

Палеонтологический журнал, 2020, № 4, стр. 109-118
Fagaceae в эоценовой палинофлоре юга Приморского края: новые данные по таксономии и морфологии
Н. Н. Нарышкина a, *, Т. А. Евстигнеева b, **
a Институт географии РАН
Москва, Россия
b ФНЦ Биоразнообразия ДВО РАН
Владивосток, Россия
* E-mail: naryshkina.natali@gmail.com
** E-mail: evstigneeva@biosoil.ru
Поступила в редакцию 14.03.2019
После доработки 03.06.2019
Принята к публикации 03.06.2019
Аннотация
С помощью СЭМ изучены дисперсные пыльцевые зерна Fagaceae из эоценовых отложений юга Приморского края. Выявлено 14 типов пыльцевых зерен Fagaceae: один из них относится к вымершему роду Eotrigonobalanus, четыре типа отнесено к подсемейству Castaneoideae, два типа – к роду Fagus и семь типов – к роду Quercus, среди которых четыре – пыльца листопадных дубов (Quercus), один – вечнозеленых дубов (Cyclobalanoides), и два типа – пыльца вечнозеленых жестколистных дубов (Heterobalanus). Присутствие пыльцы вымершего рода Eotrigonobalanus для изученного региона установлено впервые. Полученные данные свидетельствуют о значительном разнообразии семейства Fagaceae в эоцене Приморского края.
ВВЕДЕНИЕ
Семейство Fagaceae включает 10 родов: Fagus L., Nothofagus Blume, Trigonobalanus Forman, Castanopsis (D. Don) Spach., Castanea L., Lithocarpus Blume, Quercus L., Chrysolepis Hjelmquist, Colombobalanus Lozano, Cyclobalanopsis Oerst. (APG, 2003, 2009). Представители Fagaceae играют важную роль в экосистемах Северного полушария, часто доминируя в умеренных, субтропических и тропических областях с центром разнообразия в тропиках Юго-Восточной Азии (Soepadmo, 1972; Crepet, Daghlian, 1980; Меницкий, 1984, Kubitzki, 1993; Zhou, 1999; Manos, Stanford, 2001; Menitsky, 2005). Разнообразие на видовом уровне (Manos et al., 2001, 2008) равномерно распределяется между сезонными субтропическими и вечнозелеными тропическими лесами Центральной Америки (род Quercus) и лесами южной части континентальной Азии и Малайского архипелага (подсемейство Castaneoideae Oerst.). Ископаемые находки Fagaceae (цветки и плоды Archaefagacea futabensis Takahashi, Friis, Herendeen et Crane из отложений раннего коньяка Японии; Takahashi et al., 2008) известны с конца мелового периода. Пыльцевые зерна Archaefagacea futabensis по морфологическим признакам схожи с таковыми у современных Castaneoideae. Однако заметную роль в растительных сообществах представители Fagaceae стали играть только в палеогене (Grímsson et al., 2015, 2016). В эоцене сформировались все современные роды этого семейства. Наиболее типичными представителями семейства Fagaceae в эоценовых палинофлорах юга Приморского края являлись Quercus gracilis Boitz., Q. graciliformis Boitz., Q. conferta Boitz., Q. forestdalensis Trav., Quercites sparsus (Mart.) Samoil., а также представители Fagus, Castanea и Castanopsis (Баскакова, Громова, 1982; Болотникова, 1988; Верховская, Кундышев, 1989; Павлюткин, Петренко, 2010). Отметим, что исследования пыльцевых зерен с помощью только светового микроскопа не дают полной информации, необходимой для их точной идентификации. Использование сканирующего электронного микроскопа (СЭМ) в оценке ископаемых пыльцевых зерен открывает новые перспективы, как для таксономических, так и для эволюционных и палеоэкологических исследований (Liu et al., 2007; Naryshkina, Evstigneeva, 2009; Denk et al., 2012; Evstigneeva, Naryshkina, 2012, 2013; Tekleva et al., 2014; Grímsson et al., 2015, 2016; Hofmann, Gregor, 2018; и др.).
Цель настоящей работы – выявить особенности морфологического строения ископаемых пыльцевых Fagaceae с помощью СЭМ, определить их более точное систематическое положение и установить таксономическое разнообразие буковых в эоцене юга Приморского края.
Исследование выполнено при финансовой поддержке РФФИ, проект 16-04-01241 и по теме Госзадания №0148-2019-0005 (регистрационный номер 01201352491).
МАТЕРИАЛ И МЕТОДИКА
Материалом для исследования послужили коллекционные образцы из эоценовых отложений, вскрытых скважиной 1956 (рис. 1) в пределах угловской свиты Шкотовской впадины (юг Приморского края). Ранее Н.Б. Верховская и А.С. Кундышев (1989) провели детальный анализ таксономического состава и количественных характеристик спорово-пыльцевых спектров этих отложений.
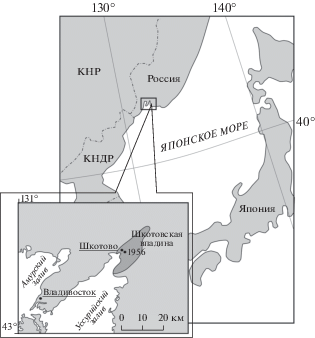
Палинологический материал был изучен с помощью сканирующего электронного микроскопа Zeiss Merlin в Центре коллективного пользования ФНЦ Биоразнообразия ДВО РАН. Для изучения особенностей строения скульптуры ископаемых дисперсных пыльцевых зерен было отобрано семь образцов с преобладанием пыльцы Fagaceae (1956 SH/345, 1956 SH/431, 1956 SH/455, 1956 SH/475, 1956 SH/480, 1956 SH/490, 1956 SH/492). Далее материал отмывали от глицерина и наносили на столик для СЭМ. Затем в напылителе Quorum Q150 объекты в вакууме покрывались хромом. Всего было исследовано 34 фоссильных пыльцевых зерна. Терминология для описания морфологии пыльцевых зерен дана ранее (Halbritter et al., 2018).
Образцы № 5 1956 SH/305, 1956 SH/315, 1956 SH/325, 1956 SH/335, 1956 SH/345, 1956 SH/405, 1956 SH/431, 1956 SH/439, 1956 SH/455, 1956 SH/475, 1956 SH/480, 1956 SH/490, 1956 SH/492 хранятся в палеоботанической коллекции № 5 (1956 SH) “Биоресурсной коллекции” ФНЦ Биоразнообразия ДВО РАН (г. Владивосток).
РЕЗУЛЬТАТЫ ЭЛЕКТРОННО-МИКРОСКОПИЧЕСКОГО ИССЛЕДОВАНИЯ ИСКОПАЕМЫХ ПЫЛЬЦЕВЫХ ЗЕРЕН
В результате детального исследования дисперсных пыльцевых зерен определены следующие представители семейства Fagaceae (табл. 1).
Таблица 1.
Распространение пыльцы Fagaceae по разрезу скважины 1956
| Морфотипы | Глубина, см | ||||||
|---|---|---|---|---|---|---|---|
| 345 | 431 | 455 | 475 | 480 | 490 | 492 | |
| Eotrigonobalanus sp. | + | + | |||||
| Castaneoideae sp. 1 (aff.Castanea) |
+ | + | |||||
| Castaneoideae sp. 2 (aff. Castanopsis cuspidata) |
+ | + | |||||
| Castaneoideae sp. 3 (aff. Castanopsis/Lithocarpus) |
+ | ||||||
| Castaneoideae sp. 4 (aff. Castanopsis/Lithocarpus) |
+ | ||||||
| Fagus sp. 1 (aff. Fagus japonica) |
+ | + | |||||
| Fagus sp. 2 | + | + | |||||
| Quercus sp.1 (aff. Quercus, секция Lobatae) |
+ | + | |||||
| Quercus sp. 2 (aff. Quercus, секция Lobatae) |
+ | + | |||||
| Quercus sp. 3 (aff. Quercus) |
+ | + | + | ||||
| Quercus sp. 4 (aff. Quercus, секция Cerris) |
+ | + | |||||
| Quercus sp. 5 (aff. Cyclobalanoides ) |
+ | ||||||
| Quercus sp. 6 (aff. Heterobalanus) |
+ | + | + | ||||
| Quercus sp. 7 (aff. Heterobalanus, секция Ilex) |
+ | + | |||||
Род Eotrigonobalanus Walther et Kvaček
Eotrigonobalanus sp. (табл. XI , фиг. 1, 2; см. вклейку), исследовано два пыльцевых зерна. Пыльцевые зерна одиночные, в очертании с экватора продолговато-сфероидальные, соотношение длины полярной оси и экваториального диаметра (P/E) = 1.3; полярная ось 20.0–21.1 мкм, экваториальный диаметр около 10.4–15.8 мкм; трехбороздные, борозды длинные, около 16.3 мкм. Скульптура поверхности морщинистая, тонкие скульптурные элементы скручиваются и переплетаются, формируя более крупные морщины – косы. Длина струй 0.7–1.4 мкм, ширина 0.1–0.2 мкм.
Рис. 2.
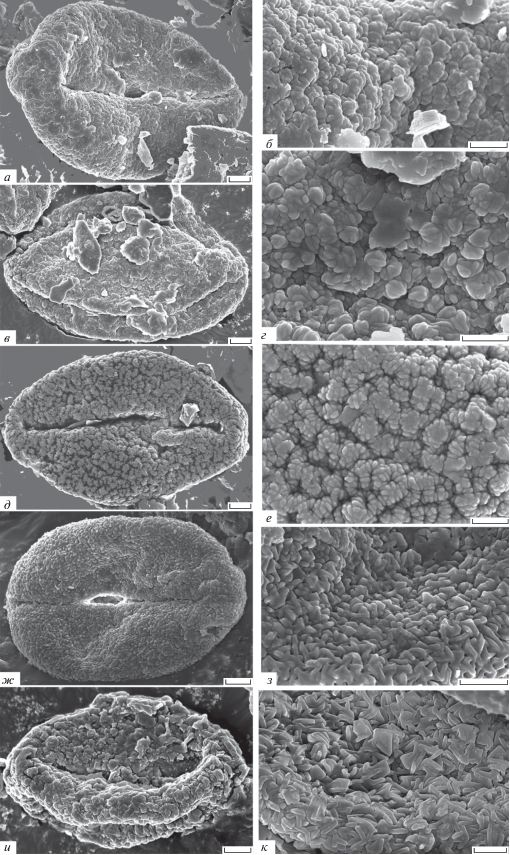
Ископаемые пыльцевые зерна Quercus, ФНЦ Биоразнообразия РАН № 5 1956; юг Приморского края; эоцен; СЭМ: а, б – Quercus sp. 3 (aff. Quercus), инт. 490 см: а – общий вид, б – скульптура поверхности; в, г – Quercus sp. 4 (aff. Quercus, секция Cerris), инт. 480 см: в – общий вид, г – скульптура поверхности; д, e – Quercus sp. 5 (aff. Cyclobalanoides), инт. 431 см: д – общий вид, е – скульптура поверхности; ж, з – Quercus sp. 6 (aff. Heterobalanus), инт. 431 см: ж – общий вид, з – скульптура поверхности; и, к – Quercus sp. 7 (aff. Heterobalanus, секция Ilex), инт. 431 см: ж – общий вид, з – скульптура поверхности. Длина масштабной линейки: а, в, д, ж, и – 2 мкм; б, г, е, з, к – 1 мкм.
Сравнение. Для ископаемых пыльцевых зерен Eotrigonobalanus характерно большое разнообразие размера, формы и типа скульптурных элементов (Grímsson et al., 2015). Пыльца Eotrigonobalanus описана из отложений палеоцена–эоцена Австрии (Hofmann, Zetter, 2010; Hofmann et al., 2011), эоцена юга Русской равнины (Викулин, Нарышкина, 2015), эоцена Западной Гренладии (Grímsson et al., 2015), палеоцена Западной Гренландии (Grimsson et al., 2016), эоцена Германии (Hofmann, Gregor, 2018), олигоцена Центральной Европы (Denk et al., 2012).
Обнаруженные нами пыльцевые зерна Eotrigonobalanus по скульптуре наиболее схожи с пыльцой из отложений олигоцена Центральной Европы (Denk et al., 2012), палеогена (средний эоцен) юга Русской равнины (Викулин, Нарышкина, 2015) и эоцена Западной Гренландии (Grímsson et al., 2015). Это первое описание пыльцы Eotrigonobalanus для юга Приморского края.
ПОДСЕМЕЙСТВО CASTANEOIDEAE OERST.
Castaneoideae sp. 1 (aff. Castanea) (табл. XI , фиг. 3, 4 ), исследовано три пыльцевых зерна. Пыльцевые зерна одиночные, в очертании с экватора продолговатые, P/E = 1.6; полярная ось 11.0–13.3 мкм, экваториальный диаметр 6.6–8.2 мкм; трехборозднооровые, борозды длинные, около 9.0 мкм. Скульптура сглажено морщинистая, образована гладкими, короткими морщинами 0.7 мкм длиной и 0.3 мкм шириной, края скульптурных элементов нечетко выраженные, формируют слабый рельеф. Между морщинами имеются перфорации.
Сравнение. Пыльцевые зерна со сглаженной скульптурой характерны для рода Castanea (Grímsson et al., 2015; Нарышкина, 2016; Нарышкина, Евстигнеева, 2017).
Castaneoideae sp. 2 (aff. Castanopsis cuspidata) (табл. ХI , фиг. 5, 6 ), исследовано три пыльцевых зерна. Пыльцевые зерна одиночные, эллипсоидальные, в очертании с полюса трехлопастные, с экватора продолговатые, P/E = 1.5; полярная ось 18.0–20.2 мкм, экваториальный диаметр 11.1–14.4 мкм; трехборозднооровые, борозды длинные, около 13.6 мкм. Скульптура поверхности морщинистая, образована узкими морщинами до 0.1 мкм и длиной до 1.0 мкм. Скульптурные элементы расположены близко друг к другу и пересекаются между собой.
Сравнение. Ископаемые пыльцевые зерна с подобным типом скульптуры были описаны из голоценовых отложений Японии и Японского моря и определены как Castanopsis cuspidata Schottky (Miyoshi, 1983; Нарышкина, Евстигнеева, 2017).
Castaneoideae sp. 3 (aff. Castanopsis/Lithocarpus) (табл. ХI , фиг. 7, 8 ), исследовано одно пыльцевое зерно. Пыльцевые зерна одиночные, в очертании с экватора продолговато-сфероидальные, P/E = 1.3; полярная ось 15.6 мкм, экваториальный диаметр 12.0 мкм; трехборозднооровые, борозды длинные, около 8.6 мкм. Скульптура поверхности крупно морщинистая, образована морщинами длиной до 1.4 мкм и шириной 0.3 мкм со сглаженным рельефом. Между морщинами имеются перфорации.
Сравнение. Пыльцевые зерна Castaneoi-deae sp. 3 крупнее по размеру, чем Castaneoideae sp. 1 и имеют более длинные скульптурные элементы. В целом скульптура современных пыльцевых зерен Castaneoideae в СЭМ описывается как струйчатая или слабо морщинистая (Van Benthem et al., 1984; Wang, Chang, 1991; Denk et al., 2012; Grímsson et al., 2015). Ранее нами для некоторых видов Castanopsis и Lithocarpus скульптура также описывалась как морщинистая, сформированная широкими и гладкими морщинами (Нарышкина, 2016; Нарышкина, Евстигнеева, 2017).
Castaneoideae sp. 4 (aff. Castanopsis/Lithocarpus) (табл. ХI , фиг. 9, 10 ), исследовано одно пыльцевое зерно. Пыльцевые зерна одиночные, в очертании с экватора продолговатые, P/E = 1.7; полярная ось 15.9 мкм, экваториальный диаметр 9.5 мкм; трехборозднооровые, борозды длинные, около 14.9 мкм. Скульптура поверхности морщинисто-струйчатая, образована морщинами длиной 1.4–2.2 мкм и шириной 0.2–0.3 мкм, с четкими границами, хорошо выраженным рельефом и хаотично пересекающимися.
Сравнение. Обоснованием для выделения Castaneoideae sp. 4 послужило отличие в длине (1.4–2.2 мкм) и степени приподнятости скульптурных элементов (морщин), формирующих поверхность и характер залегания от таковых у Castaneoideae sp. 3 (табл. ХI , фиг. 7, 8 ).
ПОДСЕМЕЙСТВО FAGOIDAE К.KOCH
Род Fagus L.
Fagus sp. 1 (aff. Fagus japonica) (табл. ХII , фиг. 1, 2; см. вклейку), исследовано два пыльцевых зерна. Пыльцевые зерна одиночные, в очертании с экватора округлые, P/E = 1.1; полярная ось 27.8–31.3 мкм, экваториальный диаметр 22.4–27.5 мкм; трехборозднооровые, борозды короткие, поры хорошо заметны. Скульптура поверхности морщинистая, образована широкими (около 0.3 мкм) морщинами, длиной до 0.1 мкм. Морщины между собой перекрещиваются или сливаются, расположены хаотично и имеют вторичную скульптуру в виде поперечных складочек.
Сравнение. Морщинистая скульптура с широкими перекрещивающимися и выступающими морщинами характерна для рода Fagus (Praglowski, 1982; Denk, 2003; Grímsson et al., 2015, 2016). Наибольшее сходство скульптуры исследованных нами ископаемых пыльцевых зерен Fagus sp. 1 было обнаружено со скульптурой пыльцевого зерна Fagus japonica Maxim. (Denk, 2003).
Fagus sp. 2 (табл. ХII , фиг. 3, 4 ), исследовано два пыльцевых зерна. Пыльцевые зерна одиночные, в очертании с экватора округлые, P/E = 1.2; полярная ось 23.6–31.3 мкм, экваториальный диаметр 23.5–26.7 мкм; трехборозднооровые, борозды короткие, поры хорошо заметны. Скульптура поверхности морщинистая, морщины 0.5–0.6 мкм шириной и длиной до 1.9 мкм. Морщины между собой перекрещиваются или сливаются, расположены хаотично и имеют вторичную скульптуру – шероховатую или гранулярную.
Сравнение. Обоснованием для выделения морфотипа Fagus sp. 2 послужило отличие размеров скульптурных элементов (морщин), формирующих поверхность, и различие во вторичной скульптуре. Морщины более длинные и широкие, чем у ископаемых пыльцевых зерен Fagus sp. 1 (табл. ХII , фиг. 1, 2). Вторичная скульптура поверхности пыльцевых зерен Fagus sp. 1 – в виде поперечных складочек, тогда как у Fagus sp. 2 она шероховатая или гранулярная.
Род Quercus L.
Quercus sp. 1 (aff. Quercus, секция Lobatae) (табл. ХII , фиг. 5, 6 ), исследовано два пыльцевых зерна. Пыльцевые зерна одиночные, эллипсоидальные, в очертании с полюса трехлопастные, с экватора продолговатые, P/E = 1.8; полярная ось 26.5–29.2 мкм, экваториальный диаметр 18.2–21.8 мкм; трехбороздные, длина борозд около 24.5 мкм. Скульптура крупноморщинистая, образована крупными, гладкими морщинами шириной более 1.5 мкм. Между морщинами имеются перфорации.
Сравнение. В эоценовых отложениях Гренландии (Grímsson et al., 2015) пыльца с подобным типом скульптуры была определена как Quercus sp. 3 (aff. Group Lobatae).
Quercus sp. 2 (aff. Quercus, секция Lobatae) (табл. ХII , фиг. 7, 8 ), исследовано два пыльцевых зерна. Пыльцевые зерна одиночные, эллипсоидальные, в очертании с полюса трехлопастные, с экватора – продолговатые, P/E = 1.6; полярная ось 28.8–30.0 мкм, экваториальный диаметр 15.4–23.3 мкм; трехбороздные, длина борозд около 20.2 мкм. Скульптура морщинистая, морщины гладкие, шириной до 0.7 мкм. Между морщинами хорошо заметны перфорации.
Сравнение. Этот тип пыльцевых зерен сходен с таковым у ряда видов Quercus секции Lobatae из Северной Америки (Solomon, 1983). Пыльца с подобным типом скульптуры была обнаружена также в эоценовых отложениях Западной Гренландии (Grímsson et al., 2015) и западной части Северной Америки (Bouchal et al., 2014).
Quercus sp. 3 (aff. Quercus) (рис. 2, а, б), исследовано шесть пыльцевых зерен. Пыльцевые зерна одиночные, в очертании с экватора продолговатые, P/E = 1.66; полярная ось 20.8–22.5 мкм, экваториальный диаметр 10.5–18.8 мкм; трехбороздные, борозды длинные, около 18.6 мкм. Скульптура бородавчато-складчатая, образована бородавками, объединяющимися в складки шириной до 0.5 мкм. Между крупными складками имеются перфорации.
Сравнение. Подобный тип скульптуры характерен для пыльцевых зерен некоторых современных видов подрода Quercus L. (листопадные дубы) подсекции Quercus (Нарышкина, 2015).
Quercus sp. 4 (aff. Quercus, секция Cerris) (рис. 2, в, г), исследовано два пыльцевых зерна. Пыльцевые зерна одиночные, в очертании с экватора продолговатые, P/E = 1.7; полярная ось 21.7–25.5 мкм, экваториальный диаметр 12.6–15.3 мкм; трехбороздные, длина борозд 20.0 мкм. Скульптура мелкобородавчатая, бородавки довольно плотно распределены по поверхности и усложнены поперечными морщинками.
Сравнение. Подобный тип скульптуры был описан для пыльцевых зерен Quercus cerris L. подсекции Cerris (Smit, 1973; Van Benthem et al., 1984; Нарышкина, 2015).
Quercus sp. 5 (aff. Cyclobalanoides) (рис. 2, д, е), исследовано два пыльцевых зерна. Пыльцевые зерна одиночные, в очертании с экватора продолговатые, P/E = 1.6; полярная ось 23.3–23.5мкм, экваториальный диаметр 13.7–14.6 мкм; трехбороздные, длина борозд около 18.7 мкм. Скульптура морщинисто-бородавчатая, образована бородавками с хорошо заметной гранулярной структурой. Бородавки, объединяясь, образуют морщинистую скульптуру.
Сравнение. Тип скульптуры (морщинисто-бородавчатый с гранулами) характерен для пыльцы некоторых современных видов вечнозеленых дубов (Liu et al., 2007; Denk, Grimm, 2009; Нарышкина, 2015).
Quercus sp. 6 (aff. Heterobalanus) (рис. 2, ж, з), исследовано четыре пыльцевых зерна. Пыльцевые зерна одиночные, в очертании с полюса трехлопастные, с экватора продолговатые, P/E = 1.7; полярная ось 17.9–21.2 мкм, экваториальный диаметр 10.8–13.6 мкм; трехбороздные, длина борозд около 15.9 мкм. Скульптура мелкоморщинистая, с вертикально или горизонтально направленными кончиками скульптурных элементов (палочковидные).
Сравнение. Ранее нами (Нарышкина, 2015) такой тип скульптуры был описан как Heterobalanus – тип, характерный для пыльцевых зерен видов подрода Heterobalanus Oerst. (вечнозеленые жестколистные дубы).
Quercus sp. 7 (aff. Heterobalanus, секция Ilex) (рис. 2, и, к), исследовано два пыльцевых зерна. Пыльцевые зерна одиночные, в очертании с экватора продолговатые, P/E = 1.9; полярная ось 18.3–22.4 мкм, экваториальный диаметр 10.6–11.5 мкм; трехбороздные, длина борозд около 13.9 мкм. Скульптура мелкоморщинистая, состоит из заостренных, вертикально направленных палочковидных скульптурных элементов.
Сравнение. Скульптура характерна для пыльцевых зерен Q. phillyreoides A. Gray и Q. engleriana Seem. подсекция Ilex (Smit, 1973; Liu et al., 2007; Нарышкина, 2015). В отличие от предыдущего пыльцевого типа Quercus sp. 6, кончики скульптурных элементов, формирующих скульптуру Quercus sp. 7, длиннее и острее.
ОБСУЖДЕНИЕ
В результате детального исследования дисперсных пыльцевых зерен из эоценовых отложений юга Приморского края выделено 14 морфотипов Fagaceae: один из них относится к вымершему роду Eotrigonobalanus, четыре отнесены к подсемейству Castaneoideae, два – к роду Fagus и семь – к роду Quercus.
Использование СЭМ позволило более точно определить систематическое положение ряда таксонов. Так, пыльцевые зерна Castaneoideae sp. 2 и Fagus sp. 1 показали наибольшее сходство с пыльцевыми зернами современных видов – Castanopsis cuspidata и Fagus japonica, соответственно. Пыльцевые зерна Quercus sp. 4 и Quercus sp. 6–7 идентифицированы до уровня секций – Cerris и Ilex.
Интерес представляет сопоставление полученных нами палинологических результатов с данными по макрофлоре Приморского края. Так, пыльцевые зерна вымершего рода Eotrigonobalanus впервые обнаружены и описаны нами для юга Приморского края. В листовой флоре раннего олигоцена Краскино (юг Приморского края) также отмечен Eotrigonobalanus sp. (Павлюткин и др., 2014). Эти данные позволяют утверждать, что Eotrigonobalanus произрастал на данной территории уже в эоцене–олигоцене. Считается, что он был типичным представителем вечнозеленых лесов палеогена и процветал в различных местообитаниях (Walther, 2000).
В палеогеновых флорах юга Приморского края широко представлено подсемейство Castaneoideae: Castaneoideae sp. 1 (aff. Castanea), Castaneoideae sp. 2 (aff. Castanopsis cuspidata), Castaneoideae sp. 3–4 (aff. Castanopsis/Lithocarpus). Пыльцевые зерна Castaneoideae sp. 1 (aff. Castanea) определены только до рода. Макрофлора более разнообразна. Так, в палеогеновых (эоцен – ранний олигоцен) листовых флорах юга Приморского края установлено несколько видов Castanea, например, C. tanaii Huzioka, C. longifolia Borsuk, C. miomollissima Hu et Chaney, C. sachalinensis Kodrul (Красилов, Алексеенко, 1977; Кундышев, Петренко, 1987; Ахметьев, 1993; Аблаев, 2000; Павлюткин, 2007; Павлюткин и др., 2014).
Пыльцевые зерна Castaneoideae sp. 2 схожи с пыльцевыми зернами современного вида C. cuspidata. В раннеолигоценовой листовой флоре Краскино были описаны представители вымершего рода Fagopsis: F. nipponica Tanai и F. primorica Pavlyutkin (Павлюткин и др., 2014). Й. Бучал и др. (Bouchal et al., 2014), описывая пыльцевые зерна Fagopsis из отложений западной части Северной Америки, указывал на их наибольшее сходство с таковыми у современных видов Castaneoideaе, в частности, по типу микроскульптуры они близки к Castanopsis cuspidata. Кроме того, из отложений верхнего олигоцена Южного Китая описана древесина Castanopsis nanningensis Huang, Jin, Quan et Oskolski, имеющая наибольшее сходство с современным видом C. cuspidatа (Huang et al., 2018). Вероятнее всего, в палеогене на территории востока Азии произрастал предковый вид C. cuspidatа.
Типы пыльцевых зерен Castaneoideae sp. 3–4 на данном этапе изучения удалось идентифицировать только до группы родов Castanopsis/Lithocarpus. Листовые отпечатки Castanopsis и Lithocarpus также известны из эоцен–олигоценовых отложений Приморского края (Кундышев, Петренко, 1987; Павлюткин, 2007; Павлюткин и др., 2014).
Род Fagus L. представлен двумя типами пыльцевых зерен – Fagus sp. 1 (aff. Fagus japonica) и Fagus sp. 2. В листовой флоре палеогена юга Приморского края также отмечено несколько видов Fagus, в том числе и близких к F. japonica. Известен, например, F. antipovii Heer (Штемпель, 1926; Аблаев, 2000; Павлюткин, Петренко, 2010; Павлюткин и др., 2014), который, по данным Т. Танаи (Tanai, 1974), является предковым видом для F. crenata Blume и F. japonica. Обнаружен также и F. paleojaponica Tanai et Onoe в отложениях эоцена–нижнего олигоцена (Красилов, Алексеенко, 1977). В раннеолигоценовой листовой флоре Краскино описан F. cf. paleocrenata Okutsu, близкий к двум видам бука: F. paleocrenata и F. stuxbergii (Nath.) Tanai (Павлюткин и др., 2014), а также F. cf. evenensis Cheleb., близкий к современному виду F. engleriana Seem, произрастающему в Центральном Китае. В настоящее время F. japonica относится к подроду Engleriana и произрастает на территории Корейского п-ова и в Японии (о-ва Хонсю, Кюсю и Сикоку). Вероятнее всего, в палеогене на юге Приморского края уже произошла дифференциация буков на уровне подродов.
Род Quercus в эоценовой палинофлоре юга Приморского края среди Fagaceae играет более заметную роль и насчитывает семь морфотипов пыльцевых зерен. Он представлен тремя линиями: листопадные дубы подрода Quercus – Quercus sp. 1–4, вечнозеленые дубы подрода Cyclobalanoides – Quercus sp. 5 и вечнозеленые жестколистные дубы подрода Heterobalanus – Quercus sp. 6–7. Особый интерес вызывают пыльцевые зерна Quercus sp. 1 и sp. 2. По морфологическим признакам выделенные морфотипы близки к некоторым видам секции Lobatae североамериканских красных дубов. Пыльцевые зерна с подобным типом скульптуры были идентифицированы с помощью СЭМ также в эоценовых отложениях Гренландии, Северной Европы и западной части Северной Америки (Bouchal et al., 2014; Grímsson et al., 2015; Denk et al., 2017). В эоценовой листовой флоре юга Приморского края описан Q. kobatakei Tanai et Yokoyama (Алексеенко, Красилов, 1980), по строению эпидермы близкий к некоторым видам подродов Protobalanus (юг Северной Америки) и Erythrobalanus (Средиземноморье). Пыльцевые зерна Quercus sp. 3 и sp. 4 (секция Cerris) могут рассматриваться как индикаторы присутствия в палеофлорах листопадных дубов.
По листовой флоре палеогена (эоцен–олигоцен) юга Приморского края к секции Cerris отнесены ископаемые Q. pseudocastanea Goepp. (Ахметьев, Манчестер, 2000), Q. ussuriensis Krysht. (Ахметьев, 1988; Tanai, Uemura, 1994; Фотьянова, 1997; Павлюткин и др., 2014) и Q. kraskinensis Pavlyutkin (Павлюткин, 2015).
Представители вечнозеленых жестколистных дубов Heterobalanus (в том числе, секция Ilex Loud.), вероятно, также присутствовали в растительности эоцена–олигоцена юга Приморского края. Пыльца этих дубов была идентифицирована нами как в нижней, так и верхней части разреза (табл. 1). Это подтверждают и данные по макрофлоре. Так, листья Q. chekryzhovii Pavlyutkin из раннеолигоценовой флоры Краскино (юг Приморского края) сравнивались с современным Q. incana Roxb. – представителем секции Ilex (Павлюткин, 2015). В этой же флоре был обнаружен отпечаток листа Quercus sp., имеющий сходство с современными видами Q. lanata Smith и Q. franchetii Scan, также входящими в состав секции Ilex (Павлюткин и др., 2014).
Таким образом, результаты исследования дисперсных пыльцевых зерен хорошо сопоставимы с данными по макрофлоре, что свидетельствует о значительном разнообразии семейства Fagaceae в эоценовой растительности Приморского края. В составе эоценовых флор Приморья были представлены почти все роды семейства: Fagus, Castanea, Castanopsis, Lithocarpus, Quercus, а также вымерший род Eotrigonobalanus. Наибольшее разнообразие отмечено для рода Quercus, что подтверждает мнение Л.И. Фотьяновой (1997) о дифференциации на подродовые категории рода Quercus к позднему эоцену на территории Востока Азии.
Список литературы
Аблаев А.Г. Биостратиграфия палеогена побережья юга Приморья. Владивосток: Дальнаука, 2000. 117 с.
Алексеенко Т.М., Красилов В.А. Строение эпидермиса и систематическое положение дубов с лопастными листьями из миоцена Приморья // Палеонтол. журн. 1980. № 3. С. 118–124.
Ахметьев М.А. Кайнозойские флоры Восточного Сихотэ-Алиня. М.: ГИН АН СССР, 1988. 48 с.
Ахметьев М.А. Фитостратиграфия континентальных отложений палеогена и миоцена внетропической Азии. М.: Наука, 1993. 143 с. (Тр. ГИН РАН. Вып.475).
Ахметьев М.А., Манчестер С.Р. Новый вид Paleocarpinus (Betulaceae) из палеогена Восточного Сихотэ-Алиня // Палеонтол. журн. 2000. № 4. С. 107–112.
Баскакова Л.А., Громова Н.С. Фитостратиграфическое расчленение палеогеновых отложений Юго-Западного Приморья // Сов. геол. 1982. № 11. С. 68–77.
Болотникова М.Д. Палинологическая характеристика и возраст угленосных отложений Чернышевского буроугольного месторождения (Южное Приморье) // Тихоокеан. геол. 1988. № 4. С. 101–105.
Верховская Н.Б., Кундышев А.С. Физиономические особенности спорово-пыльцевых спектров и их использование в стратиграфии // Кайнозой Дальнего Востока. Владивосток: ДВО АН СССР, 1989. С. 128–134.
Викулин С.В., Нарышкина Н.Н. Представители семейства Буковых (Fagaceae) в палинофлоре палеогена юга Русской равнины // Геология и биоразнообразие мезозойско-кайнозойских отложений юга России. Матер. Всеросс. научно-практ. конф. 20-24 июля 2015 г. / Ред. Пинчук Т.Н. Горячий Ключ, 2015. С. 50–56.
Красилов В.А., Алексеенко Т.М. Смена растительных сообществ в палеогене и неогене Южного Приморья / Палеоботаника на Дальнем Востоке. Владивосток: ДВНЦ АН СССР, 1977. С. 7–17.
Кундышев А.С., Петренко Т.И. О возрасте Болотнинской ископаемой флоры Южного Приморья // Палинология Востока СССР. Владивосток: ДВНЦ АН СССР, 1987. С. 53–59.
Меницкий Ю.Л. Дубы Азии. Л.: Наука, 1984. 315 с.
Нарышкина Н.Н. Морфология пыльцы восточноазиатских представителей рода Quercus (Fagaceae) // Ботан. журн. 2015. Т. 100. № 9. С. 873–885.
Нарышкина Н.Н. Морфология пыльцевых зерен некоторых видов термофильных буковых (Fagaceae) // Бюлл. БСИ ДВО РАН [Электронный ресурс]: науч. журн. Вып. 15. Владивосток: Ботан. сад–ин-т ДВО РАН, 2016. С. 54–56.
Нарышкина Н.Н. Евстигнеева Т.А. Пыльца Castaneoidae (Fagaceae) из голоценовых отложений Японского моря // Палеонтол. журн. 2017. № 3. С. 97–106.
Павлюткин Б.И. Эоценовая усть-давыдовская флора юга Приморья. Владивосток: Дальнаука, 2007. 148 с.
Павлюткин Б.И. Род Quercus (Fagaceae) в раннеолигоценовой флоре Краскино (Приморский край) // Палеонтол. журн. 2015. № 6. С. 103–110.
Павлюткин Б.И. Петренко Т.И. Стратиграфия палеоген–неогеновых отложений Приморья. Владивосток: Дальнаука, 2010. 164 с.
Павлюткин Б.И., Чекрыжов И.Ю., Петренко Т.И. Геология и флоры нижнего олигоцена Приморья. Владивосток: Дальнаука, 2014. 236 с.
Фотьянова Л.И. Лопастно-зубчатые дубы в эоцене Восточной Азии // Палеонтол. журн. 1997. № 2. С. 103–112.
Штемпель Б.М. Угловский район. Отчет о геологическом исследовании осенью 1924 г. // Матер. геол. полезн. ископ. Дальнего Востока. 1926. № 45. С. 1–88.
APG II. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II // Bot. J. Linn. Soc. 2003. V. 141. № 4. P. 399–436.
APG III. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III // Bot. J. Linn. Soc. 2009. V. 161. № 2. P. 105–121.
Bouchal J., Zetter R., Grímsson F., Denk T. Evolutionary trends and ecological differentiation in early Cenozoic Fagaceae of western North America // Amer. J. Bot. 2014. V.101. P. 1–18.
Crepet W.L., Daghlian Ch.P. Castaneoid inflorescences from the Middle Eocene of Tennessee and diagnostic Value of pollen (at the subfamily level) in the Fagaceae // Amer. J. Bot. 1980. V. 67. P. 739–757.
Denk T. Phylogeny of Fagus L. (Fagaceae) based on morphological data // Plant Syst. Evol. 2003. V. 240. P. 55–81.
Denk T., Grimm G.W. Significance of pollen characteristics for infrageneric classification and phylogeny in Quercus (Fagaceae) // Int. J. Plant Sci. 2009. V. 170. P. 926–940.
Denk T., Grimm G.W., Manos P.S. et al. An updated infrageneric classification of the oaks: review of previous taxonomic schemes and synthesis of evolutionary patterns // Oaks Physiological Ecology. Exploring the Functional Diversity of Genus Quercus L. V. 7 / Eds. Gil-Pelegrín E., Peguero-Pina J., Sancho-Knapik D. Cham: Springer, 2017. P. 13–38.
Denk T., Grímsson F., Zetter R. Fagaceae from the early Oligocene of Central Europe: Persisting new world and biogeographic links // Rev. Palaeobot. Palynol. 2012. V. 169. P. 7–20.
Evstigneeva T.A., Naryshkina N.N. Holocene vegetation changes on the north-eastern coast of the Korean Peninsula based on the palynological data // Acta Palaeobot. 2012. V. 52. № 1. P. 147–155.
Evstigneeva T.A., Naryshkina N.N. Mid-Holocene vegetation and environments on the northeastern coast of the Korean Peninsula // Botanica Pacifica. 2013. V. 2. № 1. P. 27–34.
Grímsson F., Grimm G.W., Zetter R., Denk T. Cretaceous and Paleogene Fagaceae from North America and Greenland: evidence for a Late Cretaceous split between Fagus and the remaining Fagaceae // Acta Palaeobot. 2016. V. 56. № 2. P. 247–305.
Grímsson F., Zetter R., Grimm G.W. et al. Fagaceae pollen from the early Cenozoic of West Greenland: revisiting Engler’s and Chaney’s Arcto-Tertiary hypotheses // Plant Syst. Evol. 2015. V. 301. P. 809–832.
Halbritter H., Ulrich S., Grímsson F. et al. Illustrated Pollen Terminology. Springer, 2018. 483 p.
Hofmann C.-C., Gregor H.-J. Scanning electron microscope and light microscope investigations of pollen from an atypical mid-Eocene coal facies in Stolzenbach mine (PreußenElektra) near Borken (Kassel, Lower Hesse, Germany) // Rev. Palaeobot. Palynol. 2018. V. 252. P. 41–63.
Hofmann C.-C., Mohamed O., Egger H. A new terrestrial palynoflora from the Palaeocene / Eocene boundary in the northwestern Tethyan realm (St. Pankraz, Austria) // Rev. Palaeobot. Palynol. 2011. V. 166. P. 295–310.
Hofmann C.-C., Zetter R. Upper Cretaceous sulcate pollen from the Timerdyakh Formation, Vilui Basin (Siberia) // Grana. 2010. V. 49. P. 170–193.
Huang L., Jin J., Quan C., Oskolski A.A. Mummified fossil woods of Fagaceae from the upper Oligocene of Guangxi, South China // J. Asian Earth Sci. 2018. V. 152. P. 39–51.
Kubitzki K. Fagaceae // The families and genera of vascular plant. V. 2 / Eds. Kubitzki K.J., Rohwer G., Bittrich B. N.Y.: Springer, 1993. P. 301–309.
Liu Y.-S., Zetter R., Ferguson D.K., Mohr B.A.R. Discriminating fossil evergreen and deciduous Quercus pollen: A case study from the Miocene of eastern China // Rev. Palaeobot. Palynol. 2007. V. 145. P. 289–303.
Manos P.S., Cannon C.H., Oh S.H. Phylogenetic relationships and taxonomic status of the paleoendemic Fagaceae of western North America: recognition of a new genus, Notholithocarpus // Madrono. 2008. V. 55. P. 181–190.
Manos P.S., Stanford A.M. The biogeography of Fagaceae: tracking the Tertiary history of temperate and subtropical forests of the Northern Hemisphere // Intern. J. Plant Sci. 2001. V. 162. P. 77–93.
Manos P.S., Zhou Z.-K., Cannon C.H. Systematics of Fagaceae: phylogenetic test of reproductive train evolution // Intern. J. Plant Sci. 2001. V. 162. P. 1361–1379.
Menitsky Y.L. Oaks of Asia. Enfield, New Hampshire: Sci. Publ., 2005. 549 p.
Miyoshi N. Pollen morphology of the genus Castanopsis (Fagaceae) in Japan // Grana. 1983. V. 22. P. 19–21.
Naryshkina N.N., Evstigneeva T.A. Sculpture of pollen grains of Quercus L. from the Holocene of the south of the Sea of Japan // Paleontol. J. 2009. V. 43. № 10. P. 1309–1315.
Praglowski J. Fagaceae L.: Fagoideae // World Pollen Spore Flora. V. 11. Stockholm: Almqvist and Wiksell, 1982. P. 1–28.
Smit A. A scanning electron microscopical study of the pollen morphology in the genus Quercus // Acta Bot. Neerl. 1973. V. 22. P. 655–665.
Soepadmo E. Fagaceae // Flora Malesiana. Ser. 1. 1972. V. 7. Pt 2. P. 265–401.
Solomon A.M. Pollen morphology and plant taxonomy of red oaks in Eastern North America // Amer. J. Bot. 1983. V. 70. P. 495–507.
Takanashi M., Friis E.M., Herendeen P.S., Crane P.R. Fossil flowers of Fagales from the Kamikitaba locality (Early Coniacian; Late Cretaceous) of Northeastern Japan // Intern. J. Plant Sci. 2008. V. 169. P. 899–907.
Tanai T. Evolutionary trend of the genus Fagus around the northern Pacific Basin // Symp. on Origin, Phytogeogr. of Angiosperms. Birbal Sahni Inst. Palaeobot., Spec. Publ. 1974. V. 1. P. 62–83.
Tanai T., Uemura K. Lobed oak leaves from the Tetriary of East Asia with reference to the oak phytogeography of the Northern Hemisphera // Trans. Proc. Paleontol. Soc. Japan. N.S. 1994. № 4. P. 343–365.
Tekleva M.V., Naryshkina N.N., Evstigneeva T.A. Fine structure of Quercus pollen from the Holocene sediments of the Sea of Japan // Plant. Syst. Evol. 2014. V. 300. № 8. P. 1877–1893.
Van Benthem F., Clarke G.C.S., Punt W. Fagaceae. The Northwest European pollen flora // Rev. Palaeobot. Palynol. 1984. V. 42. P. 87–110.
Walther H. Floristic relationship between North and Central America and Europe in the Eocene // Acta Univ. Carol. Geol. 2000. V. 44. P. 51–57.
Wang P.-L., Chang K.-T. The pollen morphology in relation to taxonomy and phylogeny of Fagaceae // Acta Phytotaxon. Sin. 1991. V. 29. № 1. P. 60–62.
Zhou Z.K. Fossils of the Fagaceae and their implications in systematics and biogeography // J. Syst. Evol. 1999. V. 37. P. 369–385.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал