

Палеонтологический журнал, 2019, № 1, стр. 5-14
О новых подходах изучения эволюции макротаксонов и концепции экона
И. А. Вислобокова *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: ivisl@paleo.ru
Поступила в редакцию 13.07.2017
Принята к публикации 26.09.2017
Аннотация
Приоритетные глобальный и системный подходы изучения эволюции жизни повышают интерес к интеграции макроэволюционных и экологических исследований и разработке их теоретических основ. В качестве элементарной фундаментальной единицы эволюционирующего палеобиосферного пространства–времени предложен экон. Концепция экона, единства вида и экологической ниши, позволяет решать вопросы эволюции биоразнообразия, эволюционной макроэкологии и биогеографии на пространственно-временных шкалах и в рамках системы “макротаксон–биосфера”. Поэтому особую актуальность приобретают естественные классификации организмов, отражающие подлинную филогению. Целостный подход позволит получать более обоснованные выводы по эволюции и распространению как ископаемых, так и современных организмов.
ВВЕДЕНИЕ
Глобальный и системный подходы в изучении эволюции жизни становятся приоритетными. Они необходимы для дальнейшего познания законов развертывания жизни на Земле и сохранения условий выживания человечества. Огромный массив новой информации неизбежно ведет к пересмотру некоторых базовых положений, касающихся фундаментальных принципов эволюции биоразнообразия и экосистем.
Особую актуальность приобретают изучение макроэволюции (эволюции надвидовых таксонов) и разработка закона целостности (единства) эволюции таксона и среды как теоретической основы эволюционных экологических и биогеографических исследований. Макроэволюцию начинают рассматривать в качестве самостоятельной дисциплины (Jablonski et al., 1997).
Современной наукой становится все более востребованным сочетание макроэволюционного и экологического подходов (Jablonski, 2005; Lieberman et al., 2007; Ricklefs, 2007; Cavender-Bares et al., 2009; Carotenuto et al., 2010, 2015; Losos, 2010; Castiglione et al., 2017; Lima-Ribeiro, Diniz-Filho, 2017; и др.). На смену господствовавшим в науке на рубеже ХХ и XXI вв. филогенетической систематикe (кладистике) В. Хеннига (Hennig, 1966) и факторному анализу динамики биоразнообразия Дж. Сепкоски и его коллег (Sepkoski et al., 1981) приходят новые парадигмы, связанные с новыми методологиями изучения и принципами моделирования эволюционных процессов.
Достоверность новых знаний обеспечивает мультидисциплинарный подход. Ядро такого подхода составляет эволюционная палеонтология, которая сегодня сама является многоплановой наукой, использующей данные разных дисциплин (эволюционной биологии, генетики, молекулярной биологии, экологии, биогеографии и др.) и широкий круг методов. Пионерные исследования макроэволюции продолжают опираться на данные геологии и геохронологии и имеют сильный экологический акцент.
Особый интерес для разработки теории представляют исследования, которые учитывают все аспекты эволюции макротаксонов (морфологические, генетические, таксономические, экологические, биогеографические и др.) и анализируют макроэволюцию и эволюцию сообществ в контексте изменений среды в пространстве-времени (в масштабе геологического времени).
Хорошими модельными группами для изучения основных закономерностей эволюции являются всесторонне изученные высокоорганизованные “кроновые” группы (“crown groups”) позвоночных, в частности, млекопитающие. Данные по млекопитающим всегда служили основой для новых концептуальных понятий (адаптивная радиация, пространство ресурсов и др.) и дисциплин (напр., эволюционная палеонтология, зоогеография). Одной из лучших модельных групп для изучения эволюционных процессов всегда были парнопалые (Artiodactyla) (Ковалевский, 1875; Van Valen, 1971; Вислобокова, 1990, 2006, 2008a, б; Vrba, 1992, 1993; и др.), а в последнее время стали и китопарнопалые (Cetartiodactyla). Эти группы особенно интересны для сопоставления преобразований макроэволюционного уровня, отраженных в фенотипах и геномах (Вислобокова, 2013а, б; McGowen et al., 2014; и др.).
Анализ макроэволюционных процессов с применением системного и эволюционно-экологического подходов позволил четче увидеть основные закономерности, касающиеся единства эволюции таксонов и среды, и начать разработку концепции экона (Вислобокова, 2014, 2016; Vislobokova, 2017).
В этой статье я кратко остановлюсь на двух важных для теории эволюции вопросах: 1) научной значимости (ценности) лишь естественной классификации организмов для воссоздания и анализа эволюции, в том числе ее экологических и биогеографических аспектов, и 2) сути концепции “экон” и ее перспективах для изучения эволюции биоразнообразия, в эволюционной макроэкологии и биогеографии. Эти вопросы существенны для дальнейшей разработки методологии и теоретических построений эволюционной экологии, макроэкологии и эволюционной биогеографии. Ввиду большого объема проанализированного материала, включающего и оригинальные исследования автора по эволюции млекопитающих и их сообществ, в статье отражены лишь важнейшие направления и постулаты исследований.
О КЛАССИФИКАЦИИ ОРГАНИЗМОВ И НЕОБХОДИМОСТИ КОРРЕКЦИИ МЕТОДОЛОГИИ И МЕТОДОВ ФИЛОГЕНЕТИЧЕСКИХ АНАЛИЗОВ
Естественная классификация организмов – основа правильной интерпретации данных разных дисциплин (эволюционной биологии, палеонтологии, генетики, молекулярной биологии, экологии и биогеографии). Не вызывает сомнений, что данные по эволюции организмов и их сообществ могут быть правильно представлены и интерпретированы лишь с позиции естественной классификации организмов. Каждая систематическая единица (таксон) есть реальная сущность, монофилетическая группа соподчиненных естественных единиц, содержащая предковую группу и всех ее потомков. Макроэволюция протекает в пространстве-времени через преобразование видов и отражается в таксономической иерархии надвидового уровня. Макротаксоны и их иерархическое структурирование являются следствием эволюционного преобразования “потоков жизни”, фил (Simpson, 1944, 1961; Teilhard de Chardin, 1955; Jablonski et al., 1997; Vislobokova, 2017; и др.). Создание искусственных классификаций допустимо лишь в исключительных случаях, для слабо изученных групп. Оно ни в коем случае не должно приветствоваться при изучении позвоночных, тем более млекопитающих, с их хорошо изученной морфологией и экологией (в том числе, на ископаемых материалах) и пополняющимися данными по эволюции геномов и филогеномике.
Реальные надвидовые таксоны способна выявлять пока лишь систематика, основанная на данных классической филогении и новых данных филогеномики. Широко применяемая кладистика, или филетическая таксономия, очень часто становится источником не реальных, а формальных таксонов.
Вопросам анализа современного состояния кладистики и ее недостатков посвящен ряд отдельных публикаций, в том числе отечественных (Павлинов, 1990, 2005; Расницын, 2006; Зеленков, 2012). Поэтому я остановлюсь лишь на некоторых важных моментах.
Неудачным упрощением кладистики следует признать принцип узкой дихотомии (бифуркации) узловых событий, в которую трансформировалась концепция дивергенции признаков и процесса диверсификации эволюции Ч. Дарвина. Строгая дихотомия не отвечает реалиям, установленным на ископаемых материалах. Другой важный недостаток связан с упрощенным (формальным) подходом определения иерархии таксонов. В кладистике сестринские группы (виды, роды, семейства и др.) одного уровня рассматриваются в одном систематическом ранге, ранг таксона и степень родства определяются по последовательности обособления клад, при этом учитываются только синапоморфии. Таксономическая и эволюционная оценка веса признаков часто отсутствует, а для определения статуса признака (плезиоморфный или апоморфный и его полярности) нередко используются некорректные внешние группы. Хотя иногда предполагают, что проводится анализ взаимосвязанных элементов (модусов), но он таковым в большинстве случаев не является. В результате даже при применении парсимонийного анализа схема последовательности кладистических событий остается лишь очень приблизительной картиной подлинного филогенеза группы (одной из “вероятностных” гипотез ее филогенетических связей). Слабой стороной метода является и относительность понятия “время”: время оценивается как эволюционная дистанция признаков (часто недостаточно корректных). Очевидно, что для получения более достоверной картины родства и иерархии таксонов, филогенетический анализ должен основываться на глубоком знании детальной морфологии и эволюции групп, выявлении блоков признаков, имеющих разное таксономическое значение, учете реального геологического времени и реальных экологических и биогеографических процессов. Концепция экона позволит более точно реконструировать направления эволюции таксонов.
Следует подчеркнуть, что широко распространившееся за рубежом с конца ХХ в. выделение искусственных таксономических категорий и искусственных классификаций, появляющихся в результате компьютерной обработки данных с использованием программ-дериватов концепции Хеннига, не отвечает запросам современной науки. Кладистика постепенно теряет свою “парадигмическую” значимость. В научном мире растет понимание того, что формальная кладистика ведет к искажению реальной картины исторического развития “живого мира” планеты.
В свое время внедрение кладистических методов вызвало справедливую критику со стороны таких теоретиков-корифеев эволюционной биологии, как Дж. Симпсон (Simpson, 1980), Э. Майр (Mayr, 1974, 1982, 1988), Л. Ван Вален (Van Valen, 1978). Не стали приверженцами этих “инновационных” методов многие крупные эволюционисты, изучающие позвоночных (М.Ф. Ивахненко, М.А. Шишкин, S.D. Webb и др.). К сожалению, кладизм, порождающий искусственные таксоны и связи, находит все больше адептов в нашей стране и, более того, такие исследования, к сожалению, нередко рассматриваются как приоритетные.
Что положительного привнесла кладистика, кроме возросшего интереса к морфологическим исследованиям? Ее несомненной заслугой является то, что она сохранила рациональное зерно, воспринятое от дарвинизма: (1) клады – монофилетические группы; (2) нестинг клад и их иерархическая организация (расположение). Кроме того, некоторые последователи этого направления сегодня пытаются учитывать и экологические факторы (см. ниже).
О необходимости новых компьютерных программ для филогенетических реконструкций. Искусственные таксоны бесполезны не только для систематика, являющегося приверженцем естественной классификации организмов. Они бесполезны (более того, вредны) для уточнения закономерностей эволюции жизни, а также для целей эволюционной экологии и биогеографии. На основе искусственных таксонов невозможно прослеживать тенденции эволюции биоты для выстраивания глобального мониторинга и организации действий по сохранению экосистем Земли. Современные компьютерные программы филогенетических анализов, по которым уточняется систематика (классификация) организмов, требуют совершенствования. Существующие программы кладистических анализов способны воссоздавать более или менее реальную картину родства лишь в пределах родов и семейств. Чрезмерно упрощенные кладограммы, основанные на апоморфных признаках, даже непрестанно возрастающих наборах признаков и парсимонийном анализе дают быстрый, но сомнительный результат. Их достоинство лишь в малых затратах интеллектуальных способностей и времени исследователя. Результат кладистического анализа, как правило, не отражает реальные родственные связи и может рассматриваться лишь как первое приближение к последующему глубокому изучению анализируемой группы или групп организмов. Несомненно, компьютерная обработка данных – прогрессивное направление, и оно должно развиваться. Необходимо совершенствовать программы, чтобы они лучше отражали иерархию таксонов и их родство на основе детального изучения разных аспектов эволюции организмов, их фенотипов и генотипов, и с большим эволюционным смыслом применяли принцип сортинга и нестинга. Уже сегодня необходимы более сложные программы, учитывающие принципы классической систематики и филогении и экологические аспекты эволюции.
О КОНЦЕПЦИИ ЭКОНА И ЕЕ ПЕРСПЕКТИВАХ В ЭВОЛЮЦИОННОЙ МАКРОЭКОЛОГИИ И БИОГЕОГРАФИИ
Для воссоздания истории жизни на Земле и уточнения закономерностей этого процесса, наряду с выявлением эволюционных линий на основе классического изучения филогении и новейших данных филогеномики, особый интерес представляет анализ эволюционной экологии и биогеографии. Сочетание этих дисциплин позволяет реконструировать реальную картину развития жизни на Земле и ее отдельных фрагментов.
Новые данные по эволюции разных групп организмов и все возрастающая точность привязки событий прошлого к геохронологической шкале способствуют все более глубокому пониманию целостности (единства) эволюции таксона и среды, единства генетических и эпигенетических процессов, в том числе на уровне макротаксонов.
Неразрывность взаимосвязи эволюции организмов и среды их обитания была сформулирована Дарвином (Darwin, 1859) как закон единства типа и условий существования. Особенности этой взаимосвязи продолжают интенсивно изучаться. Последовательное углубление знаний о взаимосвязях эволюции видов и их среды отразилось, в частности, в совершенствовании концепций “экологическая ниша” и “адаптивная зона”, введении понятия “экон”, активной разработке концепции конструирования ниши.
Краткий обзор основных понятий и концепций, предваряющих концепцию экона. Под термином “экологическая ниша” обычно понимают совокупность всех факторов среды (внутренних и внешних, биотических и абиотических), в пределах которых возможно существование вида в природе. Ниша – не только условия обитания вида, но и среда, где действует отбор. Подробные обзоры существующих трактовок понятия “экологическая ниша” можно найти в ряде работ (Chase, Leibold, 2003; Meszéna, Hendry, 2012; Pocheville, 2015). Ниже будут приведены лишь представления, которые являются наиболее значимыми для концепции экона.
Дж. Гриннелл (Grinnell, 1917) применил термин “ниша” для обозначения функциональной роли и положения вида в сообществе и один из первых указал на значение взаимосвязи вида и его ниши в формировании биома.
Понятие “экологическая фундаментальная ниша”, введенное Дж. Хатчинсоном (Hutchinson, 1957), определило направление многих современных исследований. Оно означает часть воображаемого многомерного пространства (гиперобъема), отдельные точки которого соответствуют состоянию среды (физических и биотических факторов), необходимому для нормального существования вида. В природе фундаментальной нише соответствует реализованная ниша. Для ее изучения широко применяется моделирование, в том числе мультишкальное и мультивекторное. Для сравнения видов достаточно нескольких операционально значимых признаков их ниш (MacArthur, 1968, 1972). Видовая специфичность ниши подразумевается и принципом Вольтерра-Гаусса (см. Hutchinson, 1957).
Термин “экон” был предложен Г. Хитвоулом (Heatwole, 1989) в качестве фундаментальной экологической единицы для вида, его функциональной сущности в нише. Термин используется и для разных компонентов вида (стадий онтогенетического и исторического развития, стиля жизни и др.), занимающих самостоятельные ниши.
Единство взаимодействий организма и среды является фундаментальным свойством эволюции (Lewontin, 1983). Активно развивающаяся, в основном на популяционном уровне, концепция конструирования ниши подчеркивает значение изменений этих взаимодействий как для конструктора (носителя) ниши, так и для среды, включая реципиентов из других ниш (см. Laland et al., 2016).
Вид и его ниша составляют эволюционирующее целое. Ниша вида включает в себя не только воздействия среды, но и воздействия вида на среду. Эти воздействия отражаются в адаптации к разнообразным условиям (окружающей среде, условиям обитания, ресурсам). Следует подчеркнуть, что виды не внедряются в экологические ниши и не переходят в другие ниши (как иногда предполагается), а меняются вместе с нишей. Нишевые взаимодействия вида трансформируются в соответствии с условиями среды и меняют их, корректируя направления своей эволюции и воздействуя на изменения среды.
Понятие “экологическая ниша” широко используется при анализе основных направлений макроэволюции и их факторов (анализ разных векторов ниши), видового сортинга (влияние ниш на сборку сообщества), моделировании (Kembel, 2009; и др.). Наиболее часто изучаемые градиенты окружающей среды (ключевые оси ниши): дивергенции по ресурсам и по пространственным дистанциям, географический контекст и др. Неточность толкования этого понятия (например, допущение освоения новой ниши, перехода из одной ниши в другую) предсказуемо ведет к неверной трактовке некоторых эволюционных процессов.
Понятие “адаптивная зона”, более широкое, чем “ниша”, было введено Симпсоном (Simpson, 1944) как комплекс экологических условий, представляющих собой возможную среду обитания для различных животных и растений и определяющих направление адаптаций надвидовых таксонов. Симпсон близко подошел к пониманию явлений, которые нашли отражение в концепциях ароморфоза и алломорфоза (идиоадаптации) выдающихся российских эволюционистов А.Н. Северцова (1925) и И.И. Шмальгаузена (1940), а также механизма основных направлений эволюции – теории стабилизирующего отбора Шмальгаузена (1941), одной из основ современной эпигенетической теории эволюции. Так, Симпсон полагал, что филетическая эволюция – эволюция эволюционных линий – длительный, направленный сдвиг средних значений.
По Симпсону, границами адаптивной зоны для конкретной группы организмов служат крайние варианты физико-географических и биотических условий (хищники, паразиты), при которых представители данной группы еще могут существовать. Эволюционируют не только животные, но и занимаемые ими адаптивные зоны, каждая из которых определяется связью населяющих ее организмов с условиями среды. Для схематического графического изображения адаптивных зон Симпсон предложил адаптивную решетку и показал положение на ней трех выделяемых им форм эволюции: видообразование (для низкого таксономического уровня, подвид, вид, род и т.д.; локальная адаптация и случайное расхождение признаков, субзональное по положению в адаптивной решетке), филетическая эволюция (для среднего – роды, подсемейства, семейства; постадаптация и веками длящаяся адаптация, зональная) и квантовую (для высокого – семейства, подотряды, отряды и т.д.; преадаптация, межзональная) (см. Simpson, 1944; Симпсон, 1949).
Более популярна трактовка “адаптивной зоны” Ван Валена (Van Valen, 1971) как ниши любого таксона (прежде всего, надвидового), части окружающей среды или пространства ресурсов, которое существует независимо от использующего его таксона (например, водная среда как адаптивная зона и т.д.). Ван Вален развил понятие “адаптивная зона” для высших таксонов, опираясь на ключевые адаптации млекопитающих на уровне отрядов. Предложенное им более широкое понятие “сфера жизни” включало и конкурентные взаимодействия. Он выдвинул гипотезу, что происхождение многих высших таксонов сопровождалось важными изменениями в их сфере жизни, которые происходили при переходе между высшими таксонами.
Понятие “адаптивная зона” продолжает использоваться для обозначения среды, с которой связана адаптивная радиация какой-либо группы организмов.
Новая концепция экона. Современный уровень знаний позволяет анализировать не абстрактное пространство адаптивной зоны, а реальное эволюционирующее экопространство взаимосвязи таксонов и среды. Понятие “экон” – вид и его экологическая ниша – было предложено мной в качестве элементарной динамической единицы эволюционирующего палеобиосферного (экосистемного) пространства (Вислобокова, 2014, 2016; Vislobokova, 2017). Экон вида – “атом” этой системы (рис. 1). Экон включает в себя две составляющие: (1) таксон как целостная биосистема и (2) сумма взаимосвязей таксона с палеобиосферным пространством (которые затрагивают все уровни организации живого). Экон может рассматриваться как несущий информацию участок, или определенный объем, эволюционирующей биосферы (экосистемы). Он несет информацию о таксоне, с его историческим прошлым и возможным будущим, детерминированным и направленным в русле продолжающегося развития планеты и ее биосферы. При анализе можно ограничиваться необходимыми для исследования параметрами.
Рис. 1.
Простейшая cхема пространственно-временной эволюции эконов и биосферы. Обозначениея: e – экон; В – биосфера; Е – экосистема. Эконы изображены точками; линиями показаны направления их эволюции, дифференциации и сегрегации. Плоскости B1, B2, B3 отражают последовательные этапы эволюции биосферы, петли Е1, E2, E3 – соответствующие этим этапам экосистемы, а дефисы – разнообразие экосистем каждого этапа.
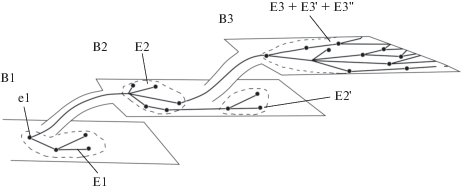
Концепция экона не подменяет и не отменяет понятий “экологическая ниша” и “адаптивная зона”, а более точно отражает принцип целостности эволюционного развития таксона (организмов), единство эволюции таксона и среды. В ходе эволюции биосферы взаимодействия между эконами разных видов становятся все более эффективными и гармоничными. Хорошо известно, что в условиях сформированных сообществ конкурентные отношения между видами и борьба за существование ослабляются.
Концепция экона отражает целостность таксона и среды в ее динамике и важна как для решения теоретических вопросов, так и для моделирования процессов макроэволюции (адаптациогенеза и экогенеза) и эволюции биосферы и ее экосистем.
О некоторых перспективах концепции экона для изучения эволюции биоразнообразия, в эволюционной макроэкологии и биогеографии. Последние десятилетия многие исследования по эволюции биоразнообразия выполнялись в русле факторного анализа динамики макротаксонов: изменений уровня таксономического разнообразия (богатства), его подъемов и падений, динамики происхождения и вымирания таксонов, эффектов массового вымирания на шкале геологического времени (Sepkoski, 1997; Jablonski, 2001; и др.). Они позволили реконструировать кривые биотических событий и увязать их с изменениями на геологических рубежах. Интересные результаты по корреляции изменений таксономического разнообразия с глобальными изменениями климата дал анализ наземных млекопитающих на основе базы данных NOW (Fortelius et al., 2006). Связи и таксономические особенности эволюции сообществ на больших территориях хорошо выявляет кластерный анализ. Такой анализ был успешно применен для фаун млекопитающих неогена и плейстоцена Европы (Palombo et al., 2006). При изучении эволюции биоразнообразия все чаще используются данные филогеномики, проводится оценка морфологической и генетической дистанции между видами в пределах макротаксонов или между макротаксонами, уделяется внимание изучению взаимодействий фенотипа с физической и биотической окружающей средой, динамике сортинга и другим сторонам эволюции (Valentine, Jablonski, 2003; и др.). Возросло понимание необходимости интеграция информации по ископаемым и современным видам и разнообразию фенотипов, геномов, их реакции на окружающую среду.
Однако теоретическая проработка массива информации остается недостаточной. Приходится констатировать, что теоретические исследования значительно отстают от эмпирических. При этом фактор целостности эволюции таксона и его экологической ниши все еще недооценивается. Примечательно, что совсем недавно теоретиков призывали уделить больше внимания такому базовому вопросу, какой тип нишевой оси, скорее всего, является наиболее важным в стимулировании диверсификации (Meszéna, Hendry, 2012). Такая постановка вопроса далека от целостного подхода.
Отставание теории порождает много вопросов, в том числе касающихся механизмов формирования разнообразных и стабильных сообществ. Необходимость целостного подхода и всестороннего анализа становится актуальной как при изучении фенотипов, так и сообществ. Концепция экона отвечает этим вызовам и может стать востребованной. Эволюционная экология сообществ и экосистем может рассматриваться как интегрированное взаимодействие эконов, а эволюционная биогеография как изменения распространения эконов и экосистем в палеобиосферном пространстве-времени (эволюции биосферы на шкале геологического времени).
Сегодня перспективным направлением исследований является эволюционная макроэкология, которая представляет особый интерес как наука на стыке макроэволюционных исследований и экологии (Raia, Fortelius, 2017). Эволюционная макроэкология изучает особенности распределения видов, изменение разнообразия в пространстве и времени, его причины и закономерности, а также анализирует то, как способы жизни (моды), эволюционные особенности (фенотипическая эволюция, изменения размеров тела и др.), тренды и климатические изменения влияют на распространение (Brown, Mauer, 1989; Brown, 1995; Blackburn, Gaston, 2003; Rabosky, 2009; Raia, Fortelius, 2017). В русле этого направления анализируются разные вопросы: диверсификация и сортинг клад, конкуренция между кладами, специфика адаптаций и дифференциаций, влияние направленных изменений на диверсификацию, эволюция и филогенетика сообществ, особенности экологии метасообществ и др. (Hunt, 2007; Pearse et al., 2014; Hubert et al., 2015; Hunt et al., 2015; Silvestro et al., 2017; и др.). Разрабатываются новые модели и методы, основанные на филогенетических деревьях и кладограммах для выяснения истории клад и их особенностей. Для анализа процессов эволюции стало использоваться понятие “экопространство”, которое объемлет все: от биома до биосферы (Lidicker, 2008). Оно, несомненно, полезно. Но чтобы не уйти в область искусственных построений, не стоит отказываться от реальных понятий: биосфера и др.
Интересные результаты по сортингу признаков между кладами получили П. Полли и его коллеги (Polly et al., 2017). Они предполагают сильное влияние сортинга на распределение направлений развития сообществ. Комбинаторика, впервые выявленная Н. И. Вавиловым, по-видимому, играла важную роль на разных уровнях жизни, в том числе и биоценотическом.
Данные по ископаемым организмам показывают, что изменение таксономического разнообразия и эволюция сообществ хорошо коррелирует с экологическими модами и изменениями окружающей среды (Вислобокова, 2008а, б, 2013б; Sahney et al., 2010; Vislobokova, 2012; и др.). Разнообразие – результат эволюционного развития линий и отбора. Аналитические исследования показывают, что экологическое разнообразие наземных позвоночных больше зависит от расширения и воздействия экопространства, чем от прямой конкуренции внутри него (Sahney et al., 2010). Этот вывод основан на анализе ключевых адаптаций инвазии клад в экопространство, перехода к новому образу жизни. Анализ эволюции эконов, а не “инвазии” клад, даст более точную картину развития жизни.
Системный подход позволил прийти к выводу, что основным направляющим стимулятором (драйвером) макроэволюции служит биосфера, система взаимосвязей “макротаксон–биосфера” (Vislobokova, 2017). Элементарным звеном эволюции этой системы является экон.
Концепция экона хорошо соответствует целостности (единству) эволюции организмов и среды. Она позволяет лучше видеть единство эволюции таксона и среды, закономерности макроэволюции в пространстве-времени. Макроэволюция предстает как авторегуляторный процесс, связанный с перестройкой, дифференциацией, сортингом и нестингом эконов в палеобиосферном пространстве, эволюция которого направляется развитием планеты и ее биоты. Процесс канализируется внутренними и внешними факторами. Усиление взаимосвязей между таксонами в ходе эволюции биосферы предстает как наложение и взаимная корреляция (подгонка) эконов, близких по экологии, в биосферном пространстве–времени. Формирование сообществ и зональное распределение определяются эволюционным комбинаторным распределением (сортингом и нестингом) эконов в эволюционирующих экосистемах в соответствии с меняющимися параметрами биосферы (градиентом температур, природной зональностью и др.). Анализ может проводиться в рамках двух динамических систем: видообразование → макроэволюция; экон → → экосистема (биогеоценоз) → биосфера.
Концепция экона хорошо сочетается с классической биогеографией. В биогеографических исследованиях учитывается внутренняя и внешняя целостность вида (в том числе, репродуктивно-генетическая и эпигенетическая, обособленность генофонда), определяемая его эволюционным развитием, взаимодействиями со средой (см. Дзунино, Дзуллини, 2010). Целостность вида выступает как единство, поддерживаемое в определенных пространственно-временных границах.
Эволюционная биогеография позволяет анализировать расселение таксонов в эволюционирующем пространственно-временном континууме биосферы (ареал–биота–экосистема–биом–биосфера). Понятие “экон” будет полезно при реконструкции картин расселения (миграция, изоляция и др.), а также лучшего понимания некоторых эффектов и связей, которые существуют между отдельными видами или их комплексами.
Понятие экон хорошо согласуется с понятием ареал. Ареал – динамическая структура, часть географического пространства, в которой этот вид присутствует и взаимодействует с окружающей средой в пространстве-времени. Согласно современным представлениям, ареал макротаксона является не простой суммой ареалов ее составляющих, а реальной единицей, подчиненные элементы которой имеют связи родственного типа; ареал надвидового таксона отражает большую толерантность организмов, чем у отдельного его вида (см. Дзунино, Дзуллини, 2010). Ареал – это, по сути, географическая (пространственная) проекция экона.
Концепция экона может изменить трактовку некоторых событий эволюционной биогеографии, изменений ареалов и расселения организмов в масштабе геологического времени, а также некоторые взгляды на современное распределение организмов. Так, сегодня в биогеографии продолжает довлеть аксиома “центр происхождения-расселения”, согласно которой, зарождаясь на какой-либо ограниченной территории, вид начинает расселяться до тех пор, пока факторы среды (ландшафтные, климатические, биотические) не ограничат его распространение. Образование новых видов и надвидовых таксонов обусловлено, по мнению многих специалистов, главным образом, ситуацией, когда переселяющиеся из центра происхождения формы адаптируются к новым условиям. Расселение зависит от многих факторов (ландшафтно-климатических изменений, эволюции широтной и высотной зональности, барьеров, изоляции и др.). Эконы новых видов адаптивных направлений возникали там, где для этого существовали предпосылки, и могли иметь очень широкое площадное распространение. Подобным образом, в частности, в раннем олигоцене Центральной Азии возникли высшие жвачные. Для возникновения новых эконов необходимы, как минимум, достаточная генетическая и эпигенетическая пластичность предкового вида и наличие у него преадаптаций, соответствующих преобразованиям среды.
В качестве другого примера можно привести эффект экваториальной помпы Ф. Дарлингтона (Darlington, 1957). Он предполагает, что активное формообразование происходит в тропических экосистемах, и новые таксоны вытесняют прежние, в частности, в более высокие широты. Этот эффект может быть объяснен смещением эконов к северу из-за глобальных трансформаций природных зон и экосистем. Такие смещения хорошо прослеживаются в эволюции кайнозойских млекопитающих северного полушария на фоне глобального тренда к похолоданию. Архаичные виды многих млекопитающих чаще приурочены к тропикам и субтропикам. Распределение парнопалых в биоте хорошо совпадает с эволюцией ландшафтно-климатической зональности, а их современные архаичные виды чаще приурочены к тропикам и субтропикам.
Приуроченность эконов к определенным ландшафтно-климатическим условиям и их изменение в ходе геологической истории Земли может лучше объяснить многие явления, наблюдаемые в современной биоте. В частности, становятся более понятными такие явления, как меньшая устойчивость вида у границы ареала, мобильность ареалов (их пульсация во времени), колебания равнинно-горного распределения видов в плейстоцене.
Концепция экона определенно потребует пересмотра современных основ филогенетической биогеографии, тесно связанной с филогенетическим кладистическим анализом и оперирующей “апохориями” (по аналогии с апоморфиями). Окажутся под сомнением и некоторые выводы викариантной биогеографии, в которой принцип случайности распространения соединен с филогенетическим анализом формального кладистического типа.
ЗАКЛЮЧЕНИЕ
Интеграция макроэволюционных, экологических и биогеографических исследований сегодня является одним из актуальных направлений изучения эволюции жизни. Необходимость корректного анализа все возрастающего массива эмпирических данных по эволюции макротаксонов, биоразнообразию, макроэкологии и биогеографии требует новых теоретических разработок и совершенствования методологии исследований. Эмпирической основой таких исследований могут быть лишь реальные таксоны. Использование искусственных таксонов (продуктов формальной кладистики) снижает научную ценность реконструкций эволюции биоты прошлого и теоретических выводов.
Концепция экона (вид и его экологическая ниша) может стать важным инструментом при анализе экологических и биогеографических аспектов эволюции макротаксонов. Эта концепция позволяет рассматривать эволюцию таксонов в их единстве с меняющейся средой и лучше видеть “стрелу времени”, направленность эволюции жизни на фоне геологической истории планеты.
Эволюционная экология сообществ и экосистем может анализироваться как интегрированное взаимодействие эконов и их сортинг в соответствии с градиентами среды в палеобиосферном пространстве–времени. В эволюционной биогеографии изменение распространения таксонов и сообществ может анализироваться как пространственная динамика эконов (расширение, смещение, наложение и др.) во времени, а ареалы таксонов рассматриваться как географические (пространственные) срезы эконов.
Концепция экона, постулирующая целостность эволюции макротаксона и его среды, стимулирует более целостный подход к ее изучению и обеспечивает получение более обоснованных результатов по эволюции и распространению как ископаемых, так и современных организмов.
Список литературы
Вислобокова И.А. Ископаемые олени Евразии. М.: Наука, 1990. 208 c. (Тр. ПИН АН СССР. Т. 240).
Вислобокова И.А. Историческое развитие парнопалых (Artiodactyla) Северной Евразии и этапы эволюции их сообществ в кайнозое // Эволюция биосферы и биоразнообразия. М.: Тов-во научн. изданий КМК, 2006. С. 416–438.
Вислобокова И.А. Основные этапы эволюции сообществ парнопалых Северной Евразии в плиоцене–начале среднего плейстоцена. Часть 1 // Палеонтол. журн. 2008а. № 3. С. 76–91.
Вислобокова И.А. Основные этапы эволюции сообществ парнопалых Северной Евразии в плиоцене – начале среднего плейстоцена. Часть 2 // Палеонтол. журн. 2008б. № 4. С. 79–89.
Вислобокова И.А. О происхождении Cetartiodactyla: сравнение данных эволюционной морфологии и молекулярной биологии // Палеонтол. журн. 2013а. № 3. С. 83–97.
Вислобокова И.А. Экологическая эволюция ранних Cetartiodactyla и реконструкция ее пропущенного начального звена // Палеонтол. журн. 2013б. № 5. С. 72–88.
Вислобокова И.А. Эволюция биосферы и макроэволюция. М.: ГЕОС, 2014. 168 с.
Вислобокова И.А. Макроэволюция как системный процесс развития жизни // Палеонтология. Стратиграфия. Астробиология. К 80-летию акад. А.Ю. Розанова. М.: ПИН РАН, 2016. С. 137–165.
Дзунино М., Дзуллини А. Биогеография (эволюционные аспекты). М.: Изд-во МГУ, 2010. 317 с.
Зеленков Н.В. Кладистический анализ, эволюция и палеонтология // Современная палеонтология: классические и новейшие методы – 2012. М.: ПИН РАН, 2012. С. 9–25.
Ковалевский В.О. Остеология двух ископаемых видов из группы копытных Entelodon и Gelocus // Изв. Имп. об-ва любит. естествознан. антропол. этногр. 1875. Т. 16. С. 1–59.
Павлинов И.Я. Кладистический анализ (методологические проблемы). М.: Изд-во МГУ, 1990. 160 с.
Павлинов И.Я. Введение в современную филогенетику (кладогенетический аспект). М.: КМК, 2005. 391 с.
Расницын А.П. Летопись и кладограмма // Эволюция биосферы и биоразнообразия. М.: КМК, 2006. С. 39–48.
Северцов А.Н. Главные направления эволюционного процесса. М.: Изд-во Думнова, 1925. 153 с.
Симпсон Дж. Г. Темы и формы эволюции. М.: ИЛ, 1949. 358 с.
Шмальгаузен И.И. Пути и закономерности эволюционного процесса. М.; Л.: Изд-во АН СССР, 1940. 231 с.
Шмальгаузен И.И. Стабилизирующий отбор и его место среди факторов эволюции // Журн. общ. биол. 1941. Т. 2. № 3. Р. 307–354.
Blackburn T.M., Gaston K.J. Macroecology: concepts and consequences. Oxford: Blackwell, 2003. 442 p.
Brown J.H. Macroecology. Chicago: Univ. Chicago Press, 1995. 269 p.
Brown J.H., Maurer B.A. Macroecology: the division of food and space among species on continents // Science. 1989. V. 243. P. 1145–1150.
Carotenuto F., Barbera C., Raia P. Occupancy, range size, and phylogeny in Eurasian Pliocene to Recent large mammals // Paleobiology. 2010. V. 36. P. 399–414.
Carotenuto F., Diniz-Filho J.A.F., Raia P. Space and time: the two dimensions of Artiodactyla body mass evolution // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2015. V. 437. P. 18–25.
Castiglione S., Mondanaro A., Carotenuto F. et al. The many shapes of diversity: ecological and evolutionary determinants of biodiversity through time // Evol. Ecol. Res. 2017. V. 18. P. 25–39.
Cavender-Bares J., Kozak K.H., Fine P.V.A., Kembel S.W. The merging of community ecology and phylogenetic biology // Ecol. Lett. 2009. V. 12. P. 693–715.
Chase J.M., Leibold M.A. Ecological niches: Linking classical and contemporary approaches. Chicago: Univ. Chicago Press, 2003. 216 p.
Darwin Ch. The origin of species by means of natural selection or the preservation of favoured races in the struggle for life. L.: J. Murray, 1859. 490 p.
Darlington P.J. Zoogeography: the geographic distribution of animals. Cambridge: Harvard Univ. Press, 1957. 236 p.
Fortelius M., Eronen J., Liu L. et al. Late Miocene and early Pliocene large land mammals and climatic changes in Eurasia // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2006. V. 238. P. 219–227.
Grinnell J. The niche relationships of the California thrasher // Auk. 1917. V. 34. P. 427–433.
Heatwole H. The concept of the econe, a fundamental ecological unit // Trop. Ecol. 1989. V. 25. № 1. P. 13–19.
Hennig W. Phylogenetic systematics. Urbana: Illinois Univ. Press, 1966. 501 p.
Hubert N., Calcagno V., Ettiene S., Mouquet N. Metacommunity speciation models and their implications for diversification theory // Ecol. Lett. 2015. V. 18. P. 864–881.
Hunt G. The relative importance of directional change, random walks, and stasis in the evolution of fossil lineages // Proc. Natl. Acad. Sci. USA. 2007. V. 104. P. 18404–18408.
Hunt G., Hopkins M.J., Lidgard S. Simple versus complex models of trait evolution and stasis as a response to environmental change // Proc. Natl. Acad. Sci. USA. 2015. V. 112. P. 4885–4890.
Hutchinson G.E. Concluding remarks // Cold Spring Harbor Sym. Quant. Biol. 1957. V. 22. P. 415–427.
Jablonski D. Lessons from the past: Evolutionary impacts of mass extinctions // Proc. Natl. Acad. Sci. USA. 2001. V. 98. P. 5393–5398.
Jablonski D. Evolutionary innovations in the fossil record: the intersection of ecology, development, and macroevolution // J. Exp. Zool. 2005. V. 304B. P. 504–519.
Jablonski D., Benton M.J., Gastaldo R.A. et al. Macroevolution in the 21st Century // Paleontology in the 21st Century Workshop / Eds. Lane H. R., Lipps J., Steininger F.F., Ziegler W. Kleine Senckenbergreihe. 1997. V. 25. P. 111–119.
Kembel S.W. Disentangling niche and neutral influences on community assembly: assessing the performance of community phylogenetic structure tests // Ecol. Lett. 2009. V. 12. P. 949–960.
Laland K., Matthews B., Feldman M.W. An introduction to niche construction theory // Evol. Ecol. 2016. V. 30. P. 191–202.
Lewontin R.C. The organism as the subject and object of evolution // Scientia. 1983. V. 118. P. 63–82.
Lidicker W.Z. Levels of organization in biology // Biol. Rev. 2008. V. 83. P. 71–78.
Lieberman B.S., Miller W. III, Eldredge N. Paleontological patterns, macroecological dynamics and the evolutionary process // Evol. Biol. 2007. V. 34. P. 28–48.
Lima-Ribeiro M.S., Diniz-Filho J.A.F. Climate change, human overkill, and the extinction of megafauna: a macroecological approach based on pattern-oriented modeling // Evol. Ecol. Res. 2017. V. 18. P. 97–121.
Losos J.B. Adaptive radiation, ecological opportunity, and evolutionary determinism // Amer. Natur. 2010. V. 175. P. 623–639.
Mayr E. Cladistic analysis or cladistics classification // Z. Zool. Syst. Evol. Forsch. 1974. Bd 12. H. 2. S. 94–128.
Mayr E. The growth of biological thought: diversity, evolution, and inheritance. Cambridge: Belknap Press, 1982. 974 p.
Mayr E. Towards a new philosophy of biology: observation of an evolutionist. Cambridge: Belknap Press, 1988. 564 p.
MacArthur R. The theory of the niche // Population biology and evolution / Ed. R.C. Lewontin. Syracuse: Syracuse Univ. Press, 1968. P. 159–176.
MacArthur R. Geographical ecology: patterns in the distribution of species. N.Y.: Harper and Row, 1972. 269 p.
McGowen M.R., Gatesy J., Wildman D.E. Molecular evolution tracks macroevolutionary transition in Cetacea // Trends Ecol. Evol. 2014. V. 29. P. 336–346.
Meszéna G., Hendry A.P. Introduction to niche theory and speciation // Evol. Ecol. Res. 2012. V. 14. P. 361–363.
Pearse W.D., Purvis A., Cavender-Bares J., Helmus M.R. Metrics and models of community phylogenetics // Modern phylogenetic comparative methods and their application in evolutionary biology / Ed. L.Z. Garamszegi. Berlin: Springer, 2014. P. 451–464.
Palombo M.R., Valli A., Kostopoulos D.S. et al. Similarity relationships between the Pliocene to Middle Pleistocene large mammal faunas of Southern Europe from Spain to the Balkans and the North Pontic Region // Cour. Forsch. Senckenb. 2006. Bd 256. P. 329–347.
Pocheville A. The ecological niche: history and recent controvercy // Handbook of evolutionary thinking in the sciences / Eds. Heams T., Huneman P., Lecointre G., Silberstein M. Dordrecht: Springer, 2015. P. 547–586.
Polly P.D., Fuentes-Gonzalez J., Lawing A.M. et al. Clade sorting has a greater effect than local adaptation on ecometric patterns in Carnivora // Evol. Ecol. Res. 2017. V. 18. P. 61–95.
Rabosky D.L. Ecological limits on clade diversification in higher taxa // Amer. Natur. 2009. V. 173. P. 662–674.
Raia P., Fortelius M. Evolutionary macroecology // Evol. Ecol. Res. 2017. V. 18. P. 1–6.
Ricklefs R.E. History and diversity: explorations at the intersection of ecology and evolution // Amer. Natur. 2007. V. 170. P. S56–S70.
Sahney S., Benton M.J., Ferry P.A. Links between global taxonomic diversity, ecological diversity and the expansion of vertebrates on lands // Biol. Lett. 2010. V. 6. № 4. P. 544–547.
Sepkoski J.J. Biodiversity: past, present, and future // J. Paleontol. 1997. V. 71. P. 533–539.
Sepkoski J.J., Bambach R.K., Raup D.M., Valentine J.W. Phanerozoic marine diversity and the fossil record // Nature. 1981. V. 293. P. 435–437.
Silvestro D., Pires M.M., Quental T.B., Salamin N. Bayesian estimation of multipleclade competition from fossil data // Evol. Ecol. Res. 2017. V. 18. P. 41–59.
Simpson G.G. Tempo and mode in evolution. N.Y.: Columbia Univ. Press, 1944. 237 p.
Simpson G.G. The principle of animal taxonomy. N.Y.: Columbia Univ. Press, 1961. 247 p.
Simpson G.G. Why and how: some problems and methods in historical biology. N.Y.: Pergamon Press, 1980. 263 p.
Teilhard de Chardin P. Le phénoméne humain. P.: Du Seuil, 1955. 318 p.
Valentine J.W., Jablonski D. Morphological and developmental macroevolution: a paleontological perspective // Int. J. Develop. Biol. 2003. V. 47. № 7–8. P. 517–522.
Van Valen L. Adaptive zones and the orders of mammals // Evolution. 1971. V. 25. № 2. P. 120–128.
Van Valen L. Why not to be a cladist // Evol. Theory. 1978. V. 3. P. 285–294.
Vislobokova I.A. Giant deer: origin, evolution, role in the biosphere // Paleontol. J. 2012. V. 46. № 7. P. 643–775.
Vislobokova I.A. The concept of macroevolution in the light of modern data // Paleontol. J. 2017. V. 51. № 8. P. 799–898.
Vrba E.S. Mammals as a key to evolutionary theory // J. Mammal. 1992. V. 73. P. 1–28.
Vrba E.S. Turnover pulses, the Red Queen, and related topics // Amer. J. Sci. 1993. V. 293. P. 418–452.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал