

Палеонтологический журнал, 2020, № 4, стр. 99-108
Ископаемые организмы котлинского горизонта верхнего венда северо-запада Русской плиты (Ленинградская область)
Е. Ю. Голубкова a, *, Е. А. Кушим a, А. Б. Тарасенко a, b
a Институт геологии и геохронологии докембрия РАН (ИГГД РАН)
Санкт-Петербург, Россия
b Санкт-Петербургский горный университет (СПГУ)
Санкт-Петербург, Россия
* E-mail: golubkovaeyu@mail.ru
Поступила в редакцию 17.12.2018
После доработки 25.11.2019
Принята к публикации 02.12.2019
Аннотация
Проанализировано вертикальное распределение и реконструированы условия обитания поздневендских микро- (группа микрофоссилии) и макроскопических ископаемых организмов северо-запада Русской плиты. Выделено пять ассоциаций котлинского возраста: одна – обедненная транзитная и четыре – богатые. Проведено монографическое переизучение нитчатых водорослей Aataenia reticularis Gnilovskaya, emend. и Bicuspidata fusiformis Assejeva, emend. Последний таксон сопоставлен с цианобактериями порядка Oscillatoriales. Описан новый вид акритарх Pterospermopsimorpha rigida sp. nov.
ВВЕДЕНИЕ
В результате интенсивных исследований, проводившихся во второй половине XX в., в терригенных отложениях Восточно-Европейской платформы были обнаружены разнообразные ископаемые организмы, что позволило обосновать выделение верхнего отдела венда в объеме редкинского, котлинского и ровенского региональных горизонтов (Розанов, 1973; Волкова и др., 1979; Вендская система…, 1985; Гниловская и др., 1988; Соколов, 1997 и др.). Вместе с тем, полученные на ранних этапах изучения палеонтологические данные зачастую не имеют привязок к конкретных разрезам, а опубликованные фотографии организмов невысокого качества, либо вовсе отсутствуют, что на фоне утраты части типовых коллекций существенно затрудняет их использование для диагностики ископаемых остатков. В связи с этим, особую актуальность приобретают работы, направленные на детальное переизучение палеонтологических коллекций наших предшественников, а также стратотипических разрезов, к числу которых могут быть отнесены венд – кембрийские последовательности, вскрытые в серии обнажений и скважин Ленинградской области.
Авторы благодарны К.Е. Наговицину за помощь в фотографировании микрофоссилий. Исследования проведены при финансовой поддержке проекта РФФИ № 17-05-00498, программы НИР № 0153-2019-0003 и гранта Польского Национального научного центра № 2013/10/A/ST10/00050.
БИОСТРАТИГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА КОТЛИНСКОГО ГОРИЗОНТА
Благодаря возобновлению палеонтологических исследований на северо-западе Русской плиты, в василеостровской свите (скв. Авлога-12R, Коровье-8, Лахта-77-1, Тайцы-2, Шоткуса-1, Уткина Заводь-1, обн. БИ-1) были обнаружены разнообразные по таксономическому составу и превосходные по сохранности микро- (группа микрофоссилии) и макроскопические ископаемые организмы, что позволило обеспечить проведение настоящих исследований. На востоке Ленинградской области василеостровская свита с постепенным переходом залегает на старорусской свите редкинского горизонта и перекрывается ломоносовской свитой (рис. 1). Последняя, по существующим в настоящее время представлениям, относится к верхнему венду (ровенский горизонт)–нижнему кембрию (лонтоваский горизонт) (Якобсон, 2014), или нижнему кембрию (Вербицкий и др., 2012).
Рис. 1.
Вертикальное распространение ископаемых организмов в котлинском горизонте северо-запада Русской плиты: 1 – аргиллиты; 2 – алевролиты; 3 – песчаники; 4 – гравелиты; 5 – ассоциации ископаемых организмов: а – Leiosphaeridia sp.; б – цианобактерии Siphonophycus sp., Oscillatoriopsis sp.; в – Aataenia reticularis Gnilovskaya, emend.; г – Pterospermopsimorpha rigida sp. nov.; д – Vendotaenia antiqua Gnilovskaya; е – Primoflagella speciosa Gnilovskaya; ж – Vanavarataenia insolita Pjatiletov, з – Pomoria rhomboidalis Siverzeva; и – Bicuspidata fusiformis Assejeva, emend. Сокращения: ОСШ – Общая стратиграфическая шкала, РСС – Региональная стратиграфическая схема, МСС – Местная стратиграфическая схема, rd – редкинский горизонт, st – старорусская свита, Ш-1 – скв. Шоткуса-1, К-8 – скв. Коровье-8, Т-2 – скв. Тайцы-2, УЗ-1 – скв. Уткина Заводь-1, Л-77-1 – скв. Лахта-77-1, А-12R – скв. Авлога-12R, БИ-1 – обн. БИ-1 на р. Черная, около пос. Большая Ижора; R – регрессия, T – трансгрессия.
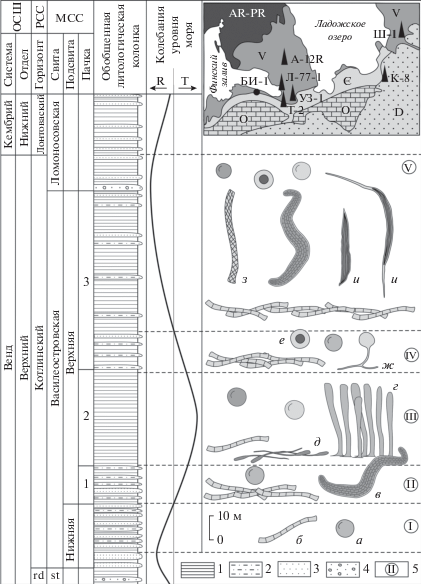
Василеостровская свита подразделяется на нижнюю и верхнюю подсвиты. Нижняя подсвита сложена отчетливым линзовидно-полосчатым переслаиванием алевритистых глин, кварцевых слюдистых алевролитов и песчаников. Мощность пелитовых разностей увеличивается снизу вверх по разрезу. В верхней части подсвиты появляются тонкие прослои сидеритов. Мощность отложений около 40 м. Микрофоссилии на этом уровне отсутствуют, либо представлены транзитными видами Leiosphaeridia crassa (Naumova) emend. Jankauskas, L. jacutica (B.V. Timofeev) emend. Mikhailova et Jankauskas, L. minutissima (Naumova) emend. Jankauskas, L. tenuissima Eisenack, Siphonophycus Schopf emend. Knoll, Swett et Mark (рис. 1, ассоциация I).
В составе верхней подсвиты выделяются три пачки. Нижняя пачка образована тонким градационным чередованием глины и серицитового волнистослоистого алевролита, количество и мощность прослоев которого уменьшается снизу вверх по разрезу. На этом уровне обнаружена вторая ассоциация, в составе которой были выделены нитчатые водоросли Aataenia reticularis Gnilovskaya, emend., Oscillatoriopsis Schopf, Siphonophycus sp., а также морфологически просто устроенные акритархи Leiosphaeridia jacutica, L. minutissima, L. tenuissima, Pterospermopsimorpha rigida sp. nov. и Navifusa Combaz, Lange et Pansart.
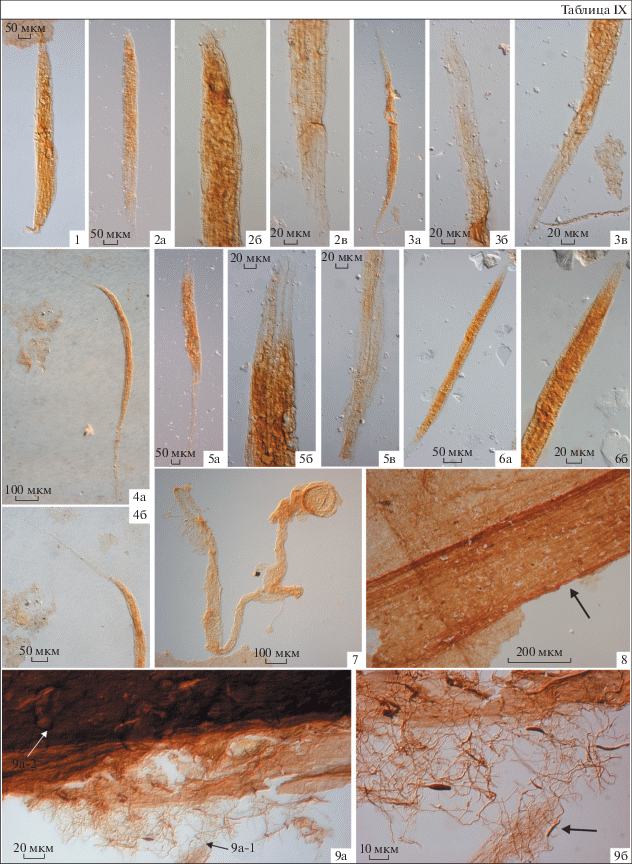
Средняя пачка сложена голубовато-серыми глинами с тонкими (1–10 см) редкими прослоями сидеритов. По всему разрезу распространены органические пленки, неопределимые пиритизированные нити и макроскопические водоросли Vendotaenia antiqua Gnilovskaya (табл. IX , фиг. 8 ; рис. 1, ассоциация III). Фрагменты вендотениевых водорослей обнаружены на субгоризонтальных межслойковых поверхностях и в органомацерате, полученном при растворении образцов на микрофоссилии. Широкое распространение в составе третьей ассоциации имеют актиномицеты Primoflagella speciosa Gnilovskaya (табл. IX , фиг. 9 ), которые, как предполагается, развивались по отмершим и захороненным в толще осадка макроскопическим водорослям и растительным пленкам. Спорадически встречаются характерные трубчатые образования Kanilovia aff. insolita A. Istchenko, вендотениевые водоросли Tyrasotaenia Gnilovskaya, микрофоссилии Leiosphaeridia minutissima, L. tenuissima, Navifusa sp., Siphonophycus sp. и некоторые новые, не описанные пока формы. На этом стратиграфическом уровне в скважине Уткина Заводь-1 дополнительно обнаружены спирально свернутые проблематики Cochleatina Assejeva emend. Burzin, которые, возможно, составляли в прижизненном положении единый организм с трубками Kanilovia insolita (Ищенко, 1983).
Верхняя пачка образована глинами с тонкими линзующимися прослоями алевролитов и тонкозернистых алевритистых песчаников. Мощность верхней подсвиты 120–140 м. В нижней части разреза установлена четвертая ассоциация. Она выделяется по появлению ?хитридиевых грибов Vanavarataenia insolita Pjatiletov (табл. IX , фиг. 7 ), совместно с которыми обнаружены единичные Aataenia reticularis, Pterospermopsimorpha rigida, а также проходящие снизу Leiosphaeridia sp., Navifusa sp. и Siphonophycus sp. В верхней части василеостровской – нижней части ломоносовской свит обнаружена пятая ассоциация, в составе которой доминируют разнообразные нитчатые водоросли Aataenia reticularis, Bicuspidata fusiformis Assejeva, emend., Oscillatoriopsis sp., Pomoria rhomboidalis Siverzeva, Siphonophycus sp., спорадически встречаются акритархи Leiosphaeridia crassa, L. jacutica, L. minutissima, L. tenuissima, Navifusa sp., Pterospermopsimorpha insolita B.V. Timofeev, emend. Mikhailova, P. rigida.
Терригенные последовательности василеостровской свиты сформировались в течение полного трансгрессивно-регрессивного цикла (рис. 1). Предполагается, что в ранневасилеостровское время осадконакопление происходило в мелководных обстановках, выше базы нормальных волнений (БНВ). Алевро-псаммитовые отложения слагали систему подводных валов, а пелитовые частицы оседали в понижениях между ними. На этом уровне были распространены эврифациальные, предположительно планктонные акритархи рода Leiosphaeridia (ассоциация I). Постепенное углубление палеобассейна в начале поздневасилеостровского времени привело к возникновению новых палеоландшафтов. На фоне изменчивого гидродинамического режима в зоне лоскутных песков накапливался алевро-пелитовый материал. На пониженных участках дна были распространены матоподобные образования, сложенные нитчатыми цианобактериями Oscillatoriopsis, Siphonophycus. Планктонные формы были представлены многоклеточными водорослями Aataenia и сфероморфными акритархами (ассоциация II). Дальнейшее углубление бассейна привело к постепенному ослаблению гидродинамики и медленному прерывистому осаждению пелитовых частиц ниже БНВ. Наличие сидеритовых прослоев может свидетельствовать об опреснении палеобассейна. На стадии диагенеза в глинистых илах в восстановительной обстановке происходило массовое захоронение органического вещества и образование псевдоморфоз пирита по нитчатым водорослям. Появление вендотениевой флоры и сопутствующих организмов третьей ассоциации отвечает максимуму трансгрессии (ассоциация III). Начавшаяся во второй половине василеостровского времени регрессия привела к постепенному смещению обстановок осадконакопления в более мелководную и высокодинамичную зону морского бассейна. Алевро-псаммитовые отложения накапливались выше БНВ, формируя подводные вдольбереговые валы. Смена обстановок привела к исчезновению вендотениевых водорослей и широкому, но кратковременному развитию ?хитридиевых грибов Vanavarataenia insolita (ассоциация IV), на смену которым, по мере обмеления бассейна, пришли разнообразные цианобактерии (ассоциация V).
В верхней подсвите василеостровской свиты обнаружены Aataenia, Bicuspidata, Kanilovia, Pomoria, Vanavarataenia, Vendotaenia, что подтверждает котлинский возраст вмещающих отложений (Вендская система.., 1985; Стратиграфическая схема.., 1996; Бурзин, 1998).
АНАЛИЗ ИЗУЧЕННЫХ ТАКСОНОВ
Первый анализируемый в работе таксон, Bicuspidata fusiformis, был обнаружен Е.А. Асеевой (1982) в каниловской серии котлинского горизонта Украины. Эти формы были описаны автором как одиночные трихомы, способные в результате продольного деления распадаться на две нити. Полученные нами данные показывают иное, чем предполагалось ранее, морфологическое строение ископаемых остатков. Водоросли сложены пучками трихомов бесклеточного строения в чехле (табл. IX , фиг. 1–6 ; рис. 2). Предполагается, что в прижизненном положении трихомы были многоклеточными, однако, в процессе фоссилизации их клеточные перегородки были разрушены. В ископаемом материале наблюдаются разнообразные по морфологии экземпляры: от коротких веретеновидных пучков, чехлы в которых плотно прилегают к трихомам (табл. IX , фиг. 1, 2) до форм, внешние оболочки которых сильно вытянуты в длину (табл. IX , фиг. 4 ). Последние, скорее всего, являются “зрелыми” нитями, в то время как формы с короткими трихомами могут быть отнесены к молодым или ювенильным экземплярам. Такое морфологическое строение характерно для нитчатых цианобактерий, близких к современным родам Microcoleus и Schizothrix. Так как в настоящее время не представляется возможным выяснить, сохранился голотип Bicuspidata fusiformis или нет, в качестве номенклатурного типа нами предлагается выделить неотип (табл. IX , фиг. 4 ; Turland et al., 2018, ст. 9.8).
Рис. 2.
Морфологическое строение и формы сохранности изученных микрофоссилий. Обозначения: к – клетка, т – трихом, ч – чехол.
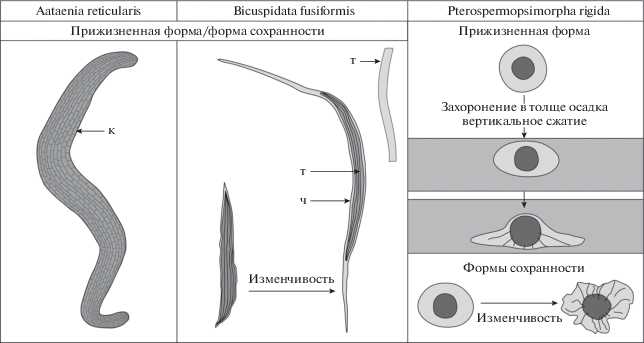
Второй таксон, Aataenia reticularis, был описан М.Б. Гниловской (1976) из котлинской свиты верхнего венда скважины Ф-165-Аа Эстонии. По литературным данным, известно всего одно изображение невысокого качества A. reticularis (голотип), поэтому его морфологическое строение и внутривидовая изменчивость оставались долгие годы не вполне понятными (Гниловская, 1976; Волкова и др., 1979; Вендская система…, 1985; Гниловская и др., 1988). Для восполнения существующих пробелов нами была изучена коллекция постоянных препаратов М.Б. Гниловской, хранящаяся в лаборатории литологии и биостратиграфии ИГГД РАН (г. Санкт-Петербург). Препарат № Аа-138-7, в мацерате которого был выделен голотип, в типовой коллекции отсутствует (утрачен). Вместе с тем, в препарате № Аа-138-10 этой коллекции обнаружен фрагмент нитчатой водоросли, который обладает теми же чертами строения, что и описанный ранее голотип. Этот экземпляр предложен нами в качестве лектотипа (табл. X , фиг. 9 ; Turland et al., 2018, ст. 9.3, 9.4). Необходимо отметить, что экземпляры, выделенные в качестве голотипа и лектотипа, были захоронены на растительных пленках и представляют собой фрагменты линейно вытянутых микроорганизмов, внешнее очертание и внутреннее строение которых плохо различимы.
В результате изучения разрезов Ленинградской обл. нами были обнаружены целые экземпляры Aataenia reticularis средней и хорошей сохранности, что позволило выделить эпитип (табл. X , фиг. 6 ; Turland et al., 2018, ст. 9.9). Анализируемые микрофоссилии сложены многорядными, многоклеточными слоевищами лентовидной формы (рис. 2). Они имеют четкие очертания и характерные плавно закругленные окончания, что позволяет относить их к планктонным формам. При захоронении в толще осадка водоросли сохраняют лентовидное строение и не распадаются на отдельные, более тонкие нити или трихомы (табл. X , фиг. 7, 8 ), что говорит об их более сложной морфологической организации по сравнению с цианобактериями. Однако отсутствие репродуктивных органов, а также каких-либо специальных и характерных структур не позволяет на данном этапе исследования определить их систематическое положение.
Последний рассмотренный в работе таксон, Pterospermopsimorpha rigida sp. nov., представлен двухслойными оболочками, форма которых в ископаемом материале варьирует от сферических или незначительно уплощенных (табл. X , фиг. 1) до сильно деформированных (табл. X , фиг. 4 ). При этом деформации, выраженные в образовании вторичных складок смятия, затрагивают преимущественно внешнюю, более тонкую и эластичную оболочку (рис. 2). Предполагается, что внутреннее тело (или оболочка) было способно сохранять объем, либо подвергалось незначительным деформациям при захоронении в толще осадка, что отличает этот таксон от выделенных ранее видов рода Pterospermopsimorpha. Анализируемые формы представлены морфологически просто устроенными гладкостенными микроорганизмами, что не позволяет определить их биологическую природу.
СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
ОТДЕЛ CYANOBACTERIA
ПОДКЛАСС NOSTOCOBACTERIDAE
ПОРЯДОК OSCILLATORIALES
Род Bicuspidata Assejeva, 1982, emend. nov.
Типовой вид – Bicuspidata fusiformis Assejeva, 1982.
Диагноз. Пучки нитчатых водорослей веретеновидной формы, состоящие из одинаковых по морфологии трихомов в чехле.
Diagnosis emended. Filamentous algae spindle-shaped, consist of uniform trichomata in the sheath.
Видовой состав. Типовой вид.
Сравнение. От позднедокембрийского рода Polythrichoides Hermann, представленного пучками трихомов бесклеточного строения, отличается веретеновидной формой, меньшим числом трихомов в пучке, а также наличием общего чехла. От вендского таксона Talakania Kolosov, сложенного многоклеточными трихомами в чехле, водоросли Bicuspidata отличаются веретеновидной формой чехла и строением трихомов.
Bicuspidata fusiformis Assejeva, 1982, emend. nov.
Табл. IX , фиг. 1–6
Bicuspidata fusiformis: Асеева, 1982, c. 12–13, табл. II , фиг. 7–9 ; Соколов, 1997, с. 109, табл V , фиг. 7, 8 .
Неотип – ИГГД РАН, № I-4; Восточно-Европейская платформа, северо-запад Русской плиты, Балтийско-Ладожская моноклиза, Ленинградская обл., р. Черная, обн. БИ-1 (N 59°56′0.27222′′; E 29°34′0.21845′′), обр. БИ-1-8А, пр. 8А/1; верхний венд, котлинский горизонт, верхняя подсвита василеостровской свиты; обозначен здесь, табл. IX , фиг. 4 .
Diagnosis emended. Uniform straight or slightly curved, unbranched trichomata, enclosed in a sheath. The length of the algae are 540–950 µm, the width are 35–70 μm. The sheath is spindle-shaped, with pointed ends. The number of filaments in the bundle is not constant, averaged 2–5. The trichomata smooth, thin, as a rule, tight to each other. The width of the trichomata are 7–12 µm. The cellular structure of the trichomata not observed.
Описание. Одинаковые по морфологии, прямые или слабо изогнутые, не ветвящиеся трихомы, заключенные в общий чехол. Длина и ширина нитей в чехле составляет, соответственно, 540–950 и 35–70 мкм. Чехол веретеновидной формы, с заостренными окончаниями. Число трихомов в пучке непостоянно, в среднем составляет два–пять. Трихомы гладкие, тонкие, как правило, плотно прилегают друг к другу, шириной 7–12 мкм. Клеточное строение трихомов не выявлено. Одиночные, предположительно планктонные водоросли, не образуют колоний.
Распространение. Украина: Волынь, каниловская свита; Подолия, жарновская и крушановская свиты каниловской серии; Россия, Ленинградская обл., верхняя подсвита василеостровской свиты; верхний венд, котлинский горизонт.
Материал. 42 экз. хорошей и средней сохранности из верхней подсвиты василеостровской свиты скважин Коровье-8 (инт. 237.6–227 м), Лахта-77-1 (инт. 32.24–26.75 м), Тайцы-2 (инт. 179–171 м), Шоткуса-1 (гл. 179.5 м) и обнажения БИ-1 (обр. БИ-1-8А, БИ-1-10, БИ-1-14) на р. Черная Ленинградской обл.
INCERTAE SEDIS
ГРУППА МИКРОФОССИЛИИ
ПОДГРУППА ACRITARCHA EVITT, 1963
Род Pterospermopsimorpha B.V. Timofeev, 1966, emend. Mikhailova et Jankauskas, 1989
Pterospermopsimorpha rigida Golubkova, Kushim et Tarasenko, sp. nov.
Табл. X , фиг. 1–4
Pterospermopsimorpha sp.: Соколов, 1997, с. 109, табл. V , фиг. 4 .
Название вида от rigidus лат. – жесткий.
Голотип – ИГГД РАН, № I-1; Восточно-Европейская платформа, северо-запад Русской плиты, Балтийско-Ладожская моноклиза, скв. Коровье-8, гл. 259.3 м, обр. Кор-8-1, пр. 1/5; верхний венд, котлинский горизонт, верхняя подсвита василеостровской свиты; обозначен здесь, табл. X , фиг. 1.
Объяснение к таблице IX
Фиг. 1–6 . Bicuspidata fusiformis Assejeva, 1982, emend.: 1 – скв. Шоткуса-1, гл. 179.5 м, обр. Шт-14, пр. 14/4; 2, 3, 5, 6 – скв. Тайцы-2, гл. 171 м, пр. 171/1; 4 – неотип № I-4, р. Черная, обн. БИ-1, обр. БИ-1-8А, пр. 8А/1.
Фиг. 7 . Vanavarataenia insolita Pjatiletov, 1985; скв. Лахта-77-1, гл. 26.75 м, обр. L26.75, пр. 26.75/3.
Фиг. 8, 9 а-2. Vendotaenia antiqua Gnilovskaya, 1971.
Фиг. 9 а-1, 9б. Primoflagella speciosa Gnilovskaya, 1979.
Фиг. 8, 9 – скв. Авлога-12R, гл. 123.5 м, обр. GDD200-25: 8 – пр. 25/9, 9 – пр. 25/7.
Россия, Ленинградская обл.; верхний венд, котлинский горизонт, василеостровская свита.
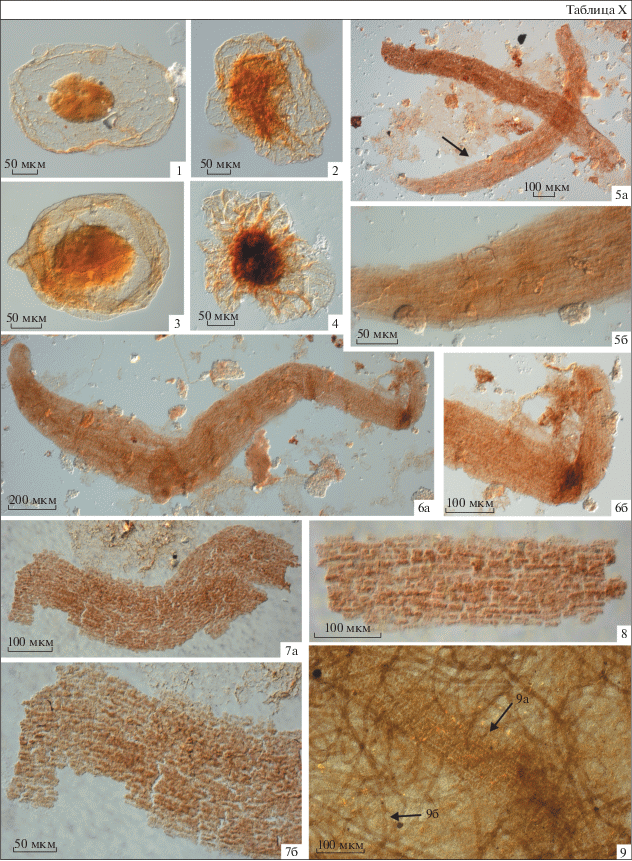
Описание. Микрофоссилии сферической формы, двухслойные. Внешняя оболочка гладкая, тонкая, эластичная, бледно-желтого цвета, часто бывает осложнена радиально расположенными вторичными складками смятия. Ее диаметр у наименее измененных форм составляет 140–305 мкм. Внешний край оболочки от плавно закругленного до сильно деформированного с неровным краем. Внутреннее тело гладкое, темно-желтого, светло-коричневого цвета, плотное, толстостенное, без вторичных складок смятия, сохраняет объем в ископаемом состоянии, диаметром 85–145 мкм.
Сравнение. От других видов рода Pterospermopsimorpha отличается неровным внешним краем оболочки, а также наличием объемного внутреннего тела.
Распространение. Украина, Волынь, каниловская свита; Россия, Ленинградская обл., верхняя подсвита василеостровской свиты; верхний венд, котлинский горизонт.
Материал. 55 экз. хорошей и средней сохранности из верхней подсвиты василеостровской свиты скважин Авлога-12R (гл. 123.5 м), Коровье-8 (инт. 259.6–205.5 м), Лахта-77-1 (инт. 32.24–23.24 м), Тайцы-2 (гл. 233 м) и обнажения БИ-1(обр. БИ-1-8А) на р. Черной Ленинградской обл.
КЛАСС VENDOTAENIOPHYCEAE
ПОРЯДОК VENDOTAENIALES
СЕМЕЙСТВО VENDOTAENIACEAE GNILOVSKAJA, 1988
Род Aataenia Gnilovskaya, 1976, emend. nov.
Типовой вид – Aataenia reticularis Gnilovskaya, 1976.
Диагноз. Многорядные многоклеточные не ветвящиеся лентовидные слоевища с плавно закругленными окончаниями.
Diagnosis emended. Thallus multiseriate, multicellular, unbranched, ribbon-like thallus. The end of the threads are smoothly rounded.
Aataenia reticularis Gnilovskaya, 1976, emend. nov.
Табл. X , фиг. 5–9 а
Aataenia reticularis: Гниловская, 1976, с. 11–12, табл. I , фиг. 6 ; Волкова и др., 1979, с. 41, табл. XLVI , фиг. 7 ; Вендская система .., 1985, с. 120, табл. XXXV , фиг. 1; Гниловская и др., 1988, с. 45–46, табл. VIII , фиг. 3 .
Лектотип – ИГГД РАН, № I-2; Восточно-Европейская платформа, северо-запад Русской плиты, Балтийско-Ладожская моноклиза, Эстония, скв. Ф-165-Аа, инт. 138-140 м, пр. Аа-138-10; верхний венд, котлинский горизонт, верхняя пачка котлинской свиты; обозначен здесь, табл. X , фиг. 9 а.
Эпитип – ИГГД РАН, № I-3; Восточно-Европейская платформа, северо-запад Русской плиты, Балтийско-Ладожская моноклиза, Ленинградская обл., скв. Тайцы-2, гл. 286.3 м, обр. Т-286.3, пр. 286.3/1; верхний венд, котлинский горизонт, верхняя подсвита василеостровской свиты; обозначен здесь, табл. X , фиг. 6 .
Diagnosis emended. Thallus large, unbranched, straight or slightly curved, ribbon-like, multiseriate and multicellular. The ends of the threads are smoothly rounded. The number of rows in the thallus from 12 to 23. The length 1450–2400 µm, the width 150–235 µm. Сells rectangular in shape, elongated, in the characteristic intracellular seals. Cells width 6–12 μm, length 7–20 μm. Cell walls fuzzy.
Описание. Крупные, не ветвящиеся, прямые или слабо изогнутые лентовидные слоевища. Окончания нитей плавно закругленные. Слоевища многорядные, многоклеточные. Наблюдаемое количество рядов в слоевище – 12–23. Общая длина и ширина целых экземпляров составляет, соответственно, 1450–2400 мкм и 150–235 мкм. Клетки однотипные, прямоугольной формы, вытянутые в длину, выделяются по характерным внутриклеточным уплотнениям, более темным по сравнению с цветом самого слоевища (табл. X , фиг. 7, 8 ). Ширина клеток 6–12 мкм, их длина 7–20 мкм. Перегородки клеток нечеткие, скорее всего, были подвержены быстрому посмертному разрушению. Водоросли одиночные, не образуют колонии.
Распространение. Восточно-Европейская платформа, северо-запад Русской плиты, Балтийско-Ладожская моноклиза; Эстония, котлинская свита; Ленинградская обл., василеостровская свита; верхний венд, котлинский горизонт.
Объяснение к таблице X
Фиг. 1–4 . Pterospermopsimorpha rigida sp. nov.: 1, 4 – скв. Коровье-8: 1 – голотип № I-1; гл. 259.3 м, обр. Кор8-1, пр. 1/5; 4 – гл. 249.5 м, обр. Кор8-6, пр. 6/3; 2 – скв. Тайцы-2, гл. 230 м, обр. Т-230, пр. 230/1; 3 – скв. Лахта-77-1, гл. 23.24 м, обр. L23.24, пр. 23.24/3.
Фиг. 5–9 а. Aataenia reticularis Gnilovskaya, 1976, emend.: 5, 6 – скв. Тайцы-2, гл. 286.3 м, обр. Т-286.3: 5 – пр. 286.3/2; 6 – эпитип № I-3; пр. 286.3/1; 7, 8 – скв. Уткина Заводь-1: 7 – гл. 172.9 м, обр. УЗ-172.9, пр. 172.9/1; 8 – гл. 134.9 м, обр. УЗ-26, пр. 26/1; 9а – лектотип № I-2, скв. Ф-165-Аа, инт. 138-140 м, пр. Аа-138-10.
Фиг. 9 б. Siphonophycus sp., скв. Ф-165-Аа, инт. 138-140 м, пр. Аа-138-10.
Фиг. 1–8 . Россия, Ленинградская обл., верхняя подсвита василеостровской свиты; фиг. 9 – Эстония, котлинская свита.
ЗАКЛЮЧЕНИЕ
В результате проведенных исследований реконструированы условия и обстановки осадконакопления в поздневендском палеобассейне северо-запада Русской плиты. В василеостровской свите Ленинградской обл. установлено пять последовательно сменяющих друг друга ассоциаций ископаемых организмов котлинского возраста: одна – обедненная транзитная и четыре – более богатые. Ассоциации выдержаны по площади в пределах изученного региона, что может быть использовано для обоснования возраста вмещающих отложений, а также для детализации местной и региональной стратиграфических схем Восточно-Европейской платформы. Диагностичными таксонами котлинского горизонта являются: Aataenia reticularis, Bicuspidata fusiformis, Kanilovia aff. insolita, Pomoria rhomboidalis, Pterospermopsimorpha rigida, Vanavarataenia insolita, Vendotaenia antiqua. Эти формы имеют определенную фациальную приуроченность в котлинское время, однако не известны в выше- и нижележащих отложениях.
Проведено монографическое переизучение стратиграфически значимых нитчатых водорослей Aataenia reticularis и Bicuspidata fusiformis, последний из которых сопоставлен с цианобактериями порядка Oscillatoriales. В составе формальной подгруппы Acritarcha группы микрофоссилии описан новый вид Pterospermopsimorpha rigida.
Список литературы
Асеева Е.А. Новые виды планктонных водорослей венда Волыни–Подолии / Систематика и эволюция древних растений Украины. Киев: Наук. думка, 1982. С. 5–16.
Вендская система. Историко-геологическое и палеонтологическое обоснование. Т. 1. Палеонтология / Ред. Соколов Б.С., Федонкин М.А. М.: Наука, 1985. 224 с.
Бурзин М.Б. Палеобиогеография позднего венда Русской платформы // Палеогеография венда–раннего палеозоя Северной Евразии. Сб. науч. трудов. Екатеринбург: УрО РАН, 1998. С. 136–146.
Вербицкий В.Р., Вербицкий И.В., Васильева О.В. и др. Государственная геологическая карта Российской Федерации. Масштаб 1 : 1 000 000 (третье поколение). Серия Центрально-Европейская. Листы 0–35 – Псков, (N‑35), 0–36 – Санкт-Петербург. Объяснительная записка. СПб.: Картогр. фабрика ВСЕГЕИ, 2012. 510 с.
Волкова Н.А., Гниловская М.Б., Палий В.В. и др. Палеонтология верхнедокембрийских и кембрийских отложений Восточно-Европейской платформы. М.: Наука, 1979. 212 с.
Гниловская М.Б. Древнейшие Metaphyta // Междунар. геол. конгресс. XXV сессия. Доклады сов. геологов. Палеонтология. Морская геология. М.: Наука, 1976. С. 10–14.
Гниловская М.Б., Ищенко А.А., Колесников Ч.М. и др. Вендотениды Восточно-Европейской платформы. Л.: Наука, 1988. 143 с.
Ищенко А.А. К характеристике вендской водорослевой флоры Приднестровья // Стратиграфия и формации докембрия Украины. Сб. научн. тр. / Ред. Рябенко В.А. Киев: Наук. думка, 1983. С. 181–203.
Микрофоссилии докембрия СССР / Ред. Янкаускас Т.В. Л.: Наука, 1989. 190 с.
Розанов А.Ю. Закономерности морфологической эволюции археоциат и вопросы ярусного расчленения нижнего кембрия. М.: Наука, 1973. 164 с.
Соколов Б.С. Очерки становления венда. М.: КМК, 1997. 156 с.
Стратиграфическая схема вендских отложений Московской синеклизы. Объяснительная записка. М.: Центр. регион. геол. центр, 1996. 46 с.
Тимофеев Б.В. Микрофитологическое исследование древних свит. Л.: Наука, 1966. 237 с.
Якобсон К.Э. Проблемы венда Восточно-Европейской платформы // Региональная геология и металлогения. 2014. № 60. С. 109–116.
Evitt W.R. A discussion and proposals concerning dinoflagellates, hystrichosphaeras and acritarchs // Proc. Natl. Acad. Sci. USA. 1963. V. 49. № 2. P. 158–164.
Turland N.J., Wiersema J.H., Barrie F.R. International Code of Nomenclature for algae, fungi, and plants (Shenzhen Code) adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017. Regnum Vegetabile 159. Glashütten: Koeltz Botanical Books, 2018. 254 p. https://doi.org/10.12705/Code. 2018
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал