; г – начальной длительности; вертикальные штрихи – значения стандартной ошибки (s.e.).
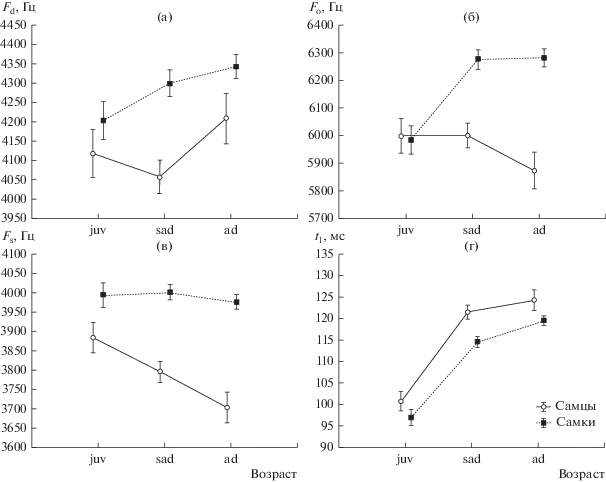
Для доминантной частоты (Fd) обнаружена достоверная связь с полом, в то время как влияние возраста в суммарной выборке недостоверно (p > 0.05) (рис. 3а). Диапазоны доминантной частоты у молодых самок и самцов существенно перекрываются. У полувзрослых самцов эта характеристика становится несколько ниже, а у взрослых выше, чем у молодых, в то время как у самок она последовательно повышается.
Значение Fd в пределах всей выборки достоверно ниже Fo (табл. 1). Однако у отдельных особей наблюдаются нетипичные значения Fd, приближающиеся к Fo. Этот феномен наблюдался у 20 животных. Число таких нетипичных сигналов в сериях колеблется от 1 до 7 (в среднем 2.35). Всего в общей выборке 47 таких сигналов (4.6%). Среднее значение Fd в этих сигналах составляет 6127 ± 72 Гц, что превышает на 2 кГц среднее значение по всей выборке без их учета (4150 ± 11). Среди сусликов, подающих сигналы с аномально высокой доминантной частотой, большинство составляют полувзрослые и взрослые самки (7 и 6 соответственно), что, вероятно, связано с преобладанием этих категорий особей в общей выборке.
ОБСУЖДЕНИЕ
Результаты нашего исследования свидетельствуют о дифференцированном формировании звукового сигнала у разных полов большого суслика в процессе роста животных. Если у молодых животных обоих полов все исследованные параметры сигнала совпадают или почти совпадают, то у полувзрослых и взрослых самцов и самок они значительно различаются. Сигнал полувзрослых самцов в целом более низкий, чем у самок, но его основные параметры (Fd и Fo) практически не отличаются от параметров сигнала молодых животных, и лишь начало крика становится несколько ниже. У полувзрослых самок, напротив, сигнал в целом выше, чем у молодых и становится практически сходным с сигналом взрослых самок по всем частотным характеристикам. Сигнал взрослых самцов существенно отличается от сигнала полувзрослых самцов и самок старших возрастных групп. Взрослые самцы начинают кричать на более низкой частоте, чем животные всех остальных половозрастных категорий (рис. 3в). Наиболее высокая область крика (Fo) у них также ниже, чем у животных других категорий (рис. 3б). При этом наибольшая энергия сигнала у взрослых самцов сосредоточена в более низком диапазоне частот (Fd), чем у взрослых самок, но в более высоком, чем у молодых и полувзрослых самцов (рис. 3а).
Практически весь диапазон частотного спектра и доминантная частота, имеющая максимальную амплитуду в спектре сигнала большого суслика, сосредоточены в начальной части сигнала, длительность которого (t1) у взрослых сусликов в среднем больше на 21.5 мс, чем у молодых. У самок всех возрастов этот показатель несколько ниже, чем у самцов соответствующих возрастных групп (рис. 3г). При этом частотный диапазон звукового сигнала (Fo–Fs) у самок с возрастом расширяется и у взрослых увеличивается на 300 Гц по сравнению с молодыми (табл. 1), в отличие от самцов, у которых диапазон остается практически одинаковым на протяжении всей жизни.
В целом, звуковой сигнал у самок большого суслика формируется в течение первого года жизни в сезон активности, до залегания в спячку, а у самцов он, вероятно, приобретает стабильные характеристики на следующий год – после первой зимней спячки. Нам не известны факторы, определяющие эти различия. Естественно предположить, что как временные, так и частотные характеристики звуковых сигналов могут зависеть от множества причин. В частности, половые различия в формировании звукового сигнала могут являться как отражением неравномерности скорости генеза отдельных органов голосового аппарата, так и различий в гормональном статусе у полов в онтогенезе, влиянием которых можно объяснить общее снижение основной частоты у самцов и ее увеличение у самок, а также расширение частотного диапазона звукового сигнала с возрастом у самок. Влияние стероидов на половую дифференциацию поведенческих реакций в период полового созревания было обнаружено у суслика Белдинга (S. beldingi) (Holekamp, Sherman, 1989). Влияние гормонального статуса на звуковую активность и структуру сигнала была неоднократно показана для многих видов млекопитающих (Никольский, 1992).
Обнаруженные нами половые различия в структуре звукового сигнала большого суслика противоречат имеющимся данным. Ранее для большого и краснощекого (S. erythrogenys) сусликов было показано отсутствие половых различий по длительности, максимальной и минимальной основным частотам (Жилин, 2002). Также не обнаружена зависимость доминантной, максимальной и минимальной основных частот от пола у крапчатых (S. suslicus) (Matrosova et al., 2011; Матросова и др., 2012) и желтых (S. fulvus) сусликов (Matrosova et al., 2007, 2011), но обнаружена у желтобрюхих сурков (Marmota flaviventris), представителей того же, что и суслики, подсемейства (Blumstein, Munos, 2005). Несовпадение результатов, полученных разными авторами на разных видах, может быть обусловлено как методическими особенностями исследований, так и различиями онтогенеза у разных видов.
Вопрос о том кричат ли молодые суслики ниже, т.е. “басовитее”, взрослых (Matrosova et al., 2007) необходимо решать с учетом выявленных половых отличий в развитии вокализации. Мы можем констатировать, что у молодых самок две изученные частотные характеристики (Fd, Fo) ниже, чем у взрослых, в то время как у самцов только доминантная частота незначительно изменяется в этом направлении, а две другие характеристики с возрастом снижаются. Таким образом, голос взрослых самок в целом выше, чем у молодых сусликов, а взрослых самцов – ниже, то есть на поставленный выше вопрос в отношении самок большого суслика можно ответить положительно, а в отношении самцов – отрицательно. Поиск возрастных различий в частотных характеристиках звуковых сигналов животных часто увязывают с проверкой принципа обратной зависимости частоты сигнала от размеров тела источника звука. Данные, имеющиеся по наземным беличьим в настоящее время противоречивы. Прямая зависимость основной частоты сигнала от возраста (=размеров тела) обнаружена у желтого суслика (S. fulvus), но не обнаружена у крапчатого (S. suslicus) суслика (Matrosova et al., 2007) и суслика Ричардсона (S. richardsonii) (Swan, Hare, 2008). При этом в другом исследовании такая зависимость у желтых сусликов не обнаружена (Matrosova et al., 2011). В то же время у малого суслика (S. pygmaeus) и у значительно более крупных, чем суслики, степного (M. bobak) и желтобрюхого (M. flaviventris) сурков найдена обратная зависимость частоты сигнала от размеров тела: молодые особи у этих видов издают более высокие звуки, чем взрослые (Blumstein, Munos, 2005; Никольский, 2007). В отношении сусликов можно предположить, что на результаты исследований оказало влияние то, что общая выборка делилась на две возрастные группы – молодых (первого года жизни) и взрослых (после первой зимовки и старше). В то время как результаты нашего исследования показали, что у полувзрослых сусликов на первом году жизни характеристики звукового сигнала сходны с сигналом взрослых особей или имеют промежуточное значение.
Исследования зависимости акустического сигнала от размеров тела у грызунов обнаружили несколько механизмов, определяющих разнообразие моделей, описывающих эту зависимость (García-Navas, Blumstein, 2016; Никольский, 2017). Полученные нами результаты также свидетельствуют в пользу того, что не только размеры тела, но и другие факторы, особенности развития морфологических структур голосового тракта, способов генерации звука, возможность управления акустическими колебаниями и др., могут оказывать значительное влияние на частотно-временные характеристики звуков, издаваемых животными (Matrosova et al., 2007; Никольский, 2017).
Частота и длительность звуковых сигналов, принадлежащих животным разного возраста, находится в зависимости от объема легких, степени развития мускулатуры, участвующей в выдохе, и морфологических особенностей звукового тракта (гортани, голосовых связок, ротовой полости), которые у молодых животных менее развиты, чем у взрослых (Hammerschmidt et al., 2000; Fitch, Hauser, 2002; Lapshina et al., 2012). Возможно, этим можно объяснить повышение с возрастом доминантной частоты и начальной длительности сигнала, требующих относительно больше энергии, расходуемой животным при звукоизвлечении, что связано с увеличением скорости продувания струи и объема выдыхаемого воздуха.
Полученные нами результаты выявляют определенные тенденции возрастных изменений звукового сигнала большого суслика, который так же, как, по-видимому, и у других видов сусликов, характеризуется широкой индивидуальной изменчивостью. На это указывают интервалы разброса значений по всем изученным показателям, которые частично перекрываются (табл. 1). Частотно-временной спектр каждого крика во многом зависит от ситуативной реакции животного и его физиологического состояния. Возможно, именно этими причинами можно объяснить продуцирование отдельными особями криков с нетипично высокой доминантной частотой. Это указывает на то, что суслики способны управлять формой акустических колебаний, перераспределяя энергию в пределах частотного спектра. Способность активно управлять структурой звуковых сигналов в процессе вокализации известна для многих видов млекопитающих (Никольский, 1984). Для нашего объекта характерен относительно широкий диапазон частотной модуляции (рис. 1), что открывает широкие возможности управления спектром сигнала.
Можно предположить, что широкая изменчивость структуры звуковых сигналов большого суслика, связанная с неравномерностью возрастных и половых изменений их отдельных характеристик, может создавать индивидуальное звучание крика и дифференцированно восприниматься конспецифическими особями. По крайней мере, звуковые сигналы взрослых самцов и самок хорошо отличаются по всем изученным частотно-временным характеристикам друг от друга и от сигнала молодых животных. Возможно, на этом основана способность различать по звуковому сигналу особей разных возрастов калифорнийским сусликом (S. beecheyi) (Hanson, Coss, 2001) и отдельных индивидуумов сусликом Ричадсона (S. richardsonii) (Hare, 1998). Также конспецифические особи могут получать информацию об эмоциональном состоянии источника сигнала, воспринимая его необычные крики, такие, как обнаруженные нами с нетипично высокой доминантной частотой.
Адаптивное значение выявленных половозрастных особенностей предупреждающего об опасности звукового сигнала большого суслика не очевидно. В связанных с данной темой работах активно обсуждается несколько аспектов селективного преимущества тех или иных описанных особенностей вокализации наземных беличьих. В частности, сближение частотного спектра сигнала молодых и взрослых особей или более низкая частота вокализации молодых рассматривается как адаптация, которая: а) улучшает распространение сигнала в открытой среде (Matrosova et al., 2007; Никольский, 2017), б) ограничивает распространение более высокого звука с целью снижения вероятности обнаружения хищником кричащего зверька или в) является вокальной мимикрией, когда молодые зверьки подражают взрослым, чтобы избежать нападения хищника (Matrosova et al., 2007). Ни одна из этих гипотез пока не получила подтверждения. Наши результаты также не поддерживают явным образом ни одну из них. По логике первой гипотезы повышение частоты сигнала с возрастом должно понижать его помехоустойчивость. Две остальные гипотезы представляются также недостаточно обоснованными, в особенности последняя (вокальной мимикрии). Учитывая многообразие способов узнавания жертвы хищником, нам трудно согласиться, что выравнивание частотных характеристик крика жертв разных возрастов представляет серьезное препятствие для хищников. Кроме того, возрастные изменения сигнала у большого суслика происходят разнонаправленно у разных полов и таким образом должны иметь взаимоисключающую адаптивную ценность. На низкий селективный пресс указывает также и широкий диапазон изменчивости по всем изученным признакам звукового сигнала.
ЗАКЛЮЧЕНИЕ
В результате нашего исследования впервые для сусликов рода Spermophilus было обнаружено, что становление звукового сигнала в постнатальном онтогенезе у самцов и самок значительно различается. Формирование на основе этого феномена широкого спектра индивидуальной изменчивости издаваемых криков может играть существенную роль во внутрипопуляционной коммуникации таких колониальных животных, как наземные беличьи. Для выяснения факторов, определяющих обнаруженные особенности половозрастной динамики частотно-временных характеристик звукового сигнала, необходимы дальнейшие исследования гормонального статуса, динамики органогенеза и физиологических функций в этой группе грызунов. Звуковой предупреждающий об опасности сигнал является хорошим модельным объектом для исследования индивидуального развития коммуникативного поведения животных.
Работа поддержана грантом Российского фонда фундаментальных исследований (№ 16-04-01826).
Список литературы
Жилин М.Е., Суслики Южного Зауралья (распространение, вокализация, межпопуляционные особенности). Дис. … канд. биол. наук. Курган: Курганский гос. ун-т, 2002. 121 с.
Клевезаль Г.А. Принципы и методы определения возраста млекопитающих. М.: КМК, 2007. 283 с.
Матросова В.А., Пиванова С.В., Савинецкая Л.Е. и др. Межпопуляционная изменчивость крика тревоги крапчатого суслика (Spermophilus suslicus, Rodentia, Sciuridae): влияние пола, возраста и массы тела // Зоол. журн. 2012. Т. 91. № 4. С. 453–453.
Никольский А.А. Фонотипы наземных беличьих Палеарктики // Млекопитающие. Тез. ко Второму Всес. Совещ. по млекопитающим. Новосибирск, 1969. С. 32–36.
Никольский A.A. Звуковые сигналы млекопитающих в эволюционном процессе. M: Наука, 1984. 199 с.
Никольский А.А. Экологическая биоакустика млекопитающих. М.: Изд-во МГУ, 1992. 120 с.
Никольский A.A. Сравнительный анализ частоты звукового предупреждающего об опасности сигнала разных возрастных групп грызунов // Зоол. журн. 2007. Т. 86. № 4. С. 499–504.
Никольский А.А. Две модели зависимости частоты звукового сигнала от размеров тела животного на примере сусликов Евразии (Mammalia, Rodentia) // Доклады Академии наук. 2017. Т. 477. № 3. С. 375–379.
Никольский А.А., Румянцев В.Ю. Изменчивость звукового сигнала сусликов группы major (Rodentia, Sciuridae, Spermophilus) как модель географического видообразования // Зоол. журн. 2004. Т. 83. № 8. С. 1008–1017.
Орлова О.С., Эстрова П.А., Калмыкова А.С. Особенности развития детского голоса в онтогенезе // Спец. образован. 2013. № 4. С. 92–104.
Blumstein D.T., Munos O. Individual, age and sex-specific information is contained in yellow-bellied marmot alarm calls // Anim. Behav. 2005. V. 69. № 2. P. 353–361.
Colvin M.A. Analysis of acoustic structure and function in ultrasounds of neonatal Microtus // Behavior. 1973. V. 44. № 3/4. P. 234–263.
Fitch W.T., Hauser M.D. Unpacking “honesty”: vertebrate vocal production and the evolution of acoustic signals // Acoustic Communication, Springer Handbook of Auditory Research. N.Y.: Springer, 2002. P. 65–137.
García-Navas V., Blumstein D.T. The effect of body size and habitat on the evolution of alarm vocalizations in rodents // Biological Journal of the Linnean Society. 2016. V. 118. № 4. P. 745–751.
Grimsley J.M.S., Monaghan J.J.M., Wenstrup J.J. Development of social vocalizations in mice // PLoS One. 2011. V. 6. Is. 3. e17460.
Hammerschmidt K., Newman J.D., Champoux M. et al. Changes in rhesus macaque ‘coo’ vocalizations during early development // Ethology. 2000. V. 106. № 10. P. 873–886.
Hanson M.T., Coss R.G. Age differences in the response of California ground squirrels (Spermophilus beecheyi) to conspecific alarm calls // Ethology. 2001. V. 107. № 3. P. 259–275.
Hare J.F. Juvenile Richardson’s ground squirrels, Spermophilus richardsonii, discriminate among individual alarm callers // Animal Behaviour. 1998. V. 55. № 2. P. 451–460.
Holekamp K.E., Sherman P.W. Why male ground squirrels disperse: a multilevel analysis explains why only males leave home // Amer. Scient. 1989. V. 77. № 3. P. 232–239.
Lapshina E.N., Volodin I.A., Volodina E.V. et al. The ontogeny of acoustic individuality in the nasal calls of captive goitred gazelles, Gazella subgutturosa // Behav. Proces. 2012. V. 90. № 3. P. 323–330.
Matrosova V.A., Blumstein D.T., Volodin I.A. et al. The potential to encode sex, age and individual identity in the alarm calls of three species of Marmotinae // Naturwissenschaften. 2011. V. 98. P. 181–192.
Matrosova V.A., Volodin I.A., Volodina E.V. et al. Pups crying bass: vocal adaptation for avoidance of age-dependent predation risk in ground squirrels? // Behav. Ecol. Sociobiol. 2007. V. 62. P. 181–191.
Morton E.S. On the occurrence and significance of motivation–structural rules in some bird and mammal sounds // The Amer. Natur. 1977. V. 111. № 981. P. 855–869.
Swan D.C., Hare J.F. Signaler and receiver ages do not affect responses to Richardson’s ground squirrel alarm calls // J. Mam. 2008. V. 89. № 4. P. 889–894.
Tembrock G. Verhaltensbiologie. Jena: VEB Gustav Fischer Verlag, 1987. 363 p.
Volodina E.V., Matrosova V.A., Volodin I.A. An unusual effect of maturation on the alarm call fundamental frequency in two species of ground squirrels // Bioacoustics. 2010. V. 20. P. 87–98.
Дополнительные материалы отсутствуют.