

Онтогенез, 2020, T. 51, № 2, стр. 132-142
Онтогенез лишайника Lobaria pulmonaria (L.) Hoffm. в растительных сообществах бореальной зоны
Р. В. Игнатенко a, b, *, В. Н. Тарасова a, Е. Ф. Марковская a
a Петрозаводский государственный университет
185910 Петрозаводск, пр. Ленина, 33, Россия
b Федеральный исследовательский центр “Карельский научный центр Российской академии наук”
185000 Петрозаводск, ул. Пушкинская, 11, Россия
* E-mail: ocean-9@mail.ru
Поступила в редакцию 08.04.2019
После доработки 29.10.2019
Принята к публикации 07.11.2019
Аннотация
Представлен онтогенез эпифитного лишайника Lobaria pulmonaria. Исследования проведены в лесных фитоценозах Карелии с давностью нарушения от 80 до 450 лет. На основе концепции дискретного описания растений и лишайников выделено 4 периода и 14 возрастных состояний в онтогенезе L. pulmonaria. Каждое онтогенетическое состояние L. pulmonaria характеризуется определенными размерами таллома, наличием и степенью развития вегетативных диаспор и плодовых тел на поверхности слоевища. Описаны возможные варианты онтогенеза вида.
ВВЕДЕНИЕ
В современной лихенологии одним из важных направлений исследований является изучение онтогенетической структуры видов (Михайлова, Воробейчик, 1999; Суетина, 2001; Михайлова, 2005; Суетина, Глотов 2010, 2015; Игнатенко, Тарасова, 2017). Выделение разнокачественных возрастных талломов способствует лучшему пониманию закономерностей онтогенетического развития и формирования популяционной структуры видов лишайников в конкретный момент времени, а также позволяет прогнозировать их развитие в будущем (Михайлова, 2005; Суетина, Глотов 2010). Данный подход особенно важен для формирования стратегии охраны при изучении редких и находящихся под угрозой исчезновения видов лишайников, нуждающихся в охране (Gauslaa, 1997; Scheidegger, 1998; Tarasova et al., 2008; Суетина, Глотов, 2010; Микрюков, 2011; Фенько, Тарасова, 2016; Игнатенко, 2018; Игнатенко, Тарасова, 2018).
Крупный эпифитный цианобионтный лишайник лобария легочная (Lobaria pulmonaria (L.) Hoffm., Lobariaceae, Ascomycotina), широко распространенный в бореальных, умеренных, горных и океанических районах мира (Yoshimura, 1971, 1998), занесен в Красную книгу Российской Федерации (2008) со статусом уязвимого вида с сокращающейся численностью (2б). L. pulmonaria – это трехкомпонентный лишайник, образованный микобионтом (аскомицетным грибом), первичным фотобионтом – эукариотической зеленой водорослью Dictyochloropsis reticulata Tschermak-Woess (Tschermak-Woess, 1995), и вторичным фотобионтом – азотфиксирующей цианобарктерией Nostoc sp. (Tschermak-Woess, 1988).
Считается, что лишайник L. pulmonaria крайне чувствителен к любому типу антропогенного воздействия и в умеренном климате приурочен к старовозрастным лесам (Andersson, Appelqvist, 1987; Rose, 1992; Gauslaa, 1994; Andersson et al., 2003; Liira, Sepp, 2009). Кроме того, имеются сведения, что этот крупный, хорошо известный в литературе лишайник, ассоциирован с другими редкими и исчезающими видами организмов, и его можно рассматривать в качестве показателя (umbrella species) их присутствия в сообществе (Campbell, Fredeen, 2004; Pykälä, 2004; Radies, Coxson, 2004; Ravera et al., 2006; Scheidegger, Werth, 2009; Nascimbene et al., 2010).
L. pulmonaria является одним из самых изученных видов лишайников: в мировой базе данных лихенологической литературы “Recent Literature on Lichens” (Culberson et al., 2019) ключевые слова с названием данного вида содержатся в 313 статьях. Несмотря на это, в отношении исследования его онтогенеза имеется еще достаточно много вопросов. Так, И.Н. Михайлова (2005) выделила онтогенетически разнокачественные талломы и разделила их на разные функционально-возрастные группы – от стерильных до фертильных (см. раздел Методы). В эту работу не были включены талломы длиной <0.2 см; вызывает сомнение положение фертильной группы в конце функционально-возрастного спектра (после сенильной группы) и не логично включение в одну стерильную группу талломов разных онтогенетических состояний.
Другие авторы – В.В. Горшков и Н.А. Семенова (2008) – на основе концепции дискретного описания лишайников (Суетина, 2001), которая в большей степени соответствует классической схеме онтогенеза растений (Работнов, 1950; Уранов, 1975), выстроили логическую последовательность онтогенетических возрастных состояний вида, но не рассматривали талломы размером <0.5 см и не дали подробного описания разных возрастных состояний.
Проведенный анализ показал, что в литературе в целом отсутствует полное описание онтогенетического развития вида L. pulmonaria, не известны морфометрические показатели талломов на ранних этапах онтогенеза. Очевидно, что имеющиеся в литературе данные требуют обобщения, унификации, проведения соответствий между выделяемыми разными авторами групп и состояний, изучения их размерной структуры, морфогенеза и экологических особенностей.
МАТЕРИАЛЫ И МЕТОДЫ
Исследование проводилось в 2014–2016 гг. в лесных сообществах Республики Карелия: в подзоне средней тайги – в заповеднике “Кивач”, национальном парке “Водлозерский”, заказнике “Кижский”, Петрозаводском городском округе; в подзоне северной тайги – в заповеднике “Костомукшский”, национальном парке “Паанаярви”.
Сбор данных осуществлялся на сети постоянных пробных площадей (ПП) размером 1 га. На каждой ПП были выполнены полные геоботанические описания фитоценозов (Андреева и др., 2002), в ходе которых оценивали сомкнутость крон деревьев (сквозистомером Ипатова) и относительную сумму площадей поперечных сечений стволов живых деревьев, сухостоя и валежа (полнотомером Биттерлиха), производили описания напочвенного покрова. Для оценки давности нарушения в сообществах использовался метод изучения возрастной структуры ценопопуляций ели (Picea spp.) (Ставрова и др., 2016).
Всего было описано 33 ПП, расположенных в различных типах леса с давностью нарушения от 80 до 450 лет. В подзоне средней тайги все изученные фитоценозы принадлежат к единому эколого-динамическому ряду, представляющему собой восстановительную динамику ельника черничного зеленомошного: 1) средневозрастные осинники чернично-злаковые (80–110 лет), 2) смешанные елово-осиновые сообщества злаково-черничные (150–190 лет); 3) ельники черничные зеленомошные субклимаксовые (210–260 лет) и 4) условно климаксовые (410–450 лет). Средневозрастные осинники в подзоне северной тайги в результате рекогносцировочных маршрутов (~110 км) не были зарегистрированы. На данной территории еловые сообщества, как правило, возобновляются через березовые леса. Поэтому исследования здесь проводились только в двух типах лесных фитоценозов: 1) смешанных елово-березовых сообществах злаково-черничных (180–200 лет) и 2) ельниках черничных зеленомошных субклимаксовых (210–270 лет).
Изучение показателей талломов L. pulmonaria выполнены методом сплошного учeта на всех субстратах (ветвях и стволах живых и мертвых деревьев) на высоте 0–2 м от земли с регистрацией характеристик местообитания (параметров деревьев и микроусловий). Для каждого таллома при помощи рамки 25 × 25 см отмечали общую площадь и площадь некрозов (см2), а также принадлежность к функционально-возрастной группе по методике И.Н. Михайловой (2005): стерильные (st), без репродуктивных структур; гипосоредиозные (s1), с маргинальными соралями; мезосоредиозные (s2), с обильными маргинальными и единичными ламинальными соралями; гиперсоредиозные (s3), с обильными ламинальными соралями; фертильные (fert), с апотециями; субсенильные (s/sen), частично разрушенные в центре с регенеративными структурами на старых лопастях; сенильные (sen) – отмирающие лопасти, с регенеративными структурами, образовавшиеся в результате распада крупного таллома.
Для изучения талломов площадью <1 см2 со стволов не менее 5 деревьев с каждой ПП отбирались образцы корки ствола дерева в местах произрастания крупных талломов (на расстоянии от 2 до 10 см) и с участков, где талломы не визуализировались (как правило, на противоположной стороне ствола, на участках корки без мхов) (на основании Разрешения № 18 на добывание объектов животного и растительного мира, принадлежащих к видам, занесенным в Красную книгу РФ от 28.03.2016 Росприроднадзора). Образцы субстрата изучались в лабораторных условиях при помощи бинокуляра (Микромед МС2), при этом регистрировали следующие признаки талломов вида (при их наличии): длина, ширина (мм), наличие ризин и шероховатостей, наличие и число выемок (шт.), форма (плоская/неплоская, округлая), число лопастей (шт.), цвет, налет, ямчатые углубления.
Начальные этапы онтогенеза L. pulmonaria (латентный период, возрастные состояния прототаллюс и протероталлюс прегенеративного периода) не были исследованы и их описание дано с учетом литературных данных, общих для лишайников в целом (Суетина, 2001, 2006; Семенова, 2005; Суетина, Глотов, 2015). Ювенильное и имматурные возрастные состояния описаны на основе результатов исследования талломов площадью <1 см2. Описания последующих онтогенетических состояний – от виргинильного до сенильного, частично или полностью, совпадают с имеющимися сведениями в литературе (Михайлова, 2005; Горшков, Семенова, 2008). Так, функционально-возрастным группам (Михайлова, 2005) соответствуют (полностью или частично) следующие онтогенетические состояния: стерильные – виргинильное 1, гипосоредиозные – виргинильное 2а, мезосоредиозные – виргинильное 2b, гиперсоредиозные – виргинильное 2с, фертильные – генеративные; субсенильные – субсенильные, сенильные – сенильные.
Размерная структура онтогенетических состояний ювенильное – сенильное изучалась на основе линейных размеров (длина, ширина) и площади талломов.
Статистическая обработка результатов выполнена на основе регрессионного анализа с использованием уравнения линейной функции: y = aХ + b (Андреева и др., 2002; Ивантер, Коросов, 2003). При наличии взаимосвязей линии на графиках проводили в соответствии с уравнением регрессии. При отсутствии достоверных изменений на графике изображали горизонтальную прямую на уровне среднего значения. Средние значения характеристик талломов L. pulmonaria сравнивали друг с другом, используя непараметрический критерий Манна–Уитни (Ивантер, Коросов, 2003).
Анализ выполнен на основе описания 4528 талломов, произрастающих на 676 субстратных единицах (отдельно стоящих или лежащих деревьях и кустарниках) (табл. 1).
Таблица 1.
Объем анализируемого материала
| Тип данных | Количество |
|---|---|
| Число пробных площадей, шт. | 33 |
| Число учетных деревьев и кустарников, шт. | 676 |
| Число описаний талломов площадью >1 см2L. pulmonaria, шт. | 3709 |
| Число описаний талломов площадью <1 см2L. pulmonaria, шт. | 819 |
| Число и площадь образцов корки, отобранных с субстратов для изучения микроталломов, шт./см2 | 632/11 967 |
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Полный жизненный цикл L. pulmonaria, включающий развитие от споры до споры, начинается с прорастания аскоспоры гриба, включает процессы лихенизации с эукариотической зеленой водорослью (Dictyochloropsis reticulata), долихенизации – с цианобактерией (Nostoc sp.), и заканчивается образованием плодовых тел, процессами старения и отмирания таллома.
Как известно, полового размножения у водорослей и цианобактерий в теле лишайника не наблюдается – оно характерно только для грибного компонента. Споры полового размножения у вида L. pulmonaria формируются в плодовых телах открытого типа (апотециях). Апотеции красно-коричневого цвета образуются на верхней поверхности вдоль ребер и краям таллома, имеют размер 2–4 мм в диаметре (Denison, 2003). В каждом апотеции содержатся сумки со спорами, количество которых превышает десятки тысяч штук. По мере созревания, споры высыпаются из сумок и, подхватываемые воздушными и водными потоками, являются легкими агентами для распространения и колонизации новых субстратов. Однако на данном этапе популяции вида проходят своеобразное “узкое горлышко” (bottle neck), связанное с необходимостью прорастающих из споры гиф гриба встретить в данном местообитании нужную водоросль для последующего процесса лихенизации (Walser et al., 2001). Возрастные состояния прототаллюс (прорастание гифы из споры) и протероталлюс (лихенизация) в данном исследовании нами не наблюдались, в виду микроскопических размеров, кратковременности протекания и технических трудностей в определении видовой принадлежности. Однако, имеющиеся в литературе данные (Суетина, 2001, 2006; Семенова, 2005; Суетина, Глотов, 2015), и общая логика хода онтогенеза предполагают их наличие, по крайней мере, в тех ценопопуляциях вида, в которых присутствуют генеративные талломы.
Полученные в настоящей работе данные позволяют внести изменения в имеющуюся в литературе схему онтогенеза L. pulmonaria (Горшков, Семенова, 2008) (табл. 2). Так, в прегенеративном периоде можно выделить 4 возрастных состояния: ювенильное, имматурное 1, имматурное 2, имматурное 3.
Таблица 2.
Онтогенетические состояния Lobaria pulmonaria
| Период | Онтогенетические состояния и их индексы | Признаки |
|---|---|---|
| Латентный | Спора гриба (sp) | Спора четырехклеточная, веретеновидная, бесцветная |
| Прегенеративный | Прототаллюс (pr) | Образование из спор гиф мицелия |
| Протероталлюс (prt) | Обвивание гифами клеток зеленой водоросли и образование зачатка слоевища | |
| Ювенильный таллом (j) | Округлый, выпуклый, каплевидной формы (накипное слоевище) | |
| Имматурное 1 (im1) | Листоватое слоевище, в форме плоской пластинки овальной формы | |
| Имматурное 2 (im2) | Слоевище продолговатой формы, с 0–1 выемкой | |
| Имматурное 3 (im3) | Сильнорассеченное слоевище (2 и > выемки), появляются ямчатые углубления (характерная “легочная” складчатость), и/или первые зачаточные лопасти, формирование цефалодий | |
| Виргинильное 1 (v1) | Таллом имеет характерную взрослую форму, хорошо сформированные лопасти с ямчатыми углублениями и ребрами; стерильный, без репродуктивных структур | |
| Виргинильное 2a (v2a) | Слоевище с маргинальными соралями, цефалодии становятся заметными на верхней и/или нижней поверхности слоевища | |
| Виргинильное 2b (v2b) | Слоевище с обильными маргинальными и единичными ламинальными соралями | |
| Виргинильное 2c (v2c) | Слоевище с обильными ламинальными соралями, с хорошо заметными цефалодиями, формируются изидии | |
| Генеративный | Генеративное (g) | Крупное слоевище с апотециями, с обильными ламинальными соралями, на старых участках таллома имеются изидии |
| Постгенеративный | Субсенильное (ss) | Частично разрушенное слоевище, на старых участках имеются изидии и лобули (регенеративные структуры, не способные к распространению) |
| Сенильное (s) | Распад крупного таллома, наличие большого количества некротических образований, отмирающие лопасти с регенеративными структурами, не способными к распространению |
Ювенильное и имматурные состояния. В ходе развития на начальных стадиях онтогенеза талломы L. pulmonaria площадью <1 см2 претерпевают ряд последовательных (качественно различных) морфологических состояний (в скобках указано число исследованных талломов): 1) накипное слоевище округлой, выпуклой, каплевидной формы (209 шт.), 2) листоватое слоевище в форме плоской пластинки овальной формы (соотношение длины к ширине 3 : 2) (182 шт.), 3) листоватое слоевище продолговатой формы (соотношение длины к ширине 7 : 3), без выемок или с одной выемкой (214 шт.), 4) сильнорассеченное слоевище с 2-мя и более выемками, с ямчатыми углублениями (“легочной” складчатостью) и/или с первыми зачаточными лопастями, с формирующимися цефалодиями (214 шт.). Выявлено, что в последовательном морфологическом ряду состояний, при переходе от округлой формы к рассеченной, средняя длина и ширина талломов L. рulmonaria увеличивается (рис. 1а, 1б, табл. 3), растет площадь талломов (рис. 2а–2г, табл. 4). Важно отметить, что округлые талломы не превышают 0.15 см в длину и 0.12 см в ширину, овальные – ≤0.45 и ≤0.24 см, продолговатые – ≤0.82 и ≤0.71 см и рассеченные – ≤1.04 и ≤0.86 см соответственно.
Рис. 1.
Средние значения длины (а) и ширины (б) талломов Lobaria pulmonaria разных морфологических форм в исследованных фитоценозах Карелии. Примечание: формы талломов обозначены следующими цифрами: 1 – округлая, 2 – овальная, 3 – продолговатая, 4 – рассеченная. Разными латинскими буквами отмечены статистически значимые различия между средними значениями при p < 0.01.
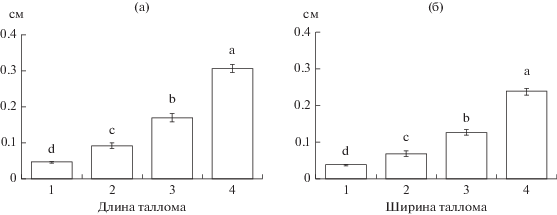
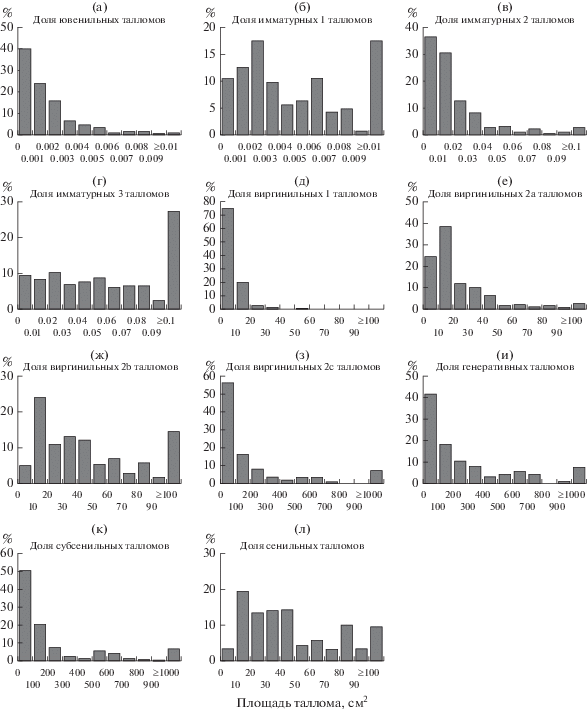
Таблица 3.
Линейные размеры талломов Lobaria pulmonaria, относящихся к разным онтогенетическим состояниям
| Онтоге- нетические состояния |
Число талломов | Длина таллома | Ширина таллома | ||||
|---|---|---|---|---|---|---|---|
| минимальные/ максимальные значения, см | среднее значение, см | CV1, % | минимальные/ максимальные значения, см | среднее значение, см | CV1, % | ||
| j | 209 | 0.01–0.15 | 0.05 ± 0.002 | 53 | 0.01–0.12 | 0.04 ± 0.001 | 52 |
| im1 | 182 | 0.01–0.45 | 0.09 ± 0.007 | 61 | 0.01–0.24 | 0.06 ± 0.005 | 49 |
| im2 | 214 | 0.03–0.82 | 0.17 ± 0.008 | 61 | 0.02–0.71 | 0.12 ± 0.007 | 65 |
| im3 | 214 | 0.04–1.04 | 0.32 ± 0.01 | 56 | 0.02–0.86 | 0.23 ± 0.01 | 57 |
| v1 | 1136 | 1–30 | 3.72 ± 0.11 | 78 | 1–13 | 2.54 ± 0.06 | 59 |
| v2a | 1117 | 2–40 | 8.01 ± 0.23 | 82 | 1–25 | 5.43 ± 0.11 | 57 |
| v2b | 279 | 2–39 | 12.15 ± 0.44 | 52 | 2–29 | 9.17 ± 0.36 | 56 |
| v2c | 192 | 2–76 | 20.16 ± 1.13 | 66 | 1–44 | 14.05 ± 0.69 | 59 |
| g | 156 | 5–91 | 29.79 ± 1.78 | 60 | 3–59 | 20.01 ± 1.19 | 59 |
| ss | 326 | 5–84 | 26.2 ± 1.09 | 67 | 1–64 | 16.8 ± 0.65 | 65 |
| s | 503 | 2–63 | 19.39 ± 0.58 | 63 | 1–49 | 13.1 ± 0.39 | 63 |
Установлено, что среди изученных признаков первыми в ходе онтогенеза появляются щетинки (опушение), ризины (на нижней стороне таллома) и шероховатости (на верхней стороне); позднее формируются выемки, и только после этого образуются выраженные ямчатые углубления.
Основываясь на полученных данных и имеющихся в литературе сведениях об онтогенезе растений и лишайников (Работнов, 1950; Уранов, 1975; Суетина, 2001; Горшков, Семенова, 2008; Суетина, Глотов, 2010; Суетина, Ямбердова, 2010 и др.), все изученные микроскопические талломы можно отнести к двум возрастным состояниям прегенеративного периода онтогенеза: округлые выпуклые талломы – к ювенильному (j) состоянию, а остальные 3 морфологические фазы, развивающиеся последовательно друг за другом, – к имматурному (im1, im2, im3 соответственно).
Талломы, находящиеся в ювенильном состоянии, не имеют признаков взрослого организма. Начиная с имматурного 1 до имматурного 3 состояний у L. pulmonaria формируется листоватое слоевище, у талломов увеличиваются линейные размеры и площадь (табл. 3, табл. 4, рис. 2а–2г), начинают появляться и постепенно развиваются характерные видовые особенности. Важно отметить, что в возрастном состоянии имматурное 3 в теле лишайника начинают формироваться цефалодии, что свидетельствует о завершении процесса лихенизации и образовании трехкомпонентной системы: гриба, зеленой водоросли и цианобактерии. Так, в работе C. Cornejo и C. Scheidegger (2013) описан процесс образования цефалодий у молодых талломов L. pulmonaria. Авторы отмечают, что данные структуры у молодых талломов могут быть как внешними, так и внутренними.
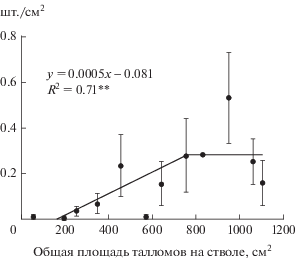
Как показали проведенные исследования, ювенильные и имматурные талломы L. pulmonaria обнаружены только в тех местообитаниях, где растут макроталломы, то есть в непосредственной близости к крупным талломам, которые, вероятно, являются материнскими. С противоположной стороны ствола не было обнаружено ни одного таллома площадью <1 см2. Важно отметить, что в лесных сообществах подзоны средней тайги на стволах Populus trеmulа с увеличением общей площади талломов на дереве с 50 до 750 см2 плотность ювенильных и имматурных талломов повышается в 30 раз: с 0.01 до 0.3 шт./см2 (рис. 3).
Рис. 3.
Плотность микроталломов Lobaria pulmonaria при разной общей площади талломов на стволах Populus trеmulа в лесных сообществах среднетаежной подзоны. Примечание: вертикальными линиями показана ошибка среднего, R2 – коэффициент детерминации; ** – уровень значимости, p < 0.01.
Виргинильные состояния. Как и у растений, формирование типичной жизненной формы лишайников, сохраняющейся на последующих этапах онтогенеза, происходит в виргинильном (v) онтогенетическом состоянии (Суетина, Ямбердова, 2010). Так, в возрастном состоянии виргинильное 1 (v1) прегенеративного периода талломы L. pulmonaria имеют характерный взрослый облик – хорошо сформированные лопасти, ямчатые углубления и ребра, но являются стерильными, без репродуктивных структур. Большинство талломов этого состояния (75%) имеют площадь ≤10 см2 (рис. 2д, табл. 4). При дальнейшем развитии лишайника в состоянии виргинильное 2a (v2a) площадь таллома увеличивается: у 74% она варьирует от 1.3 до 30 см2 и в среднем составляет 14 см2 (рис. 2е). По краям лопастей появляются вегетативные диаспоры, которые занимают до 10% площади таллома, ребра становятся более выпуклыми. На верхней и/или нижней поверхности таллома становятся заметными цефалодии, в виде коричневых или темно-коричневых выпуклостей. В онтогенетическом состоянии виргинильное 2b (v2b) значительная часть талломов (71%) имеет площадь от 10 до 70 см2 (рис. 2ж) со средним значением ~30 см2. Большое количество вегетативных пропагул формируется по краям таллома, появляются единичные ламинальные сорали. Доля площади с вегетативными диа-спорами от общей площади таллома составляет от 10 до 30%. Заканчивается прегенеративный период онтогенетическим состоянием виргинильное 2c (v2c). При этом площадь таллома в данном состоянии сильно варьирует от 6.25 до 3437 см2 (табл. 4, рис. 2з) и в среднем составляет 265.9 см2. У 78% талломов площадь превышает 40 см2. Сорали обильно распространены по краям и ребрам таллома, площадь их поверхности составляет >30%, на верхней и/или нижней поверхности таллома хорошо просматриваются цефалодии, а на старых участках таллома начинают формироваться изидии.
Генеративное состояние. L. pulmonaria размножается и распространяется преимущественно с помощью вегетативных диаспор и плодовые тела появляются редко (Gauslaa, 2006; Edman et al., 2008; Jüriado et al., 2011). Считается, что апотеции формируются только в генетически гетерогенных популяциях (Zoller et al., 1999). В изученных лесных сообществах Карелии только у 4% талломов обнаружены плодовые тела. Генеративные (g) талломы имеют достаточно крупные размеры. Так, у ~60% талломов площадь составляет ≥100 см2 (рис. 2и). Важно отметить, что апотеции формируются на богатых соралями талломах, с ярко выраженными ребрами. На старых участках таллома имеются изидии.
Субсенильное и сенильное состояния. В постгенеративном периоде у субсенильных (ss) талломов L. pulmonaria начинает разрушаться слоевище, на старых, отмирающих участках имеются изидии, а также начинают формироваться регенеративные структуры, не способные к распространению (лобули). Однако площадь у 50% талломов, принадлежащих к субсенильному состоянию, составляет ≥100 см2 (рис. 2к). В данном онтогенетическом состоянии площадь поверхности таллома с вегетативными пропагулами составляет >30%. Завершает онтогенез сенильное (s) состояние, при котором большая часть таллома разрушается, а на оставшихся лопастях имеются крупные некротические участки, а также регенеративные структуры, не способные к распространению. Площадь некрозов от общей площади таллома составляет в среднем ~20%. Большая часть талломов (~60%) этого состояния имеет площадь от 20 до 50 см2 (рис. 2л). При этом на оставшихся частях таллома присутствуют вегетативные пропагулы (сорали, изидии), которые, по-видимому, успешно распространяются до момента отмирания таллома.
Таблица 4.
Площадь таллома и доля некротических образований в талломах Lobaria pulmonaria разных онтогенетических состояний
| Онтоге- нетические состояния |
Число талломов | Площадь таллома | Площадь некрозов | ||||
|---|---|---|---|---|---|---|---|
| минимальные/ максимальные значения, см2 | среднее значение, см2 | CV, % | минимальные/ максимальные значения, см2 | среднее значение, см2 | CV, % | ||
| j | 209 | 0.0001–0.048 | 0.0022 ± 0.0003 | 171 | – | – | – |
| im1 | 182 | 0.0003–0.057 | 0.0065 ± 0.0007 | 121 | – | – | – |
| im2 | 214 | 0.001–0.76 | 0.034 ± 0.0067 | 288 | – | – | – |
| im3 | 214 | 0.001–0.89 | 0.095 ± 0.0075 | 119 | – | – | – |
| v1 | 1136 | 0.3–69 | 7.9 ± 0.26 | 110 | 0–50 | 1.2 ± 0.21 | 594 |
| v2a | 1117 | 1.25–443 | 27.0 ± 1.19 | 147 | 0–67 | 4.5 ± 0.32 | 240 |
| v2b | 279 | 3.1–544 | 58.1 ± 3.97 | 114 | 0–50 | 5.6 ± 0.58 | 174 |
| v2c | 192 | 6.25–3437 | 265.9 ± 34.84 | 182 | 0–50 | 11.4 ± 3.73 | 698 |
| g | 156 | 12.5–3750 | 330.2 ± 41.04 | 155 | 0–86 | 8.1 ± 0.92 | 142 |
| ss | 326 | 6.25–1937 | 292.7 ± 30.65 | 189 | 0–54 | 9.2 ± 0.49 | 96 |
| s | 503 | 1.9–564 | 96.6 ± 5.59 | 128 | 0–80 | 19.7 ± 0.79 | 91 |
ЗАКЛЮЧЕНИЕ
На основе изученной выборки талломов L. pulmonaria, с учетом литературных данных (Михайлова, 2005; Горшков, Семенова, 2008), в онтогенезе вида было выделено 4 периода и 14 возрастных состояний.
После объединения грибного компонента с водорослью зачаток слоевища обретает округлую, каплевидную форму (ювенильное возрастное состояние); со временем таллом становится листоватым, сильнорассеченным, появляется ямчатая бугристость. В состоянии имматурное 3 начинает происходить внедрение цианобактерий в состав таллома. Таким образом, в возрастном состоянии виргинильное 1 талломы приобретают взрослый облик, однако все еще остаются стерильными.
В ходе индивидуального развития, в возрастном состоянии виргинильное 2, таллом увеличивается в размере на его поверхности образуются сорали, развивающиеся сначала по краям лопастей, а впоследствии и на их верхней поверхности, формируются изидии, цефалодии становятся хорошо заметными на верхней и/или нижней поверхности таллома. Вегетативные диаспоры содержат в себе клетки гриба и зеленой водоросли. После отделения от материнского таллома соредии закрепляются на стволе дерева гифами гриба и образуют новый таллом. Однако для того чтобы выжить, ему необходимо включить в состав своего тела цианобактерию (Werth et al., 2006).
Когда таллом достигает крупных размеров, а его поверхность обильно покрывают сорали, он становится потенциально генеративным. В результате полового размножения на поверхности таллома образуются плодовые тела – апотеции.
С течением времени таллом начинает разрушаться. При этом на старых лопастях имеются изидии, которые могут разрастаться в лобули (регенеративные структуры не способные к распространению) (Михайлова, 2005). Заканчивается жизненный цикл лишайника распадом крупного таллома и наличием большого количества некротических образований. Важно отметить, что на поверхности старых субсенильных и сенильных талломов присутствуют в большом количестве вегетативные пропагулы, что свидетельствует о способности данного вида лишайника к вегетативному размножению до конца жизни.
В результате механического воздействия от таллома могут отделиться не только специализированные вегетативные диаспоры, но и целые фрагменты. Такие части таллома при попадании в благоприятные условия способны прикрепиться к субстрату и образовать новый таллом. В таком случае необходимость в процессе долихенизации отпадает.
Таким образом, в жизненном цикле L. pulmonaria есть несколько вариантов образования нового слоевища: от аскоспоры, вегетативной диаспоры или фрагмента таллома. При этом талломы, образовавшиеся разными способами, могут в процессе онтогенеза проходить через генеративное состояние, а могут его пропустить. Это связано с тем, что плодовые тела формируются только в генетически гетерогенных популяциях, т.е. при наличии гетероталличных талломов L. pulmonaria (Zoller et al., 1999). Установлено, что на процесс формирования апотециев оказывают влияние условия местообитания, прежде всего – давность нарушения сообщества, от которой зависит наличие подходящих условий для развития ценопопуляций и продолжительность колонизации (Игнатенко, Тарасова, 2018).
В ходе исследований стало очевидно, что крупные талломы вида в подходящих условиях среды обладают высоким потенциалом к образованию новых жизнеспособных талломов. Несмотря на то, что определить, из каких именно (половых или вегетативных) диаспор образовались те или иные ювенильные и имматурные талломы без генетических исследований невозможно, полученные данные позволяют предположить, что большая часть описанных ювенильных и имматурных талломов возникла вегетативным путем, поскольку фертильные талломы L. pulmonaria в изученных сообществах встречаются достаточно редко.
Приведенная в настоящей работе схема онтогенеза имеет ряд существенных отличий от имеющихся в литературе данных (Михайлова, 2005; Горшков, Семенова, 2008). Как показали наши данные, ювенильные талломы не превышают 0.15 см в длину и 0.12 см в ширину, а имматурные 1 – ≤0.45 и ≤0.24 см соответственно. Исходя из принципов дискретного выделения возрастных (онтогенетических) состояний у лишайников (Суетина, 2006; Суетина, Глотов, 2015) к ювенильному следует относить слоевища только накипной жизненной формы, а не в виде лопасти (чешуйки) (Горшков, Семенова, 2008), которые, на наш взгляд, являются имматурными. Момент появления цефалодиев на талломах – важнейший признак, который до этого в литературе не учитывался.
Способность к образованию ветегативных пропагул является одним из главных признаков, появляющихся в ходе онтогенеза лишайника, и, как правило, сохраняющийся в течение всей жизни таллома. Однако в работе В.В. Горшкова и Н.А. Семеновой (2008) наличие соредиев учитывается только в виргинильном 2а состоянии. В наших исследованиях, как и в работе И.Н. Михайловой (2005), при выделении виргинильных состояний (v1, v2a, v2b, v2c) ключевым признаком являлось обилие и расположение соралей, а также площадь таллома. Известно, что в онтогенетическом ряду от виргинильного 2а до виргинильного 2с увеличивается число и доля соралей, возрастают линейные размеры талломов (Михайлова, 2005). Показано, что изидии появляются на старых лопастях талломов, которые обильно покрыты соралями (виргинильное 2с). Важно, что вегетативные пропагулы продолжают существовать на старых, погибающих талломах и способны к распространению. Изидии и лобули в работе В.В. Горшкова и Н.А. Семеновой (2008) используются как отдельные признаки для выделения онтогенетических состояний виргинильное 2b и виргинильное 2с соответственно. В наших исследованиях, как и в работе И.Н. Михайловой (2005), установлено, что лобули формируются на старых, отмирающих лопастях на талломах, которые относятся к постгенеративному периоду. Таким образом, полученные данные существенно дополняют имеющуюся в литературе схему онтогенеза лишайника L. pulmonaria.
Список литературы
Андреева Е.Н., Баккал И.Ю., Горшков В.В. и др. Методы изучения лесных сообществ. СПб.: НИИ Химии СПбГУ, 2002. 240 с.
Горшков В.В., Семенова Н.В. Структура популяций Lobaria pulmonaria (L.) Hoffm. в лесах с разной давностью последнего нарушения в средней тайге Республики Коми // Современное состояние и пути развития популяционной биологии: Материалы X всероссийского популяционного семинара (Ижевск, 17–22 ноября 2008 г.). Ижевск, 2008. С. 113–115.
Ивантер Э.В., Коросов А.В. Введение в количественную биологию. Учебное пособие. Петрозаводск: Изд-во ПетрГУ, 2003. 304 с.
Игнатенко Р.В. Экология лишайника Lobaria pulmonaria (L.) Hoffm. в растительных сообществах Карелии: Автореф. дис. … канд. биол. наук. СПб.: БИН РАН, 2018. 26 с.
Игнатенко Р.В., Тарасова В.Н. Оценка состояния лишайника лобария легочная (Lobaria pulmonaria (L.) Hoffm.) в лесных сообществах с разной давностью нарушения на Северо-западе Европейской России // Экология. 2018. № 4. С. 245–253.
Игнатенко Р.В., Тарасова В.Н. Характеристика ювенильного и имматурного состояний прегенеративного периода онтогенеза охраняемого лишайника лобария легочная (Lobaria pulmonaria (L.) Hoffm.) // Современная микология в России. М.: Нац. акад. микол., 2017. Т. 6. С. 324–327.
Микрюков В.С. Популяционная экология эпифитного лишайника Lobaria pulmonaria (L.) Hoffm. на территории Урала и Сибири: Автореф. дис. … канд. биол. наук. Екатеринбург: ИЭРиЖ УрО РАН, 2011. 20 с.
Михайлова И.Н. Анализ субпопуляционных структур эпифитных лишайников (на примере Lobaria pulmonaria (L.) Hoffm.) // Вестник Нижегородского университета им. Н.И. Лобачева. 2005. № 1. Вып. 9. С. 124–134.
Михайлова И.Н., Воробейчик Е.Л. Размерная и возрастная структура популяций эпифитного лишайника Hypogymnia physodes (L.) Nyl. в условиях атмосферного загрязнения // Экология. 1999. № 2. С. 130–137.
Работнов Т.А. Жизненный цикл многолетних травянистых растений в луговых ценозах // Тр. БИН АН СССР. Сер. 3. Геоботаника. М., 1950. Вып. 6. С. 7–204.
Семенова Н.А. Критерии для выделения онтогенетических состояний лишайника Lobaria pulmonaria (L.) Th. Fr. // Актуальные проблемы биологии и экологии: Тезисы докладов XII молодежной научной конференции Института биологии Коми НЦ УрО РАН (Сыктывкар, Республика Коми, Россия 4–7 апреля 2005 г.). Сыктывкар, 2005. С. 212–213.
Ставрова Н.И., Горшков В.В., Катютин П.Н. Формирование структуры ценопопуляций лесообразующих видов в процессе послепожарного восстановления северотаежных лесов // Тр. КарНЦ РАН. Сер. Биогеография. 2016. № 3. С. 10–28.
Суетина Ю.Г. Морфологический подход в популяционно-онтогенетических исследованиях лишайников // Особь и популяция – стратегия: сборник материалов IX Всероссийского популяционного семинара (Уфа, 2–6 октября 2006 г.). Уфа, 2006. Ч. 2. С. 366–371.
Суетина Ю.Г. Онтогенез и популяционная структура Xanthoria parietina (L.) Th. Fr. в различных экологических условиях // Экология. 2001. № 3. С. 203–208.
Суетина Ю.Г., Глотов Н.В. Онтогенез и морфогенез кустистого лишайника Usnea florida (L.) Weber ex F.H. Wigg. // Онтогенез. 2010. Т. 41. № 1. С. 32–40.
Суетина Ю.Г., Глотов Н.В. Популяционно-онтогенетические исследования эпифитных лишайников // Теоретические проблемы экологии и эволюции: Шестые Любищевские чтения, 11-й Всероссийский популяционный семинар и Всероссийский семинар “Гомеостатические механизмы биологических систем” с общей темой “Проблемы популяционной экологии” (Тольятти, 6–10 апреля 2015 г.). Тольятти, 2015. С. 288–292.
Суетина Ю.Г., Ямбердова Е.И. Онтогенез и возрастно-виталитетная структура популяции лишайника Evernia prunastri (L.) Ach. // Вестник Удмуртского университета. Серия “Биология”. Науки о земле. 2010. Вып. 3. С. 44–52.
Уранов А.А. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов // Биол. науки. 1975. № 2. С. 7–34.
Фенько А.А., Тарасова В.Н. Распространение и состояние охраняемого лишайника бриория Надворника (Bryoria nadvornikiana (Gyeln.) Brodo et D. Hawksw.) в лесных сообществах Петрозаводского городского округа // Тр. КарНЦ РАН. Сер. Экологические исследования. 2016. № 4. С. 83–91.
Andersson L., Appelqvist T. Lobaria pulmonaria and Gyalecta ulmi as indicators of deciduous woodland with high nature qualities // Svensk Botanisk Tidskrift. 1987. V. 81. P. 185–194.
Andersson L., Martverk R., Külvik M. et al. Woodland Key Habitat Inventory in Estonia 1999–2002. Tartu: Regio Publishing, 2003. 192 p.
Campbell J., Fredeen A.L. Lobaria pulmonaria abundance as an indicator of macrolichen diversity in Interior Cedar-Hemlock forests of east-central British Columbia // Canadian J. Botany. 2004. V. 82. 970–982.
Cornejo C., Scheidegger C. New morphological aspects of cephalodium formation in the lichen Lobaria pulmonaria (Lecanorales, Ascomycota) // The Lichenologist. 2013. V. 45(1). P. 77–87.
Culberson W.L., Egan R.S., Esslinger T.L., Hodkinson B.P. Recent literature on lichens. 2019. http://nhm2.uio.no/ lichens/rll.html. [Presented on the Web by E. Timdal. First posted 1997.04.14, continuously updated.]
Denison W.C. Apothecia and ascospores of Lobaria oregana and Lobaria pulmonaria investigated // Mycologia. 2003. V. 95. P. 513–518.
Edman M., Eriksson A.-M., Villard M.-A. Effects of selection cutting on the abundance and fertility of indicator lichens Lobaria pulmonaria and Lobaria quercizans // Journal of Applied Ecology. 2008. V. 45(1). P. 26–33.
Gauslaa Y. Lobaria pulmonaria, an indicator of species-rich forest of long ecological continuity // Blyttia. 1994. V. 52. P. 119–128.
Gauslaa Y. Population structure of the epiphytic lichen Usnea longissima in a boreal Picea abies canopy // Lichenologist. 1997. V. 29(5). P. 455–469.
Gauslaa Y., Lie M., Solhaug K.A. et al. Growth and ecophysiological acclimation of the foliose lichen Lobaria pulmonaria in forests with contrasting light climates // Oecologia. 2006. V. 147(3). P. 406–416.
Jüriado I., Liira J., Csencsics D. et al. Dispersal ecology of the endangered woodland lichen Lobaria pulmonaria in managed hemiboreal forest landscape // Biodiversity Conservation. 2011. V. 20. P. 1803–1819.
Liira J., Sepp T. Indicators of structural and habitat natural quality in boreo-nemoral forests along the management gradient // Annales Botanici Fennici. 2009. V. 46. P. 308–325.
Nascimbene J., Brunialti G., Ravera S. et al. Lobaria pulmonaria (L.) Hoffm. as an indicator of lichen conservation importance of Italian forests // Ecological Indicators. 2010. V. 10. P. 353–360.
Pykälä J. Effects of new forestry practices on rare epiphytic macrolichens // Conservation Biology. 2004. V. 18. P. 831–838.
Radies D.N., Coxson D.S. Macrolichen colonization on 120–140 year old Tsuga heterophylla in wet temperate rainforests of central-interior British Columbia: a comparison of lichen response to even-aged versus old-growth stand structures // Lichenologist. 2004. V. 36(3–4). P. 235–247.
Ravera S., Massari G., Genovesi V. Phytoclimatic characterization of lichen habitats in central Italy // Nova Hedwigia. 2006. V. 82. P. 143–165.
Rose F. Temperate forest management: it effects on bryophyte and lichens floras and habitats // Bryophytes and Lichens in a Changing Environment / Eds. Bates J.W., Farmer A.M. Oxford: Oxford Scientific Publications, Clarendon Press, 1992. P. 211–233.
Scheidegger C., Frey B., Walser J.C. Reintroduction and augmentation of populations of the endangered Lobaria pulmonaria: methods and concepts // Lobarion Lichens as Indicators of the Primeval Forests of the Eastern Carpathians / Eds. Kondratyuk S.Y., Coppins B.J. (Kiev, 25–30 May 1998). Kiev, 1998. P. 33–52.
Scheidegger C., Werth S. Conservation strategies for lichens: insights from population biology // Fungal Biol. Rev. 2009. V. 23. P. 55–66.
Tarasova V.N., Favorskaja M.A., Shreders M.A. State of Evernia divaricata (L.) Nyl. populations in forest communities of “Kivach” strict nature reserve (South Karelia) // Field Meeting “Lichen of Boreal Forests” and the Fourth Russian Lichenogical Workshop: Proceedings. Syktyvkar, 2008. P. 183–196.
Tschermak-Woess E. Dictyochloropsis splendida (Chlorophyta), the correct phycobiont of Phlyctis argena and the high degree of selectivity or specificity involved // Lichenologist. 1995. V. 27. P. 169–187.
Tschermak-Woess E. The algal partner // CRC handbook of lichenology / Ed. Galun M. Boca Raton, FL, USA: CRC Press Inc., 1988. V. 1. P. 39–92.
Walser J.-C., Zoller S., Büchler U. Species-specific detection of Lobaria pulmonaria (lichenized ascomycete) diaspores in litter samples trapped in snow cover // Mol. Ecol. 2001. V. 10(9). P. 2129–2138.
Werth S., Wagner H.H., Gugerli F. et al. Quantifying dispersal and establishment limitation in a population of an epiphytic lichen // Ecology. 2006. V. 87(8). P. 2037–2046.
Yoshimura I. Lung lichens and their vegetation in Japan and the other regions // Lobarion Lichens as Indicators of the Primeval Forests of the Eastern Carpathians / Eds. Kondratyuk S.Y., Coppins B.J. (Kiev, 25–30 May 1998). Kiev, 1998. P. 53–63.
Yoshimura I. The genus Lobaria of Eastern Asia // J. Hattori Bot. Lab. 1971. V. 34. P. 231–364.
Zoller S., Lutzoni F., Scheidegger C. Genetic variation within and among populations of the threatened lichen Lobaria pulmonaria in Switzerland and implications for its conservation // Molec. Ecol. 1999. V. 8(12). P. 2049–2059.
Дополнительные материалы отсутствуют.