

Онтогенез, 2021, T. 52, № 4, стр. 287-294
Проявление биоморфологических и анатомических особенностей в ходе онтогенеза мышехвостника маленького Myosurus minimus L. (Ranunculaceae)
М. В. Марков *
Московский педагогический государственный университет
119991 Москва, ул. Малая Пироговская, д. 1, стр. 1, Россия
* E-mail: markovsmail@gmail.com
Поступила в редакцию 27.02.2021
После доработки 22.03.2021
Принята к публикации 27.03.2021
Аннотация
Показано своеобразие развития зародыша семени, морфологической и анатомической структуры побега и корневой системы особи у одного из самых мелких по размерам однолетних растений мышехвостника маленького на самых ранних и следующих за ними этапах онтогенеза. Обсуждается соответствие выявленной структуры (наличие у проростка коллета с ризоидами, аэренхимы в первичной коре гипокотиля и придаточных корней, вторичной гоморизии, плоской апикальной меристемы при облигатной розеточности) и поливариантности онтогенеза (наличие эфемерного варианта и варианта с зимовкой), продолжительности фаз жизненного цикла экологическому своеобразию вида: гигрофильности, минимальному размеру (минимальному числу метамеров на оси главного побега).
ВВЕДЕНИЕ
Объект нашего исследования – мышехвостник маленький Myosurus minimus L. в силу своего очевидного биоморфологического своеобразия не был обделен вниманием в ботанической литературе, хотя детального и тщательного изучения его онтогенеза, судя по всему, до сих пор не предпринималось. В тех же публикациях, которые затрагивали этот вид (Мусина и др., 1993; Нухимовский, 2020), не раз приводились либо неполные, либо ошибочные сведения. С учетом этого обстоятельства была подготовлена настоящая статья, претендующая на восполнение пробела в знаниях относительно названного объекта.
При этом, исходя из того, что доэмбриональному периоду и ранним стадиям эмбрионального периода онтогенеза мышехвостника в литературе было уделено больше внимания (Swingle, 1908; Buvat, 1989), в настоящей статье мы остановимся подробнее на более поздних стадиях эмбриогенеза и онтогенеза.
В обширном и многоликом семействе лютиковых Ranunculaceae, где представлено немало декоративных, ярких растений, есть удивительно мелкие и невзрачные виды. Иногда, как например, в названии мышехвостник маленький (Myosurus minimus L.), мелкие размеры подчеркиваются автором вида (в данном случае Карлом Линнеем) даже в видовом эпитете. В определителе П.Ф. Маевского (2014) для мышехвостника приведенные К.В. Киселевой приблизительные ориентировочные размеры (2–13 см), подчеркивают реальную мелкость, малую так называемую высоту (на деле длину главного побега) генеративных особей этого вида. Обработавший лютиковые для руководства “Сорные растения СССР” Н.В. Шипчинский (1934) приводит еще более узкую амплитуду изменчивости этого показателя – 2–8 см. Даже среди однолетних растений виды столь мелких размеров редки, и потому популяции таких видов заслуживают внимательного изучения в разных аспектах (Марков, 2020). Своих конечных размеров особи достигают в ходе онтогенеза, для которого очень характерна поливариантность, проявляющаяся в темпах роста, числе и структуре метамеров, образованных на разных этапах и зависящая от предпосылок, складывающихся в различных местообитаниях.
Малый размер особей, как морфологический признак, фигурирующий в описаниях таксонов, все равно очень приблизителен и может быть уточнен только при работе с реальными природными или модельными популяциями видов. Безусловно, даже более значимыми представляются данные по амплитуде изменчивости не только высоты, но, даже в большей степени, воздушно-сухой массы особи, к примеру, данные о пороговом размере (массе), достаточном для ее перехода в репродуктивное состояние, но таких данных нет ни в одном определителе. Не приводятся они, зачастую, даже в статьях по биологии видов, например, из серии “Биологическая флора Московской области”, хотя подобная биологическая информация была бы явно очень полезной. Ведь размеры (биомасса) особи у однолетних растений напрямую определяет мощность их репродукции, то есть величину семенной продукции. Цель настоящей работы – показать ряд особенностей биоморфологии, анатомии и популяционной биологии мышехвостника, проявляющихся на разных, и, в том числе, самых ранних, стадиях онтогенеза.
МАТЕРИАЛЫ И МЕТОДЫ
Материал (гербарий и фиксации) был собран в 4-х природных популяциях мышехвостника в Московской области. Особи, высушенные до воздушно-сухого состояния, были обработаны и взвешены на торсионных весах, а анатомию предварительно зафиксированных растений изучали на сделанных от руки или при помощи микротома срезах, обработанных флороглюцином и HCl, обеспечив возможность детального рассмотрения всех одревесневших структур, включая стенки сосудов. Семена проращивали на фильтровальной бумаге в чашках Петри в лаборатории. Появление проростков и формирование розеток и генеративных структур отслеживали как у особей, пересаженных в лабораторию, так и в природных популяциях. Все фотографии сделаны цифровой камерой Nikon Coolpix L22 с применением микроскопа и бинокулярной лупы или без дополнительного увеличения.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Протяженность многоорешка как продукта одного единственного цветка и его акропетальное нарастание как спороносного побега (стробила) приводит к тому, что апокарпные орешки разных уровней и семязачатки внутри них развиваются несинхронно и потому оказываются в момент наблюдения на разных стадиях формирования женского гаметофита, а позднее – зародыша. На рис. 1а, где приводится фото продольного среза многоорешка мышехвостника, и сопроводительных рисунках видно, что в верхней части он не закрыт, здесь продолжается рост – семязачатки, расположенные в самых верхних орешках, содержат еще лишь оформленные археспориальные клетки, тогда как семязачатки нижних орешков уже содержат зрелые зародышевые мешки. Процесс двойного оплодотворения и самых ранних стадий эмбриогенеза хорошо документирован в работе L.D. Swingle (1908), а более поздние стадии эмбриогенеза охарактеризованы и профессионально проиллюстрированы R. Buvat (1989).
Рис. 1.
(а) – 0 – продольный микротомный срез многоорешка, справа (в соответствии с их размещением на цветоложе) показаны стадии развития зародышевых мешков в семязачатках, расположенных на разных уровнях многоорешка: 1 – оформленная археспориальная клетка; 2 – восьмиядерная стадия, 3 – зрелый зародышевый мешок, с яйцевым и антиподальным аппаратами и ядром центральной клетки, готовый к процессу оплодотворения. (б) – 1 – поперечный срез многоорешка; 2 – поперечный срез семени; 3 – продольный срез семени; 4 – зародыш в полости эндосперма.
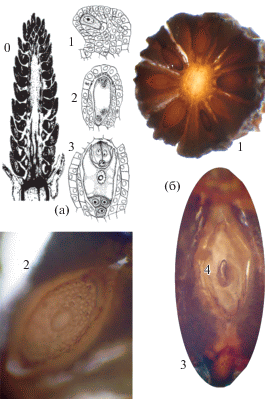
Формирующиеся орешки односеменные и содержат внутри очень мелкие семена. Отмечая, что мышехвостнику в целом присущ эмбриогенез Onagrad (=Cruciferae)-типа (Johansen, 1950), Э.С. Терехин (1996) уточняет, что базальная клетка, образованная в результате первого деления зиготы, не участвует или почти не участвует в построении тела зародыша. Краткую информацию о внутреннем строении семян находим в статье И.А. Ивановой (1966), сообщающей, что в каждом из них имеется обильный твердый эндосперм и маленький зародыш, лежащий на микропилярном конце семени и окруженный полуразрушенными лизированными клетками эндосперма. На приведенной в статье иллюстрации фигурирует лишь крайне схематичный рисунок орешка и еще более схематичное и мелкое изображение семени. Вместе с тем говорится, что, несмотря на малую абсолютную величину зародыша (0.2201 мм), он, составляя лишь 27.39% от длины эндосперма, довольно хорошо развит – имеет семядоли, центральный цилиндр в осевой части зародыша и корневой чехлик. Впрочем, перечисленные детали на представленном в статье рисунке не показаны. На рис. 1б приведены поперечный срез зрелого многоорешка, а также поперечный и продольный срезы семени. Можно видеть, насколько мелок зародыш, размещенный в полости богатого липидами эндосперма, и констатировать сложность, если не невозможность различить какие-либо детали строения зародыша. Судя по нашим данным, такая картина и такие соотношения размеров наблюдаются относительно недолго, потому что при наличии подходящих условий (доступе влаги) рост зародыша, потребляющего питательные вещества эндосперма внутри семени продолжается. После созревания многоорешка, если он целиком, падая на почву, не попадает в условия пригодные для прорастания, когда может реализоваться синаптокарпия – синхронное прорастание семян всего многоорешка (рис. 2), он быстро высыхает, так что продолжение взаимодействия эндосперм–зародыш и транзит от первого ко второму питательных веществ, способствующих доразвитию зародыша, восстанавливается, по-видимому, только после набухания семени и прекращается непосредственно перед его прорастанием.
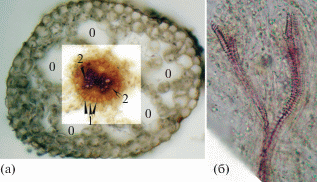
Рис. 2.
Синаптокарпия – синхронное прорастание семян из плодиков многоорешка, упавшего на влажную почву.

Отсюда понятно, почему к моменту прорастания, зародыш уже заполняет весь объем внутри семенной кожуры, а остатков эндосперма, которого к концу достижения апокарпными орешками их конечных размеров по объему было первоначально существенно больше (рис. 1б), обнаружить уже не удается.
Прорастание же семени мышехвостника начинается, как и у большинства двудольных, с появления из семенной кожуры первичного корешка (radicle) и гипокотиля с коллетом (вздутием в области контакта гипокотиля с главным корнем (Compton, 1912; Юсуфова, Марков, 2014), покрытым ризоидами (рис. 3). В этот момент можно выдавить из кожуры увеличившийся в размерах зародыш и рассмотреть его структуру достаточно детально, что и было предпринято нами (см. рис. 4). Хорошо оформленные семядоли, между которыми заметна уплощенная плюмула, первичный корешок с корневым чехликом и явственно выраженный гипокотиль, в котором просвечивают ткани центрального цилиндра, – все это полностью, но уже в другом масштабе, соответствует выше приведенному, но, к сожалению, не иллюстрированному ею в деталях, описанию, данному И.А. Ивановой.
Рис. 3.
Начальные этапы прорастания семени с появлением зародышевого корешка (1), гипокотиля (2) и коллета с ризоидами (3); (4) – семядоли пока в контакте с семенной кожурой.

Рис. 4.
Извлеченный из кожуры семени зародыш с семядолями (1), гипокотилем с просвечивающим центральным цилиндром (2) и корневым чехликом (3) (на давленном препарате).

После прорастания, происходящего по надземному типу, уже у двусемядольного проростка заметно замедление и прекращение роста главного корня, но в зоне коллета очень рано закладывается первый придаточный корень (рис. 5а). На просвет в гипокотиле в результате происходящей внутренней дифференцировки становятся, благодаря прокрашиванию, видны проводящие ткани центрального цилиндра вплоть до деталей строения кольчатых и спиральных сосудов первичной ксилемы (рис. 6б), а также проводящие пучки ответвляющиеся от центрального цилиндра – по достижении семядольного узла – центральный цилиндр разделяется на два пучка, ответвляющихся в семядоли. На поперечных срезах гипокотиля проростка на этой достаточно ранней стадии можно видеть анатомическое строение, близкое к первичному диархному строению корня (рис. 6а). В аэренхиме первичной коры фигурируют крупные полости, а в хорошо выраженной эндодерме достаточно отчетливо видны пояски Каспари. Пока еще ко внутри от эндодермы видны два фрагмента вторичной ксилемы, что свидетельствует об уже произошедшем переходе ко вторичному утолщению.
Рис. 5.
Начальные стадии формирования придаточных корней в нижней части гипокотиля проростка и ювенильных особей: зародышевый корешок (1); гипокотиль (2), семядоли (3) придаточные корни (4), настоящие листья розетки (5).

Рис. 6.
(а) – слева – поперечный срез гипокотиля проростка с аэренхимой в коре (0), эндодермой с поясками Каспари (1) и двумя фрагментами вторичной ксилемы (2). (б) – снятые на просвет кольчато-спиральные сосуды ксилемы центрального цилиндра, дихотомирующего в семядоли.
Выше упомянутый первый гипокотильный придаточный корень выступает как предвестник заложения серии придаточных корней, формирующих первую, особо важную для особей любых размеров, нижнюю гоморизную зону (рис. 7а). И центральный цилиндр гипокотиля и его ответвления переходят ко вторичному строению, а их вторичные одревесневающие проводящие ткани хорошо окрашиваются, что позволяет отчетливо увидеть кору как самого гипокотиля, так и кору его ответвлений. Вторичное утолщение с образованием крупного массива ксилемы продолжается и в гипокотиле, благодаря чему он сохраняется у особи на всю ее жизнь, и в придаточных корнях нижней зоны, каждый из которых имеет собственную кору с аэренхимой. Некоторые из них, отходя от центрального цилиндра, некоторое время гравитропно растут вертикально вниз под корой, прижимаясь к центральному цилиндру гипокотиля, что бывает хорошо видно на поперечных срезах (рис. 8). Наличие коры с хорошо выраженной аэренхимой у гипокотиля и его ветвей в нижней, гоморизной зоне представляется крайне интересной особенностью онтогенеза мышехвостника, которую вместе с наличием ризоидов на коллете, способствующих гравитропизму проростка и обеспечению его влагой еще до формирования настоящих корневых волосков (Parsons (2009, 2013), логично связать с гигрофильностью, свойственной этому виду и способностью произрастать в местообитаниях не только с достаточным, но, порой, и с избыточным увлажнением. Посевы культурных растений, подвергшиеся вымоканию, – это пример подобных местообитаний, в которых популяции мышехвостника находят оптимальные условия и могут иметь наиболее высокую плотность (рис. 9).
Рис. 7.
(a) – снятый на просвет гипокотиль (3) с нижней и верхней гоморизными зонами ювенильного растения мышехвостника: зародышевый корешок (1), зачаток придаточного корня и сформированные придаточные корни нижней гоморизной зоны (2); зачаток (5) и сформированный придаточный корень (4) верхней гоморизной зоны; просвечивающий центральный цилиндр гипокотиля (6) с ответвлениями в придаточные корни нижней гоморизной зоны; (б) – просвечивающий центральный цилиндр гипокотиля (1); зачатки придаточных корней верхней гоморизной зоны с чехликами (2) и проводящие пучки, ответвляющиеся в розеточные листья от центрального цилиндра (3). Масштабная линейка 3 мм.

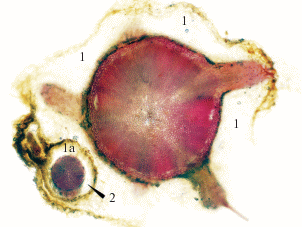
Рис. 8.
Поперечный срез нижней части гипокотиля взрослой генеративной особи с корой (1), придаточными корнями и отходящими от центрального цилиндра проводящими пучками, следующими в эти корни. Поперечный срез одного из придаточных корней (2) с его собственной корой (1а), гравитропно направлявшегося вниз под корой гипокотиля.

Вторая (верхняя) гоморизная зона формируется только у достаточно мощных особей в верхней части гипокотиля вблизи розетки, причем не всегда придаточные корни отходят здесь от узлов главного побега. На рис. 7 показана полностью развитая нижняя зона придаточных корней, включая их хорошо оформленные зачатки, а также верхняя зона придаточных корней вблизи розетки с уже сформированным придаточным корнем и также хорошим оформлением зачатков, имеющих даже сформированные внутри гипокотиля корневые чехлики (рис. 7б). Между этими двумя (нижней и верхней) зонами находится зона бескорневого гипокотиля – вероятность заложения здесь придаточных корней крайне низкая, что позволяет рассматривать ее по аналогии с известной зоной торможения побега, где почки не активны.
Любопытной особенностью мышехвостника, как облигатно розеточного растения, представляется наличие увенчивающего гипокотиль плоского или иногда даже несколько вогнутого апекса, который, подобно плоскому апексу подорожника большого (Plantago major), также никогда не теряющего своей розеточности, формирует в норме большую и плотную розетку листьев. Располагающиеся по периферии вокруг медуллярной центральной части плоской апикальной меристемы (т.н. меристемы ожидания – Buvat, 1989) листовые примордии в составе так называемого инициального кольца (Buvat, 1989), превращаются позднее в узкие листья с одной (у ювенильных особей) или тремя (у генеративных особей) жилками. Основания розеточных листьев остаются после отпадения пластинок, охватывая ось укороченного розеточного побега (рис. 10а). После удаления развитых розеточных листьев становится хорошо заметно, что диаметр розеточной части главной оси заметно больше, чем диаметр цветоноса, т.е. репродуктивной части главной оси (рис. 10б). Для анатомии поперечного среза цветоножки характерно наличие склеренхимы перицикла, расположенных по кругу коллатеральных проводящих пучков и полой сердцевины (рис. 11а). В отличие от апикальной меристемы подорожника, обеспечивающей моноподиальное нарастание главной оси при формировании только пазушных репродуктивных побегов, меристема мышехвостника изменяет свою форму при переходе особи из вегетативного в генеративное состояние и формирует верхушечный цветок (рис. 10в). Размеры этого цветка (по длине образующегося в нем многоорешка) у нормально развитых особей мало отличаются от цветков, впоследствии формирующихся на паракладиях в пазухах нижерасположенных розеточных листьев. Из сказанного ясно, что при отсутствии выраженной гетерофиллии у мышехвостника различать онтогенетические (~возрастные) состояния удается только по анатомии листьев (числу жилок), а по форме листьев различия между ними очень слабо выражены и потому плохо заметны.
Рис. 10.
(а) – укороченная и разросшаяся по диаметру ось розетки, покрытая основаниями розеточных листьев; (б) – цветонос первого цветка, завершающего рост главной оси розетки (1) инициальное кольцо листовых примордиев плоской апикальной меристемы, сформировавшей до этого розетку (2), (в) – первый цветок, увенчивающий главную ось розетки.

Рис. 11.
(а) – поперечный срез цветоноса первого цветка; (б) – цветущая особь минимального размера с живыми семядолями (1), гипокотилем (2) и двумя первыми настоящими розеточными листьями (3).

Итак, гипокотиль мышехвостника вместе с придаточными корнями, благодаря переходу ко вторичному росту, сохраняется в течение всей жизни особи, и в этом наиболее существенная особенность, отличающая мышехвостник от большинства травянистых двудольных, гипокотиль у которых очень быстро сдавливается, съеживается и потому становится у взрослой особи незаметным. По понятным причинам такую судьбу гипокотиля разделяет и еще более эфемерный атрибут гипокотиля коллет.
Поэтому весьма странным выглядит утверждение авторов атласа по анатомии растений (Schweingruber et al., 2011: 7), что для анатомических исследований и определения возраста наиболее важной частью растения является переходная зона между корнем и стеблем (которую авторы называют “collar”). Но термины “collar” и “collet” (коллар и коллет) – это синонимы для обозначения соответствующей морфологической структуры, которой присуща определенная важная функция, а не просто зоны между двумя другими структурами (Compton, 1912; Parsons, 2009; Юсуфова, Марков, 2014).
Вопреки неоднократно опубликованному мнению, что мышехвостник – это весенний эфемер, следует уточнить, что в условиях нашего умеренного климата, демонстрируя поливариантность онтогенеза, он ведет себя, не только как весенний эфемер, но и как зимующий или озимый однолетник. Во втором случае прорастание семян и формирование розеток у него наблюдается в конце лета или осенью (рис. 12), и от того, в каком состоянии особи такой осенней когорты уходят в зиму, зависит их реализуемая весной репродуктивная мощность, поскольку вегетативный рост, начавшийся осенью, весной не продолжается. Так в мае 2015 г. из-за позднего прорастания осенью предыдущего года, особи мышехвостника были очень мелкими и на стадии репродукции имели размеры близкие к минимальным (рис. 11б). Но и такие особи способны формировать 5–7 вполне жизнеспособных семян, что было доказано в наших опытах по проращиванию.

Как показано выше, розетку мышехвостника (облигатно розеточного однолетника) формирует плоский апекс с окружающим его инициальным кольцом листовых примордиев (рис. 10б). Минимальное число фитомеров взрослого типа в составе розетки, достаточное для перехода к репродукции, – всего 2 (рис. 11б). Редко у кого из растений, даже однолетних, можно наблюдать такое малое число фитомеров, образуемых в ходе онтогенеза на главной оси, а воздушно-сухая масса минимальных по размеру особей может составлять иногда менее 1 мг! У крупных особей на основе плотной главной розетки из большого числа листьев и мелких розеток из двух листьев у каждого из паракладиев (рис. 13) формируется высокая семенная продуктивность – до 1500 орешков на особь.
Рис. 13.
Мощная генеративная особь мышехвостника с гипокотилем и придаточными корнями. Внизу – паракладий – боковой побег с кроющим листом, из пазухи которого он появился (1), двумя розеточными листьями (2) и многоорешком (3).

Ранее нами уже было показано, что в популяциях именно самые мелкие одноцветковые особи в сумме формируют число орешков превышающее то, которое образуют особи других размерных классов (Марков, 1992, 2020). При этом для особей всех размерных классов характерно весьма высокое и сходное репродуктивное усилие (до 80%!), а коэффициенты в популяционно-аллометрических уравнениях близки к единице – указывая на изометрию – сохранение сходных пропорций у особей разной мощности.
ВЫВОДЫ
1. К моменту их прорастания в семенах мышехвостника нет эндосперма и присутствует только занимающий весь объем краекорешковый зародыш с первичным корешком (radicle), который первым трогается в рост при прорастании, семядолями и уплощенной плюмулой между ними.
Вопреки имеющимся в литературе сведениям о преобладающем в семени по объему эндосперме и крохотном зародыше установить, когда зародыш сумел настолько вырасти, что присутствие эндосперма в семени к моменту прорастания практически незаметно, довольно трудно.
2. Мышехвостник маленький Myosurus minimus L., не имея системы стержневого корня, который отмирает на самых ранних стадиях (стадии первичного корешка) – вскоре после прорастания семени, формирует в ходе онтогенеза систему придаточных корней, которая начинает формироваться уже на стадии 5-дневного проростка с образования первого придаточного корня на гипокотиле; продолжение формирования этой системы идет по четко выраженному плану: после формирования серии придаточных корней вблизи нижнего конца гипокотиля центр формирования новой серии многочисленных придаточных корней смещается вверх сразу к нижней части розетки, так что между названными двумя очагами формирования придаточных корней остается лишенная корней средняя и наиболее протяженная зона – некая “зона торможения”.
3. Гипокотильные придаточные корни ответвляются от центрального тяжа проводящих тканей вместе с имеющейся у них корой и сразу приобретают характерную для корней гравитропическую направленность роста.
4. Гипокотиль мышехвостника претерпевает вторичное утолщение и потому сохраняется у особи на всю ее жизнь. Проводящая система гипокотиля начинает формироваться очень рано и при условии окрашивания ее одревесневших элементов флороглюцином видна уже у зародыша, выдавленного из кожуры. Чуть позднее можно рассмотреть кольчатые и спиральные сосуды, тяжи которых дихотомически расходятся к семядолям. В коре гипокотиля есть хорошо оформленная аэренхима, что объясняет приуроченность растения к местообитаниям не только с достаточно, но и избыточно высокой влажностью почвы.
5. Дополнительной деталью, свидетельствующей о гигрофильности мышехвостника можно считать коллет, покрытый ризоидами.
6. Для мышехвостника маленького характерна поливариантность онтогенеза и наличие, помимо весеннего эфемерного варианта, варианта зимующего однолетника с перезимовкой розеток и обязательным немедленным переходом к репродукции сразу после зимовки.
Список литературы
Иванова И.А. О внутреннем строении семян лютиковых // Бюл. ГБС. 1966. Вып. 61. С. 72–79.
Маевский П.Ф. Флора средней полосы европейской части России. 11-е изд. М.: Т-во науч. изд. КМК. 2014. 635 с.
Марков М.В. Роль многоорешков в обеспечении пластичности репродуктивной сферы у Ceratocephala falсata (L.) Pers и Myosurus minimus L. // Бюл. МОИП, отд. Биология. 1992. Т. 97. Вып. 1. С. 51–67.
Марков М.В. Популяционная биология растений. М.: КМК, 2012. 387 с.
Марков М.В. “Изюминка” популяционной биологии малолетних растений или почему размеры растений также важны // Журн. Общей Биологии. 2020, Т. 81. № 6. С. 458–468.
Мусина Г.В., Барыкина Р.П., Чубатова Н.В. Myosurus minimus L. // Биологическая флора Московской области. 1993. Вып. 9(1). С. 65–73.
Нухимовский Е.Л. Основы биоморфологии семенных растений. Том 3: Теория интегральной соматической эволюции. М.: Недра. 2020. 676 с.
Терехин Э.С. Семя и семенное размножение. Мир и семья-95. СПб., 1996. 376 с.
Шипчинский Н.В. Myosurus L. Мышехвостник // Сорные растения СССР. Л.: АН СССР, 1934. Т. 2. С. 228–229.
Юсуфова В.З., Марков М.В. К анатомии и морфологии коллета как особой структуры у проростков некоторых видов растений// Труды IX международной конференции по экологической морфологии растений, посвященной памяти И.Г. и Т.И. Серебряковых. (К 100-летию со дня рождения И.Г. Серебрякова). М., 2014. Т. 2. С. 306–309.
Compton R.H. Theories of anatomical transition from root to stem // New Phytol. 1912. V. 1. P. 13–25.
Johansen D.A. Plant Embryology. N.Y., The Chronica Botanica Company. 1950. 305 p.
Parsons R.F. Hypocotyl hairs: an historical perspective // Aust. J. Bot. 2009. 57:106–108.
Parsons R.F. Adaptations to floodplains in Populus and Salix: the role of collet hairs // Trees. 2013. V. 27. P. 1–5.
Salisbury E.J. The pioneer vegetation of exposed muds and its biological features // Philosophical Transactions of the Royal Society of London. 1970. V. 259. P. 207–255. https://doi.org/10.1098/rstb.1970.0059
Schweingruber F.H., Börner A., Schulze E.-D. Atlas of Stem Anatomy in Herbs, Shrubs and Trees. Springer. 2011. V. 1. 490 p.
Swingle L.D. Embryology of Myosurus minimus // The American Naturalist. 1908. V. 42. № 501. P. 582–591.
Дополнительные материалы отсутствуют.