

Палеонтологический журнал, 2019, № 1, стр. 15-23
Особенности распределения и морфологии вида Islandiella norcrossi (Cushman) в верхнечетвертичных отложениях моря Лаптевых
Я. С. Овсепян a, c, d, *, Е. Е. Талденкова b, c
a Геологический институт РАН
Москва, Россия
b Московский государственный университет им. М.В. Ломоносова, Географический ф-т
Москва, Россия
c Арктический и антарктический научно-исследовательский институт
Санкт-Петербург, Россия
d Институт океанологии им. П.П. Ширшова РАН
Москва, Россия
* E-mail: yaovsepyan@yandex.ru
Поступила в редакцию 2.10.2017
Принята к публикации 1.11.2017
Аннотация
На основе микрофоссилий из верхнеплейстоцен-голоценовых осадков моря Лаптевых анализируется экология и объем вида Islandiella norcrossi (Cushman, 1933), одного из типичных представителей фауны секреционно-известковых бентосных фораминифер Арктики. Данный вид распространен на шельфе и континентальном склоне арктических морей, он свидетельствует о становлении нормально-морских условий с сезонным ледовым покровом и холодными арктическими водными массами. Вид также приурочен к зоне продуктивности у границы сезонных льдов. Раковины в осадках из моря Лаптевых очень многочисленны, у них наблюдается некоторая изменчивость в размере и форме камер, что, возможно, связано с суровыми условиями арктического региона (ограниченным периодом роста и размножения); встречены аберрантные модификации с дополнительным устьем. В популяции присутствуют раковины макросферической и микросферической генерации, а также ювенильные формы.
ВВЕДЕНИЕ
Цель настоящей работы – проанализировать распределение, описать и проиллюстрировать изменчивость современных и ископаемых раковин вида Islandiella norcrossi (Cushman, 1933), который является одним из самых массовых среди бентосных фораминифер Арктики. При его определении часто возникают трудности, связанные с изменчивостью морфологии раковины и особенностями диагностических признаков, которые отличают его от других видов рода Islandiella Nørvang, 1958 и от видов близкого рода Cassidulina d’Orbigny, 1826.
Подробных работ, в которых приводится описание раковины и структуры устья I. norcrossi немного (Nørvang, 1958; Knudsen, 1971; Rodrigues et al., 1980; Nomura, 1983a, b; Гуськов, 1997; Погодина, 2000; Бордунов, 2015). В основном в статьях I. norcrossi упоминается в списках фауны, редко с фотографиями (Culver, Buzas, 1985; Osterman, 1996; Wollenburg, Mackensen, 1998).
Море Лаптевых – одно из окраинных морей российской Арктики, характеризующееся суровыми природными условиями. Главными особенностями региона являются сильно выраженная сезонность, проявляющаяся летом в чрезвычайно больших объемах стока сибирских рек (Лена, Яна, Хатанга, Анабар и Оленек) и в суровых ледовых условиях в остальное время года (Добровольский, Залогин, 1982). С сезонностью связано неравномерное поступление питательных веществ и сложная структура взаимодействия различных водных масс, что отражается на составе фауны. Вероятно, природные особенности региона повлияли на некоторую изменчивость раковин вида I. norcrossi, подробное описание которого приводится в данной статье.
МАТЕРИАЛ И МЕТОДИКА
Материалом для палеонтологических исследований послужили пробы поверхностных осадков и верхнеплейстоцен-голоценовых отложений, вскрытых колонками из западной части моря Лаптевых (рис. 1, а, б). Образцы получены в российско-германских экспедициях ТРАНСДРИФТ 1998–2010 гг.
Рис. 1.
Расположение станций в акватории моря Лаптевых: а – положение колонок PS51/154-11 и PS51/159-10; б – распределение I. norcrossi (Cushman, 1933) в поверхностных образцах.
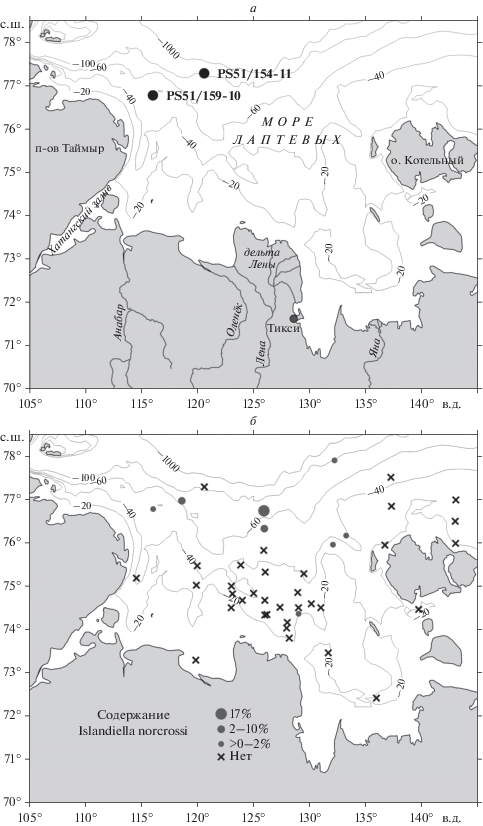
Раковины I. norcrossi в большом количестве присутствуют в двух морских колонках (рис. 1, а): PS51/154-11 (континентальный склон, гл. 270 м, керн 700 см) и PS51/159-10 (внешний шельф, гл. 60 м, керн 490 см). Отложения достаточно однородные, представлены песчано-алевритистой глиной и песчаным илом. Для разрезов сделаны возрастные модели на основе радиоуглеродных датировок (Bauch et al., 2001; Талденкова и др. 2009; Taldenkova et al., 2010; Овсепян и др., 2015; Овсепян, 2016).
При изучении современного распределения фораминифер среди 42 станций по всей акватории моря раковины I. norcrossi были найдены в 7 пробах с глубин от 45 до 120 м, а также в одной мелководной (33 м) (рис. 1, б) (Овсепян, 2016).
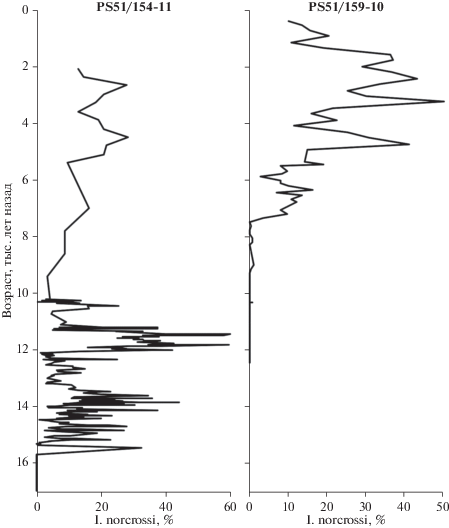
Бентосные фораминиферы изучены во фракции >63 мкм; составлена коллекция микрофоссилий № PS51-1998, которая хранится в Лаборатории новейших отложений и палеогеографии плейстоцена (географический ф-т МГУ им. М.В. Ломоносова). Графики изменения процентного содержания I. norcrossi относительно календарного возраста показаны для образцов, в которых содержание бентосных фораминифер более 100 экз. (рис. 2).
Рис. 2.
Распределение процентного содержание I. norcrossi (Cushman, 1933) в комплексе фораминфер в зависимости от возраста (тыс. лет назад) для колонок PS51/154-11 и PS51/159-10.
Авторами исследована морфология раковины, составлено подробное описание. Для изучения структуры стенки экземпляры I. norcrossi дополнительно чистились в ультразвуковой ванне. Фораминиферы сфотографированы при помощи светового бинокулярного и поляризационного микроскопов (центр оптической микроскопии ГИН РАН и лаборатория им. О.Ю. Шмидта ААНИИ), а также электронного сканирующего микроскопа (кабинет приборной аналитики ПИН РАН). Фотографии обработаны с помощью графических редакторов Helicon Focus, Corel Draw, Corel Photo-Paint, составлено 2 фототаблицы.
В статье для описания структурных элементов устья используется терминология из работ Р. Номуры (Nomura, 1983a, b). Для устья (форамена) выделяются передний край (ПК) и задний край (ЗК) по направлению нарастания раковины. Первичный язык (ПЯ) – разновидность зубной пластины, имеющей более сложное строение; это внутреннее устьевое образование фронтальной стенки, связанное широким основанием со стенкой предыдущей раковины и передним краем предшествующего форамена. Гребенчатый зуб (ГЗ) – конец первичного языка, выступающий наружу с зазубренным краем. Оба находятся на заднем краю устья. Вторичный язык (ВЯ) – структура, расположенная в переднем краю устья в месте сочленения фронтальной стенки раковины и септальной стенки смежной камеры. Губа (Г) – изогнутая краевая часть стенки раковины, образующей устье, на противоположенной стороне от первичного и вторичного языка. Апертурный гребень (АГ) – утолщение внешней части стенки предыдущей камеры в месте образования устья новой камеры.
В работе используется систематика I. norcrossi, принятая О. Гроссом, опубликованная во “Всемирной базе данных по современным фораминиферам”, на сайте www.marinespecies.org/foraminifera (Gross, 2001; Hayward et al., 2011).
РЕЗУЛЬТАТЫ
ТИП FORAMINIFERA
КЛАСС POLYTHALAMEA
ОТРЯД ROTALIIDA
СЕМЕЙСТВО CASSIDULINIDAE D’ORBIGNY, 1839
Род Islandiella Nørvang, 1958
Islandiella: Nørvang, 1958, c. 26; Feyling-Hanssen, Buzas, 1976, c. 155; Rodrigues et al., 1980, c. 49; Nomura, 1983a, c. 46; Loeblich, Tappan, 1988, c. 654; Гуськов, 1997, с. 99.
Cassilamellina: Волошинова, 1960, с. 59.
Planocassidulina: Гудина, 1966, с. 48; Бордунов, 2015, с. 90.
Типовой вид – Cassidulina islandica Nørvang, 1945; современный, прибрежные воды Исландии, обозначен автором рода.
Диагноз. Раковина шарообразная, сдавленная с боковых сторон, или линзовидная, спирально-плоскостная, двухрядная, инволютная. Контур раковины округлый, волнистый. Периферический край закругленный или приостренный, иногда с килем. В последнем обороте три–пять пар камер. Камеры объемные, на боковой стороне раковины проецируются в виде удлиненных треугольников, ромбоэдров или овалов. Швы четкие, слабо углубленные или расположены вровень с поверхностью. Стенка кальцитовая, многослойная (число слоев зависит от количества камер), оптически радиально-лучистая, прозрачная или матовая, перфорированная. Устье интермаргинальное, вытянутое вдоль периферического края в виде широкой щели, иногда треугольной формы, со скульптурными элементами, представленными губой, апертурным гребнем, первичным и вторичным языком, гребенчатым зубом. Основание первичного языка протягивается от заднего края устья к переднему краю форамена предшествующей камеры.
Видовой состав. Семь видов: I. californica (Cushman et Hughes, 1925), верхний миоцен-плейстоцен Сахалина и Камчатки, средний миоцен – современность, сев. часть Тихого океана, зап. побережье Сев. Америки (Rodrigues et al., 1980; Бордунов, 2015); I. compressa Nomura, 1983, плейстоцен, Японское море (Nomura, 1983b); I. islandica (Nørvang, 1945), неоген Сахалина и Камчатки, четвертичные отложения Севера Евразии и Сев. Америки, современные – повсеместно в Арктике, Северной Пацифике и Атлантике (Rodrigues et al., 1980; Гуськов, 1997; Бордунов, 2015); I. japonica (Asano et Nakamura, 1937), плиоцен – современность Японии, Сахалина, Камчатки, сев.-зап. часть Тихого океана (Nomura, 1983b; Бордунов, 2015); I. norcrossi (Cushman, 1933) – четвертичные отложения севера Евразии и Сев. Америки, современные – повсеместно в Арктике, Северной Пацифике и Атлантике (Rodrigues et al., 1980; Гуськов, 1997; Погодина, 2000; Polyak et al., 2002); I. subdelicata Golik, 1977, современные, Панамский залив (Golik, 1977); I. vejlensis Nørvang, 1958, верхний эоцен Сев. Европы (Nørvang, 1958).
Сравнение. У представителей рода Cassidulina d’Orbigny, 1826 схожая форма раковины и тип навивания, структура стенки зернистая, но иногда радиально-лучистая, поэтому следует обращать внимание на строение устья. У раковин Cassidulina устье в виде узкой щели, обрамленной внешними структурными элементами: губой и пластиной, которые не имеют внутреннего основания. Губу можно перепутать с гребенчатым зубом Islandiella, являющимся внешней частью первичного языка.
Замечания. Род Islandiella был выделен А. Норвангом в 1958 г. для видов, ранее относившихся к роду Cassidulina, имеющих радиально-лучистую структуру стенки и сложное строение устья с внутренними элементами, в том числе первичным языком, связанным с фораменом предшествующей камеры.
Islandiella norcrossi (Cushman, 1933)
Табл. I , фиг. 1–11, 13–15; табл. II , фиг. 1–8 (см. вклейку)
Cassidulina norcrossi: Cushman, 1933, с. 7, табл. 2 , фиг. 7 a–c.
Islandiella norcrossi: Nørvang, 1958, с. 32, фиг. 8–14; Knudsen, 1971, с. 248, табл. 8 , фиг. 1, 2; Rodrigues et al., 1980, с.55, табл. 4 , фиг. 1, 3, 5, 10; Østby, Nagy, 1982, с. 89, табл. 1 , фиг. 18; Nomura, 1983b, c. 13, табл. 2 , фиг. 2, 3, табл. 8 , фиг. 3-10; Гуськов, 1997, с. 108, табл. 3 , фиг. 1-9; Polyak et al., 2002, табл. 2 , фиг. 13; Лукина, 2004, с. 108, табл. VI , фиг. 86.
Planocassidulina norcrossi: Гудина, 1966, с. 138, табл. 6 , фиг. 2–3; Бордунов, 2015, с. 121, табл. XX , фиг. 7.
Islandiella helenae: Østby, Nagy, 1982, с. 89, табл. 1 , фиг. 17; Погодина, 2000, табл. 3 , фиг. 5.
Cassidulina teretis: Лукина, 2004, с. 108, табл. VI , фиг. 84.
Голотип – Национальный музей естественной истории (США, Вашингтон), № 26139. Происходит из поверхностных морских осадков в 5 милях от м. Борлас-Уоррен, сев.-вост. Гренландия (Cushman, 1933).
Описание. Раковина линзовидная. Периферический край с закругленным килем. Раковина инволютная c 8–10 камерами в последнем обороте, которые в проекции на боковую сторону имеют треугольную или ромбоидную форму. Швы четкие, тонкие, изогнутые, вровень с поверхностью, в пупке не сходятся. Пупочная область на обеих сторонах выпуклая, состоит из прозрачного натечного материала раковины, через который иногда видны камеры предыдущих оборотов. Устье интермаргинальное, широкое, щелевидной или треугольной формы, часто со вторичным и первичным языками, гребенчатым зубом, губой и апертурным гребнем. Стенка оптически радиально-лучистая, призматическая, прозрачная, тонкая. Средний диаметр пор 0.5 мкм, плотность в среднем 12 пор на 10 мкм2.
Размеры. Диаметр раковины 200–600 мкм, толщина 100 – 250 мкм.
Возрастные изменения. Раковины каплевидной формы малого диаметра (200 мкм) с общим количество камер 4–6 штук, можно отнести к ювенильным (табл. II , фиг. 1, 2) (морфотип 1). В том случае, когда общее число камер больше 6, и они образуют полный оборот, можно говорить о взрослых особях. При этом среди них встречаются формы, выделяющиеся большой и шаровидной начальной камерой (табл. I , фиг. 9, 10; табл. II , фиг. 3), что влияет на их контур, делая его не симметричным относительно периферического края (морфотип 2). Такие раковины, вероятно, относятся к макросферической генерации, их доля может доходить до 20% от общего количества I. norcrossi в образце.
Изменчивость. Раковины I. norcrossi из колонок с шельфа и континентального склона моря Лаптевых очень разнообразны, даже в одной пробе они могут быть представлены как ювенильными, так и взрослыми особями; у последних изменяется размер раковины, количество камер и их соотношение в последнем обороте. Стенка может быть как прозрачной, так и матовой. На табл. II представлены различные формы из одного образца с гл. 395 см в колонке PS51/154-11. Камеры имеют проекцию на боковую сторону в виде треугольников с острыми или закругленными вершинами, которые могут быть как одинаковой, так и разной формы, отмечается чередование крупных и мелких треугольников (морфотип 3: табл. I , фиг. 11; табл. II , фиг. 4). У крупных форм в последнем обороте все камеры, или только последняя, могут иметь форму округлых ромбов или овалов (морфотип 4: табл. II , фиг. 5, 6). Встречаются аберрантные (измененные) модификации (морфотип 5), когда происходит сращивание двух раковин, или появление дополнительных устьев (табл. I , фиг. 13–15; табл. II , фиг. 7, 8).
Сравнение. Отличается от других представителей рода Islandiella линзовидной (дисковидной) формой раковины и отсутствием вытянутых прямоугольных камер. Элементы устья I. norcrossi немного меньше по размеру, чем у других видов. От линзовидных раковин близкого рода Cassidulina отличается строением устья и структурой стенки.
Замечания. На материале из Зап. Сибири В.И. Гудина (1966) выделила новый род, состоящий из одного вида Planocassidulina norcrossi на основе последовательно расположенных одинаковых треугольных камер в последнем обороте, в то время как большинство авторов указывает на чередующееся расположение камер (Nomura, 1983a, b; Гуськов, 1997; Бордунов, 2015). В описании Islandiella norcrossi также отмечают изменения в размере и формы камер от округлых ромбоэдров до треугольников (Knudsen, 1971), и макросферические и микросферические генерации (Nørvang, 1958).
I. norcrossi иногда объединяют с представителями другого рода: Cassidulina neoteretis Seidenkrantz, 1995 или С. teretis Tappan, 1951 из-за схожести в морфологии (Osterman, 1996; Scott et al., 2008). Основным критерием, помимо устья (если оно обломано), служит структура стенки; при изучении в скрещенных николях кальцит раковины I. norcrossi показывает погасание, как единый кристалл (табл. I , фиг. 11), а у гранулярной стенки С. neoteretis видна зернистость (табл. I , фиг. 12).
При ревизии вида С. teretis раковины с признаками рода Islandiella выделили в отдельный вид I. helenae (Feyling-Hanssen et Buzas, 1976). От вида I. norcrossi он отличается только формой камер, при этом данный признак обладает изменчивостью на видовом уровне (Nørvang, 1958; Knudsen, 1971) и наблюдается в онтогенезе (Nomura, 1983b). Многие авторы при палеоэкологических исследованиях объединяют эти виды, обозначая их при написании через косую черту, как Islandiella norcrossi/helenae (Koç et al., 2002; Polyak et al., 2002; Rutter et al., 2002; Ślubowska-Woldengen et al., 2007; Sacher et al., 2009; Taldenkova et al., 2008, 2010) или Islandiella norcrossi s.l. (Korsun, Hald, 1998). Возможно, для описания раковин с камерами овальной и ромбоидной формы в дальнейшем следует использовать название Islandiella norcrossi forma helenae.
В связи с тем, что вид I. norcrossi был описан раньше, это название обладает приоритетом, поэтому выбрано авторами статьи.
Распространение. Плейстоцен-голоцен побережья Сев. Америки, Гренландии, севера Евразии и северо-запада Тихого океана (Камчатка, Япония). Современные представители обитают в Арктике, Северной Пацифике, в Сев. и Ц. Атлантике, в том числе, в Мексиканском заливе и Средиземном море.
Материал. Поверхностные пробы из моря Лаптевых – 193 экз. Колонка PS 51/154-11 – 23 061 экз; колонка PS 51/159-10 – 2683 экз.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ И ВЫВОДЫ
Для реконструкций условий палеосреды часто используется I. norcrossi. Изучение распределения этого вида в поверхностных образцах из моря Лаптевых подтвердило его принадлежность к группе фораминифер внешнего шельфа и континентального склона, удаленных от опресняющего влияния речного стока (см. рис. 1, б). I. norcrossi предпочитает холодные арктические водные массы в районах с сезонным ледовым покровом. В Арктике он достигает максимальной численности в интервале глубин от 50 до 1200 м на внешнем шельфе и континентальном склоне в морях Лаптевых, Карском и Баренцевом (Steinsund et al., 1994; Korsun, Hald, 1998; Polyak et al., 2002).
При палеоэкологическом анализе важно различать сходные по морфологии раковины видов I. norcrossi и С. neoteretis, так как последний обитает на больших глубинах (200–1400 м) и является индикатором атлантических водных масс (Lubinski et al., 2001; Wollenburg et al., 2004; Ślubowska-Woldengen et al., 2007). По распределению C. neoteretis в осадках колонок из моря Лаптевых выделяются периоды увеличения атлантического влияния (Taldenkova et al., 2010, 2012; Овсепян и др., 2015), в то время как по распределению I. norcrossi таких выводов сделать нельзя. Поэтому важно проводить диагностику элементов устья у линзовидных раковин семейства Сassidulinidaе и дополнительно изучать под поляризационным микроскопом структуру стенки.
В колонке PS 51/154-11 с верхней части континентального склона I. norcrossi является одним из массовых видов, который встречается во всех образцах в интервале 17–573 см, начиная с 15.4 тыс. лет назад и до современности, и показателен для нормальных морских условий (рис. 2). Максимальная численность вида в комплексе (60%) отмечается в интервале 200–300 см, соответствующем переходу от позднего дриаса к голоцену (11.2–12 тыс. лет назад). Для этого времени характерно увеличение биоразнообразия среди бентосных фораминифер; кроме того, содержание I. norcrossi может свидетельствовать не только о сезонном ледовом покрове, но и о зоне продуктивности на границе таяния морского льда (Taldenkova et al., 2012; Овсепян и др., 2015).
В колонке PS 51/159-10 раковины I. norcrossi постоянно присутствуют в верхних 100 см, в интервале 0.4–7.3 тыс. лет назад, где составляют до 50% комплекса бентосных фораминифер (рис. 2). Cмена комплексов микрофауны в колонке с внешнего шельфа рассматривается в рамках послеледниковой трансгрессии (Bauch et al., 2001), при этом интервал с преобладанием I. norcrossi соответствует периоду завершения роста уровня моря. Данный комплекс фораминифер свидетельствует о становлении современных морских условий с нормальной соленостью на внешнем шельфе (глубина 60 м), начиная с 7.3 тыс. лет назад (Талденкова и др., 2009; Овсепян, 2016).
Среди видов экологической группы внешнего шельфа и континентального склона [Astrononion gallowayi Loeblich et Tappan, 1953, Cibicides lobatulus (Walker et Jacob, 1798), Melonis barleeanus (Williamson, 1948), Stainfortiha feylingi Knudsen et Seidenkrantz, 1994] Islandiella norcrossi имеет максимальную численность в разрезах и поверхностных пробах из моря Лаптевых, поэтому более важен для палеоэкологического анализа. Обилие раковин позволяет наглядно изучить объем вида для довольно короткого интервала времени (последние 15.4 тыс. лет). Следует отметить, что особенностью фораминифер из моря Лаптевых является лучшая сохранность и большие размеры раковин, по сравнению с материалом из Баренцева моря (Чистякова и др., 2010). Вероятно, размеры фораминифер связаны с интенсивными темпами роста раковин в сокращенный летний период вегетации в суровых арктических условиях, при этом период размножения может откладываться на более поздний срок, чем в субарктических и бореальных районах. Данные наблюдения также приводились P. Фейлинг-Хансенном для Канадской Арктики (Feyling-Hanssen, 1980).
Возможно, что с интенсивным периодом роста, когда в короткий летний период поступает большое количество органики, связаны аберрантные (уродливые) формы раковин (С.А. Корсун, устн. сообщ.). Причем данные изменения не проявляются у представителей других видов из моря Лаптевых. Сросшиеся раковины I. norcrossi, или имеющие дополнительные устья, найдены в количестве более 100 экз. Появление видоизмененных раковин часто связывают с загрязнением окружающей среды тяжелыми металлами (Yanko et al., 1999), но, так как район исследований не подвергался антропогенному воздействию, эту причину можно исключить.
ЗАКЛЮЧЕНИЕ
Изучение распределения фораминифер вида Islandiella norcrossi в море Лаптевых позволяет заключить, что он является индикатором нормально-морских условий с холодными арктическими водными массами на внешнем шельфе и континентальном склоне, где отсутствует речное влияние. Вид также приурочен к зоне продуктивности у границы сезонных льдов.
Всесторонний анализ морфологии раковин показал, что вид обладает некоторой изменчивостью, выраженной в вариациях размера и формы камер из последнего оборота. Возможно, это связано с суровыми условиями этого арктического региона, ограниченным периодом роста и размножения.
В материале из моря Лаптевых I. norcrossi представлены как ювенильными, так и взрослыми особями, среди которых наблюдается изменчивость в виде раковины с одинаковыми и неодинаковыми по размеру камерами; встречены формы макросферической генерации, а также аберрантные сросшиеся модификации с дополнительным устьем.
Дальнейшие исследования будут направлены на изучение генетического материала живых фораминифер и установления степени родства для семейства Сassidulinidae из окраинных морей Российской Арктики.
* * *
Авторы выражают благодарность Х. Кассенс и Х.А. Бауху за предоставленный материал. Большое спасибо сотрудникам кабинета приборной аналитики ПИН РАН, центра оптической микроскопии ГИН РАН, лаб. им. О.Ю. Шмидта ААНИИ за помощь в фотографировании. Авторы признательны А.С. Алексееву и С.А. Корсуну за консультации и рекомендации по описанию материала.
Работа Овсепяна Я.С. поддержана из бюджетной темы ГИН РАН № 0135-2019-0057. Работа Талденковой Е.Е. поддержана субсидией Минобрнауки по проекту “Изменчивость Арктической Трансполярной системы” (СATS) (уникальный идентификатор проекта – RFMEFI61617X0076).
Список литературы
Бордунов С.И. Стратиграфия и фораминиферы неогена Восточной Камчатки. М.: ГЕОС, 2015. 148 с.
Волошинова Н.А. Успехи микропалеонтологии в деле изучения внутреннего строения фораминифер // Тр. Первого семинара по микрофауне. Л.: Гостоптехиздат, 1960. С. 48–77.
Гудина В.И. Фораминиферы и стратиграфия четвертичных отложений северо-запада Сибири. М.: Наука, 1966. 376 с.
Гуськов А.С. Исландиеллы и кассидулины (фораминиферы) Баренцева моря // Биостратиграфия и микроорганизмы фанерозоя Евразии. Тр. ХII микропалеонтол. совещ., посвященного 100-летию Д. М. Раузер-Черноусовой. М.: ГЕОС, 1997. С. 97–112.
Добровольский А.Д., Залогин Б.С. Моря СССР. М.: Изд‑во МГУ, 1982. 192 с.
Лукина Т.Г. Особенности распределения фораминифер в море Лаптевых // Фауна и экосистемы моря Лаптевых и сопредельных глубоководных участков арктического бассейна. СПб., 2004. С. 86–123 (Исследования фауны морей. Т. 54. № 62).
Овсепян Я.С., Талденкова Е.Е., Баух Х.А., Кандиано Е.С. Реконструкция событий позднего плейстоцена–голоцена на континентальном склоне моря Лаптевых по комплексам бентосных и планктонных фораминифер // Стратигр. Геол. корреляция. 2015. Т. 23. № 6. С. 964–112.
Овсепян Я.С. Позднечетвертичные фораминиферы моря Лаптевых и реконструкции изменения среды на основе палеоэкологического анализа. Дисс. … к.г.-м.н. М., 2016. 240 с.
Погодина И.А. Стратиграфия верхнечетвертичных отложений Баренцева моря по фораминиферам. Дисс. … к.г.-м.н. М., 2000. 212 с.
Талденкова Е.Е., Баух Х.А., Степанова А.Ю. и др. Изменения палеосреды шельфов морей Лаптевых и Карского в ходе послеледниковой трансгрессии (по ископаемой бентосной фауне) // Система моря Лаптевых и прилегающих арктических морей: современное состояние и история развития. М.: Изд-во Моск. ун-та, 2009. С. 384–409.
Чистякова Н.О., Иванова Е.В., Рисебробаккен Б. и др. Реконструкция послеледниковых обстановок в юго-западной части Баренцева моря по комплексам фораминифер // Океанология. 2010. Т. 50. № 4. С. 608–617.
Bauch H.A., Mueller-Lupp T., Taldenkova E. et al. Chronology of the Holocene transgression at the North Siberian margin // Global and Planetary Change. 2001. V. 31 P. 125–139.
Cushman J.A. New Arctic foraminifera collected by Capt. R.A. Bartlett from Fox Basin and off the Northeast coast of Greenland // Smithson. Misc. Coll. 1933. V. 89. № 9. P. 1–18.
Culver S.J., Buzas M.A. Distribution of Recent benthic Foraminifera off the North American Pacific coast from Oregon to Alaska // Smithson. Contrib. Mar. Sci. 1985. V. 26. P. 1–234.
Feyling-Hanssen R.W., Buzas M.A. Emendation of Cassidulina and Islandiella helenae new species // J. Foram. Res. 1976. V. 6. № 2. P. 154–158.
Feyling-Hanssen R.W. Microbiostratigraphy of young Cenozoic marine deposits of the Qivituq Peninsula, Baffin Island // Mar. Micropaleontol. 1980. V. 5. P. 153–184.
Golik A., Phleger F.B. Benthonic foraminifera from the Gulf of Panama // J. Foram. Res. 1977. V. 7. № 2. P. 83–99.
Knudsen K.L. Systematic part // Late Quaternary foraminifera from Vendsyssee, Danmark and Sandnes, Norway / Eds. Feyling-Hanssen R.W., Jorgensen J.A., Knudsen K.L., Lykke-Andersen A.L. Geol. Soc. Denmark Bull. 1971. V. 21. P. 185–291.
Koç N., Klitgaard-Kristensen D., Hasle K. et al. Late glacial palaeoceanography of Hinlopen Strait, northern Svalbard // Polar Res. 2002. V. 21. № 2. P. 307–314.
Korsun S., Hald M. Modern benthic foraminifera of Novaya Zemlya tide water glaciers, Russian Arctic // Arctic and Alpine Res. 1998. V. 30. № 1. P. 61–77.
Loeblich A.R., Tappan H. Foraminiferal genera and their classification. N. Y.: Van Nostrand Reinhold Co., 1988. 970 p.
Lubinski D.J., Polyak L.A., Forman S.L. Freshwater and Atlantic water inflows to the deep northern Barents and Kara seas since ca 13 14C ka: foraminifera and stable isotopes // Quatern. Sci. Rev. 2001. V. 20. P. 1851–1879.
Nomura R. Cassidulinidae (Foraminiferida) from the Uppermost Cenozoic of Japan (Part 1) // Tohoku Univ. Sci. Rep., 2nd ser. (Geol.) 1983a. V. 53. № 1. P. 1–101.
Nomura R. Cassidulinidae (Foraminiferida) from the Uppermost Cenozoic of Japan (Part 2) // Tohoku Univ. Sci. Rep., 2nd ser. (Geol.) 1983b. V. 54. № 1. P. 1–93.
Nørvang A. Islandiella and Cassidulina D’Orbigny // Vidensk. Medd. Dansk naturh. Foren. 1958. V. 120. P. 25–41.
Osterman L.E. Pliocene and Quaternary benthic foraminifers from Site 910, Yermak Plateau // Proc. of ODP. Sci. Results, 151: College Station, TX (Ocean Drilling Program) / Eds. Thiede J., Myhre A.M., Firth J.V. et al. 1996. P. 187–195.
Østby K., Nagy J. Foraminiferal distribution in the western Barents Sea, Recent and Quaternary // Polar Res. 1982. V. 1982. № 1. P. 53–87.
Polyak L., Korsun S., Febo L. et al. Benthic foraminiferal assemblages from the southern Kara Sea, a river-influenced Arctic marine environment // J. Foram. Res. 2002. V. 32. № 3. P. 252–273.
Rodrigues C., Hooper K., Jones P. The apertural structures of Islandiella and Cassidulina // J. Foram. Res. 1980. V. 10. № 1. P. 48–60.
Rutter F., Knudsen K., Seidenkrantz M.-S., Erikson J. Modern distribution of benthic foraminifera on the north Icelandic shelf and slope // J. Foram. Res. 2002. V. 32. № 3. P. 217–244.
Sacher M., Klitgaard-Kristansen D., Hald M. et al. Benthic foraminiferal assemblages in the Central Barents Sea: an evaluation of the effect of combining live and total fauna studies in tracking environmental change // Norw. J. Geol. 2009. V. 89. P. 149–161.
Scott D., Schell T., Rochon A., Blasco S. Modern benthic foraminifera in the surface sediments of the Beaufort shelf, slope and Mackenzie trough, Beaufort Sea, Canada: taxonomy and summary of surficial distributions // J. Foram. Res. 2008. V. 28. № 3. P. 228–250.
Ślubowska-Woldengen M., Rasmussen T.L., Koç N. Advection of Atlantic water to the western and northern Svalbard shelf since 17 500 cal yr BP // Quatern. Sci. Rev. 2007. V. 26. P. 463–478.
Steinsund P.I., Polyak L., Hald M. et al. Distribution of calcareous benthic foraminifera in recent sediments of the Barents and Kara Seas // Benthic foraminifera in surface sediments of the Barents and Kara Seas: modern and late Quaternary applications. Steinsund P.I., Dr. sci. thesis, Univ. Tromsø. 1994. P. 61–102.
Taldenkova E., Bauch H.A., Gottschalk J. et al. History of ice-rafting and water mass evolution at the North Siberian continental margin (Laptev Sea) during Late Glacial and Holocene times // Quatern. Sci. Rev. 2010. V. 29. P. 3919–3935.
Taldenkova E., Bauch H.A., Stepanova A. et al. Postglacial to Holocene history of the Laptev Sea continental margin: Palaeoenvironmental implications of benthic assemblages // Quatern. Intern. 2008. V. 183. P. 40–60.
Taldenkova E., Bauch H.A., Stepanova A. et al. Benthic community changes at the North Siberian margin in response to Atlantic water mass variability since last deglacial times // Mar. Micropaleontol. 2012. V. 96-97. P. 13–28.
Wollenburg J.E., Knies J., Mackensen A. High-resolution paleoproductivity fluctuations during the past 24 kyr as indicated by benthic foraminifera in the marginal Arctic Ocean // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2004. V. 204. P. 209–238.
Wollenburg J., Mackensen A. Living benthic foraminifers from the central Arctic Ocean: faunal composition, standing stock and diversity // Mar. Micropaleontol. 1998. V. 34. № 3–4. P. 153–185.
Yanko V., Arnold A., Parker W. Effects of marine pollution on benthic foraminifera // Modern foraminifera / Ed. B.K. Sen Gupta. N.Y.: Kluwer Acad. Publ., 1999. P. 217–235.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал