

Онтогенез, 2021, T. 52, № 2, стр. 137-145
Начальные этапы онтогенеза представителей рода Hydrangea L. в условиях культуры
Ф. К. Мурзабулатова a, *, З. Х. Шигапов a, Н. В. Полякова a
a Южно-Уральский ботанический сад-институт Уфимского федерального исследовательского центра
Российской академии наук
450080 Республика Башкортостан, Уфа, ул. Менделеева, 195, корп. 3, Россия
* E-mail: murzabulatova@yandex.ru
Поступила в редакцию 28.04.2020
После доработки 14.09.2020
Принята к публикации 21.09.2020
Аннотация
Описаны начальные этапы онтогенеза видов рода Hydrangea L. Исследования проводились на протяжении 2007–2015 гг. Цель данной работы – изучить и описать начальные этапы онтогенеза представителей рода Hydrangea в условиях культуры в Башкирском Предуралье. В ходе исследований были решены следующие задачи: определены этапы виргинильного возрастного состояния изучаемых видов гортензии; описаны признаки, характерные для каждого этапа; охарактеризованы морфологические параметры растений на каждом этапе развития; выявлены особенности развития изученных видов на начальных этапах онтогенеза. Описаны 4 этапа виргинильного возрастного состояния: проростки, ювенильные, имматурные и взрослые вегетативные растения. Этап ювенильных растений у гортензий наступает в среднем через 25 дней после прорастания семян с появлением 2-й пары настоящих листьев; имматурных – в основном на 3 году, после начала ветвления осевого побега; виргинильных – на 4–5 году жизни с началом кущения и приобретением жизненной формы взрослого растения.
ВВЕДЕНИЕ
Род Гортензия (Hydrangea L., Hydrangeaceae Dumort.) включает в себя красивоцветущие кустарники, естественный ареал распространения которых – Китай, Гималаи, Япония, Северная и Южная Америка, единично – Филиппины и о. Ява. В настоящее время род включает около 200 видов и более 350 сортов (van Gelderen, 2004). В последние годы гортензии стали чрезвычайно популярны в декоративном садоводстве и ландшафтном дизайне, и, следовательно, возникла необходимость в изучении их роста и развития в условиях конкретного региона и климатической зоны.
В настоящее время подробное анатомическое и морфологическое описание семян гортензий дано в работах L. Hufford (1995), А.Л. Тахтаджяна (2000), Т.А. Лобовой (2000), M. Morozowska, A. Woznicka, M. Kujawa (2012). Семена гортензий многочисленные, мелкие, 0.4–1.5 мм длины, 0.5–2 мм ширины, 0.3–1 мм толщины. Цвет семян от золотисто-желтого до темно-коричневого. Форма семян разнообразная: основная часть видов имеет веретеновидные, обратнояйцевидные, широкояйцевидные и овальные семена. Н.А. Коляда (2007) в своих опытах с 5 видами гортензий описала морфологию семян, количественные параметры, имеющие важное значение для познания их биологии. В частности, максимальный вес 1000 шт. семян отмечен ею у H. paniculata – 0.091 г, H. bretschneideri – 0.075 г, H. paniculata f. grandiflora – 0.060 г. В природе, по данным M. Hara и др. (1999), а также H. Kanno и K. Seiwa (2004) у H. paniculata средний вес семян составляет 1.01 г. Вероятно, такая разница в весе семян в природных условиях и при интродукции может свидетельствовать о кардинальной разнице в комплексе климатических условий, в которых происходит завязывание и созревание семян. О прорастании семян видов гортензий имеются данные в работах Н.А. Коляды (2007). По ее сведениям, в дендрарии Горно-таежной станции ДВО РАН лабораторная всхожесть семян гортензии достаточно высокая и в среднем составляет 79%, а у H. bretschneideri она достигает 90%. По данным Л.А. Семкиной и Ю.В. Цыкарева (1994), в условиях культуры на Среднем Урале H. bretschneideri обильно цветет, но всхожесть ее семян в лабораторных условиях составляет всего 37%. В условиях Ботанического сада г. Киева (Коркуленко, 2013) семена интродуцированных гортензий имеют высокую всхожесть и не нуждаются в предпосевной подготовке, лабораторная всхожесть их достигает 99.3%.
Данных по особенностям роста и морфологии проростков и ювенильных растений гортензий при интродукции в литературе нет. Имеются некоторые данные по изучению морфологии и онтогенеза цветков гортензий (Roels et al., 1997; Hufford, 2001).
М.Т. Мазуренко и А.П. Хохрякова (1977) изучали особенности прорастания семян и развития сеянцев гортензии крупнолистной (H. opuloides) в естественных местообитаниях в окрестностях г. Батуми (Грузия); в их описаниях отсутствует информация о характере роста проростков. Данными авторами установлено, что в системе побегов формирования может развиться до 6 порядков побегов, при этом основной цикл проходит всего за 3, реже 4 года. Его периоды:
1. Рост побега формирования и побегов ветвления первого порядка (первый год жизни, реже – начало второго года).
2. Развитие побегов ветвления первого-второго, реже третьего порядков (второй год жизни).
3. Отмирание верхушки системы побегов формирования, развитие и отмирание побегов ветвления третьего-шестого порядков (третий-четвертый год жизни).
Таким образом, число порядков побегов здесь больше, чем число лет основного цикла. Это связано, очевидно, с более длительным периодом вегетации в условиях теплого климата и, как следствие, закладкой и ростом большего количества почек и побегов (Мазуренко, Хохряков, 1977).
Анализ литературы по теме данной работы показывает, что биологические особенности интродуцированных видов гортензий изучались на базе коллекций крупных ботанических садов в России (г. Москва) и Украине (г. Киев). Сведения о биологии видов гортензий в других регионах носят не полный и отрывочный характер. Информации по культуре гортензий на Урале (в т.ч. на Южном Урале) и в Приуралье практически нет. Аспекты онтогенеза видов гортензий в культуре как в зарубежной, так и в отечественной литературе не освещены, а именно: отсутствует полное описание онтогенетического развития гортензий, нет данных по морфометрическим параметрам растений в каждом возрастном состоянии, не выявлены особенности индивидуального развития на каждом этапе онтогенеза. Это обусловливает необходимость обобщения существующих данных, а также их уточнения и детализации. Таким образом, актуальность и новизна представленных результатов очевидна. Цель данной работы – изучить и описать начальные этапы онтогенеза представителей рода Hydrangea в условиях культуры в Башкирском Предуралье. В соответствии с этой целью были поставлены следующие задачи:
– определить этапы виргинильного возрастного состояния изучаемых видов гортензии;
– описать признаки, характерные для каждого этапа;
– охарактеризовать морфологические параметры растений на каждом этапе развития;
– выявить особенности развития изученных видов на начальных этапах онтогенеза.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводились на базе коллекции гортензий Южно-Уральского ботанического сада-института Уфимского федерального исследовательского центра РАН (ЮУБСИ УФИЦ РАН) в период с 2007 по 2015 годы (Мурзабулатова, Полякова, 2016). Объектами исследования являлись 6 видов и 1 подвид рода Hydrangea, произрастающие на коллекционном участке декоративных кустарников (Фрутицетуме):
Подсекция Americanae
Виды: гортензия древовидная (H. arborescens L.);
гортензия лучистая (H. arborescens subsp. radiata (Walter) McClintock);
гортензия пепельная (H. cinerea Small).
Подсекция Heteromallae
Виды: гортензия метельчатая (H. paniculata Siebold);
гортензия почвопокровная (H. heteromalla D. Don);
гортензия Бретшнейдера (H. bretschneideri Dippel);
гортензия золотистожилковая (H. xanthoneura Diels).
Климатические условия района исследований имеют свои особенности и отличия от условий естественного произрастания гортензий. Климат Башкирского Предуралья континентальный и характеризуется холодной зимой и теплым летом, большой амплитудой колебаний температуры в течение года, ранними осенними и поздними весенними заморозками. В г. Уфа среднемноголетняя температура воздуха составляет +3.4°C, средняя температура января –13.5°С, абсолютный минимум достигает –48.5°С, зимой нередки оттепели, безморозный период длится в среднем 144 дня. Средняя температура июля +19.5°C, абсолютный максимум зафиксирован на уровне +40°С. Климат достаточно влажный: среднегодовое количество осадков составляет 500–590 мм, в т.ч. около 350 мм в теплый период (максимум приходится на июнь–июль). Снежный покров устанавливается в ноябре и держится в среднем 155 сут (Путенихин и др., 2017).
Как известно, в онтогенезе растений выделяют 4 периода (Серебряков, 1952, 1964; Уранов, 1967; Borchert, 1975; Hackett, 1985; Рекомендации по изучению онтогенеза.., 1990; Donovan, Ehleringer, 1991; Paiva, Machado, 2006; Wang et al., 2011; Albert et al., 2012):
– латентный (состояние семени);
– виргинильный (включающий этапы проростков, ювенильных, имматурных и взрослых вегетативных растений);
– генеративный (этапы молодых, средневозрастных и старых генеративных растений);
– сенильный (субсенильный и сенильный этапы).
Поскольку гортензии являются древесными долгоживущими растениями, мы в наших исследованиях ограничились описанием этапов виргинильного периода.
В исследованиях были задействованы по 30 экземпляров каждого изучаемого таксона. Все они были этикетированы, каждые 7 дней фиксировались изменения их частей и органов в процессе развития, отмечались признаки перехода одного этапа возрастного состояния в другой. Для проведения замеров морфологических параметров и изучения строения корневой системы растений на каждом этапе выкапывались модельные растения, которые фотографировались или зарисовывались.
Статистическая обработка данных проводилась с помощью пакета анализа Microsoft Excel и статистических программ Statistica 6.0; Statistica 10.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Анализ роста и развития гортензий при грунтовом посеве позволил описать 4 этапа виргинильного возрастного состояния: проростки, ювенильные, имматурные и взрослые вегетативные растения.
Проросток (plantules (Горышина и др., 1988)). Состояние проростков определяется с момента появления семядолей до их отмирания или же до момента появления первой пары настоящих листьев. У проростков изученных нами видов гортензий наблюдалась следующая очередность развития: рост первичного корешка > рост гипокотиля > вынос семядолей > разворачивание семядолей > рост семядолей > начало роста почечки > разворачивание и рост первой пары настоящих листьев (рис. 1, 2).


На этом этапе у некоторых представителей подсекции Heteromallae наблюдались отклонения в сторону увеличения количества семядолей (поликотилия) обычно до 3-х (рис. 3).

Начало роста корней 1-го порядка происходит на стадии роста семядолей. В период формирования почечки проростки имеют главный корень и несколько боковых корешков. Гипокотиль голый, вначале беловатый, затем постепенно приобретает зеленую окраску. Длина гипокотиля представителей подсекции Americanae до 0.4 см, а у проростков из подсекции Heteromallae – до 0.2 см. Семядольные листья светло-зеленые, голые, с еле заметной центральной жилкой. Верхушка семядольных листьев сеянцев у представителей подсекции Americanae округлая, а в подсекции Heteromallae слегка заостренная. У представителей подсекции Heteromallae междоузлия укорочены, расстояние между ними составляет до 1.5 мм. Продолжительность жизни семядольных листьев составляет в среднем от 56 дней (H. heteromalla) до 77 дней (H. paniculata) (табл. 1). Окончание вегетации сеянцев на первом году жизни знаменуется закладкой апикальной почки и завершением роста побега. Сеянцы видов из подсекции Americanae (H. arborescens, H. cinerea, H. arborescens subsp. radiata) не успевают закончить вегетацию и сформировать апикальную почку, но семядольные листья у них усыхают и опадают так же, как у видов из другой подсекции.
Таблица 1.
Основные фазы роста и развития проростков гортензий (в днях от начала прорастания)
| Вид | Начало роста | Окончание роста сеянца |
Усыхание семядолей | ||
|---|---|---|---|---|---|
| 1-й пары листьев | 2-й пары листьев | 3-й пары листьев | |||
| Подсекция Americanae | |||||
| H. arborescens | 13 ± 0.17 | 24 ± 0.42 | 40 ± 1.32 | –* | 70 ± 1.16 |
| H. cinerea | 14 ± 0.56 | 31 ± 1.64 | 40 ± 1.59 | –* | 70 ± 0.69 |
| H. arborescens subsp. radiata | 19 ± 0.47 | 35 ± 0.40 | 43 ± 0.32 | –* | 75 ± 1.91 |
| Подсекция Heteromallae | |||||
| H. paniculata | 22 ± 0.48 | 27 ± 0.29 | 35 ± 0.35 | 92 ± 0.23 | 77 ± 0.39 |
| H. bretschneideri | 12 ± 0.16 | 18 ± 0.42 | 32 ± 0.29 | 108 ± 1.32 | 57 ± 0.96 |
| H. heteromalla | 12 ± 0.13 | 17 ± 0.58 | 32 ± 0.25 | 107 ± 1.26 | 56 ± 0.88 |
| H. xanthoneura | 12 ± 0.15 | 25 ± 0.87 | 39 ± 0.97 | 97 ± 0.86 | 71 ± 0.93 |
Минимальной высотой осевого побега отличаются сеянцы H. xanthoneura (табл. 2), а максимальной – H. arborescens, у которой отмечено наибольшее количество метамеров. К концу сезона осевая часть побега одревесневает. В пазухах семядольных листьев и нижней пары листьев закладываются почки возобновления. Главный корень имеет наименьший размер у H. arborescens (в среднем 3.2 см), а самый длинный у H. heteromalla (в среднем 12.6 см), с развитием надземной части проростков увеличиваются боковые корни. Первые настоящие листья сеянцев имеют эллиптическую и яйцевидно-продолговатую форму листовой пластинки. Окраска листьев сверху ярко-зеленая, снизу – светло-зеленая.
Таблица 2.
Высота однолетних сеянцев и длина центрального корня
| Название вида | Среднее количество междоузлий, шт. | Средняя высота, см | Средняя длина центрального корня, см |
|---|---|---|---|
| Подсекция Americanae | |||
| H. arborescens | 3.7 ± 0.18 | 3.9 ± 0.34 | 3.2 ± 0.15 |
| H. arborescens subsp. radiata | 1.7 ± 0.21 | 2.1 ± 0.07 | 9.5 ± 1.21 |
| H. cinerea | 1.7 ± 0.13 | 2.0 ± 0.16 | 4.8 ± 0.38 |
| Подсекция Heteromallae | |||
| H. paniculata | 2.3 ± 0.10 | 1.7 ± 0.09 | 4.7 ± 0.32 |
| H. bretschneideri | 2.0 ± 0.14 | 1.2 ± 0.12 | 9.7 ± 0.60 |
| H. heteromalla | 1.6 ± 0.13 | 0.8 ± 0.04 | 12.6 ± 0.37 |
| H. xanthoneura | 1.6 ± 0.15 | 0.6 ± 0.12 | 6.8 ± 0.21 |
Первая пара листьев с максимальным размером зафиксирована нами у H. bretschneideri, а самая мелкая – у H. paniculata. Наиболее крупная вторая пара листьев отмечена у H. heteromalla, а минимальный размер листовой пластинки второй пары также у H. paniculata. Корневая система однолетних сеянцев характеризуется удлинением главного корня и увеличением количества боковых корешков. Наибольшей высотой в первый год жизни характеризовались H. arborescens (3.9 см в среднем) и H. paniculata (2.3 см), наименьшей – H. heteromalla (0.8 см в среднем) и H. xanthoneura (0.6 см). Максимальный диаметр стволика у корневой шейки отмечен у H. xanthoneura, H. heteromalla и H. bretschneideri (от 2.2 до 2.6 мм), минимальный – у H. cinerea (1.6 мм). Связь между высотой растений и диаметром ствола умеренная отрицательная, коэффициент корреляции r = –0.52 (p < 0.05). Дисперсионный анализ показал, что виды гортензии различаются уже на начальных этапах онтогенеза как по высоте однолетних сеянцев (F = 81.900; p < < 0.001), так по длине центрального корня (F = = 45.91; p < 0.001).
Ювенильные растения (juvenile plants (Hackett, 1985; Popov et al., 2004)). Данный этап длится с момента разворачивания второй пары настоящих листьев до начала ветвления (Серебряков, 1952; Рекомендации по изучению онтогенеза…, 1990). У представителей обеих подсекций первые настоящие листочки похожи на листья взрослых растений, но меньшего размера и имеют более редкое опушение; размер листовых пластинок увеличивается пропорционально высоте сеянцев. В этом возрастном состоянии наблюдается рост эпикотиля сеянцев подсекции Americanae, последовательное разворачивание и рост 3-го, 4-го и последующих настоящих листьев, прекращение роста семядольных листьев, прекращение апикального роста и формирование терминальной почки (только у видов подсекции Heteromallae, в подсекции Americanae терминальная почка не закладывается), изменение окраски семядолей, отмирание и опадение их, прекращение радиального роста сеянцев. Ювенильные растения имеют моноподиально (одноосно) растущий побег, корневая система состоит из главного и боковых корней.
В ювенильном возрасте (второй год жизни) самые высокие сеянцы зафиксированы у H. arborescens и H. arborescens subsp. radiata (13.0 и 13.9 см), Наименьший показатель зафиксирован у H. bretschneideri и H. heteromalla (10.0 и 10.6 см) (табл. 3), коэффициент вариации высоты видов колеблется от 7 до 61%. Наибольший диаметр стволика зафиксирован у H. bretschneideri (3.9 мм), а наименьший показатель – у H. paniculata (2.5 мм). Связь между высотой растений и диаметром ствола умеренно положительная и коэффициент корреляции r = 0.35 (p < 0.05). Сохраняется и развивается осевой корень, также наблюдается увеличение количества боковых корней до 4–5-го порядка (рис. 4). Виды, входящие в подсекцию Americanae, в условиях ботанического сада не успевают формировать верхушечные почки (без кроющих чешуй) и кончики побегов засыхают или обмерзают и у некоторых сеянцев на втором году начинается ветвление. Дисперсионный анализ подтверждает, что на стадии ювенильных растений сеянцы гортензий также имеют различия по высоте (F = 2.59; p < 0.001) и длине центрального корня (F = 17.256; p < 0.001).
Таблица 3.
Средние значения морфометрических параметров и их изменчивость у 2-летних сеянцев (ювенильный период) гортензий
| Название вида | Средняя высота растения | Средняя длина осевого корня | Средняя величина диаметра корневой шейки | Среднее количество боковых побегов, шт. | |||
|---|---|---|---|---|---|---|---|
| высота, см | CV, % | длина, см | CV, % | диаметр, мм | CV, % | ||
| Подсекция Americanae | |||||||
| H. arborescens | 13.0 ± 0.63 | 16.8 | 15.8 ± 0.6 | 11.7 | 3.5 ± 0.16 | 20.9 | 0.4 ± 0.2 |
| H. cinerea | 10.8 ± 0.83 | 22.4 | 17.6 ± 0.9 | 17.2 | 3.5 ± 0.21 | 32.4 | 0 |
| H. arborescens subsp. radiata | 13.9 ± 0.99 | 28.1 | 15.4 ± 0.7 | 15.1 | 3.7 ± 0.20 | 14.1 | 0.5 ± 0.2 |
| Подсекция Heteromallae | |||||||
| H. paniculata | 12.8 ± 1.35 | 23.0 | 9.8 ± 0.3 | 10.6 | 2.5 ± 0.28 | 39.5 | 0 |
| H. bretschneideri | 10.0 ± 0.79 | 34.2 | 12.5 ± 0.6 | 16.7 | 3.9 ± 0.19 | 22.4 | 0 |
| H. heteromalla | 10.6 ± 0.7 | 7.0 | 15.7 ± 0.3 | 12.4 | 3.7 ± 0.21 | 19.4 | 0 |
| H. xanthoneura | 12.8 ± 1.78 | 61.0 | 14.8 ± 1.10 | 24.9 | 3.8 ± 0.24 | 17.6 | 0 |
Рис. 4.
Этапы виргинильного возрастного состояния видов гортензий: проростки (p1, p2, p3, p4), ювенильные (j), имматурные (im) и взрослые вегетативные (v) растения.
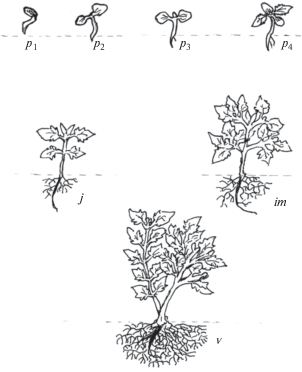
Имматурные растения (immature plants). Этот этап начинается с ветвления сеянцев (Серебряков, 1952; Рекомендации по изучению онтогенеза…, 1990) и является переходным от ювенильных растений к взрослым вегетативным. У некоторых сеянцев гортензий ветвление начинается во втором вегетационном сезоне; по нашим наблюдениям, количество боковых ветвей сеянцев различно в зависимости от принадлежности к подсекциям (табл. 4). У видов гортензий имматурный возрастной период в основном начинается на третьем году жизни. У представителей обеих подсекций наблюдается формирование и рост боковых побегов (рис. 4) наибольшее количество которых наблюдается у H. arborescens subsp. radiata (табл. 4), небольшое количество их у видов H. bretschneideri и H. heteromalla. Корневая шейка у всех сеянцев утолщается, максимальный прирост отмечен у H. bretschneideri (6.8 мм), минимальный – у H. cinerea (5.1 мм). В этом возрасте рост центрального корня у всех видов замедляется, наблюдается утолщение центрального и боковых корней и увеличение количества боковых корней. На третьем году жизни рост осевого побега продолжается, и трехлетние сеянцы гортензий достигают высоты 17.2–61.6 см и 5.3–6.8 мм в диаметре у корневой шейки (табл. 4). Максимальной высоты в этом возрасте достигли H. arborescens (65.8 см у отдельных особей) и H. bretschneideri (61.6 см), наименьший показатель зафиксирован у H. paniculata (17.2 см) и H. xanthoneura (24.5 см). Длительность имматурного возрастного состояния у видов гортензий составляет обычно один год.
Таблица 4.
Средние значения морфометрических параметров и их изменчивость у сеянцев в имматурном возрастном состоянии
| Вид | Средняя высота растения | Средняя длина осевого корня | Средняя величина диаметра корневой шейки | Количество боковых побегов, шт. | |||
|---|---|---|---|---|---|---|---|
| высота, см | CV, % | длина, см | CV, % | диаметр, мм | CV, % | ||
| Подсекция Americanae | |||||||
| H. arborescens | 65.8 ± 2.39 | 19.0 | 15.8 ± 3.1 | 16.8 | 5.9 ± 0.31 | 17.9 | 1.9 ± 0.3 |
| H. cinerea | 53.9 ± 3.60 | 19.9 | 14.3 ± 5.2 | 34.3 | 5.1 ± 0.21 | 10.4 | 1.7 ± 0.4 |
| H. arborescens subsp. radiata | 40.2 ± 6.64 | 42.0 | 16.1 ± 4.1 | 26.4 | 6.2 ± 0.42 | 27.8 | 2.0 ± 0.4 |
| Подсекция Heteromallae | |||||||
| H. paniculata | 17.2 ± 1.80 | 11.7 | 12.4 ± 4.2 | 31.8 | 5.4 ± 0.38 | 24.3 | 1.7 ± 0.3 |
| H. bretschneideri | 61.6 ± 5.59 | 56.9 | 13.4 ± 3.2 | 23.7 | 6.8 ± 0.30 | 28.0 | 0.9 ± 0.2 |
| H. heteromalla | 52.7 ± 6.39 | 30.9 | 15.5 ± 4.5 | 29.3 | 6.2 ± 0.47 | 14.9 | 0.9 ± 0.2 |
| H. xanthoneura | 24.5 ± 3.70 | 44.4 | 13.6 ± 5.4 | 39.6 | 5.3 ± 0.48 | 31.2 | 1.7 ± 0.1 |
У единичных сеянцев, особенно в подсекции Heteromallae, наблюдался ускоренный темп развития, который приводил к переходу сеянцев в генеративное состояние и к появлению зацветающих ювенильных (H. paniculata) и имматурных (H. xanthoneura) растений.
Дисперсионный анализ показал, что на стадии имматурных растений виды гортензий имеют различия по высоте (F = 18.143; p < 0.001), в то время как различий по длине центрального корня уже не отмечается (F = 1.500; p = 0.186).
Взрослые вегетативные растения (adult phases of vegetative development in plants (Wang et al., 2011)). В этот этап развития сеянцы видов гортензий переходят на 4–5 год жизни. На первичном кусте развиваются первые ростовые побеги высотой от 13.5 до 37.9 см (табл. 5), они являются побегами формирования (Мазуренко, Хохряков, 1977; Чубарь, 2013); при этом наблюдается кущение растений. У представителей обеих подсекций в основании главного побега над почвой (аэроксильно) из спящих почек (почки возобновления) (рис. 5) начинают расти побеги; в дальнейшем появляются и побеги из подземных почек (аэроксильно-геоксильные) (Капранова, 1980; Тищенко, 2004) (рис. 5).
Таблица 5.
Количество и высота побегов у взрослых вегетативных растений гортензий
| Название вида | Высота побегов, см | Количество побегов, шт. | ||
|---|---|---|---|---|
| среднее | min | max | ||
| Подсекция Americanae | ||||
| H. arborescens | 36.0 ± 5.61 | 10.3 | 59.5 | 2 |
| H. cinerea | 37.9 ± 3.76 | 15.0 | 53.6 | 2 |
| H. arborescens subsp. radiata | 27.9 ± 4.01 | 10.5 | 60.2 | 2 |
| Подсекция Heteromallae | ||||
| H. paniculata | 13.5 ± 2.77 | 5.6 | 23.0 | 2 |
| H. bretschneideri | 14.7 ± 1.15 | 9.0 | 23.7 | 2 |
| H. heteromalla | 16.3 ± 10.5 | 9.0 | 23.0 | 2 |
| H. xanthoneura | 18.5 ± 1.88 | 10.5 | 35.6 | 2 |
Рис. 5.
H. arborescens subsp. radiata. 1 – надземные побеги или побеги формирования; 2 – подземные почки; 3 – придаточные корни; 4 – главный корень.

Самая большая высота побегов формирования (будущие скелетные оси) на этапе взрослых вегетативных растений отмечена у H. cinerea (среднее значение 37.9 см), самые низкие побеги формирования на этом этапе зафиксированы у H. paniculata (среднее значение 13.5 см) (табл. 5). Количество этих побегов в подсекции Americanae насчитывается от 1 до 2 на одном растении, в подсекции Heteromallae у отдельных растений побеги формирования не появились и эти экземпляры остались в имматурном состоянии. У взрослых вегетативных растений образуется масса придаточных корней, главный корень со временем перестает среди них выделяться и теряется в общей массе корней (рис. 5).
ЗАКЛЮЧЕНИЕ
При изучении начальных этапов онтогенеза видов гортензий коллекции ЮУБСИ УФИЦ РАН выявлены особенности 4 этапов виргинильного возрастного состояния: проростки, ювенильные, имматурные и взрослые вегетативные растения. В связи с отсутствием литературных данных по изучению онтогенеза гортензий в других регионах и, следовательно, сравнений, приводятся результаты собственных наблюдений.
Максимальной высотой в первый год жизни характеризуются сеянцы H. arborescens (3.9 см) и H. paniculata (2.3 см), минимальной – H. heteromalla (0.8 см) и H. xanthoneura (0.6 см). Среди трехлетних сеянцев наименьшая изменчивость по высоте наблюдалось у H. paniculata (17.2%); наибольший показатель коэффициента вариабельности по высоте отмечен у H. bretschneideri (56.9%), а по диаметру корневой шейки наименьший показатель зафиксирован у H. cinerea (10.4%), максимальный – у H. xanthoneura (31.2%). Максимальной скоростью роста обладают сеянцы H. arborescens, медленнее всех растут сеянцы H. paniculata.
Переход проростков в ювенильные растения происходит после появления второй пары настоящих листьев, в среднем через 25 дней после прорастания. Моментом перехода от ювенильных к имматурным растениям можно считать начало ветвления в средней и верхней части первичного побега, т.е. рост побегов II порядка, который происходит в основном на 3 году жизни сеянцев. Переход к взрослым вегетативным растениям связан с началом отрастания побегов формирования из спящих почек (почки возобновления) в основании первичного побега (Мазуренко, Хохряков, 1977; Чубарь, 2013). При дальнейшем росте увеличивается размер растений за счет кущения. Главный корень в начале роста сеянцев хорошо выражен, по мере роста начинает сильно ветвиться, и теряется в общей массе корней.
Список литературы
Горышина Т.Г., Гребенщиков О. С., Ниценко А.А. Терминологический словарь по экологии, геоботанике и почвоведению: Русско-англо-немецко-французский. Л.: Изд-во Ленингр. ун-та, 1988. 250 с.
Капранова Н.Н. Сравнительное морфологическое исследование некоторых видов чубушника в онтогенезе: Автореф. дис. … канд. биол. наук. М., 1980. 26 с.
Коляда Н.А. Качество семян некоторых видов семейства Гортензиевые в дендрарии Горно-таежной станции ДВО РАН // Мат-лы Междунар. науч. конф., посвященной 75-летию со дня образования Центрального ботанического сада НАН Беларуси. Минск. 2007. Т. 1. С. 176–178.
Коркуленко О.М. Особливостi плодоношеня iнтродукованих видiв та культиварiв роду Hydrangea L. // Науковий вiсник НУБiП Украiни. Серiя: Лiсiвництво та декоративне садiвництво. Киiв. 2013. № 187–1. С. 75–80.
Лобова Т.А. Анатомо-морфологическая характеристика семян группы семейств порядка Hydrangeales: Автореф. дис. … канд. биол. наук. СПб, 2000. 23 с.
Мазуренко М.Т., Хохряков А.П. Структура и морфогенез кустарников. М.: Наука, 1977. 159 с.
Мурзабулатова Ф.К., Полякова Н.В. Коллекционный фонд рода Гортензия (Hydrangea L.) в ботаническом саду-институте Уфимского научного центра РАН // Известия Уфимского научного центра РАН. 2016. № 4. С. 69–76.
Путенихин В.П., Путенихина К.В., Шигапов З.Х. Кедр сибирский в Башкирском Предуралье и на Южном Урале: биологические и лесоводственные особенности при интродукции. Уфа: Башкирская Энциклопедия, 2017. 248 с.
Рекомендации по изучению онтогенеза в ботанических садах СССР. Киев, 1990. 378 с.
Семкина А.А., Цыкарев Ю.В. Особенности прорастания семян гортензии Бретшнейдера // Тезисы докладов совещания по семеноведению интродуцентов. Москва, 1994. С. 35–36.
Серебряков И.Г. Морфология вегетативных органов высших растений. М.: Советская наука, 1952. 391 с.
Cеребряков И.Г. Жизненные формы высших растений и их изучение. // Полевая геоботаника. М.-Л.: Наука, 1964. Т. 3. С. 146–205.
Тахтаджян А.Л. Сравнительная анатомия семян. Том 6. Двудольные. Rosidae II. СПб.: Наука, 2000. 455 с.
Тищенко М.П. Онтогенез смородины темно-пурпуровой (Ribes atropurpureum C.A. Mey) // Онтогенетический атлас лекарственных растений: Научное издание. Т. IV. Йошкар-Ола: МарГУ, 2004. С. 43–48.
Уранов А.А. Онтогенез и возрастной состав популяций // Онтогенез и возрастной состав популяций цветковых растений. М.: Наука, 1967. С. 3–8.
Чубарь Е.А. Онтогенез Дальневосточных видов рода Deutzia (Hydrangeaceae) // Ботанический журн. 2013. Т. 98. № 12. С. 1524–1541.
Albert V.A, Gustafsson M.H.G, Laurenzio L. Ontogenetic systematics, molecular developmental genetics // Molecular Systematics of Plants. 2012. P. 349–370.
Borchert R. The concept of juvenility in woody plants // Symposium on Juvenility in Woody Perennials. 1975. V. 56. P. 21–36.
Donovan L.A., Ehleringer J.R. Ecophysiological differences among juvenile and reproductive plants of several woody species // Oecologia. 1991. V. 86. № 4. P. 594–597.
Hackett W.P. Juvenility, maturation and rejuvenation in woody plants // Horticultural Reviews. 1985. P. 109–569.
Hara M., Kimura M., Hirabuki Y., Kanno H. Temporal and spatial patterns of seed production of four shrub species in a climax beech forest // Japanese J. Ecology. 1999. № 49. P. 91–104.
Hufford L. Seed morphology of Hydrangeaceae and its phylogenetic implication // Intern. J. Plant Sci. 1995. V. 156. № 4. P. 555–580.
Hufford L. Ontogeny and morphology of the fertile flowers of Hydrangea and allied genera of tribe Hydrangeeae (Hydrangeaceae) // Botanical J. Linnean Society. 2001. V. 137. № 2. P. 139–187.
Kanno H., Seiwa K. Sexual vs. vegetative reproduction in relation to forest dynamics in the understorey srub, Hydrangea paniculata (Saxifragaceae) // Plant Ecology. 2004. V. 170. № 1. P. 43–53.
Morozowska M., Woznicka A., Kujawa M. Microstructure of fruits and seeds of selected species of Hydrangeaceae (Cornales) and systematic importance // Acta Scientiarum Polonarum. Hortum Cultus. 2012. V. 11. № 4. P. 17–38.
Paiva E.A.S., Machado S.R. Colleters in Caryocar brasiliense (Caryocaraceae) ontogenesis, ultrastructure and secretion // Brazilian J. Biology. 2006. V. 66. № 1b.
Popov A.S., Popova E.V., Nikishina T.V. The development of juvenile plants of the hybrid orchid Bratonia after seed cryopreservation // Cryoletters. 2004. V. 25. № 3. P. 205–212.
Roels P., Decraene L.P.R., Smets E.F. A floral ontogenetic investigation of the Hydrangeaceae // Nordic J. Botany. 1997. V. 17. № 3. P. 235–254.
Van Gelderen C.J., van Gelderen D.M. Encyclopedia of Hydrangeas. Timber Press, Portland – Cambridge, 2004. 279 p.
Wang J.-W., Park M.Y., Wang L.-J., Koo Y., Chen X.-Y., Detlef W. MiRNA control of vegetative phase change in trees // PLoS Genetics. 2011. V. 7. № 2.
Дополнительные материалы отсутствуют.