

Палеонтологический журнал, 2020, № 5, стр. 26-37
Новые аммоноидеи из разреза пограничных отложений девона и карбона в Берчогуре (Западный Казахстан)
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Казанский государственный университет
Казань, Россия
* E-mail: 44svnikol@mail.ru
Поступила в редакцию 25.11.2019
После доработки 12.12.2019
Принята к публикации 12.12.2019
Аннотация
Изучены новые и переизучены старые коллекции аммоноидей из пограничных отложений девона и карбона разреза Берчогур в Западном Казахстане. Аммоноидеи происходят из генозоны Acutimitoceras, из слоев одновозрастных со штоккумской фауной Германии. Комплекс в основном включает виды родов Imitoceras и Acutimitoceras, с разнообразной формой начальных оборотов. Описаны новые виды Acutimitoceras alabasense и A. dzhanganense.
ВВЕДЕНИЕ
Разрез Берчогур на территории Западного Казахстана – один из немногих разрезов пограничных отложений девона и карбона с аммоноидеями на Южном Урале, включая его южное окончание – Мугоджарские горы. Аммоноидей из этого местонахождения изучали Л.С. Либрович (1940), Е.А. Балашова (1953), Л.Ф. Кузина (Barskov et al., 1984; Кузина, 1985; Барсков и др., 1988) и Б.И. Богословский (Фауна и биоcтратиграфия…, 1987). Все исследователи отмечали высокую ценность разреза в решении вопроса о границе девона и карбона, поскольку, помимо аммоноидей, в отложениях разреза Берчогур были собраны конодонты, фораминиферы, споры и пыльца, брахиоподы, кораллы, конулярии, гастроподы, трилобиты (Симаков и др., 1985; Фауна и биостратиграфия…, 1987; Барсков и др., 1988). Изучение комплексов этих организмов показало, что аммоноидей из пограничных слоев девона и карбона в этом разрезе следует относить к самым низам карбона (Фауна и биостратиграфия…, 1987), поскольку здесь установлена генозона Acutimitoceras. Разрез Берчогур интенсивно изучался в 80-х гг. прошлого века, но после принятия в 1991 г. границы девона и карбона по первому появлению конодонтов вида S. sulcata со стратотипом в разрезе La Serre (Франция), работы на нем были приостановлены. С 2009 г. граница девона и карбона вновь пересматривается, потому что в стратотипическом разрезе конодонты S. sulcata были обнаружены ниже установленного уровня, непосредственно выше литологической границы (Kaiser et al., 2009). Поскольку в подстилающих отложениях в La Serre конодонтов нет, установить там уровень границы невозможно. В настоящее время ведутся поиски нового маркера и нового стратотипа, и переизучаются опорные разрезы всех регионов. Кроме того, нужно отметить, что изменения в биоте и в палеообстановках в пограничном интервале лучше всего изучены в бассейнах Западной Европы и Марокко (Walliser, 1984; Becker et al., 2016). Наряду с этим, имеется много сведений и по разрезам в Казахстане и Средней Азии (Симаков, 1985; Барсков и др., 1988) и Китае (Ruan, 1981; Sheng in Ji, 1989; Qie et al., 2015; Zhang et al., 2019).
В связи с этим начато переизучение западноказахстанского разреза Берчогур, в ходе которого задокументировано точное положение находок аммоноидей и других ископаемых и проведены палеонтологические и седиментологические исследования. В течение двух полевых сезонов (2018 и 2019 гг.) мы проводили сборы ископаемых на разрезе и собрали новую коллекцию аммоноидей совместно с конодонтами, фораминиферами, кораллами, брахиоподами и криноидеями. Первые результаты переизучения аммоноидей изложены в этой статье.
МАТЕРИАЛ
Материалом для изучения послужили коллекции аммоноидей из Западного Казахстана, собранные в 1981–1983 гг. отрядом Палеонтологического ин-та АН CCCP под руководством Б.И. Богословского и геологического фак-та МГУ под руководством И.С. Барскова, а также сборы экспедиций Университета им. К.И. Сатпаева под руководством С.Н. Мустапаевой, в 2018–2019 гг. Все материалы собраны в левом борту сухого руч. Буртыбай (другое название Джангансай), в его меридиональном верхнем течении, в 1 км севернее нового карьера Алабас в Шалкарском р-не Актюбинской области. Материал происходит из нескольких близко расположенных местонахождений в джанганинской свите, которые традиционно именуются “Берчогур”, по соседнему пос. Берчогур (Биршогыр) и железнодорожной станции с тем же названием. Основные сборы происходят из пачки 3 разреза 3 (=BK-3) в левом борту сая Буртыбай (Джангансай) (координаты 48°34′08′′ N, 58°40′23′′ E), который в литературе и на полевых этикетках 1981–1983 гг. также называется “точка 3”, “канава БК-3”, “обр. 1”, “обр. 2”. Дополнительный материал собран из точки 5 в 10 м к северу от разреза 3 (поверхность кровли пачки 3), точки 6 в 300 м к северу и вверх по ручью от разреза по левому борту, в его широтном течении (координаты 48°34′15.77′′ N, 58°40′26.01′′ E), а также из осыпи пачки 4 разреза 3 (координаты 48°34′09.7′′ N, 58°40′22.3′′ E). Изученные коллекции хранятся в Палеонтологическом ин-те им. А.А. Борисяка (ПИН) РАН (Москва), №№ 1266, 4005, 5643.
АММОНОИДЕИ НА РУБЕЖЕ ДЕВОНА И КАРБОНА
В конце девонского периода богатые и разнообразные фауны аммоноидей существовали в бассейнах Урала, Тянь-Шаня, Китая, Западной Европы, Северной Африки и Северной Америки. Они были представлены отрядами Clymeniida, Agoniatitida и Goniatitida. Особенно выделялись клименииды, которые обладали дорсальным сифоном и имели самую разнообразную форму раковины. Агониатитиды, напротив, находились в стадии угасания. Гониатитиды были представлены в основном одним или двумя морфотипами раковины с очень инволютными оборотами и лопастной линией с неразделенной вентральной и простой боковой лопастями. Умбиликальная лопасть располагалась на умбиликальном шве. В самом конце девона все отряды аммоноидей испытали глубокий кризис, в результате которого исчезли клименииды и агониатитиды, и сильно сократилось разнообразие гониатитид. Аммоноидеи пограничных слоев девона и карбона лучше всего изучены в Германии, Австрии и Франции (Schindewolf, 1937; Vöhringer, 1960; Walliser, 1984; Korn, 1992; Becker, 1996; Bartzsch, Weyer, 1996; Korn, Weyer, 2003; Korn, Feist, 2007 и др.), где они встречаются в характерной последовательности циклических отложений, включающих хангенбергские сланцы, песчаники и известняки. Эти отложения отвечают последовательности трансгрессивных и регрессивных событий в истории Рейнского бассейна (Price, House, 1984; Becker, 1993, 1996; House, 1993; Korn, 1993, 2000; Becker et al., 2016 и др.), которые отражались на разнообразии населяющих его организмов. Основное событие в развитии биоты (крупное вымирание в конце девона, так называемое “хангенбергское событие”) связывают с затоплением шельфа и развитием малокислородных и бескислородных обстановок, которые сопровождались отложением черных сланцев Хангенберг в аммоноидной фазе UD VI-E (=середина фаменской конодонтовой фазы praesulcata). В черных сланцах встречаются гониатитиды семейства Prionoceratidae и последние клименииды. Это событие фиксируется и в других регионах мира, например, в Южном Китае и в Северной Америке (Becker et al., 2016). В целом сообщество аммоноидей и черных сланцев Хангенберг, и их аналогов крайне обедненное (роды Mimimitoceras, ?Sporadoceras, Postclymenia, Cymaclymenia) (Zhang et al., 2019). Этот уровень можно считать завершением девонского периода развития аммоноидей. За затоплением последовало резкое обмеление и новая трансгрессия (соответственно, песчаники Хангенберг и известняки Штоккум и Хангенберг), с новым комплексом аммоноидей, причем ни один из видов аммоноидей из черных сланцев не встречается в вышележащих отложениях (Walliser, 1984, 1996). В песчаниках Хангенберг найдены первые представители рода Acutimitoceras, и на этом уровне в Рейнском бассейне в настоящее время проводится нижняя граница одноименной генозоны. Массовое появление Acutimitoceras приходится на интервал известняков Штоккум (=нижней части известняков Хангенберг) и их аналогов (верхняя часть UD VI-F до LC-I A1) (Becker et al., 2016; Zhang et al., 2019). Сообщество генозоны, состоящее исключительно из прионоцератид (роды Imitoceras, Nicimitoceras и Acutimitoceras), отличается присутствием раковин с эволютными начальными оборотами, которые, вероятно, дали начало основным раннетурнейским морфотипам (напр., родам Gattendorfia, Zadelsdorfia, Voehringerites и новому отряду Prolecanitida) (Korn, Weyer, 2003 и др.). Утвержденная граница девона и карбона (D/C) (по появлению конодонтов вида Siphonodella sulcata) проходит внутри аммоноидной генозоны Acutimitoceras, выше уровня главного события вымирания (Becker et al., 2016) и, вероятно, внутри видовой зоны A. prorsum (Korn, Weyer, 2003). Новый уровень границы, который, с большой вероятностью, будет выбран по первому появлению конодонтов Protognathodus kockeli, тоже проходит внутри этой зоны, вблизи основания известняка Штоккум.
КОМПЛЕКС АММОНОИДЕЙ ГЕНОЗОНЫ ACUTIMITOCERAS ИЗ КАЗАХСТАНА
Древнейшие каменноугольные аммоноидеи известны в пограничных отложениях девона и карбона Карагандинской и Актюбинской областей Казахстана. Более древние позднефаменские сообщества аммоноидей включали представителей трех отрядов (Clymeniida, Goniatitida, Agoniatitida). Эти сообщества были очень разнообразными, и в них ведущую роль играли клименииды. В конце фамена (до конодонтовой фазы S. sulcata) все эти аммоноидеи вымерли, и их место заняли сообщества прионоцератид генозоны Acutimitoceras (роды Acutimitoceras, ?Nicimitoceras, Imitoceras). В Карагандинской обл. Либрович (1940) указывал разрезы с видами, ныне относимыми к роду Acutimitoceras: (1) р. Айна-Су (лев. приток р. Нуры), обн. 553 [?Nicimitoceras cf. ?carinatum (Schmidt, 1924), A. subbilobatum (Münster, 1839) = = вероятно, A. dzhanganense sp. nov.], обн. 1375 [A. subbilobatum (Münster, 1839) = вероятно, A. dzhanganense sp. nov.], (2) в басс. р. Шерубай-Нура (руч. Сулу), обн. 2126 [A. intermedium (Schindewolf, 1923)], (3) в окрестностях сопки Тюгельбай (обн. 1046) [A. “rotiforme” (Librovitch, 1940)], (4) Карамайсолган, обн. 1349 [A. intermedium (Schindewolf, 1923)], (5) сопка Майузек, южный клон г. Теректы (обн. 926) [A. subbilobatum (Münster, 1839) = вероятно A. dzhanganense sp. nov.], обн. 103 [A. intermedium (Schindewolf, 1923)].
В Шалкарском р-не Актюбинской обл. известны два района с аммоноидеями в пограничных отложениях девона и карбона – на востоке и на западе Берчогурской мульды. Балашова (1953) изучала аммоноидей из западной части мульды, а Кузина (1985) из северо-восточной, в районе истоков руч. Буртыбай (в его меридиональном течении, разрез 3 и осыпь). Балашова (1953) указывала следующие местонахождения аммоноидей в пределах Берчогурской мульды: (1) сухие реки Актан и Куcть-Кара – A. intermedium, A. cf. rotiforme, A. dzhanganense sp. nov. (определен Балашовой как Imitoceras subbilobatum), (2) среднее течение р. Сарыcай – A. cf. rotiforme, A. dzhanganense sp. nov. (определен Балашовой как I. subbilobatum), (3) на запад от. хребта Джан-Гана, балка Тунгулык-Булак – A. alabasense sp. nov. (определен Балашовой как I. substriatum), I. bertchogurense, A. dzhanganense sp. nov. (определен Балашовой как I. subbilobatum; (4) р. Кабакcай – A. dzhanganense sp. nov. (определен Балашовой как I. subbilobatum. Большинство этих сухих рек показано на рис. 1.
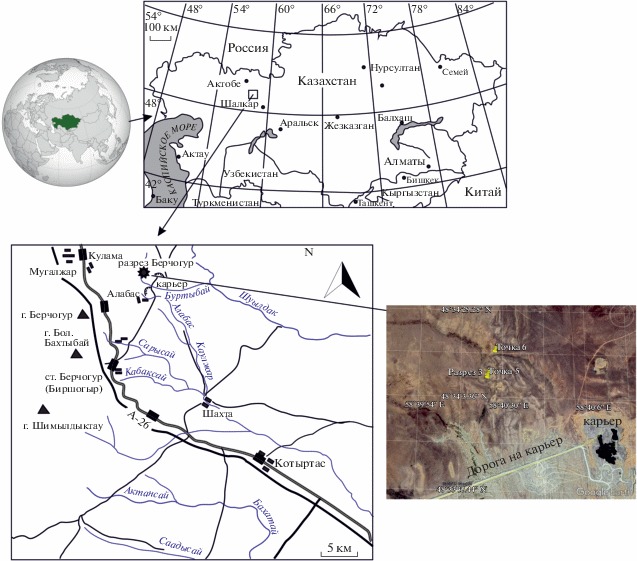
Изученное нами местонахождение “Берчогур” в восточной части мульды находится в борту сухого ручья Буртыбай (другое название Джангансай), к востоку от пос. Берчогур и Алабас (Балашова, 1953; Barskov et al., 1984; Кузина, 1985; Фауна и биостратиграфия …, 1987; Барсков и др., 1988). Разрез находится примерно в 1 км севернее нового отделения карьера “Алабас”, в левом борту сухого ручья, который пересекается дорогой, соединяющей старый и новый карьеры (рис. 1). Аммоноидеи происходят из пачек 3 и 4 разреза [см. описание разреза в: Барсков и др. (1988)].
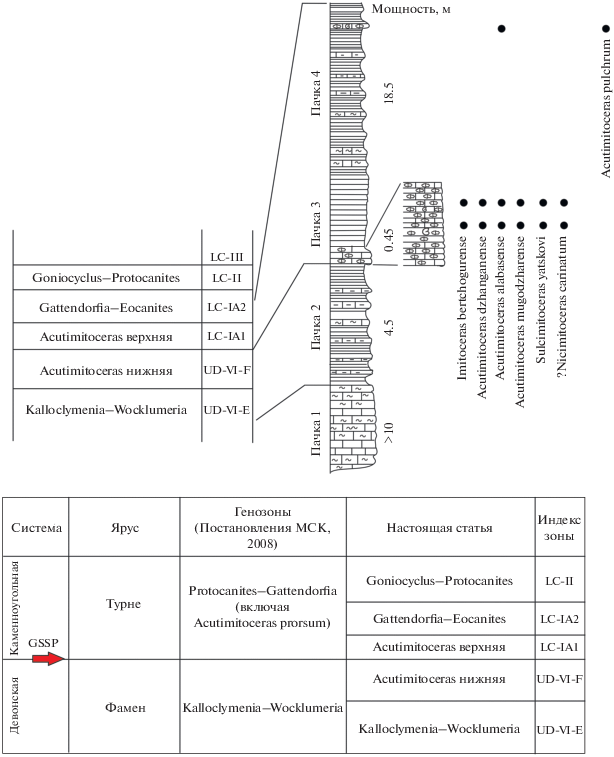
Комплекс из пачки 3 в целом соответствует фауне, описанной Кузиной (1985), и содержит Imitoceras bertchogurense (Balashova, 1953); Acutimitoceras dzhanganense sp. nov. [вид, определенный Кузиной как “I. (A). subbilobatum” (Münster, 1839)], A. alabasense sp. nov. [относимый Кузиной к “I. (A.) substriatum” (Münster, 1840)], A. mugodzharense Kusina, 1984, ?Nicimitoceras carinatum (Schmidt, 1924), Sulcimitoceras yatskovi Kusina, 1985. Этот комплекс относится к генозоне Acutimitoceras на основании находок рода–индекса. Видовую зону определить сложнее. Комплекс содержит прионоцератид с инволютными начальными оборотами, I. bertchogurense, и ?N. carinatum. Кроме того, вид A. dzhanganense похож на А. subbilobatum, который в типовой местности и в Марокко найден в отложениях зоны prorsum (Becker et al., 2016). Поэтому, вероятнее всего, комплекс из пачки 3 можно коррелировать с зоной prorsum.
Положение этой фауны относительно конодонтовой зональности в настоящее время переизучается. На сегодняшний день известно, что уровень находок конодонтов S. praesulcata на Берчогуре находится в пачке 2б, примерно в метре ниже пачки 3 с аммоноидеями, а сам интервал с фауной аммоноидей охарактеризован конодонтами S. sulcata (Barskov et al., 1984; Фауна и биостратиграфия …, 1987; Барсков и др., 1988), которые появляются в пачке 3, непосредственно ниже находок аммоноидей. Таким образом, утвержденная граница D/C, вероятно, находится в основании слоев с аммоноидеями (пачка 3). Если это действительно так, то пачку 3 можно сопоставить с верхней частью зоны prorsum, поскольку в Западной Европе вид конодонтов S. sulcata впервые появляется внутри зоны prorsum (Clausen, Korn, 2008). Известняки пачки 3 перекрываются глинистыми сланцами пачки 4 с Acutimitoceras pulchrum Kusina, 1985 [прежде определенные как A. aff. “rotiforme” (Barskov et al., 1984)] и A. alabasense (см. Кузина, 1985; Барсков и др., 1988). Возможно, слои пачки 4 можно сопоставить с более высокими горизонтами турне, но пока для этого недостаточно данных.
Сообщество аммоноидей имитоцерасовой фауны Берчогура схоже с одновозрастными сообществами других регионов тем, что включает три основных морфотипа взрослой раковины (широкую, относительно узкую, и узкую килеватую формы). Первый морфотип представлен A. alabasense, второй – I. bertchogurense и A. dzhanganense, третий – ?N. carinatum. При этом, за исключением I. bertchogurense Balashova, 1953 и A. pulchrum Kusina, 1985, раковины лишены пережимов (у последнего вида пережимы развиты только на ядре). Ювенильные раковины отличаются несколько большим разнообразием. Среди них найдены шарообразные формы с относительно широким умбиликом (A. alabasense), пахиконовые формы с широким и относительно широким умбиликом (А. mugodzharense, A. dzhanganense, S. yatskovi), пахиконовые формы с узким умбиликом (?N. carinatum), узко дискоидальные раковины с приоткрытым умбиликом (A. pulchrum). Эти основные морфотипы подразделяются на более мелкие морфотипы ювенильных форм, например, с вентральным желобком (S. yatskovi), с грубой скульптурой (А. mugodzharense) и так далее. Кроме того, Кузина (1985) указала на находку оксиконовой раковины 15 мм в диаметре (которую, к сожалению, не удалось повторить). М. Хаус (House, 1996, с. 182) оценил разнообразие ювенильных форм из известняка Штоккум (Германия), как результат значительного давления отбора на самых ранних стадиях развития аммоноидей вслед за крупными биотическими кризисами, связанными с бескислородными обстановками в девонских бассейнах. Интересно, что схожее высокое разнообразие морфотипов ранних стадий аммоноидей генозоны Acutimitoceras наблюдается в таких удаленных регионах, как Штоккум (Рейнские Сланцевые Горы), Грюн Шнейд (Grüne Schneid, Карнийские Альпы, Австрия) и Западный Казахстан. В частности, можно отметить присутствие схожих ювенильных форм, напр., с вентральным желобком (Кузина, 1985, табл. III , фиг. 10 ; House, 1996, рис. 23 А), субсфероконовых с полностью закрытым умбиликом (Кузина, 1985, рис. 1д; Schönlaub et al., 1988, табл. 2 , фиг. 4), субсфероконовых и пахиконовых с частично открытым умбиликом (здесь, экз. 4005/115; Korn, 1984, табл. 2 , фиг. 9, 15 ). Разнообразие ранних стадий скорее было связано с приспособлением к различным условиям питания и защитными функциями, а не с локомоцией, поскольку молодь аммоноидей была, вероятно, планктонной. Преобладающей эволюционной тенденцией в это время был педоморфизм, возможно, связанный с преимуществами достижения половозрелости на ранних стадиях. Педоморфизм отмечен в эволюции прионоцератид и их потомков – гаттендорфид, у которых развернутыми являются и юные, и более поздние обороты. По-видимому, выживаемость прионоцератид была связана с биологией видов, способных приспосабливаться к различным условиям обитания, а разнообразие ювенильных стадий может свидетельствовать о приспособлении к расселению молоди в различные биотопы.
БИОСТРАТИГРАФИЯ ПОГРАНИЧНОГО ИНТЕРВАЛА ПО АММОНОИДЕЯМ И МОРФОЛОГИЯ РАКОВИНЫ
Шкала аммоноидных генозон пограничного интервала девона и карбона претерпевает постоянные изменения. В основном в пограничном интервале указывают две генозоны – Acutimitoceras (частью в фамене, частью в турне) и Gattendorfia (низы турне) (напр., Becker, 1996), однако есть и другие варианты шкалы. Например, Р. Беккер и Д. Вайер (Becker, Weyer, 2004) указывают три генозоны: Stockumites (фамен), Acutimitoceras и Gattendorfia (две последние генозоны в схеме этих авторов отвечают низам турне). В большинстве схем генозона Acutimitoceras подразделяется на две видовые зоны – Acutimitoceras prorsum и Acutimitoceras acutum (Price, House, 1984; Shönlaub et al., 1988; Clausen et al., 1989; Becker, 1993; Korn, Weyer, 2003 и др.), но в некоторых шкалах зона Acutimitoceras acutum не значится, а выше зоны Acutimitoceras prorsum указывается зона Gattendorfia subinvoluta (Korn, Klug, 2015 и др.). Сообщество аммоноидей Берчогура (роды Imitoceras и Acutimitoceras) по своему таксономическому составу близко к сообществам известняка Штоккум, и на этом основании здесь тоже выделена генозона Acutimitoceras (Кузина, 1985) (рис. 2). Более детальное подразделение основано на анализе морфологии прионоцератид, которые составляют это сообщество. Установлено, что большинство прионоцератид этого интервала обладало эволютными начальными оборотами, что совершенно не характерно для позднефаменских представителей. Учитывая, что в вышележащей генозоне Gattendorfia–Eocanites большинство аммоноидей обладают эволютной раковиной во взрослом состоянии, можно предположить эволюцию по пути педоморфизма, с сохранением эволютных оборотов у взрослых раковин. Кроме того, Беккер (Becker, 1996) предложил учитывать размер раковин с килеватами оборотами, предположив, что развитие оксиконовых оборотов сдвигается на более ранние стадии в ходе филогенеза рода Acutimitoceras. Так, у вида Acutimitoceras sp., указанного Кузиной из Берчогура [Imitoceras (A.) sp., см. Кузина, 1985, табл. III , фиг. 6 ], оксиконовая раковина появляется при диаметре 15 мм, у A. acutum из турнейской зоны acutum типовой области в Германии – при диаметре 10 мм. У последнего вида оксиконовая раковина возникает на четвертом обороте, что отличает его от несколько более древнего ?N. carinatum (Schmidt, 1924), у которого оксиконовая раковина возникает на шестом обороте (Кузина, 1985, рис. 1и; Korn, 1994, рис. 37G ). Можно предположить, что сдвиг появления оксиконовой раковины на более ранние стадии в конечном счете привел к появлению Voehringerites и некоторых Gattendorfia, у которых такая раковина наблюдается во взрослом состоянии. Учитывая, что в берчогурском разрезе присутствуют аммоноидеи, у которых оксиконовая раковина появляется на шестом обороте, и которые были определены Кузиной (1985) как Imitoceras (Acutimitoceras) carinatum, можно предположить, что здесь представлены аналоги зоны prorsum. Кроме того, Кузина (1985) отметила, что I. bertchogurense напоминает вид A. prorsum и тоже предположила, что Берчогурский разрез, скорее всего, относится к зоне prorsum. Нужно отметить, что в недавних работах по фауне пограничных отложений девона и карбона вид A. carinatum отнесен к роду Nicimitoceras Korn, 1993 (Korn, Klug, 2002), возможно, потому что у рейнских представителей этого вида боковая лопасть заметно глубже вентральной, как, например, у N. subacre (Vöhringer, 1960) – типового вида Nicimitoceras. У берчогурских представителей ?N. carinatum боковая лопасть, изображенная Кузиной (1985, рис. 3б), не глубже, чем вентральная лопасть, но следует отметить, что лопастная линия топотипа вида ?N. carinatum зарисована при высоте оборота 16.5 мм (Korn, 1994), а у Кузиной – при диаметре раковины 7.6 мм, то есть, с гораздо меньшего экземпляра. Возможно, вид, описанный Кузиной как I. (A.) carinatum, представляет собой новый вид Nicimitoceras или Acutimitoceras, но пока материала для решения этого вопроса недостаточно. Аммоноидная фауна Берчогура, вероятно, может быть также сопоставлена с таковой из нижней части формации Вангиоу (Wangyou) в Китае, которая изучена в разрезах Мухуа (Muhua) (Ruan, 1981; Qie et al., 2015), где в тонкослоистых глинистых известняках отмечены A. wangyuense Sun et Shen, 1965. Аммоноидеи Acutimitoceras sp. обнаружены в этой же формации, в конодонтовой зоне S. sulcata или в основании зоны S. duplicata (Пачка C) в разрезе Гедонггуан (Gedongguan, Youjiang Basin, Changshun County) в провинции Гуйчжоу (Guizhou), Китай (Qie et al., 2015, рис. 4). Аммоноидеи с похожей морфологией раковины указываются из Фауны 1 (слой RTB10) разреза Гара Боу Тлидат (Gara Bou Tlidat) низов формации Феззу (Fezzou) в Марокко (Becker et al., 2018).
Рис. 2.
Шкала пограничных отложений девона и карбона и схематический разрез джанганинской свиты в разрезе Берчогур.
ТАКСОНОМИЯ РОДА ACUTIMITOCERAS LIBROVITCH, 1957
Либрович (1957) выделил род Acutimitoceras в сноске на с. 263 с указанием типового вида Acutimitoceras acutum (Schindewolf, 1923), но без описания и ревизии видового состава. Oн включил этот род в сем. Aganidae Smith, 1903, наряду с Imitoceras Schindewolf, 1923, Gattendorfia Schindewolf, 1920, Kazakhstania Librovitch, 1940, Irinoceras Ruzhencev, 1947, Neoaganides Plummer et Scott, 1937. Нужно отметить, что прежде Либрович (1940), который очень подробно рассматривал Imitoceras, отнес его к семейству Cheiloceratidae Frech, 1923. Обе точки зрения сейчас считаются устаревшими, и в настоящее время роды Imitoceras и Acutimitoceras относятся к семейству Prionoceratidae (Кузина, 1985; Korn, Klug, 2002 и др.). Д. Корн (Korn, 1994) установил подсемейство Acutimitoceratinae с типовым видом Acutimitoceras Librovitch, 1957, включив в него, помимо типового рода, Costimitoceras Vöhringer, 1960 (типовой вид C. ornatum Vöhringer, 1960) и Nicimitoceras Korn, 1993. Кузина (1985, с. 37) указала, что основным признаком, отличающим Acutimitoceras, Либрович считал килеватую вентральную сторону раковины. Она также отметила, что не все согласились считать этот признак определяющим, поскольку килеватая вентральная сторона появляется на взрослых стадиях многих каменноугольных таксонов. Справедливости ради нужно сказать, что Либрович (1957) ничего не написал о диагнозе нового рода, но сам факт того, что типовым родом он выбрал A. acutum, у которого килеватая вентральная сторона появляется очень рано в онтогенезе, позволяет предположить, что именно эта черта морфологии послужила основанием для выделения этого рода. Э. Фёрингер (Vöhringer, 1960) изучил морфогенез раковины A. acutum, у которого внутренние обороты оказались эволютными, и на основании этого Корн (Korn, 1984 и др.) отнес к роду Acutimitoceras всех прионоцератид с эволютными ранними стадиями. Кузина (1985) не была уверена в правильности такой трактовки, поскольку далеко не у всех прионоцератид с эволютными начальными оборотами развивается килеватая вентральная сторона, и предложила рассматривать Acutimitoceras в качестве подрода рода Imitoceras. Беккер (Becker, 1996) принял род Acutimitoceras, но подразделил его на несколько подродов. Помимо номинального подрода Acutimitoceras с типовым видом A. acutum, в составе рода им были выделены подроды Stockumites [типовой вид Imitoceras intermedium (Schindewolf, 1923)] и Streeliceras (типовой вид Imitoceras heterolobatum Vöhringer, 1960). Род Sulcimitoceras Kusina, 1985 Беккер понизил до подрода в роде Acutimitoceras на основании того, что борозда на вентральной стороне ядра [характерный признак Sulcimitoceras, указанный Кузиной (1985)], по его мнению, не может считаться родовым признаком, поскольку такую борозду можно обнаружить на многих экземплярах других таксонов, не связанных родством. Кроме того, ребра на боковых сторонах (тоже характерный признак) присутствуют, по мнению Беккера, и у других прионоцератид. Это справедливые замечания, но, кроме этих признаков, у Sulcimitoceras совершенно необычная лопастная линия с глубокой, но округленной боковой лопастью. Поэтому, как указывала Кузина, отнести этот вид к какому-то другому роду прионоцератид затруднительно. Корн и Клюг (Korn, Klug, 2002) посчитали подрод Stockumites Becker младшим синонимом Acutimitoceras Librovitch, а род Streeliceras Becker младшим синонимом рода Nicimitoceras Korn, 1993, но сохранили родовой статус Sulcimitoceras. Мы принимаем род Acutimitoceras с измененным диагнозом (прионоцератиды с эволютными начальными оборотами), но отмечаем, что систематика рода Acutimitoceras и всего семейства Prionoceratidae нуждается в основательной ревизии. Нашего материала для такой ревизии недостаточно, поскольку до сих пор отсутствуют сведения о строении начальных оборотов у ряда видов, особенно из китайских разрезов.
Автор выражает искреннюю благодарность С.Н. Мустапаевой (Ун-т им. К. Сатпаева, Алматы), А.С. Алексееву (МГУ), Е.И. Кулагиной (Ин‑т Геологии, Уфимский научный центр РАН, Уфа) и Ю.А. Гатовскому (МГУ) за участие в полевых исследованиях и полезные дискуссии, и Т.Б. Леоновой (ПИН РАН) за обсуждение рукописи и критическую правку. Полевые исследования были поддержаны грантом Министерства науки и образования республики Казахстан, № 2018/AP05131610. Исследования С.В. Николаевой были поддержаны Программой Президиума РАН и Программой Казанского ун-та по повышению конкурентоспособности ведущих российских университетов.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Род Acutimitoceras Librovitch, 1957
Acutimitoceras alabasense Nikolaeva, sp. nov.
Табл. III , фиг. 1–6 (см. вклейку)
Рис. 5.
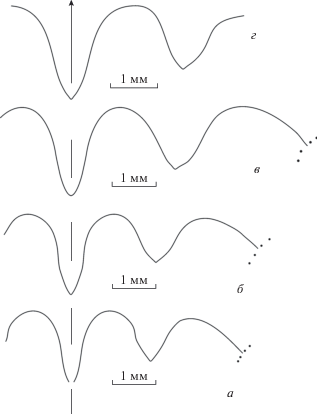
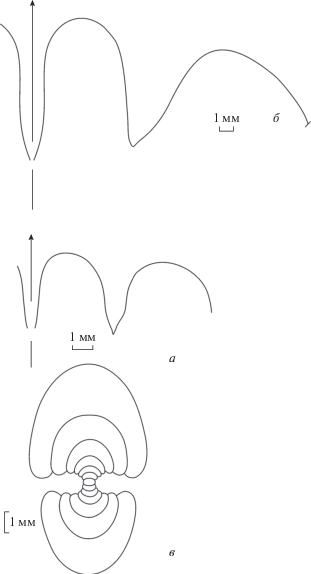
Лопастные линии и сечение Acutimitoceras dzhanganense sp. nov.: а – экз. ПИН, № 5643/1, при В = 10.5 мм, Ш = 13.4 мм; б – голотип ПИН, № 4005/126, при В = 12.2 мм, Ш = 13.4 мм; в – экз. ПИН, № 4005/62, при Д = 10.4 мм; разрез Берчогур.
Imitoceras substriatum (part.): Балашова, 1953, с. 196, табл. 12 , фиг. 1–9 .
Imitoceras (Acutimitoceras) substriatum (part.): Кузина, 1985, с. 42, табл. III , фиг. 3, 4.
?Acutimitoceras substriatum: Ruan, 1981, c. 63, табл. 11 , фиг. 26–28 .
non Goniatites striatus Münster, 1839, c. 20.
non Goniatites substriatus Münster, 1840, c. 107.
Название вида по пос. Алабас, близ которого расположен разрез.
Голотип – ПИН, № 4005/147; Казахстан, Актюбинская обл., Шалкарский р-н, к северо-востоку от пос. Берчогур (Биршогыр), левый борт сухого руч. Буртыбай, в его меридиональном верхнем течении, 1 км севернее нового карьера Алабас, разрез Берчогур; джанганинская свита; нижний карбон, генозона Acutimitoceras.
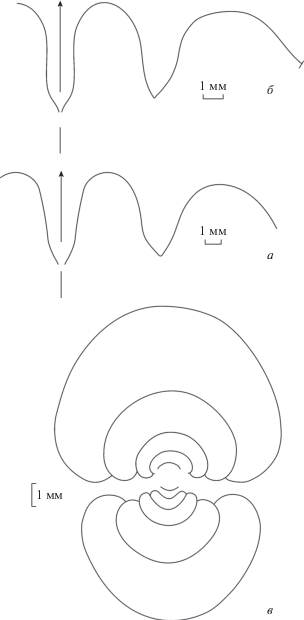
Описание. Форма (рис. 3, в). Начальные обороты слабоэволютные (рис. 3, в; Кузина, 1985, рис. 1, з). Во взрослом состоянии раковина пахиконовая, с округленной вентральной стороной, с широкими, слабовыпуклыми боковыми сторонами, постепенно сходящимися к вентральной. Умбилик закрытый.
Размеры в мм и отношения:
| Экз. № | Д | В | Ш | Ду | В/Д | Ш/Д | Ду/Д |
|---|---|---|---|---|---|---|---|
| 4005/122 | 39.8 | 20.9 | 26.8 | 1.0 | 0.53 | 0.67 | 0.03 |
| 4005/147 голотип |
33.7 | 19.8 | 22.6 | – | 0.59 | 0.67 | – |
| 4005/129 | 25.8 | 13.6 | 16.6 | – | 0.53 | 0.64 | – |
| 4005/124 | 21.3 | 10.7 | 16.1 | – | 0.50 | 0.76 | – |
| 4005/150 | 20.5 | 12.6 | 13.5 | – | 0.61 | 0.66 | – |
| 4005/123 | 18.6 | 9.4 | 14 | – | 0.51 | 0.75 | – |
| 4005/117 | 12.7 | 7.0 | 8.4 | – | 0.55 | 0.66 | – |
| 4005/115 | 9.3 | 4.2 | 7.4 | 1.0 | 0.45 | 0.80 | 0.11 |
Скульптура. На взрослых раковинах скульптура не сохранилась. Пережимов нет. На более юных раковинах заметны ламеллы (на экз. № 4005/124, при Д = 14.5 мм, на 1 мм боковой стороны приходится 4–5 ламелл), которые более или менее радиально отходят от умбилика, затем изгибаются назад, образуя широкий вентральный синус.
Лопастная линия (рис. 3, a, б; 4). Вентральная лопасть узкая, ланцетовидная, со слабовыпуклыми сторонами, наружное седло широкоокругленное. Боковая лопасть довольно узкая, со слабовыпуклыми сторонами, заметно короче вентральной лопасти. Основание боковой лопасти узко-угловатое. Наружная сторона боковой лопасти более или менее плоская, а внутренняя сторона слабо выпуклая. Второе наружное седло такой же высоты, как первое, широкое.
Сравнение. По параметрам раковины новый вид близок к ?А. substriatum (Münster, 1840) из Верхней Франконии (Бавария, Германия), от которого отличается отсутствием пережимов на всех стадиях роста. От экземпляров A. intermedium (Schindewolf, 1923), приведенных Корном (Korn, 1994, с. 47) из местонахождения Штоккум, кроме того, отличается более широкими оборотами взрослой раковины (при Д = 30 мм Ш/Д = 0.67 против 0.60). От паратипа CE 1130/29 вида A. simile (Vöhringer, 1960) из сл. 1 известняка Гаттендорф местонахождения Хоннетал (Hönnetal), Германия, и экземпляров A. cf. simile, изображенных Корном и Ваейром (Korn, Weyer, 2003) из сл. 80 разреза Хассельбах (Hasselbach), Германия, отличается формой поперечного сечения оборотов. У наших экземпляров на пятом обороте умбилик еще относительно открытый, и поперечное сечение более широкое (Ш/Д = 0.77), в то время как у сравниваемых экземпляров поперечное сечение более вытянуто в высоту (Vöhringer, 1960, рис. 7a ; Korn, Weyer, 2003, рис. 14b ).
Замечания. Балашова (1953) и Кузина (1985) описали экземпляры этого вида, отнеся их к “I. (A.) substriatum (Münster, 1840)”. При этом они указали, что на изученных экземплярах нет пережимов (ни на поверхности раковины, ни на ядре). Кузина отметила, что проводила сравнение с экземплярами ?A. substriatum, описанными Фёрингером (Vöhringer, 1960) из Хоннетала, Германия, а не с типовым материалом, который был недоступен для изучения. Корн (Korn, 1994, с. 22, рис. 12D ) опубликовал фотографию голотипа этого вида (Мюнхенский музей BSP AS Vll25) и дал его краткое описание. Голотип A. substriatum представляет собой раковину примерно 25 мм в диаметре, Ш/Д = 0.65, с пережимами на раковине и ядре. Рядом с пережимами на голотипе наблюдается небольшое вздутие. Последняя характеристика является диагностическим признаком рода Mimimitoceras, видимо, поэтому Корн под вопросом отнес вид substriatum к этому роду. Также под вопросом этот вид отнесен к Mimimitoceras Корном и Клюгом (Korn, Klug, 2002). На наших образцах начальные обороты раковины слабо эволютные. Учитывая это и то, что раковины и ядра лишены пережимов, мы относим новый вид к роду Acutimitoceras.
Материал. 10 экз. из разреза Берчогур.
Acutimitoceras dzhanganense Nikolaeva, sp. nov.
Табл. III , фиг. 7–10
Imitoceras subbilobatum (part.): Либрович, 1940, с. 13, рис. 2–4, табл. I , фиг. 1–4; Балашова, 1953, с. 191, табл. 11 , фиг. 11–16 .
Imitoceras (Acutimitoceras) subbilobatum (part.): Кузина, 1985, с. 43, табл. III , фиг. 5.
?Acutimitoceras subbilobatum: Sheng in Ji et al., 1989, c. 111, табл. 33 , фиг. 1, 2.
non Goniatites subbilobatus Münster, 1839.
Название вида по джанганинской свите.
Голотип – ПИН, № 4005/126; Казахстан, Актюбинская обл., Шалкарский р-н, к северо-востоку от пос. Берчогур (Биршогыр), левый борт сухого руч. Буртыбай, в его меридиональном верхнем течении, 1 км севернее нового карьера Алабас, разрез Берчогур; джанганинская свита; нижний карбон, генозона Acutimitoceras.
Описание. Форма (рис. 5, в). Начальные обороты эволютные (см. Кузина, 1985, рис. 1). У экз. № 4005/77, при Д = 4.8 мм, ширина умбилика около 1 мм. Во взрослом состоянии раковина пахиконовая, с относительно медленно возрастающими оборотами (W = 2.12). Вентральная сторона узкоокругленная, боковые стороны слабовыпуклые, сходящиеся к вентральной стороне. Умбилик у взрослых экземпляров закрытый.
Размеры в мм и отношения:
| Экз. № | Д | В | Ш | Ду | В/Д | Ш/Д |
|---|---|---|---|---|---|---|
| 4005/126 голотип |
45.8 | 26.1 | 23.6 | – | 0.57 | 0.52 |
| 4005/205 | 46.7 | 26.5 | 21.7 | – | 0.57 | 0.46 |
| 4005/195 | 38.0 | 20.1 | 19.2 | – | 0.53 | 0.51 |
| 5643/1 | 34.2 | 18.6 | 19.2 | – | 0.54 | 0.56 |
| 4005/133 | 23.7 | 13.9 | 13.2 | – | 0.59 | 0.56 |
| 4005/58 | 21.5 | 12.4 | 11.7 | – | 0.58 | 0.54 |
| 4005/154 | 24.1 | 13.7 | 13.0 | – | 0.57 | 0.54 |
Скульптура. На взрослых раковинах скульптура не сохранилась. Пережимов нет. На юных раковинах развиты струйки роста, прямые на боковых сторонах и образующие мелкий вентральный синус.
Лопастная линия (рис. 5, а, б). Вентральная лопасть узкая, с параллельными сторонами, наружное седло округленное, широкое. Боковая лопасть довольно узкая, с вогнутой наружной стороной и слабовыпуклой внутренней стороной, немного короче вентральной лопасти у взрослых экземпляров. Второе наружное седло такой же высоты, как первое, широкое.
Сравнение. По форме поперечного сечения новый вид близок к A. subbilobatum (Münster, 1839), от которого, в первую очередь, отличается отсутствием пережимов. Это хорошо заметно, особенно при сравнении с экземплярами, изображенными Корном из Штоккума (Korn, 1994, рис. 42a–c ) и Обер-Родингхаузена (Ober-Rödinghausen) (Korn, 1994, рис. 44d–f ). Кроме того, видно, что у казахстанских представителей боковая лопасть уже (Korn, 1994, рис. 39 ). От экземпляров A. intermedium (Schindewolf, 1923), приведенных Корном (Korn, 1994, с. 47) из местонахождения Штоккум, кроме того, отличается более узкими оборотами взрослой раковины (при Д = 30 мм Ш/Д = 0.56 против 0.60). От типовых экземпляров A. procedens Korn (Korn, 1984, с. 80, табл. 4 , фиг. 24, 25 , рис. 5B) из местонахождения Мюссенберг (Müssenberg), Германия, отличается отсутствием пережимов (у сравниваемого вида их, по крайней мере, четыре на оборот) и отсутствием бокового синуса у линий роста молодых экземпляров. От A. prorsum (Schmidt, 1925) отличается более широкой раковиной (при Д = 20 мм Ш/Д = 0.54 против 0.50), отсутствием пережимов и слабее изогнутыми линиями роста.
Замечания. Либрович (1940), Балашова (1953) и Кузина (1985) относили, по крайней мере, некоторые берчогурские экземпляры A. dzhanganense sp. nov. к виду A. subbilobatum (Münster, 1839). Тем не менее, все эти авторы отмечали, что ни у одного представителя из Берчогура они не наблюдали пережимы. Их определение было связано с недостаточной в то время изученностью типового материала вида A. subbilobatum. Помимо того, что лопастная линия голотипа этого вида была зарисована неправильно при первоначальном описании (см. Münster, 1839, табл. XVII, фиг. 1 и Либрович, 1940), было неясно, присутствуют ли пережимы на раковине и/или ядре, и каковы начальные обороты раковины. Нужно отметить также, что существовала путаница с номенклатурой этого вида, который описывали еще и как “Aganides guerichi Frech” (см. историю вопроса у Либровича, 1940). Кузина (1985), например, указывала, что, возможно, берчогурские экземпляры относятся не к subbilobatum, а к новому виду, но что в отсутствие полноценного описания лектотипа Г. Мюнстера она не решается выделить новый вид. К настоящему времени описаны хорошо сохранившиеся экземпляры A. subbilobatum (Münster, 1839) из типового местонахождения Гаттендорф (Германия) (Korn, 1984), из местонахождений Штоккум и Обер-Родингхаузен (Korn, 1994), Хассельбах (Германия) (Korn, Weyer, 2003), Мфис (Анти-Атлас, Марокко) (Bockwinkel, Ebbighausen, 2006) и других районов Западной Европы и Северной Африки, и все они имеют отчетливые пережимы на ядре. Учитывая, что ни один из экземпляров берчогурских раковин, “похожих на Acutimitoceras subbilobatum”, не имеет пережимов, можно сделать вывод, что это новый вид рода Acutimitoceras. Возможно, экземпляры из разреза Дапушанг (Dapoushang), изображенные Шенгом (Sheng in Ji et al., 1989, табл. 33 , фиг. 1, 2), также относятся к новому виду.
Материал. 11 экз. из разреза Берчогур.
Список литературы
Балашова Е.А. Гониатиты карбона Бер-Чогура // Ежегодн. Всес. Палеонтол. об-ва. Т. 14. М.–Л., 1953. С. 189–202.
Барсков И. С., Кононова Л. И., Бывшева Т. В., Алексеев А.С. Граница девона и карбона в разрезе Берчогур (Мугоджары) // Граница девона и карбона на территории СССР. Минск, 1988. С. 171–180.
Кузина Л.Ф. К ревизии рода Imitoceras // Палеонтол. журн. 1985. № 3. С. 35–48.
Либрович Л.С. Ammonoidea из каменноугольных отложений Северного Казахстана // Палеонтология СССР. Т. 4. Ч. 9. Вып. 1. М.- Л., 1940. 394 с.
Либрович Л.С. О некоторых новых группах гониатитов из каменноугольных отложений СССР // Ежегодн. Всес. Палеонтол. об-ва. 1957. Т. 16. С. 246–272.
Постановления МСК и его постоянных комиссий. СПб.: ВСЕГЕИ, 2008.
Симаков К.В. Биостратиграфия пограничных отложений девона и карбона. Вып. 9. Пограничные отложения девона и карбона Мугоджар. Препринт. Магадан: СВКНИИ ДВНЦ, 1985. 55 с.
Фауна и биостратиграфия пограничных отложений девона и карбона Берчогура (Мугоджары) / Ред. Маслов В.А. М.: Наука, 1987. 119 с.
Barskov I.S., Simakov K.V., Alekseev A.S. et al. Devonian–Carboniferous transitional deposits of the Berchogur section, Mugodzhary, USSR // Cour. Forsch.-Inst. Senckenb. 1984. V. 67. P. 207–230.
Bartzsch K., Weyer D. Acutimitoceras acutum (Schindewolf 1923) – Leitart der ersten unterkarbonischen Ammonoidea-Zone im Bohlen-Profil von Saalfeld (Thüringisches Schiefergebirge) // Beitr. Geol. Thüringen. N. F. 1996. № 3. S. 91–103.
Becker R.T. Anoxia, eustatic changes, and Upper Devonian to lowermost Carboniferous global ammonoid diversity // The Ammonoidea, Environment, Ecology, and Evolutionary Change / Ed. House M.R. N.Y.: Oxford Univ. Press, 1993. P. 115–164 (Syst. Assoc. Spec. Vol. № 47).
Becker R.T. New faunal records and holostratigraphic correlation of the Hasselbachtal D/C-boundary auxiliary stratotype (Germany) // Ann. Soc. géol. Belg. 1996. V. 117. P. 19–45.
Becker R.T., Hartenfels S., Klug Ch. et al. The cephalopod-rich Famennian and Tournaisian of the Aguelmous Syncline (southern Maïder) // Münster. Forsch. Geol. Paläontol. 2018. V. 110. P. 273–306.
Becker R.T., Kaiser S.I., Aretz M. Review of chrono-, litho- and biostratigraphy across the global Hangenberg Crisis and Devonian–Carboniferous boundary // Geol. Soc. London. Spec. Publ. 2016. V. 423. P. 355–386.
Becker R.T., Weyer D. Bartzschiceras n. gen. (Ammonoidea) from the Lower Tournaisian of Southern France // Mitt. Geol.-Paläontol. Inst. Univ. Hamburg. 2004. V. 88. P. 11–36.
Bockwinkel J., Ebbighausen V. A new ammonoid fauna from the Gattendorfia-Eocanites Genozone of the Anti-Atlas (Early Carboniferous; Morocco) // Fossil Rec. 2006. V. 9. № 1. P. 87–129.
Clausen C.-D., Korn D. 4.1.15 Höheres Mitteldevon und Oberdevon des nördlichen Rheinischen Schiefergebirges (mit Velberter Sattel und Kellerwald) // Schr. Dtsch. Ges. Geowiss. 2008. H. 52. S. 439–481 (Stratigraphie von Deutschland VIII).
Clausen C.-D., Leuteritz K., Ziegler W., Korn D. Ausgewählte Profile an der Devon/Karbon-Grenze im Sauerland (Rheinisches Schiefergebirge) // Fortschr. Geol. Rheinland und Westfalen. 1989. V. 35. P. 161–226.
House M. Earliest Carboniferous goniatite recovery after the Hangenberg Event // Ann. Soc. géol. Belg. 1993. V. 115. P. 559–579.
House M.R. Juvenile goniatite survival strategies following Devonian extinction events // Geol. Soc. Spec. Publ. 1996. V. 102. P. 163–185.
Kaiser S.I. The Devonian/Carboniferous boundary stratotype section (La Serre, France) revisited // Newslett. Stratigr. 2009. V. 43/2. C. 195–205.
Korn D. Die Goniatiten der Stockumer Imitoceras–Kalklinsen (Ammonoidea; Devon/Karbon-Grenze) // Cour. Forsch.-Inst. Senckenb. 1984. Bd 67. S. 71–89.
Korn D. Ammonoideen aus dem Devon/Karbon-Grenzprofil an der Grünen Schneid (Karnische Alpen, Österreich) // Jb. Geol. Bund. 1992. Bd 135. № 1. S. 7–19.
Korn D. The ammonoid faunal change near the Devonian–Carboniferous boundary // Ann. Soc. géol. Belg. 1993. V. 115. P. 581–593.
Korn D. Devonische und karbonische Prionoceraten (Cephalopoda, Ammonoidea) aus dem Rheinischen Schiefergebirge // Geol. und Paläontol. in Westfalen. 1994. Bd 30. S. 1–85.
Korn D. 2000. Morphospace occupation of ammonoids over the Devonian-Carboniferous boundary // Paläontol. Z. 2000. V. 74. № 3. P. 247–257.
Korn D., Feist R. Early Carboniferous ammonoid faunas and stratigraphy of the Montagne Noire (France) // Fossil Rec. 2007. V. 10. P. 99–124.
Korn D., Klug Ch. Ammoneae Devonicae // Fossilium Catalogus, I: Animalia. Pars 138 / Ed. Riegraf W. Leiden: Backhuys Publ., 2002. 375 p.
Korn D., Klug Ch. Paleozoic Ammonoid Biostratigraphy // Ammonoid Paleobiology: From Macroevolution to Paleogeography / Eds. Klug Ch., Korn D., De Baets K. et al. Dordrecht: Springer, 2015. P. 299–328.
Korn D., Weyer D. High resolution stratigraphy of the Devonian–Carboniferous transitional beds in the Rhenish Mountains // Mitt. Mus. Naturk. Berlin. Geowiss. Reihe. 2003. V. 6. P. 79–124.
Münster G.G. Nachtrag zu den Goniatiten des Fichtelgebirges // Beitr. zur Petrefactenkunde. Bayreuth: Buchner, 1839. H. 1. S. 16-31.
Münster G.G. Die Versteinerungen des Uebergangskalkes mit Clymenien und Orthoceratiten von Oberfranken // Beitr. zur Petrefactenkunde. Bayreuth: Buchner, 1840. H. 3. S. 33–121.
Price J.D., House M.R. Ammonoids near the Devonian–Carboniferous boundary // Cour. Forsch.-Inst. Senckenb. 1984. V. 67. P. 15–22.
Qie Wenkun, Liu J., Chen J. et al. Local overprints on the global carbonate δ13C signal in Devonian–Carboniferous boundary successions of South China // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2015. V. 41. P. 290–303.
Ruan Y. Devonian and earliest Carboniferous ammonoids from Guangxi and Guizhou // Mem. Nanjing Inst. Geol. Palaeontol. Acad. Sin. 1981. V. 15. P. 1–152.
Schindewolf O.H. Zur Stratigraphie und Paläontologie der Wocklumer Schichten (Oberdevon) // Abhandl. Preuss. Geol. Landesanst. N. F. 1937. Bd 178. P. 1–132.
Schönlaub H.P., Feist R., Korn D. The Devonian–Carboniferous Boundary at the section “Grüne Schneid” (Carnic Alps, Austria): A preliminary report // Cour. Forsch.-Inst. Senckenb. 1988. V. 100. P. 149–167.
Sheng H. Ammonoids // Ji Q., Wei J.R., Wang Z.J. et al. The Dapoushang section–An Excellent Section for the Devonian-Carboniferous Boundary Stratotype in China. Beijing: Sci. Press, 1989. P. 108–119.
Vöhringer E. Die Goniatiten der unterkarbonischen Gattendorfia-Stufe im Hönnetal (Sauerland) // Fortschr. Geol. Rheinland und Westfalen. 1960. Bd 3. № 1. S. 107–196.
Walliser O.H. Pleading for a natural D/C-boundary // Cour. Forsch.-Inst. Senckenb. 1984. V. 67. P. 241–246.
Walliser O.H. Global events in the Devonian and Carboniferous // Global Events and Event Stratigraphy in the Phanerozoic / Ed. O.H. Walliser. Berlin: Springer, 1996. P. 225–250.
Zhang M., Becker R.T., Ma X. et al. Hangenberg Black Shale with cymaclymeniid ammonoids in the terminal Devonian of South China // Palaeobiodiv. and Palaeoenvironm. 2019. V. 99. № 1. P. 129–142.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал