

Онтогенез, 2019, T. 50, № 4, стр. 247-252
Индивидуальный рост Anodonta beringiana (Unionidae, Bivalvia) в постличиночном онтогенезе
А. А. Зотин a, *, И. Ю. Попов b, c
a Институт биологии развития им. Н.К. Кольцова РАН
119334 Москва, ул. Вавилова, 26, Россия
b Санкт-Петербургский государственный университет
199034 Санкт-Петербург, Университетская наб., 7/9, Россия
c Национальный медицинский исследовательский центр онкологии имени Н.Н. Петрова
197758 пос. Песочный, Санкт-Петербург, ул. Ленинградская, 68, Россия
* E-mail: zotin@idbras.ru
Поступила в редакцию 16.12.2018
После доработки 15.03.2019
Принята к публикации 25.03.2019
Аннотация
Проведено исследование индивидуального линейного роста 26 особей беззубки берингийской Anodonta beringiana путем измерения последовательных годовых колец на поверхности раковины. Показано, что рост моллюсков хорошо описывается уравнением роста Берталанфи. Коэффициенты этого уравнения у разных особей достоверно не различаются и составляют в среднем k = 0.062 ± 0.011 год–1; L∞ = 155.8 ± 19.5 мм. Видовая продолжительность жизни, определяемая произведением константы роста k на максимальную продолжительность жизни моллюсков в популяции, равна 1.7. Эта величина близка к средней видовой продолжительности жизни у других видов двустворчатых моллюсков.
ВВЕДЕНИЕ
Изучение индивидуального роста связано с необходимостью одновременного определения возраста и размеров животных на разных стадиях онтогенеза. Это создает определенные трудности при проведении подобного рода исследований в естественных условиях обитания. Их удается избежать в случае двустворчатых моллюсков. Измерение длин годовых колец на поверхности раковины позволяет получить данные о размерах моллюсков для разных возрастов и построить индивидуальную кривую роста. Под длинами колец понимается расстояние между краями кольца, наиболее близкими к переднему и заднему краям раковины.
Для описания роста моллюсков в большинстве случаев используют уравнение Берталанфи (Bertalanffy, 1960):
где t – номер измеренного годового кольца, начиная от макушки раковины; t0 – возраст моллюска, для годового кольца t = 0; Lt – длина годового кольца в возрасте t + t0; L∞ – предельное значение длины раковины; k – константа роста.Использование коэффициентов уравнения (1) позволяет проводить сравнительные внутрипопуляционные, межпопуляционные и межвидовые исследования роста животных.
Было показано, что константа роста находится в обратной зависимости от максимальной продолжительности жизни животных Tmax (Bauer, 1992). Об этом также свидетельствуют данные Алимова (1981), согласно которым для двустворчатых моллюсков соотношение максимального и предельного размера приблизительно постоянно. Соответственно из уравнения (1) следует, что постоянным будет и произведение τ = kTmax. Это произведение можно назвать видовой продолжительностью жизни.
Цель данной работы – определение коэффициентов уравнения роста Берталанфи (1) и видовой продолжительности жизни для индивидуального линейного роста пресноводного двустворчатого моллюска A. beringiana.
МАТЕРИАЛЫ И МЕТОДЫ
По поводу видового и родового наименования исследуемых моллюсков существуют противоречивые сведения. Так Граф и Куммингс (Graf, Cummings, 2018) относят их к роду Sinanodonta, Старобогатов и соавторы (2004) – к роду Beringiana. В базе данных ITIS валидным родом считается Anodonta (ITIS…, 2018). В результате проведенной ревизии все виды беззубок, описанные для северо-востока России и северо-запада Северной Америки, объединены в один вид Anodonta beringiana Middendorff, 1851 (Особо…, 2018а; ITIS…, 2018). Это наименование мы будем использовать в данной работе. A. beringiana включена в Красную Книгу Магаданской обл. (Особо…, 2018б), а синонимичные виды – в Красные Книги Чукотского автономного округа (Особо…, 2018в) и Сахалинской обл. (Особо…, 2018г).
Раковины погибших моллюсков A. beringiana были собраны на северо-востоке Якутии в нижнем течении р. Колыма на расстоянии 30 км от поселка Черский (68°30.73′ с.ш. 161°30.21′ в.д.). Характерной особенностью этой местности является большое количество пойменных озер. В период весеннего половодья они объединяются с р. Колымой и частью ее притоков в единое водное пространство. Затем при понижении уровня воды в пойме формируются озера, в различной степени изолированные от русла. Часто случается, что изоляция озер нарушается из-за прорывов барьеров, и вода из озер сливается в реку. В одном из таких высохших в 2018 году озер были найдены раковины беззубок в большом числе. Только 26 из них оказались достаточно сохранившимися, чтобы можно было выполнить запланированное исследование.
Здесь уместно отметить, что двустворчатые моллюски из р. Колымы, по-видимому, еще не попали в научные коллекции и не исследовались. По крайней мере, нам не удалось найти упоминания о них в научной литературе. Возможно, они демонстрируют северный предел в распространении наяд в северо-восточной России. Характер их распространения по бассейну Колымы остается неисследованным, но удалось выяснить, что практически до самого устья они встречаются местами – небольшое число поврежденных раковин было замечено в сетях местных рыболовов в поселке Черский.
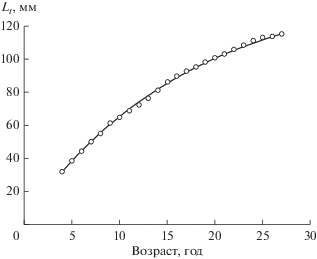
Рис. 1.
Зависимость длины годового кольца (Lt) от его возраста на примере моллюска № 8 (табл. 1). Кружки – измерения. Линия – аппроксимация уравнением роста Берталанфи (1).
Раковины фотографировали вместе с масштабной линейкой. Изображения внедряли в программу Excel. По изображению каждого моллюска измеряли длину раковины и каждого неповрежденного годового кольца с точностью 0.1 мм.
Данные аппроксимировали рекуррентной формой уравнения (1):
где Lt – длина годового кольца возраста t; Lt + 1 – длина годового кольца возраста t + 1; c = exp(–k) – коэффициент, определяющий скорость замедления роста; d = L∞/(1 – c). Аппроксимацию проводили с помощью программы Excel.Сравнение коэффициентов уравнения (2) у разных моллюсков проводили методом регрессионного анализа. Пригодность использования этого уравнения оценивали с помощью критерия нелинейности (Зотин, 2000).
Возраст моллюска вычисляли, суммируя возраст первого измеренного годового кольца (T1) и число распознаваемых годовых колец на поверхности раковины. Возраст первого измеренного годового кольца рассчитывали по уравнению (1):
Сравнение коэффициентов уравнения (2) проводили с помощью методов регрессионного анализа (Хальд, 1956).
РЕЗУЛЬТАТЫ
Линейный рост отдельных особей A. beringiana хорошо описывается уравнением (1). Пример аппроксимации полученных данных этим уравнением показан на рис. 1. Значения коэффициентов уравнений (1) и (2) и возраст исследованных моллюсков приведены в табл. 1.
Таблица 1.
Коэффициенты уравнений (1) и (2) для роста общей массы в индивидуальном постличиночном онтогенезе A. beringiana
| № моллюска | n | Уравнение (2) | Уравнение (1) | T, год | τ | ||
|---|---|---|---|---|---|---|---|
| c | d, мм | k, год–1 | L∞, мм | ||||
| 1 | 14 | 0.932 ± 0.014 | 11.0 ± 1.0 | 0.071 ± 0.015 | 161.0 ± 14.1 | 17 | 1.2 |
| 2 | 19 | 0.947 ± 0.010 | 7.4 ± 0.7 | 0.054 ± 0.011 | 139.8 ± 10.7 | 25 | 1.3 |
| 3 | 15 | 0.934 ± 0.011 | 8.7 ± 0.7 | 0.068 ± 0.012 | 132.0 ± 8.5 | 20 | 1.4 |
| 4 | 14 | 0.940 ± 0.014 | 10.1 ± 1.1 | 0.062 ± 0.015 | 168.4 ± 17.1 | 18 | 1.1 |
| 5 | 18 | 0.946 ± 0.010 | 8.4 ± 0.8 | 0.055 ± 0.011 | 155.5 ± 12.1 | 24 | 1.3 |
| 6 | 15 | 0.941 ± 0.013 | 7.5 ± 0.8 | 0.061 ± 0.014 | 127.9 ± 12.0 | 19 | 1.2 |
| 7 | 12 | 0.942 ± 0.008 | 10.2 ± 0.6 | 0.060 ± 0.008 | 175.4 ± 12.1 | 16 | 1.0 |
| 8 | 23 | 0.943 ± 0.006 | 8.2 ± 0.5 | 0.058 ± 0.006 | 145.1 ± 5.5 | 28 | 1.6 |
| 9 | 14 | 0.946 ± 0.007 | 10.3 ± 0.5 | 0.055 ± 0.007 | 192.6 ± 12.2 | 18 | 1.0 |
| 10 | 11 | 0.936 ± 0.012 | 11.4 ± 0.9 | 0.067 ± 0.013 | 176.4 ± 16.5 | 15 | 1.0 |
| 11 | 12 | 0.946 ± 0.014 | 9.6 ± 1.0 | 0.055 ± 0.014 | 178.5 ± 20.7 | 17 | 1.0 |
| 12 | 21 | 0.946 ± 0.006 | 8.4 ± 0.4 | 0.056 ± 0.006 | 154.3 ± 6.8 | 26 | 1.5 |
| 13 | 12 | 0.945 ± 0.012 | 10.3 ± 0.9 | 0.057 ± 0.013 | 186.8 ± 20.4 | 16 | 0.9 |
| 14 | 16 | 0.929 ± 0.006 | 11.0 ± 0.5 | 0.074 ± 0.006 | 155.5 ± 5.5 | 20 | 1.4 |
| 15 | 14 | 0.939 ± 0.013 | 10.4 ± 1.1 | 0.063 ± 0.014 | 169.9 ± 16.6 | 18 | 1.2 |
| 16 | 14 | 0.948 ± 0.014 | 9.2 ± 1.0 | 0.053 ± 0.015 | 178.3 ± 22.1 | 18 | 1.0 |
| 17 | 16 | 0.943 ± 0.008 | 9.1 ± 0.6 | 0.058 ± 0.009 | 160.2 ± 11.6 | 19 | 1.1 |
| 18 | 12 | 0.945 ± 0.015 | 8.7 ± 1.0 | 0.057 ± 0.016 | 156.8 ± 18.8 | 18 | 1.0 |
| 19 | 16 | 0.927 ± 0.010 | 10.8 ± 0.8 | 0.076 ± 0.011 | 148.2 ± 8.4 | 19 | 1.5 |
| 20 | 15 | 0.934 ± 0.012 | 10.0 ± 0.9 | 0.068 ± 0.013 | 152.2 ± 12.3 | 19 | 1.3 |
| 21 | 14 | 0.936 ± 0.009 | 9.6 ± 0.6 | 0.066 ± 0.010 | 150.9 ± 9.9 | 18 | 1.2 |
| 22 | 12 | 0.932 ± 0.016 | 10.4 ± 1.1 | 0.071 ± 0.017 | 152.2 ± 15.8 | 16 | 1.1 |
| 23 | 19 | 0.948 ± 0.011 | 7.5 ± 0.8 | 0.053 ± 0.011 | 144.3 ± 12.1 | 25 | 1.3 |
| 24 | 9 | 0.933 ± 0.015 | 8.0 ± 0.9 | 0.069 ± 0.016 | 119.8 ± 10.8 | 16 | 1.1 |
| 25 | 11 | 0.940 ± 0.014 | 9.0 ± 0.9 | 0.061 ± 0.015 | 151.1 ± 16.8 | 15 | 0.9 |
| 26 | 8 | 0.936 ± 0.019 | 8.2 ± 1.1 | 0.066 ± 0.021 | 127.9 ± 16.6 | 14 | 0.9 |
| Средние | 377 | 0.940 ± 0.010 | 9.3 ± 0.2 | 0.062 ± 0.011 | 155.8 ± 19.5 | – | – |
Коэффициенты c и d уравнения (2) у разных особей достоверно не отличаются друг от друга. В целом для всей исследованной популяции c = = 0.940 ± 0.010 год–1; d = 9.3 ± 0.2 мм. Соответственно коэффициенты уравнения (1) равны k = 0.062 ± 0.011 год–1; L∞ = 155.8 ± 19.5 мм. Эти значения могут быть использованы для характеристики при сравнительных межпопуляционных и межвидовых исследованиях.
Максимальный возраст (Tmax) для исследованных раковин составил 28 лет. Если принять это значение в качестве максимального возраста для всей популяции, то видовая продолжительность жизни A. beringiana составит τ = kTmax ≈ 1.7.
ОБСУЖДЕНИЕ
Для характеристики роста двустворчатых моллюсков обычно используют так называемую константу роста k из уравнения Берталанфи (Алимов, 1981; Bauer, 1992; Зюганов и др., 1993). Мы предпочитаем использовать вместо нее другую константу, названную нами коэффициент замедления роста (c), связанную с константой роста зависимостью c = exp(–k). Заметим, что если k близка к 0, то k ≈ 1 – c. Коэффициент c выгодно отличается от константы роста тем, что его статистическое распределение близко к нормальному, он входит в рекуррентную форму (2) в качестве коэффициента регрессии и, следовательно, может быть использован для сравнительных целей с помощью стандартных методов регрессионного анализа.
Для изученных моллюсков среднее c = 0.940 и, следовательно, k = 0.062 год–1. Константа роста k, согласно данным литературы, варьирует для разных видов подсемейства Anodontinae в пределах 0.127–0.315 год–1 (Алимов, 1981). Таким образом, значение константы роста A. beringiana ниже, чем у других видов беззубок и наиболее близко оно к значению k у Margaritifera (табл. 2). Известно, что для Margaritifera margaritifera константа роста находится в обратной зависимости от широты, а, следовательно, и от температуры места обитания моллюсков (Зотин, Иешко, 2017). Поэтому можно предположить, что пониженное значение k у A. beringiana связано с тем, что исследованная популяция обитает в относительно холодном водоеме.
Таблица 2.
Видовая продолжительность жизни у двустворчатых моллюсков
| Семейство | Вид | k, год–1 | Tmax, год | τ = kT | Ссылки |
|---|---|---|---|---|---|
| Mytilidae | Mytilus edulis | 0.165 | 13 | 2.1 | 1 |
| Unionidae (Anodontinae) |
Anodonta anatina | 0.185 | 12 | 2.2 | 2, 3 |
| A. seisanensis | 0.315 | 6.8 | 2.1 | 2 | |
| A. piscinalis | 0.350 | 5.8 | 2.0 | 2 | |
| Cristaria plicata | 0.127 | 22 | 2.8 | 2 | |
| Unionidae | Lanceolaria cylindrica | 0.230 | 8.6 | 2.0 | 2 |
| Unio pictorum | 0.336 | 6.7 | 2.3 | 2, 3 | |
| U. tumidus | 0.343 | 6.5 | 2.2 | 2, 3 | |
| U. crassus | 0.187 | 12 | 2.2 | 2 | |
| Margaritiferidae | Margaritifera dahurica | 0.122 | 14 | 1.7 | 2 |
| M. laevis | 0.063 | 45 | 2.8 | 2 | |
| M. margaritifera | 0.021 | 105 | 2.2 | 2,3 | |
| Dreissenidae | Dreissena polymorpha | 0.492 | 4.2 | 1.7 | 2 |
| Cyrenidae | Сorbicula purpurea | 0.336 | 5.7 | 1.9 | 2 |
| С. tibetensis | 0.580 | 4.1 | 2.4 | 2 | |
| С. fluminalis | 0.320 | 6.4 | 2.0 | 2 | |
| Astartidae | Astarte borealis | 0.411 | 7 | 2.9 | 4 |
| A. arctica | 0.485 | 6 | 2.9 | 4 | |
| Solecurtidae | Tagelus plebeius | 0.173 | 14 | 2.4 | 5 |
| Vesicomyidae | Calyptogena kilmeri | 0.25 | 7 | 1.8 | 6 |
Примечаниe. k – константа роста из уравнения (1); Tmax – максимальная продолжительность жизни; τ – видовая продолжительность жизни. Ссылки: 1 – Зотин, Озернюк, 2004; 2 – Алимов, 1981; 3 – Зотин, 2009; 4 – Селин, 2007; 5 – Lomovasky, 2003; 6 – Barry et al., 2007.
Для животных, рост которых может быть описан уравнением Берталанфи, показана отрицательная корреляция между константой роста и максимальной продолжительностью жизни (Алимов, 1981; Bauer, 1992). То есть, для этих видов τ = = kTmax может служить показателем видовой продолжительности жизни. В табл. 2 приведены значения τ для двустворчатых моллюсков. Из таблицы видно, что видовая продолжительность жизни для разных видов моллюсков варьирует незначительно от 1.7 до 2.9 (в среднем 2.2 ± 0.1). Полученное для A. beringiana значение τ соответствует нижней границе диапазона. Однако если учесть небольшую выборку исследованных моллюсков, можно предположить, что максимальная продолжительность жизни на самом деле больше 28 лет, и соответственно больше видовая продолжительность жизни. Если считать, что τ для A. beringiana соответствует среднему для двустворчатых моллюсков значению, то в этом случае оценка максимальной продолжительности жизни моллюсков в исследованной популяции составит 35 лет.
Эта величина представляет интерес в плане обсуждения долгожительства моллюсков, обитающих в северных реках. В этой связи часто привлекают внимание пресноводные жемчужницы, потому что они могут жить очень долго – до 180 лет (Helama, Valovirta, 2008). При этом самые большие рекорды отмечаются на севере ареала (Bolotov et al., 2018). Жемчужницы распространены в менее холодном климате, чем обнаруженные нами моллюски. Однако возраст последних сравнительно небольшой. Это во многом объясняется различиями среды обитания: Anodonta обитают в более продуктивных и теплых водоемах. В нашем случае в озере, которое обогащается и прогревается за счет полноводной реки, текущей с юга (в замкнутых холодных озерах окружающей тундры и лесотундры эти моллюски не обнаруживаются). Однако, несмотря на это обстоятельство, климат в нижнем течении Колымы, по-видимому, близок к экстремальному для крупных двустворчатых моллюсков. Это означает, что видовые особенности в большей степени определяют пределы продолжительности жизни, чем условия существования.
Работа осуществлена в рамках государственной программы фундаментальных научных исследований ИБР РАН № 0108-2019-0003. Материал был собран в ходе работы по гранту Санкт-Петербургского государственного университета “Урбанизированные экосистемы Арктического пояса Российской Федерации: динамика, состояние и устойчивое развитие” № 39377661.
Список литературы
Алимов А.Ф. Функциональная экология пресноводных двустворчатых моллюсков. Л.: Наука, 1981. 248 с.
Зотин А.А. Статистическая оценка параметров аллометрических уравнений // Изв. РАН. Сер. биол. 2000. № 5. С. 517–524.
Зотин А.А. Закономерности роста и энергетического обмена в онтогенезе моллюсков: Дис. … докт. биол. наук. М.: ИБР РАН, 2009. 334 с.
Зотин А.А., Иешко Е.П. Сравнительный анализ роста Margaritifera margaritifera (Bivalvia) из разных популяций Карелии и Кольского полуострова // Изв. РАН. Сер. биол. 2017. № 1. С. 5–9.
Зотин А.А., Озернюк Н.Д. Особенности роста мидии обыкновенной Mytilus edulis Белого моря // Изв. РАН. Сер. биол. 2004. № 4. С. 459–464.
Зюганов В.В., Зотин А.А., Третьяков В.А. Жемчужницы и их связь с лососевыми рыбами. М.: ЦНИИТЭИлегпром, 1993. 134 с.
Особо охраняемые природные территории (ООПТ). Anodonta beringiana Middendorff, 1851. 2018а. http://oopt.aari.ru/bio/50104.
Особо охраняемые природные территории (ООПТ). Перечень объектов животного мира, занесенных в Красную книгу Магаданской области. 2018б. http://oopt.aari.ru/rbdata/80/anim.
Особо охраняемые природные территории (ООПТ). Перечень объектов животного мира, занесенных в Красную книгу Чукотского автономного округа. 2018в. http://oopt.aari.ru/rbdata/83/anim.
Особо охраняемые природные территории (ООПТ). Перечень объектов животного мира, занесенных в Красную книгу Сахалинской области. 2018г. http://oopt.aari.ru/rbdata/81/anim.
Селин Н.И. Форма раковины, рост и продолжительность жизни Astarte arctica и A. borealis (Mollusca: Bivalvia) из сублиторали северо-восточной части острова Сахалин // Биология моря. 2007. Т. 33. № 4. С. 278–283.
Старобогатов Я.И., Прозорова Л.А., Богатов В.В., Саенко Е.М. Моллюски // Определитель пресноводных беспозвоночных России и сопредельных территорий Т. 6. Моллюски, Полихеты, Немертины. СПб.: Наука, 2004. С. 9–252.
Хальд А. Математическая статистика с техническими приложениями. М.: Изд-во иностр. лит., 1956. 664 с.
Barry J.P., Whaling P.J., Kochevar R.K. Growth, production, and mortality of the chemosynthetic vesicomyid bivalve, Calyptogena kilmeri from cold seeps off central California // Marine Ecol. 2007. V. 28. P. 169–182.
Bauer G. Variation in life span and size of the freshwater pearl mussel // J. Anim. Ecol. 1992. V. 61. P. 425–436.
Bertalanffy L. von. Principles and theory of growth // Fundamental Aspects of Normal and Malignant Growth. Amsterdam: Elsevier, 1960. P. 137–259.
Bolotov I.N., Makhrov A.A., Gofarov M.Yu., Aksenova O.V., Aspholm P.E., Bespalaya Yu.V., Kabakov M.B., Kolosova Yu.S., Kondakov A.V., Ofenböck Th., Ostrovsky A.N., Popov I.Yu., von Proschwitz T., Rudzīte M., Rudzītis M., Sokolova S.E., Valovirta I., Vikhrev I.V., Vinarski M.V., Zotin A.A. Climate change reduces suitable areas for freshwater pearl mussels throughout Europe: an indirect modeling approach // Sci. Rep. UK 2018. www.nature.com/scientificreports.
Graf D., Cummings K. The MUSSEL Project: MUSSELp. 2018. http://mussel-project.uwsp.edu/.
Helama S., Valovirta I. The oldest recorded animal in Finland: ontogenetic age and growth in Margaritifera margaritifera (L. 1758) based on internal shell increments // Memoranda Soc. Fauna Flora Fennica. 2008. V. 84. P. 20–30.
ITIS, the Integrated Taxonomic Information System. Anodonta beringiana Middendorff, 1851. 2018. https:// www.itis.gov/servlet/SingleRpt/SingleRpt?search_to-pic=TSN&search_value=79931#null.
Lomovasky B.J. The estuarine bivalve Tagelus plebeius: Recent ecology and past climate // SEPG. 2003. № 2218. P. 1–6.
Дополнительные материалы отсутствуют.