

Палеонтологический журнал, 2021, № 6, стр. 24-32
Апт–альбские устрицы юго-западных отрогов Гиссарского хребта. Статья 1: род Amphidonte Fischer de Waldheim
Е. К. Метелкин a, b, *, И. Н. Косенко a, b, c, **
a Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН
Новосибирск, Россия
b Новосибирский государственный университет
Новосибирск, Россия
c Нанкинский институт геологии и палеонтологии Китайской академии наук
Нанкин, Китай
* E-mail: MetelkinEK@ipgg.sbras.ru
** E-mail: KosenkoIN@ipgg.sbras.ru
Поступила в редакцию 25.02.2021
После доработки 18.03.2021
Принята к публикации 18.03.2021
Аннотация
Ревизованы представители рода Amphidonte Fischer de Waldheim, 1829 из апта и альба юго-западных отрогов Гиссарского хребта. В апте род представлен новым видом A. mirkamalovi sp. nov., в альбе – видом A. zachanensis Mirkamalov, 1966.
ВВЕДЕНИЕ
Апт–альбский этап хорошо выделяется в эволюции меловых устриц. Он является переходным между раннемеловым этапом (берриас–барремским), который характеризуется невысоким таксономическим разнообразием и широким распространением родов Aetostreon, Ceratostreon и Rastellum (Dhondt et al., 1999; Косенко, Метелкин, 2020; Toscano, Lazo, 2020) и первым появлением пикнодонтин (Kosenko, 2018), и позднемеловым (сеноман–маастрихтским), характеризующимся максимальным для мезозоя разнообразием устриц (только подсемейство Exogyrinae насчитывает не менее семи таксонов родового/подродового ранга: Amphidonte, Ceratostreon, Vultogryphaea, Costagyra, Exogyra, Rhynchostreon, Ilymatogyra) (Stenzel, 1971; Malchus, 1990, 1996; Aqrabawi, 1993; Dhondt et al., 1999; и др.). В апте и альбе происходит быстрая радиация экзогирин, которая приводит к появлению новых родов и видов (Malchus, 1996). В это время широко распространяются роды Amphidonte и Gryphaeostrea, появляются и быстро эволюционируют первые представители трибы Exogyrini, расширяются ареалы пикнодонтин и, в то же время, сохраняются элементы, присущие раннемеловому этапу (род Aetostreon). Вместе с тем, апт–альбские устрицы менее изучены по сравнению с ранне- и позднемеловыми, и ряд вопросов, связанных с происхождением, эволюцией и путями миграции ранних Exogyrini, до сих пор остаются дискуссионными (Malchus, 1996). В связи с этим особый интерес представляют новые данные по таксономическому составу, стратиграфическому и палеобиогеографическому распространению аптских и альбских устриц, в особенности из восточных районов Тетиса, к каковым и относятся юго-западные отроги Гиссарского хребта – устрицы оттуда менее исследованы.
Ранее меловые устрицы Гиссара изучались Х.Х. Миркамаловым (1965, 1966, 1971а, б, 1986). Им было проведено монографическое изучение меловых устриц Гиссара, описаны новые роды и виды, внесены существенные изменения в систематику и филогению экзогир, а также показано значение устриц для стратиграфии мела юго-западных отрогов Гиссарского хребта. Особое внимание Миркамалов уделял экзогирам, доминировавшим в меловых сообществах устриц Гиссара и представленным родами Exogyra, Amphidonte, Gyrostrea (в настоящее время род исключен из подсемейства Exogyrinae), Pseudogyra и Rhynchostreon. За прошедшее с выхода работ Миркамалова время изменились представления о систематике меловых устриц, а также накопились новые данные по их палеобиогеографическому и стратиграфическому распространению (Freneix, 1982; Malchus, 1990, 1996, 1998; Cooper, 1995, 1997; Seeling, Bengtson, 1999; Kosenko, 2018; Косенко, Метелкин, 2020; и др.). Кроме того, в работах Миркамалова не всегда уделялось должное внимание внутривидовой изменчивости устриц. Это приводит к необходимости ревизии апт–альбских устриц Гиссара.
Изучение нового представительного палеонтологического материала, переданного авторам из Всероссийского научно-исследовательского геологического института им. А.П. Карпинского (ВСЕГЕИ), позволяет ревизовать таксономический состав устриц из апта и альба юго-западных отрогов Гиссарского хребта и на основе проведенной ревизии уточнить сведения об их стратиграфическом и палеобиогеографическом распространении.
Настоящая статья представляет собой первую статью из цикла, посвященного ревизии апт–альбских устриц юго-западных отрогов Гиссарского хребта. В ней мы подробно рассматриваем представителей рода Amphidonte.
МАТЕРИАЛ
Материалом для исследования послужила представительная коллекция устриц из апт–сеноманских отложений юго-западных отрогов Гиссарского хребта, собранная в разные годы сотрудниками ВСЕГЕИ и в 2015 г. переданная для изучения в Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН (ИНГГ) Т.Н. Богдановой. Значительную часть коллекции составляют аптские и альбские устрицы, сеноманские устрицы немногочисленны. Коллекция происходит из семи обнажений (рис. 1), насчитывает около 650 раковин хорошей сохранности, которые характеризуются точной географической и стратиграфической привязкой (к пачкам свит, большинство из которых охарактеризовано находками аммонитов).
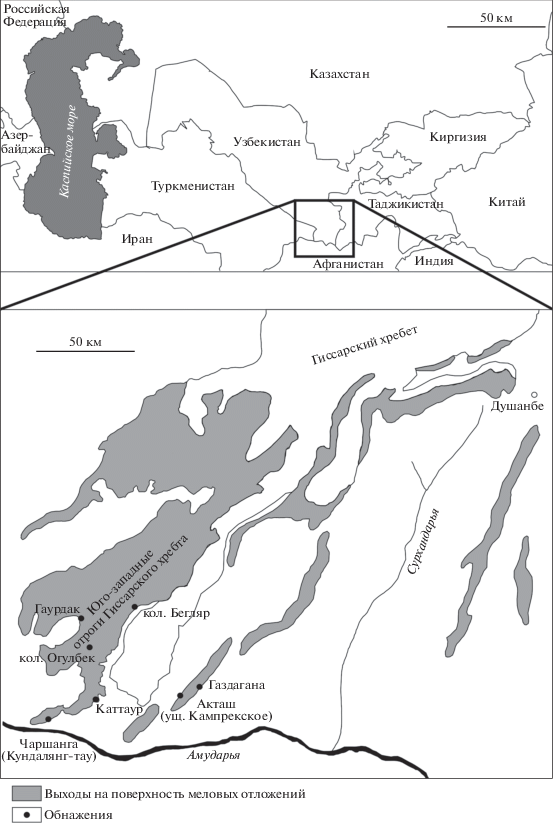
СВЕДЕНИЯ О СТРАТИГРАФИИ АПТА–СЕНОМАНА ЮГО-ЗАПАДНЫХ ОТРОГОВ ГИССАРСКОГО ХРЕБТА
Апт–сеноман юго-западных отрогов Гиссара представлен кундалянгтауской, лялимканской, калигрекской, кулькамышской, чаршангинской, каламазарской, лучакской, аккапчигайской, ширабадской и тюбегатанской свитами (Стратиграфический…, 2001).
Кундалянгтауская свита согласно залегает на мачайской свите позднебарремского возраста. Свита сложена внизу глинами серыми и темно-серыми с прослоями глинистых известняков, вверху – чередованием алевролитов, глин, зеленовато- и коричневато-серых известняков. Раннеаптский возраст свиты установлен по находкам аммонитов Deshayesites ex gr. antiquas Bogdanova, 1983.
Лялимканская свита согласно залегает на кундалянгтауской свите. Свита сложена красными и серыми алевролитами и глинами с прослоями известняков и гипсов. Мощность ее 25–70 м. Свита охарактеризована фораминиферами и остракодами Antepaijenborchella praealta turkmenica (Andreev, 1988), также содержит комплекс спор и пыльцы. По находкам микрофауны свита датирована началом среднего апта.
Калигрекская свита согласно залегает на лялимканской. Свита сложена серыми глинами, алевролитами, песчаниками с прослоями известняков и красноцветных песчаников. Мощность свиты 40–95 м. Ее среднеаптский возраст установлен по находкам аммонитов Parahoplites melchioris (Anthula, 1899).
Кулькамышская (=беглярская) свита с размывом залегает на калигрекской свите. Свита сложена конгломератами, гравелитами, песчаниками с прослоями глин и ракушняков. Мощность 10–20 м. Позднеаптский возраст свиты установлен по находкам аммонитов Acanthohoplites nolani (Seunes, 1887).
Чаршангинская свита согласно залегает на кулькамышской свите. Чаршангинская свита подразделена на две подсвиты. Нижняя сложена темно-серыми глинами с конкрециями мощностью 30–75 м. Ее позднеаптский возраст установлен по находкам аммонитов Hypacanthoplites jacobi Collet, 1907, H. karlukensis Luppov, 1961, H. elegans Fritel, 1906. Верхняя подсвита сложена темно- и зеленовато-серыми глинами с прослоями алевролитов и песчаников, в основании с фосфоритовой галькой и переотложенными (из нижней подсвиты) ядрами аммонитов, морских двустворчатых моллюсков и гастропод. Ее мощность 10–35 м. По находкам аммонитов Douvilleiceras scabrosum Casey, 1962 возраст подсвиты установлен как начало раннего альба.
Каламазарская свита согласно залегает на чаршангинской. Каламазарская свита сложена темно- и зеленовато-серыми глинами с прослоями песчаников, известняков, ракушняков, участками с прослоями красновато-коричневых алевролитов. Мощность 25–85 м. Раннеальбский возраст свиты установлен по находкам аммонитов Douvilleiceras mammilatum (Schlotheim, 1813), Cleoniceras mangyshlakensis Luppov, 1949.
Лучакская свита с размывом залегает на каламазарской. Лучакская свита сложена переслаивающимися зеленовато-серыми карбонатными глинами и ракушняками. Мощность 50–120 м. Среднеальбский возраст свиты установлен по находкам аммонитов Hoplites cf. baylei Spath, 1925, H. dentatus (Sowerby, 1821), Epihoplites trapezoidalis Luppov, 1961. Верхняя часть относится к верхнему альбу по находкам Anahoplites rossicus.
Аккапчигайская свита согласно залегает на лучакской свите. Свита сложена глинами темно- и зеленовато-серыми с тонкими (0.1–0.4 м) прослоями ракушняков. Мощность свиты 30–135 м. Позднеальбский возраст свиты установлен по находкам аммонитов Mortoniceras inflatum gibbosum Spath, 1930, Hysteroceras carinatum Spath, 1922, Semenovites michalskii (Semenov, 1899).
Ширабадская свита согласно залегает на аккапчигайской. Свита сложена известняками, песчаниками серыми, зеленовато-серыми с прослоями глин, в средней части – с прослоями гипсов и красных глин. Мощность 20–60 м. Позднеальбский возраст свиты установлен по аммонитам Karamaiceras kolbajensis Sokolov, 1967.
Тюбегатанская (=дербентская) свита согласно залегает на ширабадской. Свита сложена серыми и зеленовато-серыми глинами с прослоями (0.3–1.5 м) песчаников и ракушняков. Мощность 60–190 м. Сеноманский возраст свиты установлен по находкам аммонитов Karamaites gaurdakensis Luppov, 1963 и Mediasiaceras beliakovae Iljin, 1975.
ОБСУЖДЕНИЕ
В результате изучения коллекции уточнены сведения о таксономическом составе устриц в апте–сеномане юго-западных отрогов Гиссарского хребта и об их стратиграфическом распространении (рис. 2).
Рис. 2.
Стратиграфическое распространение устриц в апте–сеномане юго-западных отрогов Гиссарского хребта.

Из апта определены Pseudogyra pennata Mirkamalov, 1971, Ceratostreon trigonalis (Mirkamalov, 1971), Oscillopha eos (Coquand, 1869), Amphidonte mirkamalovi sp. nov., Costagyra luppovi (Mirkamalov, 1963), Aetostreon sp. и Ceratostreon sp. Также Миркамалов (1966) указывал на присутствие устриц Aetostreon ex gr. caucasicum (Mordvilko, 1960) [в настоящее время это видовое название признано синонимом вида Aetostreon mordvilkoae (Muzapharova, 1953)] (Атлас…, 2004).
Из альба определены Pseudogyra pennata, Ceratostreon trigonalis, Costagyra luppovi, Oscillopha eos и Amphidonte zachanensis Mirkamalov, 1966. Кроме того, Миркамалов (1966) описал из альба вид Costagyra canensis (Mirkamalov, 1966).
Из сеномана определены Oscillopha dichotoma (Bayle, 1849), Amphidonte sp., Costagyra luppovi, Pycnodonte (s. l.) sp. и Rhynchostreon sp.
Ниже представлены результаты ревизии представителей рода Amphidonte (рис. 2).
ОПИСАНИЕ ТАКСОНОВ
НАДСЕМЕЙСТВО OSTREOIDEA RAFINESQUE, 1815
СЕМЕЙСТВО GRYPHAEIDAE VIALOV, 1936
ПОДСЕМЕЙСТВО EXOGYRINAE VIALOV, 1936
Род Amphidonte Fischer de Waldheim, 1829
Диагноз. Гладкие или с концентрической скульптурой роста экзогиры с хоматами, развитыми вдоль всего внутреннего периметра смычного края. Для микроструктуры характерно присутствие правильной листоватой структуры “селедочная кость”.
Amphidonte zachanensis Mirkamalov, 1966
Табл. III , фиг. 6–12 ; табл. IV , фиг. 1–10 (см. вклейкy)
Amphidonta conica: Винокурова, 1963, с. 246, табл. VII , фиг. 3–7 .
Amphidonte conica: Миркамалов, 1966, с. 68, табл. VII , фиг. 6–8 , табл. IX , фиг. 8–12 , табл. X , фиг. 7–8 , табл. XIV , фиг. 11–12 .
Amphidonte arduennensis: Миркамалов, 1966, с. 70, табл. VIII , фиг. 7–11 ; 1986, с. 63, табл. XVIII, фиг. 2–3.
Amphidonte arduennensis var. zeravschanica: Миркамалов, 1966, с. 71, табл. VIII , фиг. 12–17 .
Amphidonte zachanensis: Миркамалов, 1966, с. 71, табл. VIII , фиг. 1–6 ; 1986, с. 63, табл. IX , фиг. 1–2, табл. XVIII , фиг. 8 .
Amphidonte chaliotidea: Миркамалов, 1986, с. 62, табл. VI , фиг. 5, 8 , табл. XVIII , фиг. 7 .
Голотип – Ин-т геологии и разведки нефтяных и газовых месторождений (ИГИРНИГМ), № 19; Узбекистан, юго-западные отроги Гиссарского хребта, пос. Дербент; средний альб (Миркамалов, 1966, табл. VIII , фиг. 3 ).
Описание. Раковина от маленьких до средних размеров (высота раковины от 10 до 54 мм, длина от 11 до 50 мм), вытянутая в высоту, треугольно-овальной формы, неравносторонняя, неравностворчатая.
Левая створка сильно выпуклая (коэффициент выпуклости Вып/Д от 0.4 до 0.86; среднее значение – 0.65), от треугольно-овального до овального и уховидного очертания. Передненижний край округлый, задний прямой, иногда вогнутый или слегка выпуклый. Макушка массивная, прижата к заднему краю створки, не выступает за плоскость смыкания створок. Площадка прирастания от небольшого до среднего размера, расположена вблизи макушки. От макушки к нижнему краю протягивается киль. Киль становится четким по мере роста раковины: у маленьких раковин он округлый, у крупных становится острым, иногда бугорчатым за счет сильно выраженных чешуй нарастания. Скульптура раковины представлена концентрическими линиями нарастания, с возрастом переходящими в грубые чешуи роста, которые образуют на киле бугорки, более обозначенные у крупных раковин. Примакушечная полость хорошо развита. Под замочной площадкой у заднего края имеется небольшая парадонтальная выемка. Отпечаток мускула-замыкателя треугольно-овальной формы, расположен вблизи заднего края и слегка смещен к нижнему краю. Смычной край узкий. По периметру смычного края имеются хорошо проявленные прямые хоматы. Отпечаток мускула Квенштедта на изученном материале не наблюдался.
Правая створка слабовыпуклая, уховидного очертания. Передненижний край округлый, задний край прямой или слегка выпуклый. Передний край створки утолщен за счет плотно прижатых друг к другу слоев нарастания раковины. К переднему краю створки зачастую приурочены реликтовые хоматы. Скульптура створки представлена концентрическими линиями роста. Между замочной площадкой и внутренней частью створки имеется перегиб. Смычной край узкий, сзади он расширяется и образует небольшую платформу, покрытую пустулообразными хоматами. Вдоль внутреннего периметра смычного края имеются хорошо выраженные прямые хоматы. На задневерхнем крае под замочной площадкой имеется зубовидный выступ, соответствующий парадонтальной выемке на левой створке. Зубовидный выступ покрыт хоматами. Отпечаток мускула-замыкателя небольшого размера, треугольно-округлой формы, расположен вблизи заднего края и слегка смещен к нижнему краю створки.
Размеры в мм и отношения:
| № экз. | В | Д | Вып | В/Д | Вып/Д |
|---|---|---|---|---|---|
| 2105/42 | 26 | 19 | 12 | 1.39 | 0.65 |
| 2105/51 | 45 | 34 | 28 | 1.32 | 0.81 |
| 2105/60 | 21 | 19 | 13 | 1.11 | 0.68 |
| 2105/78 | 27 | 21 | 13 | 1.24 | 0.61 |
| 2105/114 | 21 | 19 | 12 | 1.06 | 0.63 |
| 2105/115 | 21 | 18 | 11 | 1.19 | 0.64 |
| 2105/172 | 25 | 21 | 12 | 1.16 | 0.57 |
| 2105/179 | 50 | 42 | 18 | 1.17 | 0.44 |
| 2105/186 | 46 | 34 | 24 | 1.38 | 0.71 |
| 2105/190 | 30 | 25 | 17 | 1.18 | 0.65 |
| 2105/198 | 27 | 22 | 15 | 1.27 | 0.69 |
| 2105/202 | 36 | 30 | 25 | 1.18 | 0.82 |
| 2105/223 | 24 | 20 | 14 | 1.19 | 0.72 |
| 2105/238 | 47 | 38 | 31 | 1.24 | 0.82 |
Изменчивость. Возрастной изменчивости подвержена скульптура роста. У маленьких раковин она представлена тонкими линиями нарастания, в то время как у крупных раковин скульптура роста грубая, представлена чешуями роста. Кроме того, с возрастом изменяется степень выраженности киля: у маленьких раковин он округлый, у крупных становится хорошо выраженным. Степень выпуклости раковин также меняется с возрастом: мелкие раковины относительно более выпуклые по сравнению с крупными.
Внутривидовая изменчивость затрагивает в основном очертания раковин. Очертания мелких раковин изменяются от треугольно-овальных, вытянутых в высоту, до треугольно-овальных, вытянутых в длину. Очертания крупных раковин более однообразны, все они вытянуты в высоту.
Сравнение. A. zachanensis наиболее близок к широко распространенному в сеномане Европы (за исключением Средиземноморья) виду A. haliotideum (Sowerby, 1813) (Sowerby, 1813, с. 67, табл. 25 , фиг. 1–5 ; Собецкий, 1977, с. 160–161, табл. 15 , фиг. 1–3 ). A. zachanensis отличается от этого вида более широкой раковиной, сильно развитым, зачастую бугорчатым килем, меньшим размером площадки прирастания, менее выраженной задней платформой и грубой концентрической скульптурой роста.
От A. humboldtii Fischer de Waldheim, 1829 (Fischer de Waldheim, 1829, с. 177, фиг. 1; Собецкий, 1977, с. 159–160, табл. 14 , фиг. 15 ), распространенного в сеномане Русской платформы, отличается развитым килем и грубой концентрической скульптурой роста.
По наличию хорошо обозначенного киля описываемый вид похож на A. malchusi Cooper, 1997 (Cooper, 1997, с. 10–14, рис. 10–12 ) из верхов альба (зона Stoliczkaia dispar) Юго-Восточной Африки, от которого отличается менее завернутой макушкой, грубой концентрической скульптурой роста, наличием выраженной площадки прирастания, отсутствием радиальной борозды (сулькуса) и теребратулоидной складки на нижнем крае раковины.
От A. mirkamalovi sp. nov. отличается менее завернутой макушкой, треугольно-овальной формой отпечатка мускула-замыкателя и более выраженным килем. От A. conica описываемый вид отличается более обозначенным килем, треугольно-овальной формой отпечатка мускула-замыкателя и характером скульптуры роста. У A. conica скульптура представлена концентрическими линиями нарастания, а для A. zachanensis характерны более проявленные грубые чешуи нарастания.
Замечания. Экземпляры A. zachanensis c хорошо развитым килем напоминают представителей рода Aetostreon Bayle, 1878. От них описываемый вид отличается наличием хомат по всему периметру смычного края, субтреугольным отпечатком мускула-замыкателя, смещенным к заднему краю, гораздо более закрученными правыми створками, а также выраженным выступом переднего края правой створки, образованным наслаивающимися друг на друга слоями раковины.
Распространение. Альб юго-западных отрогов Гиссарского хребта.
Материал. 131 раковина из каламазарской свиты (Кампрекское ущелье – 38 экз., колодец Бегляр – 34 экз., Гаурдак – 36 экз., колодец Огулбек – 23 экз.); 39 раковин из лучакской свиты (Кундалянгтау – 24 экз., Бегляр – 4 экз., Гаурдак – 11 экз.); две раковины из верхов лучакской свиты (Кампрекское ущелье).
Amphidonte mirkamalovi Metelkin et Kosenko, sp. nov.
Табл. III , фиг. 1–5
Название вида – в память палеонтолога Х.Х. Миркамалова, специалиста по стратиграфии мела и меловым двустворчатым моллюскам Cредней Азии.
Голотип – ЦКП “Геохрон” (ИНГГ СО РАН), № 2105/11; Узбекистан, юго-западные отроги Гиссарского хребта, Кампрекское ущелье; верхний апт, чаршангинская свита. ZooBank LSID: urn:lsid:zoobank.org:act:D39AA260-D1FF-4206-96DB-4EFEEB16AB48.
Описание. Раковина маленького размера (от 16 до 32 мм в высоту, средняя высота – 24 мм), вытянутая в высоту (коэффициент вытянутости в высоту В/Д варьирует от 1.02 до 1.21; среднее значение – 1.13), уховидно-овального очертания, очень толстостенная, неравностворчатая, неравносторонняя.
Левая створка сильновыпуклая (коэффициент выпуклости от 0.46 до 0.72; среднее значение – 0.59), треугольно-овального очертания. Передненижний край округлый, задний прямой или слабовогнутый. Макушка очень массивная, закрученная в геликоидальную спираль, прижата к заднему краю. Площадка прирастания отсутствует. От макушки к задненижнему краю протягивается округлый киль, развитый одинаково на всех стадиях онтогенеза. Скульптура раковины представлена концентрическими линиями роста. Примакушечная полость хорошо выражена. Отпечаток мускула-замыкателя небольшого размера, округлой формы. Он расположен ближе к заднему краю и равноудален от верхнего и нижнего краев. Смычной край узкий, немного расширяется на заднем крае створки. Имеется небольшая парадонтальная выемка. По периметру смычного края расположены прямые хоматы. На заднем крае в месте расширения створки они становятся червеобразными. Отпечаток мускула Квенштедта на изученном материале не наблюдался.
Правая створка слабовыпуклая, имеет уховидную форму. Передний край створки утолщен за счет плотно прижатых друг к другу слоев нарастания раковины. К переднему краю створки зачастую приурочены реликтовые хоматы. Скульптура правой створки представлена концентрическими линиями роста. Смычной край узкий, у заднего края образует расширение. Прямые хоматы расположены по внутреннему периметру смычного края. На задневерхнем крае имеется зубовидный выступ, соответствующий парадонтальной выемке на левой створке. На поверхности зубовидного выступа имеются хоматы. Отпечаток мускула-замыкателя небольшого размера, имеет трапециевидно-округлую форму, равноудален от верхнего и нижнего края.
Размеры в мм и отношения:
| № экз. | В | Д | Вып | В/Д | Вып/Д |
|---|---|---|---|---|---|
| 2105/1 (Голотип) | 32 | 28 | 16 | 1.15 | 0.58 |
| 2105/2 | 19 | 18 | 9 | 1.09 | 0.53 |
| 2105/4 | 26 | 24 | 13 | 1.09 | 0.53 |
| 2105/5 | 28 | 25 | 17 | 1.14 | 0.70 |
| 2105/13 | 26 | 25 | 11 | 1.02 | 0.46 |
| 2105/14 | 23 | 20 | 12 | 1.16 | 0.61 |
| 2105/19 | 21 | 18 | 11 | 1.15 | 0.63 |
| 2105/21 | 24 | 20 | 13 | 1.16 | 0.63 |
| 2105/23 | 23 | 19 | 9 | 1.21 | 0.48 |
| 2105/28 | 18 | 16 | 11 | 1.14 | 0.67 |
| 2105/29 | 19 | 17 | 12 | 1.16 | 0.72 |
Сравнение. Описываемый вид отличается от A. zachanensis толстостенностью раковины, массивной, завернутой в геликоидальную спираль макушкой, сглаженным округлым килем и более тонкой концентрической скульптурой роста.
От широко распространенного в верхах нижнего мела – сеномане Северного полушария вида A. conica (Sowerby) (Sowerby, 1813, с. 69, табл. 26 , фиг. 3 ) описываемый вид отличается более массивной макушкой, закрученной в геликоидальную спираль, вытянутыми в высоту очертаниями раковины и более сглаженным килем.
Замечания. Сильно закрученная геликоидальная макушка левой створки делает описываемый вид похожим на виды рода Ilymatogyra Stenzel, 1971 (Stenzel, 1971, с. N1119; Malchus, 1990, с. 119; Aqrabawi, 1993, с. 69), но отличается от них хоматами, развитыми вдоль всего периметра смычного края, и присутствием простой листоватой структуры “селедочная кость” (herringbone regular foliated microstructure) в микроструктуре раковины.
Материал. 34 левые створки и четыре правые створки хорошей сохранности из типового местонахождения.
* * *
Авторы благодарны Т.Н. Богдановой за переданную для изучения коллекцию устриц, а также за полезные советы, помощь в подборе литературы и замечания при работе над гиссарской коллекцией.
Работа выполнена при поддержке Китайской академии геологических наук (проект DD20190009), Национального фонда естественных наук Китая (проект 41730317) и проекта ФНИ № 0331-2019-0004 “Палеонтология, стратиграфия, биогеография бореальных и смежных с ними палеобассейнов и комплексное обоснование усовершенствования региональных стратиграфических схем мезозоя и кайнозоя Сибири”. Работа является вкладом в проект UNESCO-IUGS 679.
Список литературы
Атлас важнейших групп фауны мезозойско-кайнозойских отложений Северного Кавказа и Предкавказья. СПб.: Изд-во ВСЕГЕИ, 2004. 126 с.
Винокурова Е.Г. Меловые двустворчатые и головоногие моллюски из юго-западных отрогов Гиссарского хребта // Стратиграфия и палеонтология Узбекистана и сопредельных районов. Ташкент: Изд-во АН УзССР, 1963. С. 233–262.
Косенко И.Н., Метелкин Е.К. Раннемеловые устрицы Мангышлака: таксономический состав, стратиграфическое и палеобиогеографическое распространение // Палеонтол. журн. 2020. № 3. С. 20–31.
Миркамалов Х.Х. Устрицы альбских отложений юго-западных отрогов Гиссарского хребта // Бюлл. МОИП, отд. геол. 1965. Т. 40. № 6. С. 97–107.
Миркамалов Х.Х. Экзогиры, их систематика и значение для стратиграфии меловых отложений юго-западных отрогов Гиссарского хребта. Ташкент: ФАН УзССР, 1966. 133 с.
Миркамалов Х.Х. Устрицы окузбулакской свиты (баррем) юго-западных отрогов Гиссарского хребта // Бюлл. МОИП, отд. геол. 1971а. Т. 46. № 1. С. 114–123.
Миркамалов Х.Х. Новые виды семейства устричных юго-западных отрогов Гиссарского хребта // Палеонтол. журн. 1971б. № 2. С. 36–40.
Миркамалов Х.Х. Стратиграфия и фауна меловых отложений юго-западных отрогов Гиссарского хребта. Ташкент: ФАН УзССР, 1986. 104 с.
Собецкий В.А. Двустворчатые моллюски позднемеловых платформенных морей юго-запада СССР. М.: Недра, 1977. 256 с.
Стратиграфический словарь Узбекистана. Ташкент: ГИДРОИНГЕО, 2001. 580 с.
Aqrabawi M. Oysters (Bivalvia-Pteriomorphia) of the Upper Cretaceous rocks in Jordan. Palaeontology, stratigraphy and comparison with the Upper Cretaceous oysters of Northwest Europe // Mitt. Geol.-Paläontol. Inst. Univ. Hamburg. 1993. Bd 75. P. 1–135.
Cooper M.R. Exogyrid oysters (Bivalvia: Gryphaeoidea) from the Cretaceous of southeast Africa. Part 1 // Durban Mus. Novit. 1995. V. 20. P. 1–48.
Cooper M.R. Exogyrid oysters (Bivalvia: Gryphaeoidea) from the Cretaceous of southeast Africa. Part 2 // Durban Mus. Novit. 1997. V. 22. P. 1–31.
Dhondt A.V., Malchus N., Boumaza L. et al. Cretaceous oysters from North Africa: origin and distribution // Bull. Soc. Géol. Fr. 1999. V. 170. № 1. P. 67–76.
Fischer de Waldheim G. Sur les fossils des corps organisés // Bull. Soc. Imp. Natur. Moscou. 1829. V. 1. № 2. P. 27–32.
Freneix S. Disparité microstructurale du test entre Gryphaeostrea et Gyrostrea (huitres du Crétacé–Miocène). Novelle classification du genres // Ann. Paléontol. (Invertébr.). 1982. V. 66. № 3. P. 67–134.
Kosenko I.N. The origin of the Pycnodonteinae and relationship between gryphaeas and true pycnodontes // Acta Palaeontol. Pol. 2018. V. 63. № 4. P. 769–778.
Malchus N. Revision der Kreide-Austern (Bivalvia: Pteriomorphia) Ägyptens (Biostratigraphie, Systematik) // Berlin. Geowiss. Abh. Reihe A. 1990. Bd 125. S. 1–231.
Malchus N. Palaeobiogeography of Cretaceous oysters (Bivalvia) in the Western Tethys // Mitt. Geol.-Paläontol. Inst. Univ. Hamburg. 1996. Bd 77. P. 165–181.
Malchus N. Multiple parallel evolution and phylogenetic significance of shell chambers and chomata in the Ostreoidea (Bivalvia) // Bivalves: An Eon of Evolution. Kansas: Univ. Calgary Press, 1998. P. 393–407.
Seeling J., Bengston P. Cenomanian oysters from the Sergipe Basin, Brazil // Cret. Res. 1999. V. 20. № 6. P. 747–765. https://doi.org/10.1006/cres.1999.0190
Sowerby J. The Mineral Conchology of Great Britain. L.: Meredith, 1813. 234 p.
Stenzel H.B. Oysters // Treatise on Invertebrate Paleontology. Part N. Mollusca 6 (Bivalvia). V. 3 / Ed. Moore R.C. Boulder: Geol. Soc. America, Univ. Kansas Press, 1971. P. N953–N1124.
Toscano A.G., Lazo D.G. Taxonomic revision and palaeobiogeographic affinities of Berriasian–Valanginian oysters from the Vaca Muerta and Mulivhinco formations, southern Mendoza, Neuquen Basin, Argentina // Cret. Res. 2020. V. 109. 104358. https://doi.org/10.1016/j.cretres.2019.104358
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал