

Палеонтологический журнал, 2019, № 3, стр. 59-65
Орнаментация и микроструктура раковины Orbiculoidea magnifica Mergl (Brachiopoda, Lingulida) из нижнего девона Тимано-Печoрской впадины
Т. Н. Смирнова a, *, Ю. А. Гатовский a, **, Е. А. Жегалло b, ***, Г. Т. Ушатинская b, ****
a Московский государственный университет им. М.В. Ломоносова
Москва, Россия
b Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: smirnovatatiana76@bk.ru
** E-mail: ustas62bk.ru
*** E-mail: ezheg@paleo.ru
**** E-mail: gushat@paleo.ru
Поступила в редакцию 02.07.2018
После доработки 05.07.2018
Принята к публикации 05.07.2018
Аннотация
Впервые описаны хитиново-фосфатные брахиоподы из нижнедевонских отложений лохковского яруса Тимано-Печорской впадины. Вместе с брефической (личиночной) раковиной на нескольких экземплярах обнаружен протегулюм (эмбриональная раковина). Впервые описана микроструктура раковины раннедевонского представителя рода Orbiculoidea – O. cf. magnifica Mergl, 2001. Установлены отличия микроструктуры раковины на разных возрастных стадиях.
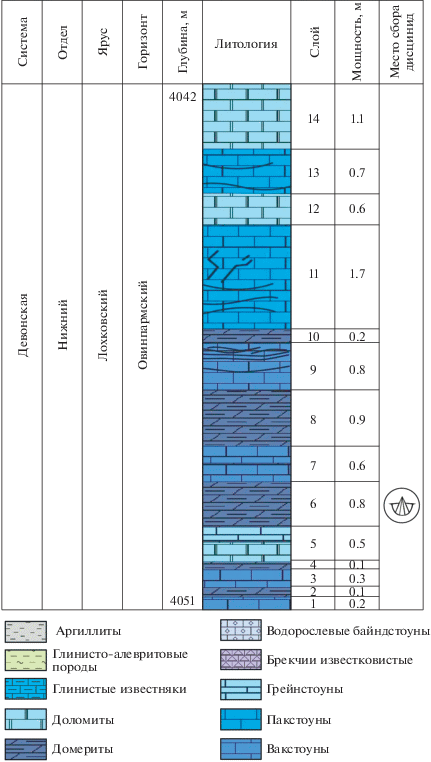
Брахиоподы найдены в отложениях лохковского яруса овинпармского горизонта северо-восточной части Тимано-Печорской впадины (северо-восточная часть Ненецкого автономного округа). Материал получен из керновых образцов скважины Варкнавтской, из доломитовых мергелей на глубине 4042–4051 м (рис. 1, 2). Скважина пробурена на территории Садаягинской ступени Хорейверской впадины.
Род Orbiculoidea d’Orbigny, 1847 известен с раннего ордовика. Девонские представители рода упоминаются в ряде публикаций. У нескольких видов этого рода было обнаружено наличие протегулюма и брефической раковины (Freeman, Landelius, 1999). О. Вильямс и М. Кусак (Williams, Cusack, 1997) при изучении микроструктуры раковин позднепалеозойских дисцинацей установили, что большая часть коллекции состоит из перекристаллизованных экземпляров, хорошая сохранность раковины имеется только у Orbiculoidea nitida (Phillips, 1836) из нижнего карбона Англии и Шотландии. Детали микроструктуры были подробно описаны этими авторами, проведено сравнение с известными палеозойскими и современными дисцинидами. Три вида Orbiculoidea описаны М. Мергл (Mergl, 2001) из девонских отложений Богемии (Чешская Республика). Мергл дает подробное описание наружного строения O. bohemica (Barrande, 1879) из нижнего девона, пражского яруса и O. tarda (Barrande, 1879) из среднего девона, нижней части живетского яруса. В результате изучения коллекции И. Барранда, хранящейся в Национальном музее в Праге, Мергл была проведена ревизия систематического состава силурийских и девонских видов рода Orbiculoidea. Подробные сведения относительно орнаментации и микроструктуры раковины раннедевонского вида O. cf. magnifica Mergl, 2001 из Тимано-Печорской впадины приводятся в настоящей статье.
В коллекции Ю.А. Гатовского, описанной в настоящей статье, имеются раковины хитиново-фосфатных брахиопод, представленные 7 створками, из них 3 створки спинные и 4 створки брюшные. Спинные створки имеют правильные округлые очертания, макушка смещена по направлению к заднему краю на треть длины створки (рис. 3, а). В месте протегулюма имеется округлый участок диаметром около 90 мкм. Он хорошо выделяется в рельефе макушечной части, хотя четкой границы с брефической раковиной не наблюдается (рис. 3, б). Брефическая раковина округлой формы, ее диаметр около 200 мкм (рис. 3, в). Первичный слой сохранился удовлетворительно на большей части раковины. В месте брефической раковины он представлен плотным покрытием толщиной 10–12 мкм (рис. 3, г). Первичный слой имеет тонко гранулированную структуру, величина гранул 0.1 мкм. На поперечном сколе первичного слоя в районе макушки видны мало заметные вертикальные элементы (рис. 4, а). В районе брефической раковины верхний прослой вторичного слоя менее плотный по сравнению с плотным покрытием первичного слоя, он имеет неровный, бугорчатый рельеф, местами видны сильно разрушенные многочисленные поры размером до 1.5 мкм. На поперечных сколах прослоя вторичного слоя видны четкие вертикальные пластины (рис. 4, б). На поверхности прослоя видна четко гранулированная структура, размеры гранул до 0.5 мкм (рис. 4, в). Верхний прослой первичного слоя взрослой части спинной створки характеризуется наличием туберкул и валиковидных образований (рис. 4, г). Первичный слой тонко гранулированный, пористый, размер пор до 1.5 мкм. Сильно разрушенные туберкулы имеют диаметр 100–120 мкм, расстояние между туберкулами 100–120 мкм. На поперечном срезе одной туберкулы можно проследить неправильно-многоугольные очертания, близкие к восьмиугольнику. Эта туберкула состоит из параллельных пластин толщиной до 2 мкм (рис. 4, г). Валиковидные образования хорошо выражены в передней половине створки, имеют длину от 100 до 300 мкм, ширину до 150 мкм и вместе с туберкулами дают слабоволнистый рельеф поверхности спинной створки. На переднем крае полоса шириной около 100 мкм характеризуется отчетливым волнистым рельефом. Выпуклые поверхности расположены неравномерно, имеют ширину от нескольких десятков до 100 мкм. Пережимы между выпуклыми поверхностями узкие, глубокие, создают морщинистость (рис. 4, е). Описание подобного рода морщинистости приводится Вильямсом и Кусаком (Williams, Cusack, 1997), которые объясняют образование этих структур деятельностью периостракума. Периостракум современных дисцинид состоит из апатитовых гранул, покрытых протеином, который находится в растворе acid glycosaminoglycans (GAG). В периоды стресса и напряжения изменения в деятельности периостракума могут дать волнистый рельеф на поверхности первичного слоя. Сходную морщинистость авторы наблюдали у современных Glottidia pyramidata Simpson, 1860, найденной у берегов Флориды, и Lingula adamasi Dall, 1873 из Японии. Морщинистость первичного слоя у ископаемых акротретоидных брахиопод наблюдалась у Angulotreta triangulata Palmer, 1954 из верхнего кембрия Монтано (Williams, Holmer, 1992). Более глубокий прослой вторичного слоя взрослой части спинной створки представлен концентрическими рельефными образованиями шириной 150–170 мкм (табл. VI , фиг. 1, см. вклейку). В поперечном сечении рельефные концентрические образования имеют вид полой трубки неправильной овальной формы; длина сечения 170 мкм, ширина сечения 120 мкм (табл. VI , фиг. 2); толщина всего прослоя до 180 мкм. Более глубокие прослои вторичного слоя на спинной створке можно видеть на поврежденных участках первичного слоя. В районе макушки на месте брефической раковины видна неровная поверхность сферолитового прослоя. Сферолиты имеют правильную шаровидную форму, их размеры 1.6–1.8 мкм. Видны поры размерами 1.5–1.7 мкм (табл. V I, фиг. 3). Вторичный слой в районе повреждения взрослой части спинной створки изображен на табл. V I, фиг. 2. Слой пористый, рыхлый, при разрушении дает тонкие, разрозненные пластинки (табл. VI , фиг. 4).
Рис. 3.
Orbiculoidea magnifica Mergl, 2001, спинные створки, наружные поверхности: а, в – экз. ПИН, № 5609/1: а – целая спинная створка, наружная поверхность, имеются протегулюм и брефическая раковина; шкала 2 мм; в – макушечная часть спинной створки, хорошо виден протегулюм и часть брефической раковины; шкала 200 мкм; б, г – экз. ПИН, № 5609/2: б – почти целая створка, наружная поверхность, имеются протегулюм и брефическая раковина, на первичном слое различаются туберкулы и линии роста; шкала 1 мм; г – первичный слой на брефической раковине; шкала 200 мкм.
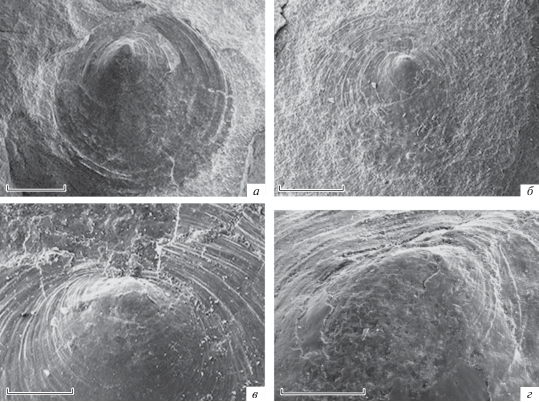
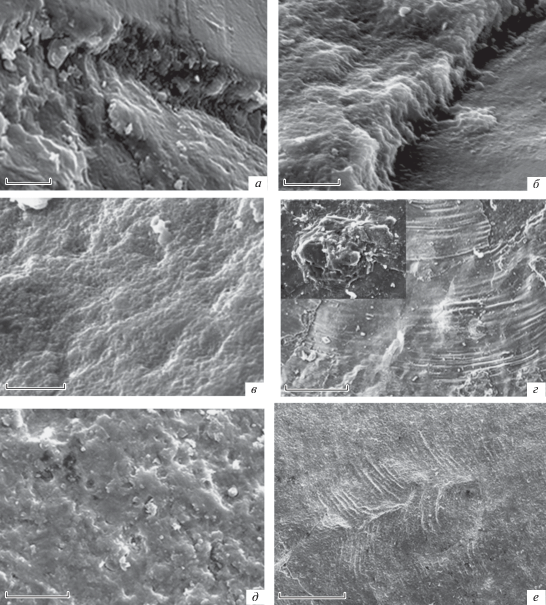
Рис. 4.
Orbiculoidea magnifica Mergl, 2001, наружная поверхность спинной створки: а, б, д, е – экз. ПИН, № 5609/1: а – вертикальные элементы на поперечном сколе первичного слоя; шкала 200 мкм; б – вертикальные элементы и участок сферолитового слоя под первичным слоем; шкала 200 мкм; д – частично разрушенный сферолитовый слой с хорошо выраженными порами; шкала 20 мкм; е – морщинистость в районе первичного слоя; шкала 20 мкм; в, г – экз. ПИН, № 5609/2: в – частично разрушенный сферолитовый слой с хорошо выраженными порами; шкала 20 мкм; г – на месте брефической раковины три слоя, образовавшиеся во время роста раковины, с увеличением дано строение туберкулы, состоящей из тонких пластиночек; шкала 50 мкм.
Брюшная створка правильной округлой формы, уплощенная, имеет размеры 4.2–4.3 мкм. На наружной поверхности видны четко выраженные 6 или 7 концентрических скульптурных образований, возможно, являющихся линиями роста (табл. VI , фиг. 5 ). Туберкулы низкие, правильной округлой формы, выражены нечетко, их размеры до 70 мкм. Наружная поверхность брюшной створки плотная, пористая, размеры пор около 1 мкм. Под плотным слоем наблюдается слой со сферолитами, обычно расположенными беспорядочно (табл. VI , фиг. 6 ). Сферолиты имеют правильную шарообразную форму, их размеры 1–1.2 мкм, на одном участке слоя сферолиты имеют размеры до 2.5 мкм (табл. VI , фиг. 7 ). На поверхности первичного слоя видны радиальные полоски, состоящие из 2–4 рядов ячеек, образовавшихся в результате деятельности пузырьков периостракума. Расстояние между рядами ячеек отличается в пределах нескольких десятков микрон на разном удалении от переднего края створки (табл. VI , фиг. 8 ). Наиболее отчетливо они прослеживаются в передней половине брюшной створки. По предположению Вильямса и др. (Williams et al., 1998), пузырьки были заполнены жидкостью с гликопротеиновыми ядрами, как у современных теребратулид, или жидкостью, близкой к acid glycosaminoglycans (GAGs). Сходная картина рядного расположения ячеек на поверхности первичного слоя наблюдалась на примере Rowellella rugosa Goryansky, 1969 из нижнего ордовика России. У O. cf. magnifica следы от пузырьков могут быть полыми, они имеют форму овалов длиной 4 мкм и шириной 3 мкм (табл. VII , фиг. 1, см. вклейку), или заполненными сферолитами. Часто сферолиты группируются по три, заполняя ячейки, тогда они формируют овальные образования (табл. VII , фиг. 2). Если сферолиты собираются по четыре, они образуют скопления неправильно шаровидной формы (табл. VII , фиг. 3). Размеры скоплений сферолитов, заполняющих ячейки, не превышают 4 мкм, размеры отдельных сферолитов около 1–1.5 мкм. Существенным элементом орнаментации брюшной створки являются рельефные, концентрические образования шириной до 25 мкм, толщиной 10 мкм, которые помещаются в неглубоких желобах (табл. VII , фиг. 4). Они представляют собой плотные образования, состоящие из сцементированного, гранулированного, тонкопористого материала. Гранулы имеют неправильно многоугольную форму, их размеры 0.2–0.3 мкм, размеры пор 0.1 мкм (табл. VII , фиг. 5 ). Листрий сохранился в разной степени на всех брюшных створках, он протягивается от середины брюшной створки, немного не доходя до заднего края створки, средняя длина его 1.5 мкм (табл. VII , фиг. 6 ). Листрий состоит из трех частей – средней в виде стержня с ровными краями и двух боковых частей, имеющих удлиненную, полукруглую форму с вытянутыми задними краями. Средняя часть сзади немного выступает за пределы боковых частей (табл. VII , фиг. 7 ). Листрий выходит из круглого форамена диаметром около 100 мкм (табл. VII , фиг. 8 ).
ЗАКЛЮЧЕНИЕ
В течение длительной геологической истории с ордовика по настоящее время у дисцинид наблюдается неоднократное появление протегулюма. Брефическая раковина, как правило, хорошо сохраняется у всех изученных дисцинид. Г. Фриман и И. Ланделиус (Freeman, Landelius, 1999) отмечали наличие протегулюма у раннедевонского вида Orbiculoidea sp. Изучение среднедевонских дисцинид O. telleri Cleland, 1911, O. sp. 2, “Discina” sp. и “D.” media показало отсутствие протегулюма. У среднедевонского вида Orbiculoidea sp. 2 был обнаружен протегулюм диаметром 90 мкм. Фриман и Ланделиус приводят таблицу наличия брефической раковины и протегулюма у ряда видов из кембрия, ордовика, карбона, одного третичного вида и двух современных видов дисцинид. Авторы пришли к выводу, что в процессе геологической истории могло изменяться в эмбриогенезе время формирования мантии. Подтверждением этого положения служит отсутствие протегулюма у современных дисцинид (Chuang, 1977) и наличие протегулюма у позднеюрских дисцинид (Смирнова и др., 2017а, б). Обнаруженный нами протегулюм у Orbiculoidea cf. magnifica является второй находкой протегулюма в раннем девоне у рода Orbiculoidea.
Основное отличие палеозойских дисцинид, в данном случае раннедевонских Orbiculoidea cf. magnifica и раннекаменноугольных Orbiculoidea nitida, от мезозойских дисцинид заключается в функции периостракума. У палеозойских дисцинид пузырьки периостракума оставляли глубокие следы на всей поверхности первичного слоя, у мезозойских дисцинид такие следы отсутствуют. В cтроении первичного слоя отмечаются существенные отличия, связанные с деятельностью мантии. Рельефные образования на первичном слое, представленные туберкулами и морщинистыми участками, развитые у позднепалеозойских дисцинид, не наблюдались у мезозойских дисцинид. Строение вторичного слоя у дисцинид палеозоя и мезозоя идентично.
Коллекция хранится в Палеонтологическом ин-те им. А.А. Борисяка (ПИН) РАН, № 5609. Структура раковинного вещества изучалась с использованием электронного сканирующего микроскопа TESCAN VEGA IIXMU (TESCAN) в ПИН РАН. Работа выполнялась при поддержке грантов РФФИ №№ 19-04-01027 и 17-04-00324.
Список литературы
Смирнова Т.Н., Ушатинская, Г.Т., Жегалло Е.А., Панченко И.В. Первые находки представителей семейства Discinidae (Brachiopoda, класс Lingulata) в верхнеюрских отложениях Западной Сибири // Палеонтол. журн. 2017а. № 2. С. 26–33.
Смирнова Т.Н., Ушатинская Г.Т., Жегалло Е.А., Панченко И.В. Микроструктура раковинного вещества Discinisca suborbicularis sp. nov. (класс Lingulata) из верхней юры Западной Сибири // Палеонтол. журн. 2017б. № 5. С. 27–36.
Chuang Shou-hwa. Larval development in Discinisca (Inarticulate Brachiopoda) // Amer. Zool. 1997. V. 19. № 1. P. 39–53.
Freeman G., Lundelius. J. Changes in the timing of middle formation and larval life history traits in linguliform and craniiform brachiopods // Lethaia. 1999. V. 32. № 2. P. 197–217.
Mergl M. Lingulate brachiopods of the Silurian and Devonian of the Barrandian (Bohemia Czech Republic) // Acta Musei Nat. Pragae. Ser. B. Hist. Natur. 2001. V. 57. № 1–2. 49 p.
Williams A., Cusack M. Brachiopod (lingulid) shell mediation in clay formation // Lethaia. 1997. V. 29. № 4. P. 349–360.
Williams A., Cusack M., Buckman J. Chemica-structural phylogeny of the discinoid brachiopod shell // Phil. Trans. Roy. Soc. London. Ser. B. 1998. V. 353. № 1378. P. 2005–2038.
Williams A., Holmer L. Ornamentation and shell structure of acrotretoid brachiopods // Palaeontology. 1992. V. 35. Pt 3. P. 657–692.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал