

Палеонтологический журнал, 2019, № 4, стр. 73-86
Остатки динозавров из верхнего мела Крыма
А. О. Аверьянов a, b, c, *, А. В. Лопатин d
a Зоологический институт РАН
Санкт-Петербург, Россия
b Казанский (Приволжский) федеральный университет
Казань, Россия
c Санкт-Петербургский государственный университет
Санкт-Петербург, Россия
d Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: dzharakuduk@mail.ru
Поступила в редакцию 10.07.2018
После доработки 12.07.2018
Принята к публикации 13.09.2018
Аннотация
Переизучение голотипа Riabininohadros weberae из верхнего мела (верхний маастрихт) Крыма (Беш-Кош) позволило определить неизвестные ранее элементы бедренной, таранной и пяточной костей. Этот таксон характеризуется комплексом примитивных признаков, наблюдаемых у игуанодонтий и базальных орнитисхий, и его филогенетическое положение определяется как Styracosterna indet. Вторая находка динозавров из Крыма (Алешино) представляет собой фрагментарный скелет, включающий шейные и грудные позвонки. Он может принадлежать продвинутому игуанодонтиду или примитивному гадрозавроиду. Таким образом, в Крыму в маастрихте существовало не менее двух видов динозавров.
Первая находка динозавра в Крыму была сделана Г.Ф. Вебер в 1934 г. на вершине горы Беш-Кош в окрестностях Бахчисарая (Бахчисарайский р-н, Республика Крым) в морских глауконитовых известняках верхнего маастрихта (Вебер, 1937; Рябинин, 1945, 1946). А.Н. Рябининым были описаны фрагмент бедренной кости, неполные большая и малая берцовые кости, три предплюсневые кости (astragalus, tarsale II и tarsale III), две плюсневые кости (metatarsale II и III) и первая фаланга второго пальца стопы, относящиеся, очевидно, к одному экземпляру (Рябинин, 1945). По определению Рябинина, кости принадлежат птицетазовому динозавру (Ornithischia). Рябинин сравнивал данную находку с известными на то время орнитоподами из позднего мела Европы, преимущественно с Orthomerus transsylvanicus (Nopcsa, 1900) из маастрихта Румынии, и выделил ее в новый вид Orthomerus weberi Riabinin, 1945. Видовое название было исправлено на weberae, поскольку вид был назван в честь Гертруды Федоровны Вебер, известного специалиста по ископаемым иглокожим (Несов, 1995). В настоящее время румынский гадрозавр называется Telmatosaurus transsylvanicus и считается одним из самых примитивных представителей семейства (Weishampel et al., 1993). В последних сводках по динозаврам крымский таксон рассматривается как nomen dubium, а данный материал определяется как Hadrosauridae incertae sedis (Brinkmann, 1988; Horner et al., 2004; Dalla Vecchia, 2012). Первоначально данная находка относилась к датскому ярусу, но позднее был установлен ее позднемаастрихтский возраст (Jeletzky, 1962).
Вторая находка динозавра в Крыму была сделана в долине р. Улаклы близ исчезнувшего ныне с. Алешино (Балта-Чокрак) в Бахчисарайском р-не Республики Крым, располагавшегося на равном расстоянии между селами Скалистое и Глубокий Яр. Здесь в рыхлом слое маастрихтского песчаника на границе с датскими известняками А.И. Трушников в 1965 г. нашел фрагментарный скелет орнитопода, состоящий из частично сочлененных шейных и грудных позвонков и грудных ребер (Лопатин и др., 2018). Скелет принадлежал молодому экземпляру, у которого большинство невральных дуг не срослось с телами позвонков. Данная находка лишь недавно была детально изучена и здесь описывается впервые.
Описанный материал хранится в Палеонтологическом ин-те им. А.А. Борисяка РАН (ПИН) в Москве. Работа выполнена при поддержке Российского научного фонда (проект 19-14-00020). Частично работа выполнена за счет средств субсидии, выделенной в рамках государственной поддержки Казанского (Приволжского) федерального ун-та в целях повышения его конкурентоспособности среди ведущих мировых научно-образовательных центров и Программы фундаментальных исследований Президиума РАН “Эволюция органического мира. Роль и влияние планетарных процессов”. Авторы благодарны Д.В. Григорьеву за фотографирование костей, проф. Г.Д. Зюссу (Национальный музей естественной истории, Вашингтон) – за консультации.
НАДОТРЯД DINOSAURIA
ОТРЯД ORNITHISCHIA
ПОДОТРЯД ORNITHOPODA
ИНФРАОТРЯД ANKYLOPOLLEXIA
ANKYLOPOLLEXIA INCERTAE SEDIS
Род Riabininohadros Ulansky, 2015
Типовой вид – Orthomerus weberae Riabinin, 1945.
Диагноз. Экстенсорная (передняя) борозда между дистальными мыщелками бедренной кости открыта с латеральной стороны. На дистальном латеральном мыщелке бедренной кости обширная депрессия для передней крестообразной связки. Перемычка между дистальными мыщелками бедренной кости расположена близ переднего края кости. Латеральная лодыжка большой берцовой кости заметно длиннее медиальной лодыжки и направлена дистально. Передний восходящий отросток таранной кости расположен близ латерального края. На его передней поверхности отсутствует дорсальная депрессия. Задний восходящий отросток таранной кости отсутствует. Фасетка для малой берцовой кости на пяточной кости каплевидной формы, с заостренным задним концом. Проксимальная поверхность третьей плюсневой кости с глубокой вырезкой вдоль постеромедиального края. Проксимальная фаланга второго пальца стопы с негинглимоидным дистальным суставом и с гребнем на задней поверхности.
Видовой состав. Типовой вид.
Riabininohadros weberae (Riabinin, 1945)
Orthomerus weberi: Рябинин, 1945, с. 5, рис. 1, табл. 1З2 ; 1946, с. 66, рис. 1.
Рис. 1.
Riabininohadros weberae (Riabinin, 1945), голотип ЦНИГР музей, № 5751; Россия, Республика Крым, Беш-Кош; верхний мел (верхний маастрихт): а–г – фрагмент дистального эпифиза левой бедренной кости, вид с латеральной стороны (а), сзади (б), с медиальной стороны (в) и с дистального конца (г); д–и – левая большая берцовая кость, вид сзади (д), с медиальной стороны (е), спереди (ж), с латеральной стороны (з) и с дистального конца (и). Обозначения: cc – кнемиальный гребень; eg – экстенсорная борозда; lc – латеральный мыщелок; lcc – депрессия для передней крестообразной связки; lm – латеральная лодыжка; mc – медиальный мыщелок; mm – медиальная лодыжка.

Таблица 1.
Промеры (в мм) костей стопы Riabininohadros weberae, голотип ЦНИГР музей, № 5751: L – длина кости; PL – длина проксимального эпифиза; PW – ширина проксимального эпифиза; DL – длина дистального эпифиза; DW – ширина дистального эпифиза
| L | PL | PW | DL | DW | |
|---|---|---|---|---|---|
| II плюсневая кость | 203 | 32 | 14 | 35 | 67 |
| III плюсневая кость | 253 | 66 | 107 | 53 | 82 |
| Проксимальная фаланга II пальца | 68 | 60 | 37 | 36 | 63 |
Orthomerus weberae: Несов, 1995, с. 111.
Riabininohadros weberi: Уланский, 2015, с. 5.
Голотип – Центральный научно-исследовательский геолого-разведочный музей им. Ф.Н. Чернышевского (ЦНИГР музей), № 5751, фрагментарный скелет, включающий фрагмент бедренной кости, фрагмент малой берцовой кости, большую берцовую кость, таранную, пяточную, вторую и третью плюсневые кости и проксимальную фалангу второго пальца стопы; Россия, Республика Крым, Бахчисарайский р-н, гора Беш-Кош восточнее г. Бахчисарай; неназванные слои верхнего маастрихта (верхний мел).
Описание (рис. 1–5). Согласно описанию Рябинина (1945), на фрагменте диафиза бедренной кости (ЦНИГР музей № 1/5751) сохранился четвертый трохантер. В настоящее время эта часть кости утрачена. Сохранился также фрагмент дистального конца левой бедренной кости (ЦНИГР музей № 4/5751), который Рябинин принял за таранную кость (astragalus). На этом фрагменте сохранились латеральный мыщелок и передняя часть медиального мыщелка (рис. 1, а–г). Латеральный мыщелок при взгляде с дистальной стороны имеет овальную форму; его задний конец немного отклонен латерально. С латеральной стороны вентральный край латерального мыщелка имеет полукруглую форму. На латеральной поверхности латерального мыщелка примерно посередине расположен мощный вертикальный гребень. Борозда вдоль заднего края этого гребня для m. iliofibularis практически не выражена. На медиальной поверхности латерального мыщелка имеется обширная и глубокая депрессия для передней крестообразной связки (lig. cruciati cranialis; рис. 1, в), у других орнитопод обычно намного меньшая или плохо выраженная. Медиальный мыщелок значительно выступает вперед по сравнению с латеральным. Очевидно, что латеральный мыщелок был существенно короче в переднезаднем направлении, чем медиальный мыщелок. Вследствие этого экстенсорная (передняя) борозда между мыщелками практически полностью открыта с латеральной стороны. Флексорная (задняя) борозда между мыщелками была очень глубокая. Вследствие этого перемычка между дистальными мыщелками располагалась у переднего конца кости, а не посередине, как у большинства орнитопод.
Рис. 2.
Riabininohadros weberae (Riabinin, 1945), голотип ЦНИГР музей, № 5751; Россия, Республика Крым, Беш-Кош; верхний мел (верхний маастрихт): а–в – проксимальный фрагмент правой малой берцовой кости, вид с проксимального конца (а), с латеральной стороны (б) и с медиальной стороны (в); г–ж – левая таранная кость, вид с дорсальной стороны (г), спереди (д), с вентральной стороны (е) и с медиальной стороны (ж); з–к – левая пяточная кость, вид с дорсальной стороны (з), с медиальной стороны (и) и с латеральной стороны (к). Обозначение: aap – передний восходящий отросток.

Рис. 3.
Riabininohadros weberae (Riabinin, 1945), голотип ЦНИГР музей, № 5751; Россия, Республика Крым, Беш-Кош; верхний мел (верхний маастрихт); левая вторая плюсневая кость, вид с дистального конца (а), с проксимального конца (б), с медиальной стороны (в), спереди (г), с латеральной стороны (д) и сзади (е).
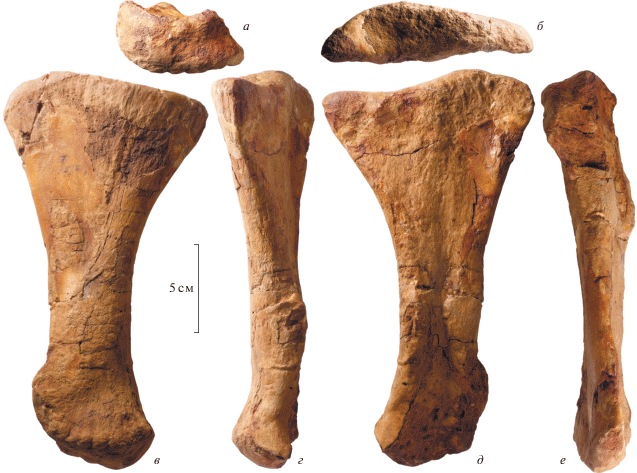
Рис. 4.
Riabininohadros weberae (Riabinin, 1945), голотип ЦНИГР музей, № 5751; Россия, Республика Крым, Беш-Кош; верхний мел (верхний маастрихт); левая третья плюсневая кость, вид с дистального конца (а), с проксимального конца (б), сзади (в), с медиальной стороны (г), спереди (д) и с латеральной стороны (е).
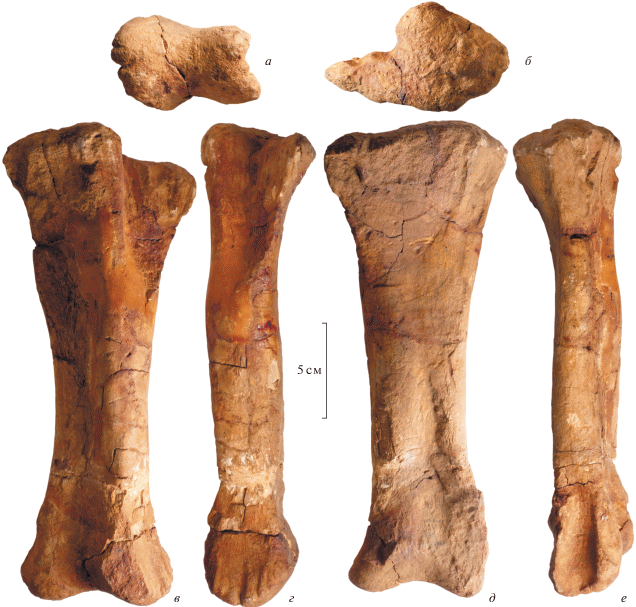
Рис. 5.
Riabininohadros weberae (Riabinin, 1945), голотип ЦНИГР музей, № 5751; Россия, Республика Крым, Беш-Кош; верхний мел (верхний маастрихт); левая проксимальная фаланга второго пальца стопы, вид с дистального конца (а), с проксимального конца (б), с латеральной стороны (в), сзади (г), с медиальной стороны (д) и спереди (е).

На большой берцовой кости (ЦНИГР музей № 2/5751) практически полностью разрушен проксимальный конец (рис. 1, д–и). Основание кнемиального гребня находится на медиальной стороне кости в проксимальной четверти ее длины. На латеральной поверхности диафиза в его проксимальной трети находится отчетливая продольная борозда для кровеносного сосуда (рис. 1, з). Как отметил Рябинин (1945), эта борозда практически идентична борозде на большой берцовой кости “Orthomerus” dolloi из маастрихта Нидерландов (Seeley, 1883, рис. 3, A). Наиболее узкая часть диафиза кости расположена в его дистальной трети. Дистальный эпифиз умеренно расширен. Медиальная лодыжка направлена медиально и немного дистально, латеральная лодыжка – преимущественно дистально. Латеральная лодыжка заметно длиннее медиальной и направлена дистально. На ее латеральной поверхности имеется отчетливо выраженная фасетка для малой берцовой кости. Дистальный конец кости при взгляде с дистального края узкий, треугольной формы, с вогнутой передней и выпуклой задней поверхностями.
В имеющемся материале не удалось найти образец ЦНИГР музей № 3/5751, который Рябинин определял как дистальный конец малой берцовой кости. Однако, среди не определенных Рябининым фрагментов костей удалось идентифицировать проксимальный конец правой малой берцовой кости (рис. 2, а–в). Проксимальный конец широкий в переднезаднем направлении и очень узкий медиолатерально. Проксимальный край кости прямой. Его передний конец вытянут в длинный отросток, обособленный от диафиза кости. На медиальной стороне имеется обширная, но неглубокая депрессия для сочленения с латеральным и постеролатеральным мыщелками большой берцовой кости.
Таранная кость (astragalus, ЦНИГР музей № 6/5751) была описана Рябининым как tarsale III. На кости отсутствуют медиальный и переднемедиальный края (рис. 2, г–ж), которые были разрушены после описания Рябинина (1945: табл. 1, фиг. 5). Дорсальная поверхность кости, контактирующая с латеральной лодыжкой большой берцовой кости, умеренно вогнутая. Вентральная поверхность кости, контактирующая с дистальными тарзальными костями, сильно выпуклая. Массивный, но невысокий дорсальный (восходящий) отросток расположен вдоль переднего края близко к латеральному краю. На передней стороне этого отростка нет четко выраженной дорсальной депрессии, характерной для гадрозавроидов (см., напр., Prieto-Márquez, Norell, 2010, рис. 17, H ). Другим отличием от таранной кости гадрозавроидов является отсутствие заднего восходящего отростка. Вся латеральная поверхность кости занята фасеткой для пяточной кости (calcaneum) бобовидной формы. Переднезадний диаметр кости максимальный в области дорсального отростка. Сохранившийся медиальный край кости приподнят дорсально и образует, очевидно, гребень между фасетками для латеральной и медиальной лодыжек большой берцовой кости. По форме таранная кость очень близка таковой у Mantellisaurus atherfiledensis из баррема-апта Европы (Norman, 1986, рис. 59, C ), отличаясь только выпуклой, а не вогнутой задней поверхностью.
Пяточная кость (calcaneum, ЦНИГР музей № 5/5751) была описана Рябининым как tarsale II. Кость небольшая, полулунной формы при взгляде с медиальной или латеральной стороны (рис. 2, з–к). Дорсальная поверхность, контактирующая с малой берцовой костью, каплевидной формы, с расширенным передним концом и заостренным задним концом. Эта поверхность вогнута в переднезаднем направлении. Медиальная поверхность кости эродирована. Вследствие этого фасетки для контакта с латеральной лодыжкой большой берцовой кости и с таранной костью неразличимы. Каплевидная по форме фасетка для малой берцовой кости у крымского динозавра вполне соответствуeт форме дистальной поверхности малой берцовой кости у Mantellisaurus (Norman, 1986, рис. 59, D ). Среди гадрозароидов наиболее близкое строение пяточной кости наблюдается у Gilmoreosaurus mongoliensis из сантона Китая (Prieto-Márquez, Norell, 2010, рис. 17, I – K).
Вторая плюсневая кость (metatarsale II; ЦНИГР музей № 7/5751) сохранилась полностью (рис. 3). Кость треугольной формы, с расширенным проксимальным концом и закругленным дистальным концом. Проксимальный конец широкий в переднезаднем направлении, почти вдвое шире дистального конца, и очень узкий в медиолатеральном направлении. Наиболее высокая часть проксимального конца расположена близ его заднего края. Кпереди от этого возвышения проксимальная поверхность кости немного вогнутая. Медиальная поверхность кости слабовыпуклая, латеральная поверхность, контактирующая с третьей плюсневой костью, плоская в проксимальной части и слабовогнутая в латеральной части. На медиальной поверхности нет следов крепления первой плюсневой кости. На передней стороне кости, примерно в одной трети ее длины от дистального конца имеется мощное морщинистое возвышение (фланг), к которому, видимо, крепилась связка, соединяющая вторую и третью плюсневые кости. Дистальный конец кости округлый при взгляде с медиальной стороны. Его сочленовная поверхность сильно скошена, так что ее задний край находится намного проксимальнее переднего. Дистальная сочленовная поверхность полулунной формы, с притупленным передним концом, вогнутым латеральным краем и выпуклым медиальным краем. Задний конец слегка подразделен неглубокой бороздкой на медиальный и латеральный мыщелки.
Третья плюсневая кость (metatarsale III; ЦНИГР музей № 8/5751), также полностью сохранилась (рис. 4). Кость сравнительно длинная и стройная. Проксимальный конец немного шире (медиолатерально) дистального конца. Проксимальный конец ромбовидной формы, с вырезкой в постеромедиальном углу. Эта вырезка распространяется дистально вдоль медиальной поверхности кости, где она выклинивается примерно на уровне трети длины от проксимального конца. Латеральная половина проксимального конца кости образует возвышение над медиальной частью. Проксимальный конец кости немного отогнут вперед по сравнению с диафизом. Диафиз кости наиболее узкий (медиолатерально) близ основания дистального эпифиза. Дистальный эпифиз кости слабо расширен медиолатерально. Он образует седловидную поверхность для сочленения с проксимальной фалангой третьего пальца. Эта поверхность немного заходит на переднюю поверхность кости. Медиальный и латеральный мыщелки разделены неглубокими межмыщелковыми бороздами с передней и задней стороны. Медиальный мыщелок отчетливо длиннее латерального в переднезаднем направлении. Медиальная поверхность медиального мыщелка морщинистая, с тремя-четырьмя короткими, но отчетливыми вертикальными гребнями. На латеральной поверхности латерального мыщелка имеется глубокая продольная борозда.
Из фаланг имеется только проксимальная фаланга второго пальца (ЦНИГР музей № 9/5751; рис. 5). Фаланга очень короткая и массивная, ее проксимодистальная длина почти равна переднезаднему диаметру. Проксимальная сочленовная поверхность ромбовидной формы, плоская. Ее медиолатеральный диаметр равен примерно половине переднезаднего диаметра. Медиальная поверхность фаланги слабовогнутая, латеральная выпуклая. Дистальная сочленовная поверхность негинглимоидная, седловидная спереди и гребневидная сзади, где она переходит на заднюю поверхности фаланги. Эта “гребнеобразная тыльная поверхность кости” была отмечена в описании Рябинина (1945, с. 6).
Размеры в мм. Промеры плюсневых костей и проксимальной фаланги второго пальца см. в табл. 1.
ОПИСАНИЕ ФРАГМЕНТАРНОГО СКЕЛЕТА ОРНИТОПОДА ИЗ С. АЛЕШИНО
Шейные позвонки экз. ПИН, № 5619 опистоцельные. От второго шейного позвонка (axis) сохранились левая невральная дуга и остистый отросток (рис. 6, а–в). Диапофиз в виде небольшого бугорка, расположен в центре невральной дуги. Сочленовная поверхность постзигапофиза вытянута в переднезаднем направлении и ориентирована вентролатерально. Отверстие неврального канала сравнительно крупное. На задней стороне невральной дуги, между постзигапофизами и остистым отростком, имеется обширная депрессия. Остистый отросток длинный в переднезаднем направлении. Его передняя часть сильно выступает вперед за уровень презигапофизов (большей частью разрушены), тогда как задняя часть не выдается назад за уровень постзигапофизов. Остистый отросток второго шейного позвонка по форме больше напоминает таковой у Iguanodon bernissartensis (Norman, 1980, рис. 24, a ), чем у Mantellisaurus atherfiledensis (Norman, 1986, рис. 25 ) (оба таксона известны из баррема-апта Западной Европы). Эпипофиз редуцирован примерно в такой же степени, как у Iguanodon (более развит у Mantellisaurus).
Рис. 6.
Styracosterna indet., экз. ПИН, № 5619; Россия, Республика Крым, Алешино; верхний мел (верхний маастрихт): а–в – невральная дуга второго шейного позвонка, вид сверху (а), с латеральной стороны (б) и сзади (в); г–ж – невральная дуга третьего шейного позвонка, вид с дорсальной стороны (г), с латеральной стороны (д), спереди (е) и сзади (ж); з, и – передний шейный позвонок, вид с дорсальной стороны (з) и спереди (и); к, л – передний шейный позвонок, вид сзади (к) и с латеральной стороны (л); м – три сочлененных тела шейных позвонков, вид с вентральной стороны. Обозначения: di – диапофиз; pa – парапофиз; poz – постзигапофиз; prz – презигапофиз.

Изолированная сравнительно длинная невральная дуга без остистого отростка, очевидно, относится к третьему шейному позвонку (рис. 6, г–ж). На дорсальной стороне имеются параллельные продольные гребни, идущие почти до конца постзигапофизных отростков. На дорсальной стороне постзигапофиза этот гребень контактирует с другим, более коротким гребнем, соответствующим, видимо, эпипофизу. Сочленовные поверхности зигапофизов крупные, овальной формы, вытянуты в переднезаднем направлении. Они ориентированы под углом около 45° к горизонтальной поверхности. Презигапофизы расположены на мощных и коротких презигапофизных отростках. Постзигапофизные отростки также мощные, существенно более длинные, расходятся под углом примерно 60°. Между основаниями постзигапофизных отростков расположена сравнительно крупная интерпостзигапофизная пластина. Поперечный отросток очень короткий, направлен преимущественно назад, без четкой сочленовной поверхности диафиза. Крупный презигапофиз на презигапофизном отростке, короткий поперечный отросток и параллельные гребни на дорсальной поверхности невральной дуги характерны для третьего шейного позвонка у Iguanodon bernissartensis (Norman, 1980, рис. 25, c ).
Два целых передних шейных позвонка (рис. 6, з–л) близки по строению и, возможно, являются четвертым и пятым шейными позвонками. Позвонки сравнительно короткие, длина тел позвонков меньше их ширины. Сочленовные поверхности тела позвонка овальной формы, уплощены дорсовентрально. Вентральная сторона тел позвонков разрушена, но, очевидно, на ней имелся продольный гребень. На одном из позвонков сохранился крупный парапофиз, расположенный близ невроцентрального шва, ближе к заднему краю позвонка. Парапофиз овальной формы, вытянут в переднезаднем направлении и помещен на коротком возвышении. Невральный канал крупный, но меньше половины высоты тела позвонка, округлой или овальной формы. На невральной дуге имеются крупные постзигапофизные и поперечные отростки (диапофизы). Сочленовные поверхности презигапофизов расположены в основании поперечных отростков, по бокам от неврального канала. Их сочленовные поверхности ориентированы дорсомедиально. Основания диапофизов, на которых расположены презигапофизы, направлены дорсолатерально. Затем диапофизы резко изгибаются вентролатерально и заканчиваются небольшими сочленовными поверхностями для бугорка ребра. Эта дистальная часть поперечного отростка заметно короче проксимальной части. Постзигапофизные отростки направлены постеролатерально, угол между ними составляет около 90°. Сочленовные поверхности постзигапофизов сохранились на одном из позвонков (рис. 6, к, л). Они крупные, овальной формы, ориентированы вентролатерально. Остистый отросток отсутствует. На обоих позвонках имеется небольшая, но четко выраженная интерпостзигапофизная пластина, расположенная в основании постзигапофизных отростков.
Имеются также изолированные опистоцельные тела трех шейных позвонков, сохранившихся в сочленении (рис. 6, м). На первых двух позвонках задняя сочленовная поверхность существенно шире передней. На вентральной поверхности тел второго и третьего позвонков имеется довольно широкий эродированный продольный гребень. На последнем позвонке вентральный гребень более узкий. На среднем позвонке имеется небольшая фасетка парапофиза близ переднего края и открытого невроцентрального шва. На боковой поверхности тел позвонков имеются крупные питательные отверстия.
Грудные позвонки представлены изолированными телами и невральными дугами с открытыми невроцентральными швами. Почти все невральные дуги относятся к передним грудным позвонкам, поскольку фасетки парапофизов расположены на ножке невральной дуги под поперечным отростком, а не на поперечном отростке. Только на экземпляре, изображенном на рис. 7, л, м, на ножке невральной дуги нет парапофиза, который находился, очевидно, на обломанном поперечном отростке. Этот позвонок, очевидно, относится к задним грудным позвонкам.
Рис. 7.
Styracosterna indet., экз. ПИН, № 5619; Россия, Республика Крым, Алешино; верхний мел (верхний маастрихт): а–в – невральная дуга первого грудного позвонка, вид с дорсальной стороны (а), спереди (б) и с латеральной стороны (в); г–е – невральная дуга переднего грудного позвонка, вид с дорсальной стороны (г), спереди (д) и с латеральной стороны (е); ж–к – невральная дуга переднего грудного позвонка, вид с дорсальной стороны (ж), спереди (з), сзади (и) и с латеральной стороны (к); л, м – невральная дуга заднего грудного позвонка, вид сзади (л) и с латеральной стороны (м); н–р – тело грудного позвонка, вид с дорсальной стороны (н), с вентральной стороны (о), с латеральной стороны (п) и спереди (?) (р). Обозначения: di – диапофиз; pa – парапофиз; poz – постзигапофиз; prz – презигапофиз.

Одна невральная дуга (рис. 7, а–в) принадлежит, очевидно, первому грудному позвонку (Norman, 1980, рис. 35 ). Презигапофиз расположен на поперечном отростке, как на шейных позвонках. Его сочленовная поверхность крупная, овальной формы, вытянута в поперечном направлении. Она расположена под углом 40° к вертикальной оси. В отличие от шейных позвонков, поперечный отросток дистальнее презигапофиза, мощный и направлен дорсолатерально, лишь немного отклоняясь вентрально от направления его проксимальной части. Поперечный отросток имеет в сечении форму параллелограмма, одна из граней которого проходит по середине передней поверхности (рис. 7, б). Постзигапофизные и остистый отростки обломаны. Фасетка парапофиза расположена в основании поперечного отростка.
Более задние грудные позвонки имеют достаточно обособленные презигапофизные отростки, направленные вперед. Их сочленовные поверхности тесно сближены и вытянуты в переднезаднем направлении. Две невральные дуги (рис. 7, г–е) близки по строению и, очевидно, располагались рядом. Невральная дуга сравнительно длинная, с узким в поперечном отношении основанием, что свидетельствует о сравнительно узком теле позвонка. Такие тела позвонков характерны для средних грудных позвонков (Norman, 1980, рис. 35, 38 ). Поперечные отростки мощные, направлены больше назад, чем латерально. Их поперечное сечение четырехгранное, с ребрами на задней и латеральной стороне. Постзигапофизы не сохранились на обоих позвонках. Парапофиз расположен под латеральным гребнем остистого отростка, либо в основании этого гребня перед ним. Между презигапофизами расположен четкий медиальный гребень, который доходит сзади до основания остистого отростка. Дистальная часть остистого отростка обломана.
Постзигапофизы сохранились на двух грудных позвонках (рис. 7, и–м). На заднем грудном позвонке они разделены глубокой щелью, открытой снизу (рис. 7, л), на переднем грудном позвонке – неглубокой депрессией (рис. 7, и). Сочленовные поверхности постзигапофизов крупные, вытянуты в переднезаднем направлении, ориентированы под углом 37° или 60° к вертикальной оси. Экземпляр, изображенный на рис. 7, ж–к, – единственный грудной позвонок, на котором сохранилась полностью невральная дуга, с пре- и постзигапофизами (поперечные и остистые отростки большей частью обломаны). Невральная дуга очень длинная и узкая, с близко расположенными пре- и постзигапофизами. Длина невральной дуги (между концами зигапофизов) вдвое больше длины ножки невральной дуги, которая, очевидно, соответствует длине тела позвонка. Это значит, что постзигапофизы были расположены позади заднего края тела позвонка. Крупный парапофиз расположен в основании поперечного отростка.
Имеются три тела грудных позвонков с открытыми невроцентральными швами (рис. 7, н–р). Тела сжаты в поперечном направлении. Сочленовные поверхности овальной формы, их высота заметно превышает длину. Боковые стороны тел позвонков вогнуты в фронтальной плоскости. На боковой стороне имеются небольшие васкулярные отверстия. На вентральной стороне имеется узкий продольный гребень.
Четыре близких по размерам двухголовчатых ребра (рис. 8, а, б) являются, очевидно, передними грудными ребрами (одно ребро правое, три других – левые). Головка, бугорок и дистальная часть ребра не сохранились. Бугорок ребра, очевидно, располагался на коротком отростке. Тело ребра умеренно изогнуто; его медиальная сторона вогнутая, латеральная – выпуклая, с продольным гребнем. Проксимальный фрагмент правого ребра с крупным овальным бугорком, расположенным на теле ребра без выраженного отростка, и с менее изогнутым в проксимальной части телом ребра (рис. 8, в, г), является, видимо, одним из задних грудных ребер. Почти полное левое одноголовчатое ребро (рис. 8, д, е), с длинным отростком головки ребра и коротким продольным гребнем на латеральной стороне, может быть последним грудным ребром.
Рис. 8.
Styracosterna indet., экз. ПИН, № 5619; Россия, Республика Крым, Алешино; верхний мел (верхний маастрихт): а, б – правое переднее грудное ребро, вид с медиальной (а) и с латеральной стороны (б); в, г – правое заднее грудное ребро, вид с медиальной (в) и с латеральной стороны (г); д, е – левое заднее грудное ребро, вид с латеральной (д) и с медиальной стороны (е). Обозначения: ca – головка ребра; tu – бугорок ребра.
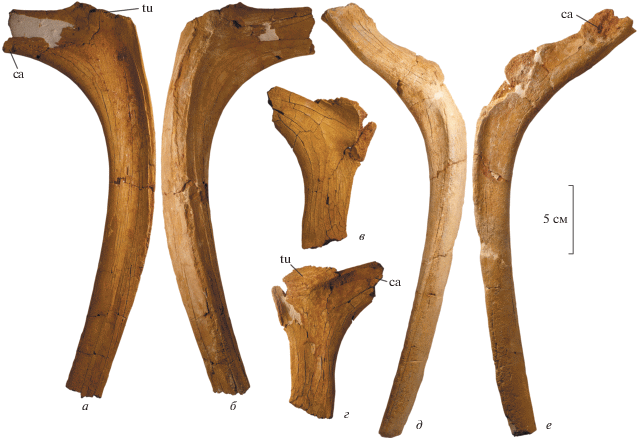
В материале имеются также два небольших фрагмента дорсального края подвздошной кости.
ОБСУЖДЕНИЕ
Посткраниальные остатки крымских динозавров демонстрируют несколько филогенетически значимых признаков, которые обсуждаются в данном разделе.
1) Тела шейных позвонков опистоцельные. Среди орнитопод платиафмицельные или слабо опистоцельные шейные позвонки характерны для гипсилофодонтид, рабдодонтид, дриозаврид и камптозаврид (Norman, 2004; Norman et al., 2004; Carpenter, Wilson, 2008). Шейные позвонки с отчетливо опистоцельными телами являются синапоморфией для клады Styracosterna, включающей игуанодонтид и гадрозавров (Norman, 2014).
2) Тела грудных позвонков амфиплатицельные. У игуанодонтид первый грудной позвонок опистоцельный, более задние – амфиплатицельные. У гадрозаврид тела всех грудных позвонков умеренно опистоцельные (Norman, 1986, 2014).
3) Достаточно необычно строение дистального эпифиза бедренной кости рябининогадра. Передние концы медиального и латерального мыщелка находятся на разных уровнях, передняя межмыщелковая (экстенсорная) борозда полностью открыта с латеральной стороны. У орнитопод исходно имелась неглубокая экстенсорная борозда, которая постепенно углублялась за счет расширения мыщелков в переднезаднем направлении (Barrett et al., 2011). Для Hadrosauridae характерна глубокая экстенсорная борозда, часто закрытая концами мыщелков на переднем конце, которые иногда срастаются, превращая экстенсорную борозду в туннель (Norman, 1998, 2014; Sues, Averianov, 2009). В частности, экстенсорная борозда замкнута в туннель у Telmatosaurus transsylvanicus из маастрихта Румынии (Weishampel et al., 1993, рис. 6, C).
4) В связи с редукцией латерального мыщелка бедренной кости перемычка между дистальными мыщелками расположена близ переднего конца кости, а не посередине, как у большинства орнитопод.
5) Другим необычным признаком рябининогадра является глубокая вырезка на проксимальном конце третьей плюсневой кости вдоль ее постеромедиального края, распространяющаяся дистально на проксимальную треть длины кости. На второй плюсневой кости нет выроста, который мог бы соответствовать этой вырезке. На реконструкции Рябинина (1945: рис. 1, B) третья плюсневая кость расположена посередине второй плюсневой кости. При этом щель между двумя костями достигает максимального размера. По строению второй и третьей плюсневых костей рябининогадр ближе всего к игуанодонтиду Hypselospinus fittoni из валанжина Великобритании (Norman, 2014). У последнего, однако, отмеченная вырезка очень неглубокая.
6) На проксимальной фаланге второго пальца стопы дистальная сочленовная поверхность переходит в гребень на постеромедиальном крае кости. Подобный гребень встречается у примитивных гадрозавроидов, например, у Eolambia caroljonesa из сеномана США и Gilmoreosaurus mongoliensis из сантона Китая (Gilmore, 1933, рис. 20 ; McDonald et al., 2012, рис. 35, C ). Данный гребень, по-видимому, служил для дополнительной опоры задней конечности.
Признаки 1 и 2 позволяют определить находку из Алешино (экз. ПИН, № 5619) до клады Styracosterna. Опистоцелия грудных позвонков может быть плохо выражена у некоторых гадрозаврид. Этот экземпляр, очевидно, принадлежит негадрозавроидному представителю клады Styracosterna.
Строение задней конечности Riabininohadros weberae достаточно своеобразно и не имеет аналогов среди известных игуанодонтий. Перечисленные выше признаки 3–5 являются, видимо, аутапоморфиями Riabininohadros weberae. Другим редким признаком крымского таксона является длинная, направленная дистально латеральная лодыжка большой берцовой кости. Уникальны обширная депрессия для передней крестообразной связки на латеральном мыщелке дистального эпифиза бедренной кости и каплевидная форма фасетки для малой берцовой кости на пяточной кости рябининогадра. Однако, плохая изученность пяточных костей у базальных игуанодонтий пока не позволяет считать последний признак аутапоморфией Riabininohadros weberae. Признак 6 встречается у базальных гадрозавроидов. Также достаточно примитивно строение таранной кости у Riabininohadros, которая отличается от таковой гадрозавроидов латеральным положением переднего восходящего отростка, отсутствием депрессии на его передней стороне и отсутствием заднего восходящего отростка. Совокупность приведенных признаков позволяет определить филогенетическое положение Riabininohadros weberae как негадрозавроидного представителя клады Styracosterna.
Фауна орнитопод позднего мела Европы достаточна своеобразна, она включает в себя как продвинутых представителей Hadrosauridae, так и реликтовые таксоны, такие как базальный гадрозаврид Telmatosaurus и базальные орнитоподы Rhabdodontidae (Dalla Vecchia, 2012; Ősi et al., 2012). Riabininohadros четко отличается от рабдодонтид отсутствием ямки для коллатерального лигамента на проксимальной фаланге стопы, а от Telmatosaurus – незамкнутой экстенсорной бороздой дистального эпифиза бедренной кости (Weishampel et al., 1993, 2003; Ősi et al., 2012). Также опистоцельные шейные позвонки экз. ПИН, № 5619 не позволяют отнести эту находку к рабдодонтидам. Таким образом, Riabininohadros является еще одним реликтовым элементом в позднемеловой фауне Европы, соответствующим по уровню эволюционного развития раннемеловым игуанодонтиям. Очевидно, что этот таксон смог дожить до самого конца позднего мела только благодаря островной изоляции. Riabininohadros weberae и экз. ПИН, № 5619 – самые поздние находки динозавров в России.
Список литературы
Вебер Г.Ф. От Ялты через д. Коккоз до Бахчисарая // Международный Геологический конгресс. XVII сессия. Южная экскурсия. Крымская АССР / Ред. Моисеев А.А. М., Л.: Главн. редакция геол.-развед. лит-ры, 1937. С. 48–58.
Лопатин А.В., Аверьянов А.О., Алифанов В.Р. Новые данные о динозаврах Крыма // Докл. Акад. наук. 2018. Т. 482. № 6. С. 736–738.
Несов Л.А. Динозавры Северной Евразии: новые данные о составе комплексов, экологии и палеобиогеографии. СПб.: Изд-во СПб. гос. ун-та, 1995. 156 с.
Рябинин А.Н. Остатки динозавра из верхнего мела Крыма // Палеонтология и стратиграфия. 1945. № 4. С. 4–10.
Рябинин А.Н. Новые находки ископаемых рептилий в Крыму // Природа. 1946. № 11. С. 65–66.
Уланский Р.Е. Riabininohadros, новый род для гадрозавра из маастрихта Крыма, Россия // Dinologia. 2015. С. 1–10.
Barrett P.M., Butler R.J., Twitchett R.J., Hutt S. New material of Valdosaurus canaliculatus (Ornithischia: Ornithopoda) from the Lower Cretaceous of southern England // Spec. Pap. in Palaeontol. 2011. V. 86. P. 131–163.
Brinkmann W. Zur Fundgeschichte und Systematik der Ornithopoden (Ornithischia, Reptilia) aus der Ober-Kriede von Europa // Documenta Naturae. 1988. Bd 45. P. 1–157.
Carpenter K., Wilson Y. A new species of Camptosaurus (Ornithopoda: Dinosauria) from the Morrison Formation (Upper Jurassic) of Dinosaur National Monument, Utah, and a biomechanical analysis of its forelimb // Ann. Carnegie Museum. 2008. V. 76. № 4. P. 227–263.
Dalla Vecchia F.M. An overview of the latest Cretaceous hadrosauroid record in Europe // Hadrosaurs / Eds. Eberth D.A., Evans D.C. Bloomington, Indianapolis: Indiana Univ. Press, 2012. P. 268–297.
Gilmore C.W. On the dinosaurian fauna of the Iren Dabasu Formation // Bull. Amer. Museum Natur. History. 1933. V. 67. P. 23–78.
Horner J.R., Weishampel D.B., Forster C.A. Hadrosauridae // The Dinosauria. 2nd edn. / Eds. Weishampel D.B., Dodson P., Osmólska H. Berkeley, Los Angeles, L.: Univ. California Press, 2004. P. 438–463.
Jeletzky J.A. The allegedly Danian dinosaur-bearing rocks of the globe and the problem of the Mesozoic-Cenozoic boundary // J. Paleontol. 1962. V. 36. № 5. P. 1005–1018.
McDonald A.T., Bird J., Kirkland J.I., Dodson P. Osteology of the basal hadrosauroid Eolambia caroljonesa (Dinosauria: Ornithopoda) from the Cedar Mountain Formation of Utah // PLoS One. 2012. V. 7. № 10. P. e45712.
Norman D.B. On the ornithischian dinosaur Iguanodon bernissartensis of Bernissart (Belgium) // Mém. Inst. Roy. Sci. Natur. Belg. 1980. T. 178. P. 1–105.
Norman D.B. On the anatomy of Iguanodon atherfieldensis (Ornithischia: Ornithopoda) // Bull. Inst. Roy. Sci. Natur. Belg., Sci. Terre. 1986. V. 56. P. 281–372.
Norman D.B. On Asian ornithopods (Dinosauria: Ornithischia). 3. A new species of iguanodontid dinosaur // Zool. J. Linn. Soc. 1998. V. 122. № 1–2. P. 291–347.
Norman D.B. Basal Iguanodontia // The Dinosauria. 2nd edn. / Eds. Weishampel D.B., Dodson P., Osmólska H. Berkeley, Los Angeles, L.: Univ. California Press, 2004. P. 413–437.
Norman D.B. On the history, osteology, and systematic position of the Wealden (Hastings group) dinosaur Hypselospinus fittoni (Iguanodontia: Styracosterna) // Zool. J. Linn. Soc. 2014. V. 173. № 1. P. 920189.
Norman D.B., Sues H.-D., Witmer L.M., Coria R.A. Basal Ornithopoda // The Dinosauria. 2nd edn. / Eds. Weishampel D.B., Dodson P., Osmólska H. Berkeley, Los Angeles, L.: Univ. California Press, 2004. P. 393–437.
Ősi A., Prondvai E., Butler R.J., Weishampel D.B. Phylogeny, histology and inferred body size evolution in a new rhabdodontid dinosaur from the Late Cretaceous of Hungary // PLoS One. 2012. V. 7. P. e44318.
Prieto-Márquez A., Norell M.A. Anatomy and relationships of Gilmoreosaurus mongoliensis (Dinosauria: Hadrosauroidea) from the Late Cretaceous of Central Asia // Amer. Museum Novit. 2010. № 3694. P. 1–49.
Seeley H.G. On the dinosaurs from the Maastricht beds // Quart. J. Geol. Soc. London. 1883. V. 39. P. 246–253.
Sues H.-D., Averianov A.O. A new basal hadrosauroid dinosaur from the Late Cretaceous of Uzbekistan and the early radiation of duck-billed dinosaurs // Proc. Roy. Soc. Ser. B: Biol. Sci. 2009. V. 276. № 1667. P. 2549–2555.
Weishampel D.B., Jianu C.-M., Csiki Z., Norman D.B. Osteology and phylogeny of Zalmoxes (N. G.), an unusual euornithopod dinosaur from the latest Cretaceous of Romania // J. Syst. Palaeontol. 2003. V. 1. № 2. P. 65–123.
Weishampel D.B., Norman D.B., Grigorescu D. Telmatosaurus transsylvanicus from the Late Cretaceous of Romania: the most basal hadrosaurid dinosaur // Palaeontology. 1993. V. 36. № 2. P. 361–385.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал