

Онтогенез, 2022, T. 53, № 1, стр. 31-45
Изменения окраски редкого глубоководного ската Ричардсона Bathyraja richardsoni (Arhynchobatidae, Rajiformes, Chondrichthyes) в онтогенезе
А. М. Орлов a, b, c, d, e, f, *, Н. И. Рабазанов d, f, А. И. Никифоров g
a Институт океанологии им. П.П. Ширшова РАН (ИО РАН)
117218 Москва, Россия
b Всероссийский научно-исследовательский институт рыбного хозяйства и океанографии (ВНИРО)
107140 Москва, Россия
c Институт проблем экологии и эволюции им. А.Н. Северцова РАН (ИПЭЭ РАН)
119071 Москва, Россия
d Дагестанский государственный университет (ДГУ)
367000 Махачкала, Россия
e Томский государственный университет (ТГУ)
634050 Томск, Россия
f Прикаспийский институт биологических ресурсов ДФИЦ РАН (ПИБР ДФИЦ РАН)
367000 Махачкала, Россия
g Московский государственный институт международных отношений (университет) МИД Российской Федерации – МГИМО
Москва, Россия
* E-mail: orlov@vniro.ru
Поступила в редакцию 20.05.2021
После доработки 15.09.2021
Принята к публикации 20.09.2021
- EDN: MATTEW
- DOI: 10.31857/S0475145022010050
Аннотация
Исследованы особенности окраски дорсальной и вентральной сторон редкого глубоководного ската Ричардсона Bathyraja richardsoni (Arhynchobatidae) в диапазоне длин от 14 до 146 см и прослежены ее онтогенетические изменения. Представлены новые данные по окраске эмбрионов, неполовозрелых, созревающих и половозрелых особей. Показано, что вариации окраски дорсальной стороны незначительны, в то время как окраска вентральной стороны может значительно варьировать. Полученные данные могут быть использованы в таксономических и популяционных исследованиях, при разработке ключей для определения видов, подготовке фаунистических сводок и определителей, облегчить понимание особенностей окраски и ее изменений у глубоководных животных, обитающих в условиях постоянной темноты, а также расширить знания об отдельных аспектах макро- и микроэволюции глубоководных скатов.
ВВЕДЕНИЕ
Фауна глубоководных скатов даже наиболее изученной северной части Атлантического океана исследована недостаточно хорошо. Многие северо-атлантические виды описаны относительно недавно (во второй половине прошлого столетия) и известны до сих пор по небольшому числу находок (Orlov et al., 2006; Porteiro et al., 2017). К таким видам относится и скат Ричардсона Bathyraja richardsoni, впервые описанный из вод Новой Зеландии (Garrick, 1961; Garrick, Paul, 1974). Ареал данного вида расположен в северной и центральной Атлантике (от северо-восточной Канады и юго-западной Гренландии до Азорских о-вов и Мавритании) и в юго-западной Пацифике в водах Тасмании и Новой Зеландии (Ebert, Stehmann, 2013; Kulka et al., 2015; Porteiro et al., 2017), где он известен по разрозненным поимкам. Скат Ричардсона – один из наиболее крупных видов глубоководных скатов, достигающий длины 175 см (Sulak et al., 2009; Ebert, Stehmann, 2013). Наибольшей численности он достигает в районе Срединно-Атлантического хребта (САХ) (Fossen et al., 2008) и является одним из наиболее глубоководных северо-атлантических скатов, поимки которого известны с глубин 501–3055 м с диапазоном максимальной встречаемости между 2100 и 2400 м (Orlov et al., 2006; Porteiro et al., 2017).
Скаты являются одной из наиболее уязвимых групп морских рыб по отношению к антропогенному воздействию (Dulvy, Reinolds, 2002), и отдельные страны и природоохранные организации уделяют повышенное внимание вопросам охраны и сохранения как хрящевых рыб в целом, так и скатов в частности (Орлов, 2005, 2008; Cavanagh, Gibson, 2007; Орлов, Шеверницкий, 2008; Орлов, Литвинов, 2010; Goodwin, 2012). По версии Международного союза охраны природы (МСОП) скат Ричардсона отнесен к категории “Least Concern” (Kulka et al., 2015), т.е. в настоящее время угрозы антропогенного характера для него отсутствуют.
Несмотря на то, что данный вид является довольно многочисленным в сравнении с другими глубоководными скатами северной Атлантики, до сих пор его биология остается изученной недостаточно (Tempelman, 1973b; Orlov et al., 2006), а в большинстве опубликованных работ описываются преимущественно особенности морфологии данного вида (Garrick, 1961; Forster, 1965; Tempelman, 1973a, 1973b; Garrick, Paul, 1974; Stehmann, Merrett, 2001; Orlov et al., 2006, 2010; Orlov, 2011; Orlov, Cotton, 2011). Между тем, опубликованные сведения об окраске ската Ричардсона разрознены и ограниченны. Штеманн и Мерретт (Stehmann, Merrett, 2001) описали окраску пяти извлеченных из яйцевых капсул эмбрионов. Ранее сведения об окраске нескольких крупных половозрелых скатов Ричардсона были описаны для экземпляров из вод Новой Зеландии (Garrick, 1961), северо-восточной (СВА) (Forster, 1965) и северо-западной (СЗА) Атлантики (Tempelman, 1973b), позднее обобщенные в региональных фаунистических сводках (Sulak et al., 2009; Ebert, Stehmann, 2013). В целом, в указанных работах для окраски взрослых особей указывались такие общие признаки: дорсальная сторона тела однотонная, от пепельной до коричневой, с небольшими светлыми точками или пятнами и темной каймой на крыльях и нижних лопастях брюшных плавников (V); вентральная сторона тела заметно темнее дорсальной, с темной каймой на крыльях и нижних лопастях V и наличием белых участков вокруг рта, ноздрей и клоаки, на хвосте и у его основания, между грудными (P) и V, на нижних лопастях V и вершинах птеригоподиев.
Отсутствие опубликованных данных по окраске среднеразмерных скатов Ричардсона не позволяло исследовать закономерности изменений его окраски на протяжении всего жизненного цикла и выявить ее внутривидовую и онтогенетическую изменчивость. Подобные сведения имеют важное значение для таксономических и популяционных исследований, а также при разработке ключей для определения видов и подготовке фаунистических сводок и определителей. Кроме того, разработка данной проблемы может представлять более широкий фундаментальный интерес в плане осмысления особенностей окраски и ее изменений у глубоководных животных, обитающих в условиях постоянной темноты, а также способствовать лучшему пониманию отдельных аспектов макро- и микроэволюции (происхождение, расселение, формирование популяционных структур и др.) глубоководных скатов.
Цель работы – анализ всех доступных (собственных и литературных) данных по окраске ската Ричардсона, позволяющих проследить ее онтогенетические изменения в пределах всего видового ареала.
МАТЕРИАЛЫ И МЕТОДЫ
Материалом для анализа вариаций окраски ската Ричардсона послужили 20 экземпляров обоих полов общей длиной от 14 до 146 см, исследованных первым автором в течение 2005–2010 гг. в различных музеях (табл. 1):
Таблица 1.
Окраска экземпляров ската Ричардсона Bathyraja richardsoni различной длины (1 – BMNH 1999.2.2.4; 2 – BMNH 1999.2.2.1; 3 – BMNH 1999.2.2.3; 4 – BMNH 1999.2.2.2; 5 – ZMUB 17600; 6 – MNHN 1999-1156; 7 – ZMUB 19522-3; 8 – BMNH 1999.10.1.1; 9 – ZMUB 19522-2; 10 – ZMUB 19535; 11 – ZMUB 19522-1; 12 – экземпляр из вод Азорских о-вов, 13 – ZMUB 19514-4; 14 – BMNH 1999.10.1.2; 15 – ZMUB 19514-1; 16 – ZMUB 19514-3; 17 – ZMUB 19514-2; 18 – ZMUB 19364; 19 – ZMUB 19448; 20 – экземпляр из северо-западной Атлантики (Tempelman, 1973a); 21 – ZMUB 19476; 22 – экземпляр из вод Канарских о-вов (Brito et al., 1998); 23 – экземпляр из северо-восточной Атлантики (Forster, 1965); 24 – экземпляр из вод Азорских о-вов; 25 – экземпляр из северо-западной Атлантики (Tempelman, 1973a); 26 – экземпляр из вод Новой Зеландии (Garrick, 1961); 27 – ZMUB 19528; 28 – экземпляр из северо-восточной Атлантики (Forster, 1965); № – номер признака по порядку; “+” – признак присутствует, “–” – признак отсутствует, нд – нет данных
| Группа | Эмбрионы | Неполовозрелые | Созревающие | Половозрелые | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Номер экземпляра | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | |
| Общая длина (TL), см/пол | 18 ♀ |
22 ♂ |
24 ♀ |
25 ♂ |
29 ♀ |
35 ♂ |
45 ♀ |
52 ♂ |
53 ♀ |
53 ♂ |
55 ♂ |
60 ♀ |
63 ♀ |
69 ♂ |
69 ♀ |
78 ♂ |
84 ♂ |
85 ♀ |
101 ♀ |
103 ♀ |
106 ♀ |
110 ♀ |
113 ♂ |
127 ♂ |
130 ♂ |
133 ♀ |
142 ♀ |
146 ♀ |
|
| № | Признак (частота встречаемости, %) | Дорсальная сторона тела | |||||||||||||||||||||||||||
| 1 | Общая окраска от пепельной до коричневой (100.0) | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | нд | + | + | + | + |
| 2 | Наличие небольших светлых точек или пятен (96.4) | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | – | + | нд | + | нд | + | + |
| 3 | Наличие темной каймы на крыльях и нижних лопастях V (96.4) | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | – | + | нд | + | + | + | + |
| № | Признак (частота встречаемости, %) | Вентральная сторона тела | |||||||||||||||||||||||||||
| 4 | Общая окраска пепельно-коричневая, темнее дорсальной поверхности, темная кайма на крыльях и нижних лопастях V (100.0) | + | + | + | + | + | + | + | + | + | + | + | нд | + | + | + | + | + | + | + | + | + | нд | + | + | + | нд | + | + |
| 5 | Белый участок вокруг рта (100.0) | + | + | + | + | + | + | + | + | + | + | + | нд | + | + | + | + | + | + | + | + | + | нд | + | + | + | + | + | + |
| 6 | Белый участок вокруг ноздрей (100.0) | + | + | + | + | + | + | + | + | + | + | + | нд | + | + | + | + | + | + | + | + | + | нд | + | + | + | + | + | + |
| 7 | Белый участок у входа в клоаку (92.4) | – | – | + | + | + | + | + | + | + | + | + | нд | + | + | + | + | + | + | + | + | + | нд | + | + | + | + | + | + |
| 8 | Белый участок у основания хвоста (80.8) | + | + | + | + | + | + | + | + | + | + | + | нд | + | + | + | – | – | + | + | + | – | нд | – | – | + | + | + | + |
| 9 | Белый участок между P и V (73.1) | + | + | + | + | – | + | – | + | + | + | – | нд | + | – | + | – | + | + | + | + | + | нд | – | + | + | + | + | – |
| 10 | Белый участок на нижних лопастях V (76.9) | + | + | + | + | + | + | + | + | + | + | + | нд | + | – | – | – | – | + | – | + | + | нд | – | + | + | + | + | + |
| 11 | Белое пятно на брюхе (38.5) | а | а | б | а | + | в | + | – | – | – | – | нд | + | – | г | – | д | – | – | – | – | нд | – | – | – | – | – | – |
| 12 | Белые пятна на хвосте (35.7) | е | + | + | + | + | e | + | + | + | + | – | нд | + | – | нд | – | – | – | – | – | – | нд | – | – | – | – | – | – |
| 13 | Белые участки на вершинах птеригоподиев (80.0) | + | + | + | + | – | + | – | – | + | + | + | + | ||||||||||||||||
а – у эмбрионов сохраняются светло окрашенные участки на месте желточного мешка, дополнительные светлые участки видны на уровне от 1-х жаберных щелей до окончания диска; б – у эмбрионов сохраняются светло окрашенные участки на месте желточного мешка, дополнительные светлые участки стреловидной формы с вершиной у желточного мешка и основанием у нижних лопастей V; в – белое пятно между 5-ми жаберными щелями; г – белое пятно на уровне между 2-й и 5-й жаберными щелями; д – белое пятно неправильной формы на уровне между ртом и 3-й жаберной щелью; е – хвост полностью белый.
Зоологический музей университета г. Берген, Норвегия: ZMUB 19528 (TL 142 см, самка, 2534 м), ZMUB 17600 (TL 29 см, самка, 2534 м), ZMUB 19514-3 (TL 78 см, самец, 1911 м), ZMUB 19514-4 (TL 63 см, самка, 1911 м), ZMUB 19522-3 (TL 45 см, самка, 2364 м), ZMUB 19522-1 (TL 55 см, самец, 2364 м), ZMUB 19522-2 (TL 53 см, самка, 2364 м), ZMUB 19535 (TL 53 см, самец, 2534 м), ZMUB 19514-1 (TL 69 см, самка, 2280 м), ZMUB 19514-2 (TL 84 см, самец, 1911 м), ZMUB 19476 (TL 106 см, самка, 2288 м), ZMUB 19448 (TL 101 см, самка, 2534 м), ZMUB 19364 (TL 85 см, самка, 2951 м) – все из вод САХ;
Национальный музей естественной истории, г. Париж, Франция: MNHN 1999-1156 (TL 35 см, самец, 1975 м, СВА, восточнее Гибридских о-вов);
Британский музей естественной истории, г. Лондон, Великобритания: BMNH 1999.10.1.1 (TL 52 см, самец, 2441 м, глубина и место поимки неизвестны), BMNH 1999.10.1.2 (TL 69 см, самец, 2441 м, глубина и место поимки неизвестны), BMNH 1999.2.2.1 (TL 22 см, самец, 1541 м, СВА восточнее Ирландии), BMNH 1999.2.2.2 (TL 25 см, самец, 1541 м, СВА восточнее Ирландии), BMNH 1999.2.2.3 (TL 24 см, самка, 1541 м, СВА восточнее Ирландии), BMNH 1999.2.2.4 (18 см, самка, 1541 м, СВА восточнее Ирландии).
Кроме того, для сравнения привлечены литературные данные об окраске, фотографии или рисунки экземпляров из вод Новой Зеландии (Garrick, 1961), СВА (Forster, 1965) и СЗА (Tempelman, 1973a), а также Азорских о-вов (фото Гуи Менезес (Gui Menezes)).
Анализ вариаций окраски проводили на основании изучения свежевыловленных особей (САХ), их описаний из опубликованной литературы, рисунков (воды Новой Зеландии) или фотографий (воды Канарских и Азорских о-вов, СВА и СЗА) и фиксированных экземпляров из музейных коллекций (СВА).
Особенности полового созревания ската Ричардсона не изучены, а о размерах, при которых он созревает, ничего не известно. Основываясь на опубликованных сведениях, а также данных о соотношении длины птеригоподиев и нижней лопасти V (Орлов, 2006) у исследованных самцов, можно заключить, что длина эмбрионов составляет 14–25 см, неполовозрелых – 35–69 см, созревающих – 78–85 см и половозрелых – свыше 113 см. При этом длина, при которой созревают самцы и самки крупных северо-тихоокеанских и южно-африканских скатов, практически не различается, а половая зрелость наступает при достижении 60% максимальной длины (Орлов, 2006; Ebert et al., 2007; Орлов, Смирнов, 2015). В связи с этим, в дальнейшем при обсуждении полученных результатов для обозначения степени половой зрелости особей ската Ричардсона в исследованном размерном ряду мы условно выделяем следующие группы: эмбрионы – 18–25 см, неполовозрелые – 29–69 см, созревающие и половозрелые – 78–105 см, половозрелые – выше 105 см (последняя составляет 60% от максимально известной длины).
РЕЗУЛЬТАТЫ
Результаты анализа представлены в табл. 1, включающей 13 признаков – особенностей окраски. В анализ не включен ранний эмбрион длиной 142 мм BMNH 1999.2.2.4 (рис. 1), который был окрашен одноцветно и не имел характерных для вида особенностей окраски как в свежем (Stehmann, Merrett, 2001), так и фиксированном состоянии (наши данные). Описание окраски проиллюстрировано фотографиями скатов, представляющих размерный ряд 14, 18, 25, 35, 52, 69, 84, 106 и 127 см (рис. 1–9).
Рис. 1.
Ранний эмбрион ската Ричардсона Bathyraja richardsoni длиной 14 см (BMNH 1999.2.2.4): а – дорсальная сторона, б – вентральная сторона.

Рис. 2.
Дорсальная сторона крупной самки ската Ричардсона Bathyraja richardsoni длиной 106 см (ZMUB 19476): 1 – грудные плавники (крылья), 2 – нижние лопасти V.

Рис. 3.
Вентральная сторона половозрелого самца ската Ричардсона Bathyraja richardsoni длиной 127 см из вод Азорских о-вов (фото Гуи Менезес): 1 – рот, 2 – ноздри, 3 – крылья (грудные плавники), 4 – нижние лопасти V, 5 – клоака, 6 – птериподии).

Рис. 4.
Эмбрион ската Ричардсона Bathyraja richardsoni длиной 18 см (BMNH 1999.2.2.4): а – дорсальная сторона, б – вентральная сторона: 1 – остатки желточного мешка, 2 – белые участки на нижних лопастях V, 3 – светлая нижняя поверхность хвоста, 4 – белые участки на птеригоподиях.

Рис. 5.
Созревающий самец ската Ричардсона Bathyraja richardsoni длиной 84 см (ZMUB 19514-2): а – дорсальная сторона, б – вентральная сторона.

Рис. 6.
Неполовозрелый самец ската Ричардсона Bathyraja richardsoni длиной 35 см (MNHN 1999-1156): а – дорсальная сторона, б – вентральная сторона.
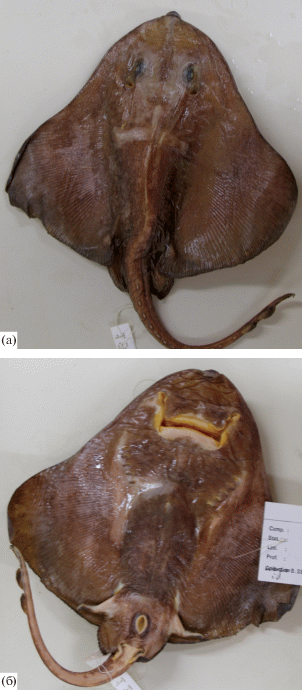
Рис. 7.
Неполовозрелый самец ската Ричардсона Bathyraja richardsoni длиной 69 см (BMNH 1999.10.1.2): а – дорсальная сторона, б – вентральная сторона.

Рис. 8.
Неполовозрелая самка ската Ричардсона Bathyraja richardsoni длиной 53 см (ZMUB 19522-2): а – дорсальная сторона, б – вентральная сторона: 1 – пятно на брюхе, 2 – белый участок у основания хвоста.
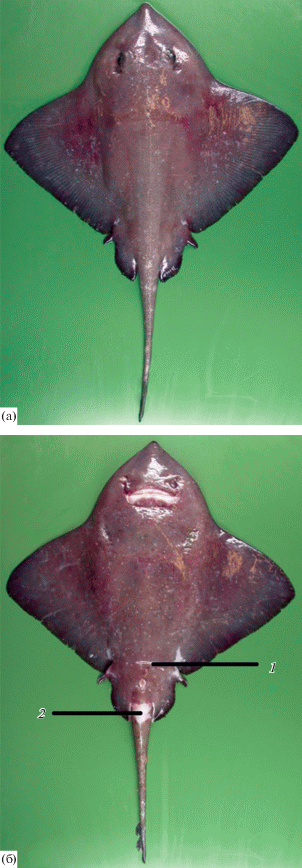
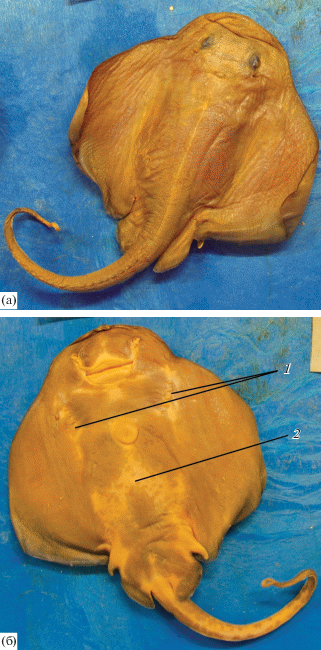
Рис. 9.
Эмбрион ската Ричардсона Bathyraja richardsoni длиной 24 см (BMNH 1999.2.2.3): а – дорсальная сторона, б – вентральная сторона: 1 – белые участки в районе жаберных щелей, 2 – светлый участок стреловидной формы.
Дорсальная сторона тела
Вариации окраски дорсальной поверхности ската Ричардсона в сравнении с таковыми вентральной можно рассматривать как незначительные. Для всех исследованных особей данного вида (без учета экземпляров, у которых по отдельным признакам данные отсутсвовали) независимо от их размера, пола и состояния половой зрелости была характерна однотонная окраска дорсальной стороны от пепельной до коричневой (см. табл. 1). Подавляющее большинство особей на дорсальной поверхности диска имело более темную кайму по его краю и на нижних лопастях V (96.4%), а также небольшие светлые точки или пятна (96.4%) (рис. 2). Исключение в этом отношении представляла самка длиной 110 см (№ 22 в табл. 1) из вод Канарских о-вов (Brito et al., 1998), у которой (судя по фото) оба рассматриваемых признака отсутствовали.
Вентральная сторона тела
Вариации окраски вентральной стороны ската Ричардсона были более разнообразны. У всех исследованных скатов без исключения независимо от длины, пола и половой зрелости вентральная сторона в целом была пепельно-коричневой. При этом она была обычно темнее дорсальной, а на краях крыльев и нижних лопастей V имелась темная кайма. Также все исследованные особи характеризовались наличием белых участков вокруг рта, ноздрей и от ноздрей до углов рта (рис. 3). У подавляющего большинства особей (92.3%) отверстие клоаки было белым (см. рис. 3), за исключением двух эмбрионов длиной 18 и 22 см (№ 1 и 2, в табл. 1 и рис. 4б, 6б). Нижняя сторона хвоста была полностью белой у 2 эмбрионов – самки TL 18 см и самца TL 35 см (№ 1 и 6 соответственно). У прочих особей низ хвоста в разной степени пигментирован (до полностью темного – рис. 3 и 5); причем у некоторых из них у его основания имелся белый участок (рис. 4б, 6). Около трети исследованных экземпляров (35.7%) характеризовались наличием светлых пятен на хвосте (рис. 8б), за исключением уже упоминавшихся выше двух особей, у которых хвост с вентральной стороны был полностью белый.
У 73.1% особей белый участок отмечен в месте соединения P и V плавников (рис. 3, 4б, 6б, 7б, 8б, 9б), за исключением семи самок и самцов длиной от 29 до 146 см (рис. 5б). У несколько большего числа особей (76.9%) белые участки отмечены на нижних лопастях V (рис. 4б, 9б), за исключением шести самцов и самок длиной от 69 до 113 см (рис. 5б, 8б). Чуть больше трети исследованных экземпляров длиной до 69 см имели белые пятна на брюхе (рис. 8б), форма, расположение и размеры которых варьировали. У эмбрионов длиной 18, 22 и 24 см сохранились светло окрашенные участки на месте желточного мешка (рис. 4б, 9б), а также имелись дополнительные светлые участки вокруг жаберных щелей (рис. 9б). У эмбриона длиной 24 см (№ 3) имелись дополнительные светлые участки, образующие фигуру стреловидной формы, с вершиной у места прикрепления желточного мешка и основанием у нижних лопастей V (рис. 9б). У самца длиной 35 см (№ 6) белое пятно отмечено между пятыми жаберными щелями (рис. 6б). У самца длиной 69 см (№ 14) белое пятно находилось на уровне между вторыми и пятыми жаберными щелями (рис. 7б). Кроме того, у 80.0% самцов имелись белые участки на птеригоподиях (рис. 3, 4б, 5б, 6б, 7б), за исключением экземпляров длиной 53, 69 и 78 см (№ 10, 14, 16).
Особенности окраски различных размерных групп
В целом, независимо от размера и пола (за исключением эмбриона TL 14 см) все исследованные особи имели однотонную от пепельной до коричневой окраску дорсальной стороны с незначительными вариациями в виде наличия у некоторых экземпляров более темной каймы по краям P и V, а также мелких точек и пятен на диске. Вариации окраски вентральной стороны тела были более значительными, хотя все изученные экземпляры характеризовались ее более темной окраской в сравнении с дорсальной стороной и наличием темной каймы на крыльях и нижних лопастях V.
Эмбрионы. Наши детальные исследования изученных ранее Штеманом и Мерреттом (Stehmann, Merrett, 2001) эмбрионов в фиксированном состоянии (№ 1–4 табл. 1) выявили некоторые дополнительные особенности их окраски. Эмбрион длиной 14 см имел одноцветную бледную окраску как в фиксированном, так и свежевыловленном состоянии (Stehmann, Merrett, 2001). У двух других эмбрионов TL 18 см (№ 1) и 22 см (№ 2) отсутствовало белое пятно у входа в клоаку. У первого из указанных экземпляров, кроме того, вся нижняя поверхность хвоста была белой, в отличие от большинства экземпляров, у которых светлый участок отмечен только у основания хвоста. У четырех из рассматриваемых экземпляров имелись белые пятна на брюхе различной конфигурации и местоположения, которые отсутствовали у скатов длиной свыше 85 см. Среди эмбрионов у всех самцов дорсальная поверхность птеригоподиев была светлой.
Неполовозрелые экземпляры. Согласно полученным нами данным, у всех неполовозрелых особей отмечено наличие белых участков вокруг рта, ноздрей, у входа в клоаку и у основания хвоста. В то же время, некоторые особенности их окраски заметно отличались от таковой созревающих и половозрелых экземпляров. Так, у некоторой части особей TL 29–69 см (4 из 11 экз.) отсутствовал белый участок между P и V при наличии его у всех особей меньшей длины и большинства более крупных экземпляров. У двух скатов длиной 69 см (№ 14 и 15) не отмечено белых участков на нижних лопастях V. У четырех особей TL 52–55 см (№ 8–11) и 69 см (№ 14) отсутствовали белые пятна на брюхе, присутствуя у большинства созревающих и половозрелых экземпляров. У скатов TL 55 см (№ 11) и 69 см (№ 14) не отмечено белых пятен на хвосте. У двух самцов TL 53 (№ 10) и 69 см (№ 14) отсутствовали белые участки на вершинах птеригоподиев, присутствуя у всех более мелких и большинства более крупных экземпляров.
Созревающие экземпляры. Для окраски вентральной стороны тела всех созревающих особей было характерно наличие белых участков вокруг рта, ноздрей и входа в клоаку. У одного из пяти исследованных экземпляров (№ 16) отсутствовало белое пятно между P и V. У трех из пяти особей (№ 16–17 и 19) не отмечено белых участков на нижних лопастях V. У всех созревающих экземпляров отсутствовали белые пятна на брюхе, за исключением самца TL 84 см (№ 17). У всех скатов рассматриваемой группы отсутствовали белые пятна на хвосте. Из двух изученных созревающих самцов (№ 16 и 17) лишь у одного вершины птеригоподиев были окрашены светло.
Половозрелые экземпляры. Как и подавляющее большинство особей других размерных групп, у половозрелых экземпляров отмечены белые участки вокруг рта и ноздрей и у входа в клоаку. Белые участки у основания хвоста имелись у трех из семи (№ 21, 23–24) изученных скатов. Наличие белых участков между P и V отмечено у двух (№ 23 и 28) из семи экземпляров. Нижние лопасти V были окрашены светло только у одной особи (№ 23). Белые пятна на брюхе и нижней поверхности хвоста ни у одного половозрелого ската не отмечены. При этом у всех изученных самцов данной группы вершины птеригоподиев были светло окрашенными.
Вариации окраски в зависимости от пола
Анализ вариаций окраски вентральной стороны тела 14 самок и 12 самцов ее принципиальных различий у особей разных полов не выявил. Отсутствие белого участка у входа в клоаку отмечено у двух наиболее мелких разнополых эмбрионов (№ 1 и 2). Белые участки у основания хвоста отсутствовали у одной крупной самки (№ 21) и четырех самцов TL 78–127 см (№ 16, 17, 23–24). Не было белых участков между P и V у трех самок TL 29, 45 и 146 см (№ 5, 7, 28) и 4 самцов TL 55, 69, 78, 113 см (№ 11, 14, 16, 23). Белые участки на нижних лопастях V не отмечены у двух самок TL 69 и 101 см (№ 15 и 19) и четырех самцов TL 69–113 см (№ 14, 16–17, 23). Белые пятна на брюхе отсутствовали у восьми самок TL 53 и >85 см, в то время как среди самцов данный признак не был выражен у экземпляров TL 52–78 см и 113–130 см. Белые пятна на хвосте отсутствовали у семи самок TL > 85 см и семи самцов >55 cм.
Вариации окраски в зависимости от глубины
У особей ската Ричардсона, пойманных на различных глубинах в диапазоне 1541–2951 м, отмечены некоторые различия особенностей окраски вентральной стороны тела. При их анализе указанный диапазон был разделен на три отрезка: 1541–1975 м (8 особей), 2125–2441 м (13 особей) и 2534–2951 м (5 особей). Скаты, у которых отсутствовали белые участки у входа в клоаку, обнаружены только на глубинах <2000. Белые участки у основания хвоста отсутствовали у двух экземпляров с глубин <2000 м и у трех – с глубин 2000–2500 м, но не отмечены у особей с глубин >2500 м. Белые участки между P и V отсутствовали у двух скатов, пойманных на глубине <2000 м, четырех – с глубин 2000–2500 м и одного с глубин >2500 м. На нижних лопастях V белые участки не обнаружены у трех особей с глубин <2000 м, двух – с глубин 2000–2500 м и одной с – глубин >2500 м. Белых пятен на брюхе не было у двух экземпляров, добытых на глубинах <2000 м, у десяти – на глубинах 2000–2500 м и у четырех – на глубинах >2500 м. Белые пятна на хвосте отсутствовали у трех особей с глубин <2000 м, у восьми – с глубин 2000–2500 м и у трех – с глубины >2500 м. Отсутствие белых вершин птеригоподиев обнаружено только у двух самцов с глубин менее 2000 м и одного с глубин свыше 2500 м.
Вариации окраски в зависимости от района
Небольшой объем материалов из вод Новой Зеландии (1 экз.) и СЗА (2 экз.) не позволяют их использовать в сравнительном анализе. Более объемные выборки из вод СВА (9 экз.), САХ и Азорских о-вов (14 экз.) предоставляют возможность сравнения окраски вентральной стороны тела рассматриваемого вида в указанных районах. Особи, у которых отсутствовал белый участок у входа в клоаку (2 экз.), отмечены только в СВА. Отсутствие белого участка у основания хвоста было более характерно для скатов из вод САХ (4 экз.), нежели СВА (1 экз.). Также у особей из вод САХ чаще отсутствовали белые участки между P и V (4 экз.) в сравнении с СВА (2 экз.), а также – на нижних лопастях V – 5 и 1 экз. соответственно. Белых пятен на брюхе не отмечено у 10 скатов из вод САХ и трех из СВА. Отсутствие белых пятен на хвосте отмечено у девяти особей из вод САХ и двух – из СВА. Попутно отметим, что у всех изученных нами скатов из вод Новой Зеландии и СЗА также не было белых пятен на брюхе и хвосте. Наличие белых вершин птеригоподиев отмечено только у трех самцов из вод САХ.
ОБСУЖДЕНИЕ
Имеющиеся в литературе сведения об окраске ската Ричардсона ограничиваются описанием таковой эмбрионов (Stehmann, Merrett, 2001) и взрослых половозрелых особей (Garrick, 1961; Forster, 1965; Tempelman, 1973a; Sulak et al., 2009; Ebert, Stehmann, 2013). Данные об особенностях окраски неполовозрелых и созревающих особей данного вида в опубликованных источниках до сих пор отсутствовали за исключением самки из района САХ (Orlov, 2011), имеющей некоторые аномалии во внешнем строении и окраске. Таким образом, имеющиеся до сих пор в литературе сведения не давали возможности описать онтогенетические изменения окраски ската Ричардсона в полном объеме, хотя упоминания о наличии таковых в опубликованной ранее литературе встречались (Orlov, Cotton, 2011).
Описывая окраску извлеченных из яйцевых капсул эмбрионов TL 14–25 см, Штеман и Мерретт (Stehmann, Merrett, 2001) отмечали, что она в целом соответствует таковой взрослых особей, но у первых окраска дорсальной и вентральной сторон отличаются сильнее в сравнении с последними. При этом у эмбрионов нижняя поверхность окрашена заметно темнее, в то время как у взрослых различия окраски дорсальной и вентральной сторон выражны не столь отчетливо. Наши исследования окраски фиксированных эмбрионов не обнаружили заметных различий между дорсальной и вентральной сторонами, однако обнаружили дополнительные особенности последней, на которые не было обращено внимания ранее и которые отсутствуют у взрослых особей.
Об особенностях окраски среднеразмерных скатов Ричардсона (неполовозрелых и созревающих) до сих пор сведения отсутствовали. Полученные нами данные по окраске среднеразмерных скатов Ричардсона значительно дополняют имеющиеся сведения и позволяют проследить онтогенетические изменения окраски рассматриваемого вида в полном объеме.
Сведений об окраске взрослых половозрелых особей ската Ричардсона в опубликованной литературе достаточно (Garrick, 1961; Forster, 1965; Tempelman, 1973a; Sulak et al., 2009; Ebert, Stehmann, 2013). Основная окраска дорсальной стороны, согласно нашим данным, принципиально ничем не отличалась от описанной ранее (Garrick, 1961; Forster, 1965; Tempelman, 1973a; Sulak et al., 2009; Ebert, Stehmann, 2013). Наличие на дорсальной стороне небольших светлых точек или пятен, а также темной каймы на краях диска и нижних лопастях V отмечены и нами у большинства проанализированных особей, за исключением самки длиной 110 см (№ 22) из вод Канарских о-вов (Brito et al., 1998). О наличии белых участков вокруг рта и ноздрей сообщали многие авторы (Garrick, 1961; Forster, 1965; Tempelman, 1973b). Наши данные в отношении крупных скатов полностью соответствуют указанным наблюдениям.
Результаты проведенного нами анализа позволяют существенно уточнить отдельные особенности окраски ветральной поверхности крупных экземпляров ската Ричардсона. Наличие белого участка у основания хвоста отмечено целым рядом авторов (Garrick, 1961; Forster, 1965; Tempelman, 1973b; Sulak et al., 2009; Ebert, Stehmann, 2013). Однако при этом одни авторы указывают на наличие белого участка у основания хвоста, другие – на наличие белых полос (пятен) на хвосте. По нашим данным, оба указанных признака следует рассматривать как независимые. Наличие белого участка у основания хвоста характерно не для всех крупных скатов, поскольку он не обнаружен у особей длиной 106–127 см (№ 21, 23–24), а белые пятна на хвосте зарегистрированы преимущественно у молодых экземпляров.
Наличие пятна у входа в клоаку ранее отмечалось рядом авторов (Garrick, 1961; Forster, 1965; Tempelman, 1973b). Согласно нашим данным, данный признак характерен для окраски всех крупных скатов рассматриваемого вида.
Еще один признак, который характерен для окраски взрослых скатов Ричардсона – наличие белого участка между P и V (Garrick, 1961; Tempelman, 1973b). По нашим данным, данный признак отсутствовал у самца длиной 113 см (№ 23) и самки длиной 146 см (№ 28).
Некоторые авторы (Sulak et al., 2009; Ebert, Stehmann, 2013) обращают внимание на наличие белых участков на нижних лопастях V. Среди крупных особей, изученных нами, данный признак отутствовал только у самки длиной 101 см (№ 19) и самца длиной 113 см (№ 23).
На наличие белых участков вокруг жаберных щелей, которые на приведенных изображениях практически не видны, указывали немногие авторы (Garrick, 1961; Tempelman, 1973b). Только на рисунке голотипа (Garrick, 1961) отчетливо видны узкие белые участки вокруг жаберных отверстий. Нашими исследованиями наличие белых участков вокруг жаберных щелей у исследованных крупных экземпляров не выявлено.
Еще одним признаком, характерным для крупных самцов ската Ричардсона, являются белые вершины птеригоподиев (Tempelman, 1973b). Наши исследования показали, что данный признак хорошо выражен у крупных (длиной свыше 84 см) самцов.
Последний признак окраски крупных особей, который упоминается в опубликованной литературе, – это светлые участки, расположенные вдоль срединной линии тела (Sulak et al., 2009; Ebert, Stehmann, 2013). Что из себя представляют данные участки, в указанных источниках не объясняется. Мы предполагаем, что речь в данном случае может идти о белых пятнах на брюхе в центральной части диска, которые, по нашим данным, у особей различной длины могут иметь разные местоположение и конфигурацию (см. выше), а также о белых участках у основания хвоста и вдоль него.
Таким образом, анализ литературных и собственных данных показывает, что отдельные признаки окраски ската Ричардсона сохраняются у него на протяжении всего жизненного цикла и практически не зависят от размера, пола, глубины и района. К ним относятся цвет обеих сторон тела, наличие светлых точек или пятен на дорсальной поверхности диска, наличие темной каймы на крыльях и нижних лопастях V с обеих сторон тела, белые участки вокруг рта и ноздрей.
Практически у всех особей (TL > 24 см) имеется белый участок у входа в клоаку, отсутствуя у лишь самых мелких эмбрионов (№ 1–2). У большей части особей имеется белый участок у основания хвоста, отсутствуя у нескольких созревающих (№№ 16–17) и половозрелых (№ 21, 23–24). Среди неполовозрелых скатов, в отличие от эмбрионов, появляются экземпляры (№ 5, 7, 11, 14), у которых отсутствует белый участок между P и V, что изредка отмечается также у созревающих (№ 16) и половозрелых (№ 23, 28) скатов. Белые участки на нижних лопастях V исчезают у неполовозрелых особей TL > 69 см (№ 14–15), что отмечено также у большей части созревающих (№ 16–17, 19) и одного половозрелого экземпляра (№ 23). Белые пятна на брюхе исчезают у неполовозрелых особей TL > 52 см (исключение № 13 TL 63 см) и не обнаруживаются как среди созревающих (№ 17 TL 84 см), так и половозрелых скатов. Белые пятна на хвосте характерны только для молодых экземпляров, они исчезают при TL > 55 см и не отмечаются у созревающих и половозрелых особей (исключение № 14 TL 69 см). Наличие белых участков на птеригоподиях характерны для большинства изученных самцов за исключением двух неполовозрелых (№ 10, 14) и одного созревающего (№ 16) экземпляров. На протяжении жизненного цикла данный признак подвержен существенным изменениям: если у эмбрионов и неполовозрелых самцов вентральная поверхность птеригоподиев целиком светлая, то у созревающих и половозрелый экземпляров окрашенными светло остаются только вершины птеригоподиев.
В целом, по окраске самцы и самки ската Ричардсона принципиально друг от друга не отличались. Однако можно выделить четыре признака, по которым окраска особей разных полов имела заметные различия: белый участок у основания хвоста (92.9% у самок против 66.7% у самцов), белый участок между P и V (78.6% против 66.7%), белый участок на нижних лопастях V (85.8% против 66.7) и белое пятно на брюхе (42.9% против 33.3%). Следует отметить, что рассматриваемые четыре признака у особей разных полов проявлялись по-разному. Если белый участок у основания хвоста имелся у всех самок (исключение № 21 TL 106 см), то он не отмечен у всех крупных самцов TL > 78 см (исключение № 25 TL 130 см). Практически у все крупные самки TL > 103 см имели белые участки на нижних лопастях V (без исключения) и между P и V (исключение № 28 TL 146 см), в то время как у более половины крупных самцов TL > 55 см (57.1%) эти признаки не были выражены. Наличие белых пятен на брюхе отмечалось преимущественно среди молодых экземпляров обоих полов. Можно предположить, что обнаруженные различия в окраске крупных самцов и самок, призваны облегчить поиск полового партнера в условиях низкой численности вида в условиях полного отсутствия света.
Принципиальных различий особенностей окраски ската Ричардсона в зависимости от глубины выявить не удалось. Можно лишь отметить, что эмбрионы, у которых отсутствовали белые участки у входа в клоаку, отмечены лишь на глубинах <2000 м. Вероятно, это связано с тем, что подобных находок на других глубинах пока нет. Среди скатов, пойманных на глубине >2500 м, не отмечено особей, у которых отсутствовал белый участок у основания хвоста. Белые участки на вершинах птеригоподиев имелись у всех скатов, пойманных на глубинах 2000–2500 м, в отличие от экземпляров с бóльших и меньших глубин. Наличие или отсутствие остальных изученных признаков отмечено у особей не зависимо от глубины поимки. Скорее всего, обнаруженные различия окраски на разных глубинах носят случайный характер, поскольку у рассматриваемого вида, в отличие от других глубоководных скатов, например, северо-тихоокеанских (Orlov et al., 2006b), онтогенетические вертикальные миграции не выражены, и размеры особей с глубиной не связаны (Orlov et al., 2006a).
Принципиальные различия в окраске ската Ричардсона в разных районах не обнаружены. Тем не менее, по некоторым признакам особи из вод САХ и СВА различались. Так частота встречаемости составила, соответственно, 71.4 и 88.9% для белых участков у основания хвоста, 64.3 и 77.8% – для белых участков между P и V, 64.3 и 88.9% – для белых участков на нижних лопастях V, 28.6 и 66.7% – для белых пятен на брюхе, 28.6 и 77.8% – для белых пятен на хвосте, 50 и 100% – для белых вершин птеригоподиев. Объяснить причины обнаруженных географических различий в окраске особей ската Ричардсона пока не представляется возможным. Однако имеются сведения о специфике окраски в водах САХ и СВА и других глубоководных скатов, например, ската Йенсена Amblyraja jenseni, бледного ската Bathyraja pallida и ската Кукуева Rajella kukujevi (Orlov, Cotton, 2013, 2015; Orlov, 2014).
Окраска свежевыловленных и фиксированных в формалине особей ската Ричардсона несколько различается, хотя и незначительно. Гаррик (Garrick, 1961) указывал, что после фиксации в формалине окраска особей становится несколько темнее. Темплмэн (Tempelman, 1973b) полагал, что после фиксации половина особей не меняет свою окраску, а у половины становится темнее вентральная поверхность, особенно по краям диска. Наши результаты показали, что после фиксации интенсивность окраски дорсальной и вентральной сторон эмбрионов практически не различается. Наши исследования глубоководных северо-атлантических скатов (Orlov, Cotton, 2013, 2015; Orlov, 2014; настоящее сообщение), основанные на изучении как свежевыловленных, так и фиксированных образцов, показывают, что и те, и другие вполне пригодны для анализа особенностей их окраски.
ЗАКЛЮЧЕНИЕ
Приведенные в данной статье и ранее опубликованные сведения о наличии видоспецифической окраски вентральной стороны у глубоководных северо-атлантических скатов ставят закономерный вопрос о ее биологическом смысле у видов, обитающих в условиях перманентного отсутствия света. По нашему мнению, данный феномен является эволюционным приобретением, направленным на облегчение поиска полового партнера в условиях низкой численности. Известно, что спариванию у различных видов надотряда Batoidea предшествует наплывание одного партнера над другим с последующим захватом самцом челюстями края крыла самки (Tricas, 1980; Luer, Gilbert, 1985; Yano et al., 1999; Chapman et al., 2003). Как происходит процесс спаривания у глубоководных скатов, неизвестно, но вряд ли он принципиально отличается от описанного выше. В условиях полной темноты и низкой численности успешность каждой попытки спаривания имеет высокое значение для выживания вида. Поэтому репродуктивный успех во многом зависит от корректной видовой идентификации партнерами друг друга. Вероятно, видоспецифические особенности окраски вентральной стороны тела глубоководных скатов служат своеобразным маркером в системе опознавания “свой–чужой”, что позволяет половым партнерам безошибочно определять не только принадлежность к одному виду, но и готовность к спариванию. Данная гипотеза вызывает закономерный вопрос о том, каким образом половые партнеры различают рисунок на вентральной поверхности тела? Известно, что различные виды скатообразных (Rajiformes) способны издавать слабоэлектрические сигналы (Baron et al., 1992; Барон, 1994; New, 1994; Sinceros et al., 1998; Sinceros, Tricas, 2002), используя их в том числе и для поиска полового партнера (Tricas et al., 1995). Можно предположить, что и глубоководные скаты в поисках походящего объекта для спаривания используют аналогичный механизм, который позволяет им различать особенности рисунка на вентральной стороне тела. Однако не следует исключать и роль зрения в этом процессе, поскольку наличие зрительного пигмента в глазах глубоководных видов, включая ската Ричардсона (Douglas et al., 1995; Lisney et al., 2012), свидетельтсвуют о том, что оно в жизни рассматриваемого вида играет не последнюю роль.
Список литературы
Барон В.Д. Возможная роль электрорецепции в поведении морских слабоэлектрических скатов Raja clavata (Rajidae) // Сенсор. сист. 1994. Т. 8. № 3–4. С. 147–161.
Орлов А.М. Итоги совещания специалистов ФАО и Всемирного фонда охраны природы “Сохранение и управление промыслом глубоководных видов хрящевых рыб” // Биология моря. 2005. Т. 31. № 1. С. 68–70.
Орлов А.М. К обоснованию промысловой меры дальневосточных скатов (сем. Rajidae) на примере массовых западноберинговоморских видов // Тр. ВНИРО. 2006. Т. 146. С. 252–264.
Орлов А.М. Красный список Международного союза охраны природы и природоохранный статус хрящевых рыб западной части Северной Пацифики // Вопр. ихтиологии. 2008. Т. 48. № 4. С. 575–576.
Орлов А.М., Литвинов Ф.Ф. Международные усилия по оценке природоохранного статуса хрящевых рыб Мирового океана // Тр. ВНИРО. 2010. Т. 149. С. 92–114.
Орлов А.М., Смирнов А.А. Новые данные о половом диморфизме и репродуктивной биологии щитоносного ската Bathyraja parmifera в северо-западной части Тихого океана // Вопр. ихтиологии. 2011. Т. 51. № 5. С. 614–627.
Орлов А.М., Шеверницкий Д.А. Природоохранный статус хрящевых рыб Северо-Восточной Атлантики // Бюлл. Моск. об-ва испыт. природы. Отд. биол. 2008. Т. 113. Вып. 4. С. 27–32.
Baron V.D., Sokolova L.S., Mikhailenko N.A. Identification of spinal electromotoneurons in the ray Raja clavata (Rajidae) // Neurosci. 1992. V. 48. № 2. P. 397–403.
Brito A., Pascual-Alayón P.J., Rabanal R. et al. Peces cartilaginosos de Canarias. Los tiburones de los fondos profundos y su aprovechamiento pesquero. Santa Cruz de Tenerife: Consejería de Agricultura, Pesca y Alimentación. Gobierno de Canarias, 1998. 171 p.
Cavanagh R.D., Gibson C. Overview of the Conservation Status of Cartilaginous Fishes (Chondrichthyans) in the Mediterranean Sea. Gland, Switzerland and Malaga, Spain: IUCN Species Survival Commission Shark Specialist Group, 2007. 42 p.
Chapman D.D., Corcoran M.J., Harvey G.M. et al. Mating behavior of southern stingrays, Dasyatis americana (Dasyatidae) // Environ. Biol. Fish. 2003. V. 68. № 3. P. 241–245.
Douglas R.H., Partridg J.C., Hope A.J. Visual and lenticular pigments in the eyes of demersal deep-sea fishes // J. Compar. Physiol. A. 1995. V. 177. № 1. P. 111–122.
Dulvy N.K., Reynolds J.D. Predicting extinction vulnerability in skates // Conserv. Biol. 2002. V. 16. № 2. P. 440–450. https://doi.org/10.1046/j.1523-1739.2002.00416.x
Ebert D.A., Compagno L.J., Cowley P.D. Aspects of the reproductive biology of skates (Chondrichthyes: Rajiformes: Rajoidei) from southern Africa // ICES J. Mar. Sci. 2008. V. 65. № 1. P. 81–102. https://doi.org/10.1093/icesjms/fsm169
Ebert D.A., Stehmann M.F.W. Sharks, batoids and chimaeras of the North Atlantic // FAO Spec. Catalogue Fish. Purpose. № 7. Rome: FAO, 2013. 523 p.
Forster G.R. Raja richardsoni from the continental slope off south-west England // J. Mar. Biol. Assoc. U.K. 1965. V. 45. P. 773–777. https://doi.org/10.1017/S0025315400016581
Fossen I., Cotton C.F., Bergstad O.A., Dyb J.E. Species composition and distribution patterns of fishes captured by longlines on the Mid-Atlantic Ridge // Deep-Sea Res. II. 2008. V. 55. P. 203–217. https://doi.org/10.1016/j.dsr2.2007.09.004
Garrick J.A.F. Studies on New Zealand Elasmobranchii. Part XIII. A new species of Raja from 1,300 fathoms // Trans. R. Soc. N.Z. 1961. V. 88. Pt. 4. P. 743–748.
Garrick J.A.F., Paul L.J. The taxonomy of New Zealand skates (suborder Rajoidea), with descriptions of three new species // J. R. Soc. N.Z. 1974. V. 4. Iss. 3. P. 345–377. https://doi.org/10.1080/03036758.1974.10419402
Goodwin H.B. Skate and ray management in the Northwest Atlantic: an overview of current management and recommendations for conservation. MS Thesis. Halifax: Dalhousie University, 2012. 114 p.
Kulka D.W., Orlov A., Barker A.S. Bathyraja richardsoni. The IUCN Red List of Threatened Species. 2015: e.T63127A70709214. Downloaded on 19 May 2020.https://doi.org/10.2305/IUCN.UK.2015-4.RLTS. T63127A70709214.en
Lisney T.J., Theiss S.M., Collin S.P., Hart N.S. Vision in elasmobranchs and their relatives: 21st century advances // J. Fish Biol. 2012. V. 80. № 5. P. 2024–2054. https://doi.org/10.1111/j.1095-8649.2012.03253.x
Luer C.A., Gilbert P.W. Mating behavior, egg deposition, incubation period, and hatching in the clearnose skate, Raja eglanteria // Environ. Biol. Fish. 1985. V. 13. № 3. P. 161–171.
New J.G. Electric organ discharge and electrosensory reafference in skates // Biol. Bull. 1994. V. 187. № 1. P. 64–75.
Orlov A.M. Record of tailless Richardson’s ray Bathyraja richardsoni (Garrick, 1961) (Rajiformes: Arhynchobatidae) caught off the Mid-Atlantic ridge // Pan-Am. J. Aquat. Sci. 2011. V. 6. № 3. P. 232–236.
Orlov A.M. New data on rare deep-water Mid-Atlantic skate Rajella kukujevi (Rajidae) // J. Ichthyology. 2014. V. 54. № 5. P. 317–337. https://doi.org/10.1134/S0032945214030102
Orlov A.M., Cotton C.F. Sexually dimorphic morphological characters in five North Atlantic deepwater skates (Chondrichthyes: Rajiformes) // J. Mar. Biol. 2011. V. 2011. Art. ID 842821. 18 p. https://doi.org/10.1155/2011/842821
Orlov A.M., Cotton C.F. New data on rare deepwater North Atlantic skate Bathyraja pallida (Forster, 1967) (Arhynchobatidae, Rajiformes) // J. Ichthyology. 2013. V. 53. № 7. P. 465–477. https://doi.org/10.1134/S0032945213040048
Orlov A.M., Cotton C.F. New data on the rare deep-sea skate Amblyraja jenseni (Rajidae) from the North Atlantic Ocean // J. Ichthyology. 2015. V. 55. № 4. P. 478–496. https://doi.org/10.1134/S0032945215040086
Orlov A., Cotton C., Byrkjedal I. Deepwater skates (Rajidae) collected during the 2004 cruises of R.V. “G.O. Sars” and M.S. “Loran” in the Mid-Atlantic Ridge area // Cybium. 2006. V. 30. № 4 (suppl.). P. 35–48.
Orlov A., Tokranov A., Fatykhov R. Common deep-benthic skates (Rajidae) of the northwestern Pacific: basic ecological and biological features // Cybium. 2006. V. 30. № 4 (suppl.). P. 49–65.
Orlov A.M., Cotton C.F., Shevernitsky D.A. Sexual dimorphism of external morphological characters in some deepwater skates (Rajidae, Rajiformes, Chondrichthyes) of the North Atlantic // Moscow Univ. Biol. Sci. Bull. 2010. V. 65. № 1. P. 40–44. https://doi.org/10.3103/S0096392510010086
Porteiro F.M., Sutton T.T., Byrkjedal I. et al. Fishes of the Northern Mid-Atlantic Ridge collected during the MAR-ECO cruise in June-July 2004: an annotated checklist // Arquipelago. Life Mar. Sci. Suppl. 2017. Suppl. 10. iii +126 p.
Sisneros J.A., Tricas T.C., Luer C.A. Response properties and biological function of the skate electrosensory system during ontogeny // J. Comp. Physiol. A. 1998. V. 183. № 1. P. 87–99.
Sisneros J.A., Tricas T.C. Ontogenetic changes in the response properties of the peripheral electrosensory system in the Atlantic stingray (Dasyatis sabina) // Brain Behav. Evol. 2002. V. 59. № 3. P. 130–140.
Stehmann M.F.W., Merrett N.R. First records of advanced embryos and egg capsules of Bathyraja skates from the deep north-eastern Atlantic // J. Fish Biol. 2001. V. 59. P. 338–349. https://doi.org/10.1111/j.1095-8649.2001.tb00134.x
Sulak K.J., MacWhirter P.D., Luke K.E. et al. Identification guide to skates (Family Rajidae) of the Canadian Atlantic and adjacent regions // Can. Tech. Rept. Fish. Aquat. Sci. 2009. № 2850. 34 p.
Tempelman W. The skate, Raja richardsoni Garrick, 1961, assigned to Bathyraja // J. Fish. Res. Board Can. 1973a. V. 30. P. 1729–1732.
Tempelman W. First records, description, distribution, and notes on the biology of Bathyraja richardsoni (Garrick) from the Northwest Atlantic // J. Fish. Res. Board Can. 1973b. V. 30. P. 1831–1840.
Tricas T.C. Courtship and mating-related behaviors in myliobatid rays // Copeia. 1980. V. 1980. № 3. P. 553–556.
Tricas T.C., Michael S.W., Sisneros J.A. Electrosensory optimization to conspecific phasic signals for mating // Neurosci. Letters. 1995. V. 202. № 1–2. P. 129–132.
Yano K., Sato F., Takahashi T. Observations of mating behavior of the manta ray, Manta birostris, at the Ogasawara Islands, Japan // Ichthyol. Res. 1999. V. 46. № 3. P. 289–296.
Дополнительные материалы отсутствуют.