

Палеонтологический журнал, 2019, № 2, стр. 84-93
Систематическое положение Palaeortyx (Aves, ?Phasianidae) и замечания по эволюции Phasianidae
Н. В. Зеленков *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: nzelen@paleo.ru
Поступила в редакцию 04.06.2018
После доработки 05.07.2018
Принята к публикации 16.07.2018
Аннотация
Проанализирована морфология посткраниального скелета олигоцен-миоценовых ископаемых курообразных из рода Palaeortyx в сравнении с современными представителями Phasianoidea. В частности, проведено сравнение с современным африканским родом Ptilopachus, недавно отнесенным к американскому семейству Odontophoridae, в которое первоначально помещали Palaeortyx. Показано, что у Palaeortyx отсутствуют продвинутые признаки Odontophoridae, а все отмеченное сходство (включая глубокую fossa pneumotricipitalis dorsalis плечевой кости) представляет плезиоморфии Odontophoridae+Phasianidae или более высокой клады. Существенного сходства с Ptilopachus также не обнаружено. Наибольшее сходство отмечается между Palaeortyx и Rollulinae, однако оно трактуется как плезиоморфное для Phasianidae s.s. Таким образом, Palaeortyx, вероятнее всего, представляет наиболее примитивных Phasianidae – до разделения на подсемейства Rollulinae и Phasianinae. Обсуждается эволюционная история рода и семейства Phasianidae в целом.
Ископаемый род Palaeortyx включает несколько видов мелких (с современного перепела или меньше) и среднего размера (крупнее серой куропатки) курообразных птиц, известных из нижнего олигоцена – верхнего миоцена Европы, Северной Азии и Африки (Mourer-Chauviré, 1992; Mlíkovský, 2002; Göhlich, Mourer-Chauviré, 2005; Göhlich, Pavia, 2008; Mayr, 2009). Наряду с родом Schaubortyx, Palaeortyx традиционно считается древнейшим несомненным представителем современного семейства Phasianidae sensu lato (Mourer-Chauviré, 1992; Mayr, 2009, 2017). Богатое видовое разнообразие Palaeortyx в олигоцене-миоцене (для этого временного интервала предполагается радиация многих современных линий Phasianidae: Stein et al., 2015) делают представителей Palaeortyx хорошими кандидатами на роль предков или ближайших родственников всех или отдельных эволюционных линий современных фазановых (см. Потапов, 1999). В то же время устоявшихся представлений о родственных связях и систематическом положении Palaeortyx нет.
Род был первоначально (Milne-Edwards, 1869) отнесен к американским (но см. ниже) зубчатоклювым куропаткам (Odontophorinae) – в первую очередь, из-за наличия глубокой дорсальной fossa pneumotricipitalis плечевой кости (см. ниже). Систематическое положение зубчатоклювых куропаток среди Galliformes долгое время оставалось крайне дискуссионным (см. обзор в: Cox et al., 2007); чаще всего их считали подсемейством в составе семейства Phasianidae s.l. (которое также включало цесарок, Numidinae), но иногда придавали ранг отдельного семейства. В настоящее время их выделяют в отдельное семейство Odontophoridae, поскольку современные молекулярные исследования (Cox et al., 2007; Hosner et al., 2015; Stein et al., 2015; Wang et al., 2017) помещают зубчатоклювых куропаток на филогенетическое древо курообразных (рис. 1) в качестве сестринской группы к Phasianidae s.s. При этом примечательно, что современные представления о филогенетическом положении Odontophoridae не были предположены ни одним из систематиков XIX–XX столетий (см. обзор в: Crowe, 1988; Sibley, Ahlquist, 1990).
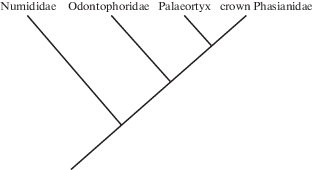
Позднее систематическое положение Palaeortyx было детально рассмотрено П. Бальманом (Ballmann, 1969). Он посчитал отнесение Palaeortyx к Odontophorinae ошибочным, сославшись на то, что глубокая дорсальная ямка плечевой кости независимо развилась у Odontophoridae и некоторых Phasianinae (она отсутствует у двух родов Odontophoridae, считавшихся примитивными, но присутствует у Ammoperdix и Coturnix из Phasianidae; см. Holman, 1961; Mayr et al., 2006). Таким образом, Бальман не посчитал наличие ямки существенным признаком для установления родства этих птиц, при этом отметил сходство между Palaeortyx и современными лесными куропатками Arborophila в строении тарсометатарсуса. Родственные отношения фазановых в то время были очень плохо понятны, и близкими родственниками Arborophila представлялись несколько родов, среди которых Ptilopachus (ныне относят к Odontophoridae). Бальман отметил, что ни один из этих родов не имеет настолько специализированной плечевой кости, как у Palaeortyx, ввиду чего он посчитал, что у этого ископаемого рода нет близких родственников в современной фауне. Тем не менее, Palaeortyx был использован для калибрации филогении Galliformes как представитель кроновых Odontophoridae (van Tuinen, Dyke, 2004).
Со времен публикации Бальмана долгое время не было попыток предпринять ревизию систематического положения рода. Только Г. Майр с соавт. (Mayr et al., 2006) предположил, что Palaeortyx, по-видимому, является базальной формой по отношению к современным фазановым, от которых он отличается плезиоморфными пропорциями конечностей (плечевая кость по длине примерно равна бедренной, как у палеогеновых курообразных). Он также отметил, что глубокая дорсальная ямка плечевой кости имеется у некоторых примитивных курообразных (см. Mourer-Chauviré, 1992) и поэтому является плезиоморфией Galliformes.
В последнее время филогения фазановых была существенно пересмотрена на основании молекулярных данных. Недавно было обнаружено, что два вида африканских курообразных, традиционно помещаемых в фазановых, в действительности представляют зубчатоклювых куропаток (Cohen et al., 2012). Эта неожиданная находка расширила ареал семейства Odontophoridae, которое до этого было представлено только американскими формами. Расхождение Odontophoridae и Phasianidae датируется концом эоцена – олигоценом (Hosner et al., 2015; Stein et al., 2015), при этом предполагается, что африканские зубчатоклювые куропатки Ptilopachus представляют собой результат олигоценовой дисперсии Odontophoridae через Берингию (Hosner et al., 2015). Современный разорванный ареал зубчатоклювых куропаток, действительно, позволяет ожидать их ископаемых находок в олигоцене или неогене Евразии и ставит вопрос переоценки систематического положения некоторых неогеновых родов фазановых, таких как Palaeortyx. Попытка подобной переоценки предпринята в настоящей работе.
Предпринятые попытки реконструировать филогению фазановых кладистическими методами (напр., Dyke et al., 2003) на основе морфологических признаков пока нельзя назвать удачными (см. Зеленков, 2009). Высокий уровень гомопластичного сходства между отдельными родами Phasianidae (при общем морфологическом однообразии группы) оставляют фактически единственный подход – анализировать морфологические признаки, опираясь на хорошо поддерживаемые молекулярные гипотезы о родстве. Ранее подобный подход уже был предпринят нами для коракоида фазановых (Zelenkov, Panteleyev, 2015).
МАТЕРИАЛ И МЕТОДЫ
В настоящей работе изучены материалы, относимые к Palaeortyx и другим ископаемым фазановым, из ряда олигоцен-миоценовых местонахождений Западной и Центральной Европы, хранящиеся в коллекциях Национального музея естественной истории в Париже, Лионского университета, Университета Монпелье (Франция), музеев естественной истории в Штутгарте и Берлине, Палеонтологической коллекции Мюнхена (далее BSP; Германия), а также Музея естественной истории Вены (Австрия). Важнейшие материалы, представленные многими сотнями костных остатков, происходят из серии местонахождений Керси (палеоген) и Сан-Жеран-ле-Пюи (нижний миоцен) во Франции и местонахождений Винтерсхоф-Вест (нижний миоцен) в Германии. Сохранность единственного полного скелета Palaeortyx из олигоцена Германии не позволяет проводить детальное остеологическое сравнение (Mayr et al., 2006), поэтому были использованы хорошо сохранившиеся разрозненные материалы. Для сравнения использовались сравнительно-остеологические коллекции современных птиц Палеонтологического ин-та им. А.А. Борисяка РАН (далее ПИН; Москва), Национального музея естественной истории в Вашингтоне (США), Сенкенбергского исследовательского ин-та (Франкфурт-на-Майне), Музея естественной истории в Штутгарте, а также Музея естественной истории в Барселоне (Испания).
АНАЛИЗ ОСТЕОЛОГИИ
Ниже приведен обзор наиболее характерных остеологических признаков представителей рода Palaeortyx по отдельным элементам скелета и их сравнение с состоянием у основных представителей отряда Galliformes.
Коракоид. Традиционно коракоид считался довольно однообразно устроенным у курообразных птиц (Holman, 1964) и по этой причине почти никогда не учитывался в диагнозах ископаемых таксонов (за исключением нескольких ископаемых семейств; см. Mourer-Chauviré, 1992). Яркая отличительная черта примитивных курообразных – наличие выемчатой cotyla scapularis; это несомненно плезиоморфный признак, сближающий Galliformes с их сестринской группой Anseriformes. У всех современных семейств Galliformes cotyla scapularis плоский, что также справедливо и для Palaeortyx и указывает на их эволюционную продвинутость. Плоский cotyla также позволяет отличать Palaeortyx от ископаемого семейства Paraortygidae, которые схожи с современными Phasianidae и могут представлять их предков.
Palaeortyx отличается от современных Numididae морфологией processus procoracoideus. У Numida и Acryllinum этот отросток имеет очень широкое каудальное основание с выпуклой кромкой. Дорсальная поверхность отростка у всех Numididae формирует плоскую поверхность, расположенную каудомедиальнее cotyla scapularis. У Palaeortyx дорсальная поверхность скошена и вышеупомянутой площадки нет, как у Odontophoridae и Phasianidae.
Н.В. Зеленков и А.В. Пантелеев (Zelenkov, Panteleyev, 2015) показали, что строение коракоида довольно вариабельно среди фазановых, при этом оказывается возможным сопоставить эволюцию этого элемента с молекулярной филогенией семейства. Наиболее примитивные из современных фазановых Rollulinae характеризуются довольно однотипным строением коракоида, в котором стержень кости начинает плавно, но равномерно отклоняться медиально (хорошо видно с дорсальной стороны), начиная примерно с уровня cotyla scapularis (рис. 2, г). Такую же морфологию имеет древнейший ископаемый представитель подсемейства из позднего миоцена Венгрии (Zelenkov, 2017). При этом impressio bicipitalis у Rollulinae совсем или практически не выступает медиально относительно медиальной кромки стержня (рис. 2, г). Palaeortyx отличается от Rollulinae более прямым стержнем, очень слабо отклоняющимся медиально в своей краниальной части, при этом impressio bicipitalis всегда заметно выступает медиально относительно медиальной кромки стержня. При виде с вентральной стороны коракоид Palaeortyx не демонстрирует схожести с какими-то из современных Phasianidae. В то же время, общие контуры кости у крупных видов Palaeortyx схожи с таковыми Rollulinae при виде с дорсальной стороны.
Рис. 2.
Коракоиды Palaeortyx и некоторых современных Phasianidae с вентральной стороны: a – Callipepla gambelii (Odontophoridae), экз. остеологической коллекции ПИН РАН 48-7-1, современный; б – Ptilopachus petrosus (Odontophoridae), экз. остеологической коллекции ПИН РАН 49-100-1, современный; в – Palaeortyx prisca, экз. BSP, № 18103; местонахождение Винтерсхоф-Вест, Бавария; нижний миоцен; г – Rollulus rouloul (Phasianidae; Rollulinae), экз. остеологической коллекции ПИН РАН 49-99-2, современный; д – Pternistis erckelii, экз. остеологической коллекции ПИН РАН 49-26-2, современный; е – Coturnix japonica, экз. остеологической коллекции ПИН РАН 49-67-1, современный. Обозначения: ib – impressio bicipitalis; pa – processus acrocoracoideus. Длина масштабной линейки 1 см.
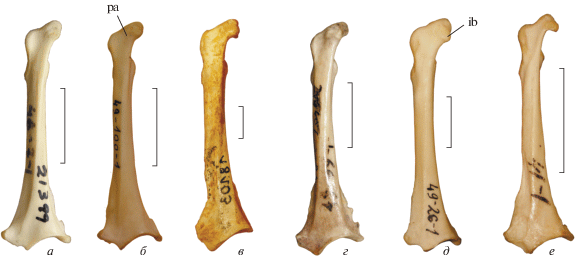
У Odontophoridae (включая Ptilopachus) коракоид также не в полной мере схож с таковым Palaeortyx (рис. 2, a, б). Processus acrocoracoideus также обычно отклоняется медиально, хотя при этом стержень может быть относительно ровным. Таким образом, по морфологии коракоида не удается установить родство Palaeortyx, который может оказаться или базальными Phasianidae, или примитивным Odontophoridae.
Глубокая дорсальная fossa pneumotricipitalis (fossa tricipitalis sensu Ballmann, 1969) плечевой кости. Присутствие этой ямки, расположенной дорсальнее crus dorsale fossa, характерно для очень небольшого числа птиц (она есть только у некоторых довольно продвинутых групп среди Charadriiformes и Passeriformes). Она отсутствует у стволовых (Presbyornithidae) и современных представителей Anseriformes – сестринского отряда для Galliformes, но при этом хорошо развита уже у древнейших палеогеновых курообразных из семейств Gallinuloididae и Paraortygidae, в скелете которых еще отмечается сходство с гусеобразными (Mourer-Chauviré, 1992; Mayr, 2000, 2006). Это позволяет считать наличие глубокой дорсальной ямки в проксимальной части плечевой кости апоморфией отряда Galliformes. В то же время, эта ямка отсутствует или очень плохо выражена у некоторых примитивных современных семейств (Cracidae, Megapodiidae, Numididae) и ископаемых Quercymegapodiidae. Все эти семейства считаются более продвинутыми по сравнению с Gallinuloididae и Paraortygidae (см. Mourer-Chauviré, 1992; Mayr, 2009), поэтому исчезновение этой ямки у них может быть вторичным. В пользу вторичности исчезновения fossa tricipitalis у Quercymegapodiidae, Cracidae, Megapodiidae и Numididae говорит тот факт, что поверхность, занятая этой ямкой у более примитивных форм, у данных семейств плоская или даже слегка вогнутая. У птиц, вовсе не имеющих данной ямки, эта поверхность выпуклая, а зачастую в этой позиции даже имеется умеренно или хорошо выраженный каудальный гребень стержня.
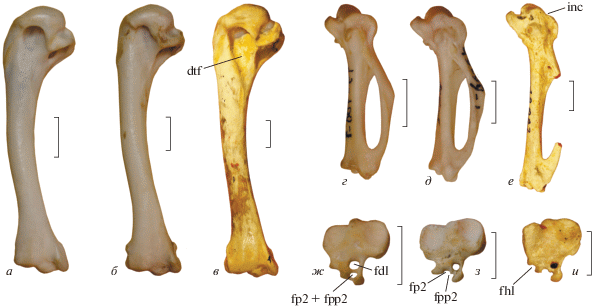
Нельзя установить точно, произошло ли исчезновение ямки единовременно в эволюции курообразных, или же независимо в четырех указанных выше семействах – оба варианта представляются равновероятными. Однако эта ямка вновь “появляется” (и при этом очень хорошо развита) у более продвинутого семейства Odontophoridae, при этом она отсутствует даже у некоторых представителей и этой группы (роды Dendrortyx и Odontophorus; Holman, 1964). Бальман (Ballmann, 1969) считал эти роды примитивными, и на этом основании посчитал этот признак (наличие глубокой дорсальной ямки) возникшим вторично в эволюции Odontophoridae. В то же время, современные кладистические исследования не подтверждают примитивность родов Dendrortyx и Odontophorus (Hosner et al., 2015; Stein et al., 2015). Кроме того, у недавно “открытых” современных африканских Odontophoridae (род Ptilophachus; Cohen et al., 2012) также имеется очень хорошо выраженная глубокая fossa tricipitalis (наши данные, рис. 3). Ptilophachus представляется сестринским родом по отношению к остальным Odontophoridae (Hosner et al., 2015); таким образом, наличие глубокой хорошо выраженной ямки можно считать исходным состоянием для Odontophoridae. Но для того, чтобы ответить на вопрос, является ли этот признак синапоморфией Odontophoridae, или же он представляет собой плезиоморфное состояние, унаследованное от общего предка Odontophoridae + Phasianidae, нужно рассмотреть строение плечевой кости у Phasianidae, которые представляют сестринскую группу для Odontophoridae (Wang et al., 2013).
Рис. 3.
Избранные кости Palaeortyx и некоторых современных Phasianidae: а, г, ж – Ptilopachus petrosus (Odontophoridae), экз. остеологической коллекции ПИН РАН 49-100-1, современный; б, д, з – Rollulus rouloul (Phasianidae; Rollulinae), экз. остеологической коллекции ПИН РАН 49-99-2, современный; в, е, и – Palaeortyx prisca, экз. BSP, № 18046; местонахождение Винтерсхоф-Вест, Бавария; нижний миоцен. a–в – плечевые кости с каудальной стороны; г–е – карпометакарпусы с вентральной стороны; ж–и – тарзометатарзусы с проксимальной стороны (показан гипотарзус). Обозначения: dtf – fossa pneumotricipitalis dorsalis; inc – вырезка в вентральном крае trochlea carpalis; fdl – канал m. flexor digitorum longus; fhl – борозда m. flexor hallucis longus; fp2 – борозда (канал) m. flexor perforans digiti II; fpp2 – борозда (канал) flexor perforatus et perforans digiti II. Длина масштабной линейки 0.5 см.
Самые примитивные из современных Phasianidae – представители подсемейства Rollulinae, объединяющего мелких лесных фазановых, встречающихся в тропиках Старого Света. Для Rollulinae характерна хорошо выраженная дорсальная fossa tricipitalis, заходящая под caput humeri – однако при этом она остается довольно мелкой (ее глубина, измеренная на уровне каудальной границы caput humeri, как минимум в три раза уступает ширине ямки на этом же уровне). Среди фазановых из второго подсемейства, Phasianinae, мелкая и умеренно выраженная ямка встречается у представителей разных линий, в частности, у примитивных (Wang et al., 2013) родов Lophophorus, Ithaginus и Polyplectron. Довольно сильно эта ямка развита у Pucrasia, где она, правда, не заходит под caput humeri. Широкое распространение умеренно выраженной fossa tricipitalis у разных фазановых говорит о том, что исходное состояние для Phasianinae могло быть сравнимо с таковым у Rollulinae (хорошо выраженная, но неглубокая ямка). При этом состояние у современных Phasianinae представляется более продвинутым, поскольку последовательные сестринские группы Odontophoridae и Rollulinae имеют более глубокую fossa tricipitalis. Немаловажно заметить, что предок современных Phasianidae мог все же иметь глубокую ямку, как у сестринских Odontophoridae.
Глубокая fossa tricipitalis среди Phasianinae имеется только у трибы Coturnicini, при этом у Tetraogallus и Pternistis, считающихся наиболее примитивными родами в этой группе (Wang et al., 2013), эта ямка еще не заходит под caput humeri; у более продвинутого Alectoris caput humeri уже нависает над ямкой (при этом глубина этой ямки заметно варьирует у представителей рода). У Ammoperdix и Coturnix (sensu lato) fossa tricipitalis особенно глубока (несомненно вторично) и сравнима с таковой у примитивных Galliformes и современных Odontophoridae.
Таким образом, само по себе наличие или отсутствие глубокой fossa tricipitalis не является достаточным критерием для отнесения той или иной ископаемой формы к определенному семейству курообразных, однако у фазановых эта ямка возникает только у одной группы в трибе Coturnicini, которая легко идентифицируется и по другим элементам скелета (в частности, по коракоиду; Zelenkov, Panteleyev, 2015). Поэтому, судя только по наличию глубокой fossa tricipitalis, Palaeortyx может оказаться как представителем Odontophoridae, так и представителем стволовых Phasianidae, а также членом неназванной клады (внутри трибы Coturnicini), включающей современных Alectoris, Ammoperdix и Coturnix sensu lato (что совершенно не поддерживается другими признаками).
Отсутствие пневматизации fossa pneumotricipitalis плечевой кости. Диагностичной особенностью рода Palaeortyx является отсутствие пневматизации fossa pneumotricipitalis (Göhlich, Mourer-Chauviré, 2005). Этот признак однозначно указывает на примитивность рода, поскольку у современных Phasianidae и Odontophoridae эта фосса всегда пневматизована. Несколько хуже пневматизация выражена у Cracidae и Numididae, но полностью отсутствует, как и у Palaeortyx, у примитивного ископаемого семейства Paraortygidae (Mourer-Chauviré, 1992). Таким образом, по данному признаку Palaeortyx выглядит даже более примитивным, чем Numididae и сближается с ископаемыми Paraortygidae.
Карпометакарпус. Современные фазановые и родственные им курообразные имеют в общем довольно однообразно устроенный карпометакарпус. Характерной особенностью продвинутых Galliformes (Odontophoridae + Phasianidae) является наличие хорошо развитого processus intermetacarpalis, который имеется и у Palaeortyx. У Numididae также вентральная часть trochlea carpalis не разделена на два полублока вырезкой, которая обычно хорошо развита у Phasianidae и имеется у Palaeortyx. У Odontophoridae эта вырезка развита слабее, чем у Phasianidae и Palaeortyx, и каудальная часть trochlea carpalis у них не сильно сдвинута дистально (рис. 3). По этим признакам Palaeortyx более продвинут, чем Numididae и Odontophoridae.
Современные представители Odontophoridae + + Phasianidae различаются, в основном, степенью изогнутости metacarpalia minor, а также тем, насколько она выступает дистально относительно metacarpalia major. Среди опубликованных или изученных нами материалов по Palaeortyx не оказалось ни одного с полностью сохранившейся metacarpalia minor, однако о степени ее изогнутости у Palaeortyx можно судить по очертаниям ее кудального края в области дистального симфиза. По морфологии этой области Palaeortyx схож как с Odontophoridae, так и с некоторыми фазановыми, например, Rollulinae. В целом, для Odontophoridae и Rollulinae характерна более выгнутая каудально metacarpalia minor, однако сходное состояние (по-видимому, плезиоморфное) иногда встречается и у других фазановых, например, у Polyplectron или Pternistis.
Бедренная кость у Palaeortyx довольно выпрямленная, особенно дорсовентрально, что хорошо заметно при виде с латеральной стороны. Кривизна бедренной кости варьирует у современных Phasianidae, но у Phasianinae стержень бедренной кости обычно заметно изогнут каудально в своей дистальной трети. Прямая бедренная кость, как у Palaeortyx, характеризует современных Rollulinae и Numididae. У Odontophoridae, включая Ptilophachus, она более изогнута. Таким образом, по этому признаку Palaeortyx может оказаться Rollulinae, стволовым Phasianinae или стволовым Phasianidae.
Тарзометатарзус современных Phasianidae характеризуется наличием хорошо развитого медиального плантарного гребня (crita plantaris medialis), который в некоторых случаях может нести шпору и обычно тянется в проксимальном направлении, соединяясь с медиальным гребнем гипотарзуса. Медиальный плантарный гребень отсутствует у Odontophoridae и Numididae, но есть у примитивных фазановых (Rollulinae) и у большинства Phasianinae, поэтому может считаться синапоморфией Phasianidae. Опять таки, исчезновение этого гребня среди Phasianidae характерно для Ammoperdix, Coturnix (sensu lato) и Tetraogallus (то есть у Coturnicini), а также родственных им Gallus и Bambusicola. У Alectoris присутствие этого гребня варьирует. Несомненное отсутствие этого гребня у Palaeortyx (см. Ballmann, 1969) указывает на возможность отнесения этого рода либо к Coturnicini, либо к Odontophoridae. Это также может рассматриваться как примитивное состояние для Galliformes, поскольку гребень практически не выражен у Numididae и отсутствует у более примитивных семейств (современные Megapodiidae и Cracidae, ископаемые Quercymegapodiidae, Paraortygidae). И хотя строение проксимального эпифиза тарзометатарзуса позволяет различать различные роды фазановых (см. также Ballmann, 1969), практически не удается выявить признаки, которые бы однозначно отличали Odontophoridae от Coturnicini. Возможно, диагностичным можно считать развитие у Palaeortyx глубоких fossa parahypotarsales (Göhlich, Mourer-Chauviré, 2005). Эти отпечатки сильно выражены у исследованных Odontophoridae и, в частности, у Ptilopachus. Если медиальная ямка бывает хорошо развита у фазановых (например, у Rollulus, Galloperdix), то латеральная почти всегда мелкая или совсем не выражена, в то время как у Odontophoridae она обычно лучше развита. Однако хорошо развитые отпечатки имеются и у примитивных фазановых Rollulinae, а также у Numididae, Cracidae и Megapodiidae. Таким образом, обе диагностичные черты Palaeortyx, отсутствие сильно выраженного медиального плантарного гребня и наличие хорошо развитых fossa parahypotarsales, могут рассматриваться как особенности продвинутых курообразных в целом (Numididae + Odontophoridae + Phasianidae).
Строение hypotarsus зачастую очень диагностично у птиц (см. Mayr, 2016). В основном, именно по строению гипотарзуса Бальман (Ballmann, 1969) сближал Palaeortyx с Arborophila. Odontophoridae имеют особенное строение гипотарзуса: у них он высок дорсоплантарно, сухожилия fp2 и fpp2 проходят обычно в замкнутом канале (см. Mayr, 2016), который при этом располагается плантарнее канала fdl (рис. 3). Высокий гипотарзус с замкнутым каналом fp2 и fpp2, как у Odontophoridae – несомненно продвинутое состояние среди курообразных, поскольку у всех примитивных семейств (в том числе, у Palaeortyx) он низкий, а вместо канала fp2 и fpp2 присутствуют одна или две незамкнутые борозды (рис. 3). Продвинутое состояние гипотарзуса, характеризующее Odontophoridae, возникает и у некоторых фазановых (напр., у Bambusicola и Francolinus), однако нет сомнений, что примитивное состояние для Phasianidae было иное – низкий гипотарзус с двумя более или менее разделенными бороздами fp2 и fpp2 (без формирования закнутого канала), при этом борозда fp2 смещена в большей или меньшей степени дорсальнее (располагается глубже) относительно борозды fpp2, и обычно несколько погружается в тело гипотарзуса, а в некоторых случаях (Mayr, 2016) даже может замыкаться. Такое строение гипотарзуса характерно для Numididae и наиболее примитивных фазановых Rollulinae, и именно таким образом устроен гипотарзус у Palaeortyx (по этой структуре ископаемый род, действительно, ближе всего Rollulinae). Относительное положение борозд fp2 и fpp2 сохраняется у многих фазановых (например, Crossoptilon, Syrmaticus, Tetraogallus) при изменении пропорций тела гипотарзуса. В частности, у представителей трибы Coturnicini (за исключением Tetraogallus) гипотарзус становится дорсоплантарно выше (как у Odontophoridae), а борозды fp2 и fpp2 сливаются в одну, расположенную плантарнее канала fdl.
Еще одна диагностическая черта строения гипотарзуса – положение борозды fhl. Майр (Mayr, 2016) отмечает, что у курообразных она ориентирована латерально, за исключением некоторых таксонов (напр., Alectoris), у которых она ориентирована плантарно. Исходным для Galliformes состоянием, по-видимому, следует считать косую (плантолатеральную) ориентацию этой борозды, как у Megapodiidae, Cracidae и Numididae. У некоторых Phasianidae (Pavo, Tetraogallus) эта борозда также ориентирована больше плантарно, чем латерально. У исследованных нами экземпляров Alectoris она ориентирована или латерально, или плантолатерально. Однако у большинства Phasianidae (в том числе у примитивных Rollulinae) и Odontophoridae она ориентирована строго латерально и обычно огорожена с плантарной стороны хорошо развитым гребнем. У Palaeortyx эта борозда примитивным образом ориентирована плантолатерально, а гребень, обрамляющий ее с плантарной стороны, выражен слабо.
Таким образом, строение гипотарзуса не позволяет отнести Palaeortyx ни к семейству Odontophoridae, ни к продвинутым Phasiandae из трибы Coturnicidae, с которыми этот род сближается по строению плечевой кости. Характерно сходство с Rollulinae в строении гипотарзуса, однако оно тоже не полное, и ориентация борозды fhl указывает на примитивность по отношению к современным Rollulinae или фазановым в целом. Однако тот факт, что примитивное устройство гипотарзуса наблюдается также у Numididae, может указывать на базальное положение Palaeortyx по отношению к кладе Phasianidae + Odontophoridae.
Пропорции конечностей. Ранее отмечалось (Mayr et al., 2006), что для Palaeortyx характерны особые пропорции конечностей: длина бедренной кости примерно равна длине плечевой, в то время как у подавляющего большинства Phasianidae и Odontophoridae бедренная кость заметно (иногда существенно) длиннее плечевой. Сходные с Palaeortyx пропорции все же встречаются у Tetraoninae, а также у отдельных Odontophoridae (Cyrtonyx) и Phasianinae (Ammoperdix). Примечательно, что у Ptilopachus бедренная кость длинная, так же как и у Coturnix (видимо, этот эффект обусловлен вторичным удлинением плечевой кости у этого рода) и примитивных Rollulinae. В то же время, бедренная кость короткая у примитивных курообразных, а также у современных Numididae (Mayr et al., 2006), поэтому можно предположить, что у Palaeortyx сохранилось примитивное состояние. Это не исключает трактовки рода как стволового представителя Odontophoridae, Phasianidae или даже до расхождения этих семейств.
ОБСУЖДЕНИЕ
Систематическое положение Palaeortyx. Приведенный выше анализ морфологических признаков подтверждает отнесение Palaeortyx к надсемейству Phasianoidea (ранее в ранге семейства Phasianidae; Ballmann, 1969; Mourer-Chauviré, 1992; Mayr et al., 2006). При этом ряд признаков Palaeortyx соответствует эволюционному уровню Numididae или даже ископаемых стволовых курообразных Paraortygidae, то есть указывает на примитивность рода по отношению к Phasianidae. Однако Palaeortyx все же эволюционно более продвинут, чем Numididae и Paraortygidae, что следует из строения карпометакарпуса (наличие processus intermetacarpalis и строение trochlea carpalis), а также коракоида – быстро эволюционирующего у курообразных элемента скелета. Сходство Palaeortyx с Odontophoridae и, в частности, с базальным родом Plitopachus, незначительно (за исключением дорсальной fossa tricipitalis плечевой кости). Расхождение Ptilophachus и Odontophoridae Нового Света датируется, по молекулярным данным, поздним эоценом-олигоценом (Hosner et al., 2015), что соответствует нахождению каких-то мелких курообразных (возможно Odontophoridae; но см. Mayr, 2009) в палеонтологической летописи Северной Америки уже в раннем олигоцене (Olson, 1985). Тот факт, что Ptilophachus схож с современными новосветскими Odontophoridae по основным обсуждаемым в этой работе признакам, говорит о том, что морфологический тип семейства Odontophoridae (включающий продвинутое строение гипотарзуса), по-видимому, уже сложился к олигоцену. С учетом этого, слабое сходство олигоцен-миоценовых Palaeortyx с Odontophoridae указывает на то, что Palaeortyx – все же более продвинутая форма, чем Odontophoridae, и определенное наблюдаемое сходство с этим семейством по некоторым признакам (например, глубокая дорсальная ямка плечевой кости), скорее всего, плезиоморфно. Palaeortyx выглядит более продвинутым по сравнению с Odontophoridae по строению trochlea carpalis карпометакарпуса.
Наибольшее сходство Palaeortyx проявляет с примитивными современными фазановыми Rollulinae, однако при этом, несомненно, выглядит заметно более примитивным, чем современные Rollulinae. У фазановых очень диагностично строение коракоида, который у Palaeortyx отличается от такового Rollulinae. Положение канала fdl в гипотарсусе Palaeortyx также указывает на примитивность рода по отношению к современным Rollulinae. Примитивна и очень глубокая дорсальная ямка плечевой кости, а также отсутствие пневматизации вентральной ямки. Эти примитивные черты, при отсутствии каких-либо продвинутых признаков Rollulinae, указывают на базальное положение Palaeortyx по отношению ко всем современным Phasianidae. Таким образом, наиболее вероятно, что Palaeortyx представляет стволовых фазановых, то есть, еще до разделения на Rollulinae и Phasianinae (рис. 1).
Эволюционная история Palaeortyx. К концу олигоцена Palaeortyx уже представлен в Западной Европе несколькими видами (Mourer-Chauviré, 1992; Mayr, 2009). В раннем миоцене Palaeortyx имели наибольший ареал и найдены, помимо Европы, в Намибии (Mourer-Chauviré, 2008), а также в Прибайкалье (Тесаков и др., 2015). Их широкое распространение может быть связано с миоценовым климатическим оптимумом на рубеже раннего и среднего миоцена (Zachos et al., 2001), однако уже в конце среднего миоцена (около 15 млн. л. н.) начинаются заметные колебания климата, после которых наступает эпоха похолодания, сопровождающаяся возникновением аридного пояса в Центральной Азии (Flower, Kennett, 1994). В этой связи следует отметить, что в конце среднего миоцена в Монголии Palaeortyx нет, но присутствует другой род Tologuica (Зеленков, Курочкин, 2009). Tologuica похожа на Palaeortyx строением плечевой кости, медленно эволюционирующей у фазановых, но зато надежно отличается, в частности, морфологией коракоида, который, как отмечалось выше, довольно вариабелен у фазановых и может служить маркером эволюционных изменений в этой группе птиц. Примечательно, что по строению коракоида Tologuica схожа с неназванными фазановыми из среднего миоцена Намибии (наши данные) и, таким образом, может представлять собой африканских мигрантов-обитателей аридных ландшафтов, вместе с другими группами птиц впервые появившихся в Азии именно в это время.
В позднем миоцене Palaeortyx практически не известен в палеонтологической летописи и, по-видимому, мог сохраниться только в отдельных рефугиумах, таких как острова на месте современного Апеннинского полуострова (Ballmann, 1976; Göhlich, Pavia, 2008). Позднемиоценовые находки Palaeortyx в местонахождениях Венгрии не подтвердились (Зеленков, 2016), при этом важно, что Palaeortyx нет уже в местонахождении Рудобаня, датируемом началом позднего миоцена (Zelenkov, 2017). Вопрос родственных связей Palaeortyx с другими родами Phasianidae остается открытым. Предполагается, что этот ископаемый род может быть предком современных Francolinus s.l. (Потапов, 1999), однако в таком случае, учитывая морфологическую примитивность Palaeortyx, следовало бы ожидать, что Francolinus представляют собой наиболее примитивных из современных Phasianinae, что не подтверждается молекулярной филогенетикой (Kimball, Braun, 2014; Stein et al., 2015; Hosner et al., 2017; Cai et al., 2018).
Происхождение и ранняя эволюция Phasianidae. Новейшие молекулярные датировки дивергенции Phasianidae относительно сестринских Odontophoridae довольно сильно различаются: от середины эоцена (Hosner et al., 2017; Cai et al., 2018) до середины олигоцена (Prum et al., 2015). Присутствие стволовых фазановых Palaeortyx в палеонтологической летописи уже в раннем олигоцене не исключает возможности того, что дивергенция семейства и правда имела место в эоцене. Майр (Mayr, 2009, 2017) предположил, что Palaeortyx могут быть потомками примитивных Phasianidae – мигрантов из Азии, проникших в Европу при закрытии Тургайского пролива. Для подтверждения этой гипотезы данных пока очень мало – Telecrex grangeri, похожий на современных Numididae и отнесенный к этой группе Олсоном (Olson, 1974), может оказаться представителем одного из стволовых семейств курообразных (Mourer-Chauviré, 1992; Mayr, 2009). Раннеолигоценовые таксоны Palaeorallus aliensis и Gobihierax edax из Центральной Монголии, недавно отнесенные к Galliformes, известны по очень фрагментарным и слабо диагностичным остаткам и также пока не определены до уровня семейства (Зеленков, Курочкин, 2015). Раннеэоценовые курообразные Монголии, пока остающиеся не изученными, по-видимому, относятся к стволовым семействам (наши данные).
Биогеографический анализ современного разнообразия фазановых показал, что семейство Phasianidae могло возникнуть в Африке; там же должны были иметь место и базальные этапы дивергенции внутри семейства (Cai et al., 2018). Известные эоценовые курообразные Африки (роды Namaortyx, Chambiortyx и Scopelortyx) не могут быть отнесены к Phasianidae и, таким образом, не поддерживают данную гипотезу. Однако олигоценовые фазановые Африки остаются пока вовсе неизвестными. Нельзя исключить вероятность того, что ранние этапы эволюции Phasianidae, действительно, имели место в Африке в олигоценовую эпоху. Этот сценарий подразумевает миграцию Palaeortyx или их предков в Западную Европу именно из Африки.
По молекулярным данным, базальная дивергенция Phasianidae, то есть расхождение Rollulinae и Phasianinae, датируется поздним эоценом (Hosner et al., 2017), границей эоцена и олигоцена (Cai et al., 2018), или олигоценом (Prum et al., 2015). Олигоценовая датировка выглядит более правдоподобно, с учетом того, что из олигоценовых отложений пока не известны какие-либо несомненные представители обоих подсемейств: древнейший представитель Rollulinae описан из низов верхнего миоцена Венгрии (Zelenkov, 2017), древнейшими представителями Phasianinae могут оказаться представители трибы Tetraonini из раннего миоцена Северной Америки (Olson, 1985). В Евразии древнейшими несомненными Phasianinae, скорее всего, являются роды Palaeocryptonyx, Tologuica, Miogallus и Lophogallus, известные только начиная со среднего миоцена (см. Mlíkovský, 2002; Зеленков, Курочкин, 2009, 2010). В целом, следует отметить, что этап эволюции фазановых от примитивных Palaeortyx до древнейших представителей современных триб остается наименее изученным (по палеонтологическим данным).
Список литературы
Зеленков Н.В. Филогенетический анализ некоторых неогеновых родов фазановых (Aves: Phasianidae) // Палеонтол. журн. 2009. № 4. С. 77–82.
Зеленков Н.В. Ревизия неворобьиных птиц Полгарди (Венгрия, верхний миоцен). 2. Galliformes // Палеонтол. журн. 2016. № 6. С. 79–91.
Зеленков Н.В., Курочкин Е.Н. Неогеновые фазановые (Aves: Phasianidae) Центральной Азии. 1. Род Tologuica gen. nov. // Палеонтол. журн. 2009. № 2. С. 86–92.
Зеленков Н.В., Курочкин Е.Н. Неогеновые фазановые (Aves: Phasianidae) Центральной Азии. 3. Роды Lophogallus gen. nov. и Syrmaticus // Палеонтол. журн. 2010. № 3. С. 79–87.
Зеленков Н.В., Курочкин Е.Н. Класс Aves // Ископаемые рептилии и птицы. Часть 3 / Pед. Е.Н. Курочкин, А.В. Лопатин, Н.В. Зеленков. М.: ГЕОС, 2015. С. 86–290.
Потапов Р.Л. Происхождение и эволюция фауны курообразных птиц (Galliformes) Эфиопской области // Зоол. журн. 1999. Т. 78. С. 301–302.
Тесаков А.С., Сыромятникова Е.В., Данилов И.Г. и др. Успехи изучения миоценовых позвоночных местонахождения Тагай (остров Ольхон, озеро Байкал) // Палеонтология Центральной Азии и сопредельных регионов. Междунар. конф. к 45-лет. СРМПЭ. Тез. докл. М., ПИН РАН. С. 75–77.
Ballmann P. Les oiseaux de la Grive-Saint-Alban (Isere) // Geobios. 1969. V. 2. P. 157–204.
Ballmann P. Fossil birds from the Neogene of Gargano Peninsula (Italy), part two // Scripta Geol. 1976. V. 38. P. 1–59.
Cai T., Fjeldså J., Wu Y. et al. What makes the Sino-Himalayan mountains the major diversity hotspots for pheasants? // J. Biogeogr. 2018. V. 45. № 3. P. 640–651.
Cohen C., Wakeling J.L., Mandiwana-Neudani T.G. et al. Phylogenetic affinities of evolutionarily enigmatic African galliforms: the Stone Partridge Ptilopachus petrosus and Nahan’s Francolin Francolinus nahani, and support for their sister relationship with New World quails // Ibis. 2012. V. 154. № 4. P. 768–780.
Cox W.A., Kimball R.T., Braun E.L. Phylogenetic position of the new world quail (Odontophoridae): eight nuclear loci and three mitochondrial regions contradict morphology and the sibley-ahlquist tapestry // Auk. 2007. V. 124. № 1. P. 71–84.
Crowe T.M. Molecules vs morphology in phylogenetics: a non-controversy // Trans. R. Soc. South Africa. 1988. V. 46. № 4. P. 317–334.
Dyke G.J., Gulas B.E., Crowe T.M. Suprageneric relationships of galliform birds (Aves, Galliformes): a cladistic analysis of morphological characters // Zool. J. Linn. Soc. 2003. V. 137. P. 227–244.
Flower B.P., Kennett J.P. The middle Miocene climatic transition: East Antarctic ice sheet development, deep ocean circulation and global carbon cycling // Palaeogeogr., Palaeoclimatol., Palaeoecol. 1994. V. 108. P. 537–555.
Göhlich U.B., Mourer-Chauviré C. Revision of the phasianids (Aves: Galliformes) from the Lower Miocene of Saint-Gerand-Le-Puy (Allier, France) // Palaeontology. 2005. V. 48. № 6. P. 1331–1350.
Göhlich U.B., Pavia M. A new species of Palaeortyx (Aves: Galliformes: Phasianidae) from the Neogene of Gargano, Italy // Oryctos. 2008. V. 7. P. 95–108.
Holman J.A. Osteology of gallinaceous birds // Quart. J. Florida Acad. Sci. 1964. V. 27. № 3. P. 230–252.
Hosner P.A., Braun E.L., Kimball R.T. Land connectivity changes and global cooling shaped the colonization history and diversification of New World quail (Aves: Galliformes: Odontophoridae) // J. Biogeogr. 2015. V. 42. № 10. P. 1883–1895.
Hosner P.A., Tobias J.A., Braun E.L., Kimball R.T. How do seemingly non-vagile clades accomplish trans-marine dispersal? Trait and dispersal evolution in the landfowl (Aves: Galliformes) // Proc. Roy. Soc. B: Biol. Sci. 2017. V. 284: 20170210.
Kimball R.T., Braun E.L. Does more sequence data improve estimates of galliform phylogeny? Analyses of a rapid radiation using a complete data matrix // Peer J. 2014. V. 2: e361.
Mayr G. A new basal galliform bird from the middle Eocene of Messel (Hessen, Germany) // Senckenb. lethaea. 2000. V. 80. № 1. P. 45–57.
Mayr G. New specimens of the early Eocene stem group galliform Paraortygoides (Gallinuloididae), with comments on the evolution of a crop in the stem lineage of Galliformes // J. Ornithol. 2006. V. 147. № 1. P. 31–37.
Mayr G. Paleogene fossil birds. Berlin, Heidelberg: Springer-Verlag, 2009. 262 p.
Mayr G. Variations in the hypotarsus morphology of birds and their evolutionary significance // Acta Zool. 2016. V. 97. № 2. P. 196–210.
Mayr G. Avian evolution. The fossil record of birds and its paleobiological significance. Chichester, West Sussex: John Wiley & Sons, 2017. 293 p.
Mayr G., Poschman M., Wuttke M. A nearly complete skeleton of the fossil galliform bird Palaeortyx from the late Oligocene of Germany // Acta Ornithol. 2006. V. 41. № 2. P. 129–135.
Milne-Edwards A. Recherches anatomiques et paléontologiques pour servir à l’histoire des oiseaux fossiles de la France. P.: G. Masson, 1869.
Mourer-Chauviré C. The Galliformes (Aves) from the Phosphorites du Quercy (France): systematics and biostratigraphy // Natur. Hist. Mus. Los Angels Co., Sci. Ser. 1992. № 36. P. 67–95.
Mourer-Chauviré C. Birds (Aves) from the Early Miocene of the Northern Sperrgebiet, Namibia // Mem. Geol. Surv. Namibia. 2008. V. 20. P. 147–167.
Olson S.L. Telecrex restudied: a small Eocene guineafowl // Wilson Bull. 1974. V. 86. № 3. P. 246–250.
Olson S.L. The fossil record of birds // Avian Biology. Vol. 8 / Eds. Farner D.S., King J.R., Parkes K.C. N.Y.: Acad. Press, 1985. P. 79–238.
Prum R.O., Berv J.S., Dornburg A. et al. A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing // Nature. 2015. V. 526. № 7574. P. 569–573.
Sibley C.G., Ahlquist J.E. Phylogeny and classification of birds: a study in molecular evolution. New Heaven, L.: Yale Univ. Press, 1990. 976 p.
Stein R.W., Brown J.W., Mooers A.Ø. A molecular genetic time scale demonstrates Cretaceous origins and multiple diversification rate shifts within the order Galliformes (Aves) // Mol. Phyl. Evol. 2015. V. 92. P. 155–164.
van Tuinen M., Dyke G.J. Calibration of galliform molecular clocks using multiple fossils and genetic partitions // Mol. Phyl. Evol. 2004. V. 30. P. 74–86.
Wang N., Kimball R.T., Braun E.L. et al. Assessing phylogenetic relationships among Galliformes: A multigene phylogeny with expanded taxon sampling in Phasianidae // PLoS One. 2013. V. 8. № 5: e64312.
Wang N., Kimball R.T., Braun E.L. et al. Ancestral range reconstruction of Galliformes: the effects of topology and taxon sampling // J. Biogeogr. 2017. V. 44. № 1. P. 122–135.
Zachos J., Pagani M., Sloan L. et al. Trends, rhythms, and aberrations in global climate 65 Ma to present // Science. 2001. V. 292. P. 686–693.
Zelenkov N.V. The revised avian fauna of Rudabànya (Hungary, Late Miocene) // Paleontología y evolución de las Aves / Eds. Acosta Hospitaleche C., Agnolin F., Haidr N. et al. Buenos-Aires: Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, 2017. P. 253–266.
Zelenkov N.V., Panteleyev A.V. Three bird taxa (Aves: Anatidae, Phasianidae, Scolopacidae) from the Late Miocene of the Sea of Azov (Southwestern Russia) // Paläontol. Z. 2015. Bd 89. № 3. P. 515–527.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал