

Палеонтологический журнал, 2022, № 1, стр. 75-84
Стрекозы семейства Kennedyidae (Odonata: Archizygoptera) из среднего-верхнего триаса Киргизии
А. С. Фелькер *
Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
* E-mail: felkafelka95@gmail.com
Поступила в редакцию 24.12.2020
После доработки 13.04.2021
Принята к публикации 20.04.2021
- EDN: OIUHZX
- DOI: 10.31857/S0031031X22010056
Аннотация
Из средне–верхнетриасовых отложений киргизского местонахождения Джайлоучо (Мадыген) описано два новых вида стрекоз семейства Kennedyidae: Kennedya madygensis sp. nov. и K. ferganensis sp. nov. Они были выделены на основе сборов 2007 и 2009 гг. Дается переописание вида K. carpenteri Pritykina, 1981. Отдельно обсуждаются особенности строения узелка кеннедиид.
ВВЕДЕНИЕ
Урочище Мадыген расположено в предгорьях Туркестанского хребта, близ одноименного кишлака на юго-западе Киргизии (Баткенский р-н, Ошская обл.). Местонахождение является одним из самых крупных по количеству ископаемых остатков насекомых (более 23 тыс.). Оно было открыто в 1933 г. Е.А. Кочневым, который отнес к Мадыгену примерно 10 км2 окружающей территории, позже поделенных на несколько площадей по сторонам света (Shcherbakov, 2008). Насекомоносные слои входят в состав мадыгенской свиты, представленной преимущественно чередованием обломочных пород с подчиненным количеством пластов углей и глинистых сланцев (Сикстель, 1960; Shcherbakov, 2008). По остаткам флоры возраст свиты установлен как ладин–карнийский (Добрускина, 1982), однако большая часть остатков насекомых представлена видами, характерными для карния (Shcherbakov, 2008). В то же время, в отложениях довольно многочисленны докарнийские формы из отрядов Blattodea, Odonata, Orthoptera, Protorthoptera s.l., что не позволяет делать однозначные выводы о времени образования основных насекомоносных слоев и предполагает использование неопределенной датировки ладин–карний в качестве наиболее приемлемой (Притыкина, 1981; Shcherbakov, 2008). Опубликовано немало работ с более подробными данными по истории изучения, геологическому строению и палеонтологическому составу комплекса местонахождения Мадыген (Сикстель, 1960; Притыкина, 1981; Добрускина, 1982; Копылов, 2014; Dobruskina, 1995; Shcherbakov, 2008; Voigt et al., 2017).
Наибольшее число ископаемых остатков насекомых (более 15 тыс. экз.) известно из отложений северной площади Мадыгена, носящей собственное название Джайлоучо. Именно в этой части местонахождения сосредоточен практически весь комплекс стрекоз (Притыкина, 1981). В работе 1981 г. Л.Н. Притыкина разделяет его на три группы по геологическому распространению: эндемики Джайлоучо (сем. Triadophlebiidae, Mitophlebiidae, Zygophlebiidae, Xamenophlebiidae и Batkeniidae), палеозойские реликты, известные из других регионов (сем. Triadotypidae и Kennedyidae) и стрекозы, распространенные в более молодых мезозойских отложениях (сем. Protomyrmeleontidae и Triassolestidae). В последние годы состав первой группы претерпел значительные изменения. Ранее считавшиеся эндемичными триадофлебииды и зигофлебииды сейчас известны из триасовых отложений Оренбуржья (местонахождение Петропавловка, оленекский ярус) и Тунчуаня, Китай (обн. Кишуйхе, ладинский ярус) (Zheng et al., 2017a, b; Фелькер, Василенко, 2019; Shcherbakov et al., 2020).
К палеозойским реликтам Джайлоучо относится единичный экземпляр триасовых представителей инфраотряда Meganeuromorpha – Reisia sogdiana (Pritykina, 1981), сем. Triadotypidae, также известных из Франции и Германии (Притыкина, 1981; Nel et al., 2001). Однако большую часть группы составляют самые молодые представители кеннедиид: Kennedya gracilis Pritykina, 1981 и K. carpenteri Pritykina, 1981. Само семейство Kennedyidae, включаемое в инфраотряд Kennedyomorpha подотряда Archizygoptera, объединяет небольших стебельчатокрылых стрекоз, характерным признаком которых является разреженное жилкование крыльев. Большинство представителей семейства найдено в пермских отложениях Северной Америки (кунгурских местонахождениях Эльмо и Мидко в США), а также Европы: местонахождения Чекарда (Пермский край, кунгурский ярус), Тюлькино (Пермский край, уфимский ярус), Сояна (Архангельская обл., казанский ярус), Китяк (Кировская обл., казанский ярус), Исады (Вологодская обл., северодвинский ярус), Вязовка (Оренбургская обл., северодвинский ярус) (Tillyard, 1925; Carpenter, 1931, 1933, 1939, 1947; Nel et al., 2012; Felker, 2020; Фелькер, 2021б). Единичные кеннедииды найдены в пограничной пермо-триасовой толще западносибирского местонахождения Бабий Камень (Фелькер, 2021а). В большинстве вышеуказанных местонахождений, за исключением Исад, Эльмо и Мидко, представители этого семейства относительно редки и известны только по единичным экземплярам.
В отложениях Джайлоучо представлен довольно крупный комплекс кеннедиид, включающий четыре вида, два из которых были описаны Притыкиной (1981). Однако, учитывая появление нового материала кеннедиид из этого местонахождения, а следовательно, и изменение части диагностических признаков, вид K. carpenteri нуждается в переописании (Nel et al., 2012; Felker, 2020; Фелькер, 2021б).
Ревизуемые в настоящей статье экземпляры типовой серии K. carpenteri были собраны в ходе экспедиций Каратау-Мадыгенского отряда Палеонтологического ин-та АН СССР (ПИН) (начальник отряда А.Г. Шаров) в 1962 г. (экз. ПИН, № 2069/37), 1964–1965 гг. (экз. ПИН, №№ 2240/1798 и 2555/616) и 1967 г. (экз. ПИН, №№ 2785/2, 6). Экз. ПИН, №№ 5330/80 и 5343/195 найдены экспедиционными отрядами ПИН РАН под руководством Е.Н. Курочкина (2007 г.) и А.С. Шмакова (2009 г.).
Все описываемые и ревизуемые виды происходят из местонахождения Мадыген (Джайлоучо), Центральная Азия, Ошская обл., Баткенский р-н, северная площадь урочища Мадыген, на запад-юго-запад от горы Курбан-Таш; средний–верхний триас, ладинский–карнийский ярусы, мадыгенская свита.
При описании материала использованы следующие обозначения жилок: А – анальная жилка; An1, An2 – антенодальные поперечные жилки; Arc – дужка; Asn – заметная поперечная жилка, расположенная между R1 и R2 на интервале от дужки до узелка; br – опорная жилка птеростигмы; C – костальная жилка; CuA – передняя кубитальная жилка; СuP – задняя кубитальная жилка; IR2, IR3 – интеркалярные радиальные жилки; МА – передняя медиальная жилка; N – узелок; n – нодальная жилка (опорная жилка узелка); Pn – постнодальные поперечные жилки; Psn – постсубнодальные поперечные жилки; Pt – птеростигма; q – дискоидальная ячейка; R1, R2, R3, R4+5 – ветви радиуса; Rs – радиальный сектор; Sc – субкостальная жилка; sn – субнодальная поперечная жилка; sq – субдискоидальная ячейка.
Типовой материал хранится в лаб. артропод ПИН РАН, колл. №№ 2069, 2240, 2555, 2785, 5330, 5343.
Автор выражает благодарность Д.В. Василенко за помощь в подготовке рукописи, Р.А. Ракитову за помощь в фотографировании материала на электронном сканирующем микроскопе TeScan Vega3 и А.П. Расницыну за прочтение рукописи и ценные замечания.
Работа поддержана грантом Российского научного фонда 21-14-00284.
ОТРЯД ODONATA
ПОДОТРЯД ARCHIZYGOPTERA
СЕМЕЙСТВО KENNEDYIDAE TILLYARD, 1925
Род Kennedya Tillyard, 1925
Kennedya carpenteri Pritykina, 1981
Табл. IX , фиг. 1–5 (см. вклейку)
Рис. 1.
Измерения крыла кеннедиид: 1 – длина крыла (в скобках указана та же характеристика для реконструированного крыла паратипа Kennedya carpenteri Pritykina ПИН, №2785/6 с учетом деформации сжатия и паратипа ПИН, № 2555/616 – с учетом растяжения); 2 – максимальная ширина на уровне основания IR3; 3 – узелковая ширина; 4 – расстояние от Arc до N; 5 – расстояние от N до Pt; 6 – длина Pt; 7 – ширина Pt.
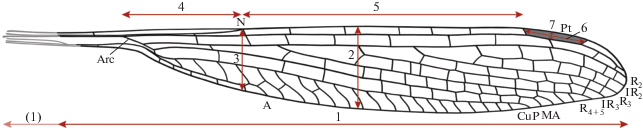
Рис. 2.
Kennedya carpenteri Pritykina, 1981, экземпляры типовой серии, прорисовки: а – голотип ПИН, № 2785/2; б – паратип ПИН, № 2069/37; в – паратип ПИН, № 2785/6; г – паратип ПИН, № 2555/616; Киргизия, Джайлоучо (Мадыген); ладинский–карнийский ярусы, мадыгенская свита. Обозначения названий жилок см. в тексте. Масштабный отрезок – 2 мм.
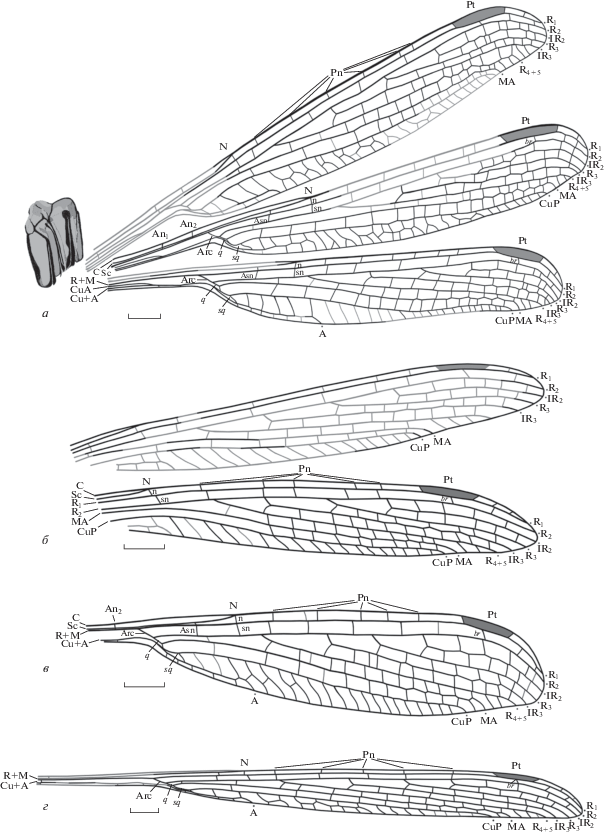
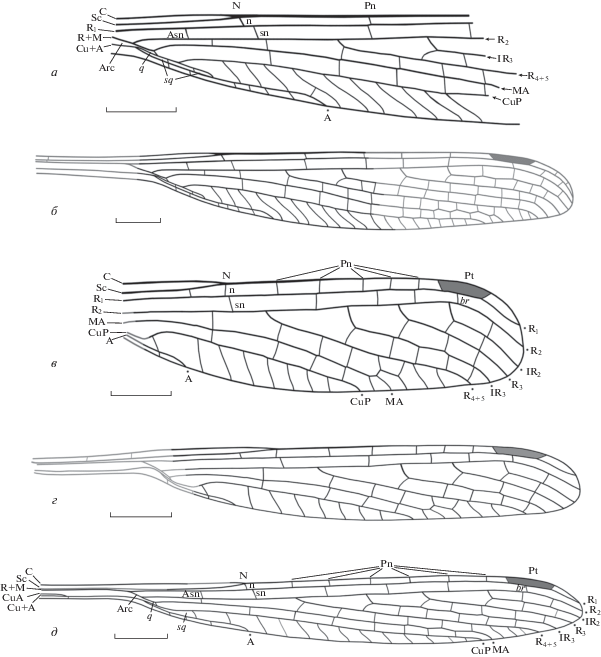
Рис. 3.
Паратип Kennedya carpenteri Pritykina, 1981 и новые виды стрекоз семейства Kennedyidae из средне–верхнетриасовых отложений Киргизии, Джайлоучо (Мадыген); ладинский–карнийский ярусы, мадыгенская свита: а, б – K. carpenteri Pritykina, 1981, паратип ПИН, № 2240/1798: а – прорисовка остатка, б – реконструированное крыло (восстановленные элементы жилкования показаны серым); в, г – K. ferganensis sp. nov., голотип ПИН, № 5343/195: в – прорисовка остатка, г – реконструированное крыло; д – K. madygensis sp. nov., голотип ПИН, № 5330/80, прорисовка остатка (реконструированные элементы жилкования показаны серым). Обозначения названий жилок см. в тексте. Масштабный отрезок – 2 мм.
Kennedya carpenteri: Притыкина, 1981, с. 34, 35, 84, 85, рис. 22 , табл. VII , фиг. 4, 5 , табл. XIII , фиг. 4 ; Nel et al., 2012, с. 15; Felker, 2020, с. 735, 736, 741; Фелькер, 2021а, с. 55, 58; 2021б, c. 45–47, рис. 3, а–д.
Голотип – ПИН, № 2785/2; позитивный и негативный отпечатки трех частично наложенных друг на друга крыльев и птероторакса хорошей сохранности; местонахождение Мадыген (Джайлоучо); средний–верхний триас, ладинский–карнийский ярусы, мадыгенская свита.
Описание (рис. 2, а–г; 3, а, б). N полный по строению, n и sn находятся прямо под ним без смещения друг относительно друга (на экз. ПИН, №№ 2555/616, 2069/37 и 2240/1798 sn немного смещена в дистальном направлении; см. Изменчивость). Asn и продолжающая ее в следующем поле поперечная жилка расположены на одной прямой. Постнодальное и постсубнодальное поля одинаковые по ширине на интервале от N до Pt. Pn жилок не менее 5, Psn – не менее 3, расположены с заметным смещением друг относительно друга. Pt довольно крупная, в длину равна двум–трем нижележащим ячейкам между R2 и Rs; по форме – трапециевидная, ее задний край слегка выпуклый. Опорная жилка Pt (br) расположена под ее серединой.
Дужка (Arc), по-видимому, расположена немного дистальнее An2, расстояние от ее начала до Asn в 1.5 раза больше расстояния от Asn до N. Дискоидальная ячейка (q) открытая (на экз. ПИН, №№ 2555/616 и 2240/1798 слегка деформирована, см. Замечания). Передний край q в полтора раза короче длины Arc. Субдискоидальная ячейка (sq) слегка изогнута и заметно расширена в дистальной части, ее базальный край расположен немного дистальнее уровня ответвления МА от Rs (на экз. ПИН, №№ 2555/616 и 2240/1798 sq деформирована, см. Замечания). Основания ветвей радиуса расположены примерно на одинаковом расстоянии друг от друга. Расстояние между N и основанием R4 + 5 может слегка изменяться на разных экземплярах типовой серии (см. Изменчивость). Основание R4 + 5 расположено немного дистальнее N, основание IR3 – в проксимальной трети расстояния между N и Pt, основание R3 – в дистальной трети того же расстояния, основание IR2 – заметно проксимальнее Pt.
A выходит на край крыла на уровне N или немного дистальнее его, имеет Y-образное соединение с несколькими поперечными жилками на заднем крае крыла. Вершина CuP расположена на уровне проксимальной трети Pt, вершина МА находится на уровне середины Pt, вершины остальных жилок расположены дистальнее Pt. Между дистальной стороной sq и окончанием А находятся от 2 до 5 поперечных жилок; между вершинами А и CuP на задний край крыла выходят не менее 17 поперечных жилок, некоторые из них образуют двойные ряды ячеек; между вершинами CuP и МА – 1–2 поперечные жилки, между МА и R4 + 5 – 2–3 поперечные жилки. Кубито-анальное поле широкое, втрое больше поля между А и задним краем крыла. Интеркалярные жилки и дистальная часть МА слегка зигзагообразные. Поперечное жилкование в дистальной части крыла заметно учащенное.
Размеры в мм (см. таблицу). Измерения показаны на рис. 1.
| Экз. ПИН, № | Признаки | ||||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| 2785/2 (п. кр.) | 34.5 | 4.5 | 3.3 | – | 15.5 | 3.5 | 0.4–0.6 |
| 2785/2 (1-е з. кр.) | 29.3 | – | – | 6.0 | 12.8 | 3.0 | 0.4–0.5 |
| 2785/2 (2-е з. кр.) | 29.6 | 4.7 | 3.9 | 6.4 | 13.2 | 3.1 | 0.4–0.5 |
| 2785/6 | 22.3 (25.1) | 5.1 (3.7) | 3.6 (3.0) | 4.9 | 11.4 | 2.6 | 0.3–0.4 |
| 2555/616 | 38.2 (28.8) | 3.1 (4.2) | 2.2 (3.3) | 6.4 (4.9) | 17.6 (13.4) | 2.9 (2.3) | 0.3 (0.4) |
| 2069/37 | 21.6 (28.2) | 3.4 | 2.5 (2.7) | – | 13.2 | 2.7 | 0.4 |
| 2240/1798 | 11.6 (24.6) | 2.9 (3.4) | 2.1 (2.4) | 4.3 | – | – | – |
Изменчивость. В размерах, а также строении и соотношении некоторых крыловых структур экземпляров типовой серии прослеживается ряд отличий. Реконструированная длина крыльев экземпляров этой серии варьирует в пределах от 24.6 до 29.3 мм (см. Размеры). По-видимому, это вписывается в рамки возможной внутривидовой изменчивости, характерной для рода Kennedya. Так, голотип K. mirabilis из местонахождения Эльмо имеет размеры 44.0 × 5.8 мм; в то же время, экз. MCZ, № 3971 того же вида из указанного местонахождения практически вдвое меньше, его размеры – 25.0 × 3.3 мм (Carpenter, 1933).
Среди вариаций жилкования выделяется различное положение sn относительно n в узелке, а следовательно, и особенности формирования N (см. Обсуждение). На голотипе (рис. 2, а) и экз. ПИН, № 2785/6 (рис. 2, в) sn лежит на одной прямой с n; на экз. ПИН, №№ 2069/37 (рис. 2, б), 2555/616 (рис. 2, г) и 2240/1798 (рис. 3, а) sn смещена дистальнее n и N.
Второй характеристикой, изменяемой внутри экземпляров типовой серии, является положение Asn и основания R4 + 5 относительно N. Так, на голотипе расстояние между N и основанием R4 + 5 втрое (на экз. ПИН, № 2785/6 – вдвое) меньше расстояния между Asn и N, а на экз. ПИН, №№ 2555/616, 2069/37 и 2240/1798 – N равноудален от Asn и основания R4 + 5.
Сравнение. Ревизуемый вид больше других схож с позднепермским видом K. suchonensis в размерах и некоторых особенностях жилкования, в частности, положении основания R4 + 5 и количестве Pn и Psn жилок, но отличается от него открытой q, большей длиной CuP и МА и частым поперечным жилкованием в дистальной части крыла. С K. azari, K. ivensis, K. pritykinae и K. suchonensis сходен формой Pt, с K. gracilis и K. suchonensis – положением br, с K. fraseri и K. suchonensis – длиной и формой А. От всех известных видов отличается большим числом Pn и Psn жилок; от всех видов, за исключением K. suchonensis, – большей длиной и проксимальным положением основания IR2. От большинства известных кеннедий, за исключением K. kedrovkensis и K. suchonensis, отличается большей шириной кубито-анального поля.
Замечания. Несколько экземпляров типовой серии (паратипы ПИН, №№ 2069/37, 2240/1798 и 2555/616) подверглись деформации растяжения вмещающей породы – обычному явлению для находок из Джайлоучо. Методика реконструкции исходного облика остатков, основанная на вычислении усредненных пропорций соотношения длины и ширины ископаемых и современных ксиелид (Hymenoptera, Xyelidae) предложена в работе Д.С. Копылова (2014). Схожим способом получены реконструированные размеры преобразованных крыльев вышеуказанных экземпляров K. carpenteri. В качестве основы для реконструкции были взяты наиболее полные экземпляры типовой серии похожего по форме и жилкованию крыла позднепермского вида K. suchonensis, а также голотип K. carpenteri (по-видимому, недеформированный), для которого известны расположенные в разных плоскостях крылья, по форме и размерам сходные с крыльями кеннедий из других местонахождений.
Экз. ПИН, № 2555/616 (рис. 2, г) заметно вытянут. Вследствие этого некоторые структуры крыла видоизменены: так, наблюдается деформация дискоидальной области крыла. В частности, жилки CuP и R + M (Arc) в составе базального края q заметно сближены, что приводит к образованию клиновидно-вытянутой формы дискоидальной ячейки, сходной с таковой у позднепермского вида K. suchonensis (Исады, северодвинский ярус). В то же время, несмотря на сильное сближение CuP и R + M, дискоидальная ячейка этого экземпляра остается базально открытой, в отличие от той же структуры у северодвинского вида (Фелькер, 2021б). Следует отметить, что сближение CuP и R + M в базальной области q отмечается только на трех экземплярах мадыгенских стрекоз: двух из типовой серии K. carpenteri (экз. ПИН, №№ 2240/1798 и 2555/616), а также голотипе K. gracilis (экз. ПИН, № 2785/23). У остальных кеннедиид с сохранившейся переходной областью от стебелька к крыловой пластинке (голотип и экз. ПИН, № 2785/6 типовой серии K. carpenteri, голотип K. madygensis) q имеет обычную форму с параллельными не сближенными передним и задним краями.
Сильное сжатие экз. ПИН, № 2785/6 (рис. 2, в), вероятно, также обусловлено деформацией вмещающей породы.
Судя по всему, экз. ПИН, № 2240/1798 (рис. 3, а) является аберрантным, вследствие изменения характерного для кеннедиид строения sq. Обычно эта структура представлена цельной треугольной пластинкой, ограниченной с переднего и заднего краев ветвями CuP и А. Дистальный край sq по сути является обычной, слегка наклоненной поперечной жилкой, в интерпретации Р. Тильярда носящей название cua (Tillyard, 1925). У экз. ПИН, № 2240/1798 область, топологически соответствующая sq, имеет овально-вытянутую форму и состоит из трех пластинок.
Материал. Голотип и паратипы ПИН, №№ 2069/37, 2240/1798, 2555/616 и 2785/6 из типового местонахождения.
Kennedya ferganensis Felker, sp. nov.
Табл. IX , фиг. 6
Название вида от Ферганской долины, на юге которой находится местонахождение.
Голотип – ПИН, № 5343/195; позитивный отпечаток, представленный средней частью крыла, и негативный – практически полный; местонахождение Мадыген (Джайлоучо); средний–верхний триас, ладинский–карнийский ярусы, мадыгенская свита.
Описание (рис. 3, в, г). N – полный по строению, n расположена прямо под ним, sn немного смещена в дистальном направлении. Asn находится в 5 раз ближе к уровню вершины А, чем к N. Pn жилок 5, Psn – 3, расположены со смещением друг относительно друга. Pt небольшого размера, трапециевидной формы; передний и задний ее края параллельны друг другу, дистальный край наклонен заметно сильнее проксимального, br расположена на уровне проксимальной трети Pt. Основания ветвей радиуса расположены дистальнее N. Основание R4 + 5 находится вдвое ближе к N, чем к основанию IR3. Основания IR3, R3 и IR2 равноудалены друг от друга. Основание IR2 расположено под проксимальной четвертью Pt.
А короткая, выходит на край крыла заметно проксимальнее N и даже Asn. Вершина CuP находится на уровне дистальной трети расстояния между N и Pt, примерно под основанием IR3. Вершина МА расположена проксимальнее Pt, примерно на уровне основания R3. Между дистальной стороной sq и окончанием А поперечных жилок нет, между вершинами А и CuP на задний край крыла выходят 7 простых поперечных жилок, между вершинами CuP и МА – 1 поперечная жилка, между МА и R4 + 5 – 3, первая из которых Y-образная. IR3 слегка зигзагообразная в дистальной части, остальные продольные жилки практически прямые.
Размеры в мм. Длина сохранившегося остатка (дистальная половина крыла) – 13.4 (примерная длина реконструированного крыла – 17.8), максимальная ширина сохранившегося фрагмента (скорее всего, постбуриально расширенного) – 3.5, узелковая ширина – 3.2. Расстояние от N до Pt – 7.0. Длина Pt – 1.6, ширина Pt – 0.3.
Сравнение. С K. ivensis новый вид сходен очень короткой А, c K. tyulkinensis – положением br. От всех видов рода Kennedya, за исключением K. tyulkinensis, новый вид отличается меньшими размерами крыла и относительно короткой CuP; от K. carpenteri, K. madygensis, K. suchonensis и K. tyulkinensis – меньшей длиной IR2 и дистальным положением ее основания; от K. azari, K. gracilis, K. ivensis, K. mirabilis, K. pritykinae – базальным положением основания IR2; от K. carpenteri, K. ivensis, K. mirabilis и K. tyulkinensis – смещением sn относительно n и узелка.
Замечания. Базальная часть крыла примерно до уровня sq не сохранилась. Вероятно, крыло слегка деформировано – растянуто по ширине.
Материал. Голотип.
Kennedya madygensis Felker, sp. nov.
Табл. IX , фиг. 7
Название вида от местонахождения и урочища Мадыген.
Голотип – ПИН, № 5330/80; позитивный и негативный отпечатки почти целого крыла хорошей сохранности; местонахождение Мадыген (Джайлоучо); средний–верхний триас, ладинский–карнийский ярусы, мадыгенская свита.
Описание (рис. 3, д). N – полный, n находится прямо под ним, sn смещена в дистальном направлении. N равноудален от Asn и продолжающей ее в следующем поле поперечной жилки в проксимальном направлении, и первой пары Pn и Psn – в дистальном. Постнодальное поле немного уже постсубнодального на участке от N до Pt. Pn жилок 6, Psn – 3. Pn и Psn жилки смещены друг относительно друга. Pt небольшая, трапециевидная, передний и задний ее края параллельны друг другу, дистальный и проксимальный края имеют практически одинаковый наклон, br расположена на уровне дистальной трети Pt.
Arc средней длины, расстояние от ее начала до Asn в 1.5 раза больше расстояния от Asn до N; q открытая. Передний край q вдвое короче Arc. Субдискоидальная ячейка узкая, длинная, слегка изогнутая, ее базальный край расположен на уровне ответвления МА от Rs. Основания ветвей радиуса расположены на равном расстоянии друг от друга, дистальнее N. Основание R4 + 5 расположено в проксимальной 1/5 части расстояния между N и Pt, основание IR3 – на середине этого расстояния, основание R3 – в дистальной трети того же расстояния, основание IR2 – немного проксимальнее Pt.
А прямая, короткая, ее вершина находится на уровне N. CuP длинная, ее вершина расположена немного проксимальнее Pt. MA слегка зигзагообразная в дистальной части, ее вершина находится на уровне основания IR2. Вершины остальных продольных жилок расположены дистальнее Pt. Между дистальной стороной sq и окончанием А находится одна поперечная жилка, между вершинами А и CuP – 11 простых поперечных жилок, между вершинами МА и R4 + 5 – 3 поперечные жилки. Задний край крыла слегка расширен. Кубито-анальное поле неширокое. IR3 слегка зигзагообразная, остальные радиальные и интеркалярные жилки прямые.
Размеры в мм. Длина сохранившегося остатка – 20.6 (примерная длина реконструированного крыла – 22.0), максимальная ширина сохранившегося фрагмента (скорее всего, слегка суженного) – 2.5, узелковая ширина – 1.6. Расстояние от Arc до N – 4.3, от N до Pt – 9.9. Длина sq – 2.1, Pt – 1.8, ширина Pt – 0.2.
Сравнение. Новый вид наиболее похож на K. kedrovkensis и K. pritykinae по форме и длине А, но отличается от них меньшей шириной кубито-анального поля, базальным положением Asn и удлиненной sq. С K. ferganensis и K. gracilis сходен формой Pt, с K. azari, K. ivensis и K. mirabilis – длиной CuP и шириной заднего края крыла. От K. tyulkinensis и K. ferganensis новый вид отличается положением br и большими размерами крыла, от остальных известных видов, за исключением K. kedrovkensis – заметно меньшими. От K. carpenteri и K. suchonensis отличается меньшими размерами IR2, от остальных видов, кроме K. kedrovkensis (для которого этот признак не известен) – заметно большими. От всех видов, за исключением K. carpenteri и K. suchonensis, отличается большим числом Pn жилок, от K. carpenteri, K. ivensis, K. mirabilis и K. tyulkinensis – смещением sn относительно n и узелка.
Материал. Голотип.
ОБСУЖДЕНИЕ
Переописание вида K. carpenteri, предложенное в настоящей статье, вызвано выявлением нескольких интересных особенностей, связанных с изменением строения некоторых крыловых структур среди экземпляров типовой серии. Наиболее важные различия сосредоточены в нодальной области. На голотипе и паратипе ПИН, № 2785/6 субнодальная жилка (sn) продолжает нодальную (n) без смещения, а на паратипах ПИН, №№ 2069/37, 2555/616 и 2240/1798 sn расположена заметно дистальнее n и N, образуя так называемый “несовершенный” ступенчатый узелок.
Узелок, наряду с Arc и первичными антенодальными жилками, является одной из наиболее древних структур, известной для большинства современных и ископаемых стрекоз, за исключением меганеврид. У Ф. Фрезера он трактуется как дистальное окончание костантенодального комплекса крыла и представляет нешарнирное соединение С (косты) и Sc (субкосты) в месте перегиба последней к переднему краю крыла (Tillyard, Fraser, 1938). У всех современных и большинства ископаемых групп в состав узелка входят нодальная (n) и субнодальная (sn) поперечные жилки, обеспечивающие соединение основной структуры с радиальными жилками (R1 и R2). Сложное строение узелка, по сути представляющего собой целый комплекс расположенных в определенном порядке продольных (С; Sc) и поперечных (n; sn) жилок, предполагает невозможность его независимого образования в разных группах стрекоз.
Различное положение и соотношение n и sn, по-видимому, отражает несколько стадий формирования узелка у ископаемых стрекоз. Так, у первых карбоновых протозигоптер (Jacquesoudardia magnifica и Bechlya ericrobinsoni) и раннепермских кеннедиид перпендикулярная sn смещена относительно n. У известных исключительно из пермских отложений пермагрионид sn заметно наклонена к n и всегда продолжает ее без смещения. При этом у самых ранних кунгурских и казанских представителей родов Solikamptilon Zalessky, 1948 и Permolestes Martynov, 1932 sn имеет заметно большую длину и наклон, чем у уржумского Epilestes Martynov, 1937 и вучапинского Permagrion Tillyard, 1928 (Tillyard, 1928; Nel et al., 2012). Вероятно, это объясняется постепенным преобразованием sn из простой постнодальной поперечной жилки в часть узелковой структуры и “подтягиванием” ее к уровню N (Tillyard, Fraser, 1938). У большинства мезо-кайнозойских и современных стрекоз укороченные sn и n расположены без смещения и входят в состав N. Среди них выделяется группа Archizygoptera, у представителей которой наблюдается довольно примитивное ступенчатое (sn смещена относительно n) и даже неполное (n и sn отсутствуют) строение N. Наиболее многочисленные и разнообразные архизигоптеры – протомирмелеонтиды – характеризуются сильной редукцией продольного жилкования при заметном увеличении поперечного. Их узелок представлен только основной структурой продольных жилок (завершение С и перегиб Sc), не несущей n и sn. По-видимому, именно отсутствие настоящего полного узелка является основным фактором заметного усиления поперечного жилкования (An и Pn) переднего края крыла. У остальных архизигоптер, в состав которых сейчас включают и семейство Kennedyidae (Nel et al., 2012), наблюдаются разные типы строения узелка: как с выравненными (“настоящий” узелок), так и со смещенными n и sn (ступенчатый, “несовершенный” узелок). Так, первый тип представлен у ранне- и среднепермских видов Kennedya azari Nel et al., 2012, K. ivensis Nel et al., 2012, K. tyulkinensis Felker, 2020, Progoneura kityakensis Felker, 2020, а также двух экземпляров типовой серии триасового K. carpenteri (Притыкина, 1981; Nel et al., 2012; Felker, 2020). Ступенчатый узелок, со смещенной sn, присутствует у остальных кеннедиид и большинства известных архизигоптер (сем. Voltzialestidae, Luiseiidae и др.). Притыкина (1981) отмечает, что этот признак (полное строение узелка) носит “приспособительный” некоррелируемый характер в разных группах стрекоз. То есть, у протозигоптер, мезо-кайнозойских и современных стрекоз этот признак сохраняется на довольно высоких уровнях, вплоть до надсемейства (Tillyard, Fraser, 1938; Nel et al., 2012). В то же время, у архизигоптер наблюдаются разные по степени сформированности узелки (ступенчатый и совершенный) даже в пределах одного вида. По-видимому, это связано с тем, что их узелок имеет довольно примитивное строение, при котором sn еще не входит в его состав и, по сути, является обычной постсубнодальной поперечной жилкой (у Protomyrmeleontidae, соответственно, обе жилки – n и sn – не являются узелковыми структурами). Это вполне подтверждает предположение об эволюции узелка от ступенчатого, характерного для карбоновых, ранне- и среднепермских архизигоптер через узелок пермских пермагрионид, с “подтягиваемой” sn, к совершенному – у современных стрекоз. Присутствие несовершенного узелка, со смещенной или, по-видимому, отсутствующей sn, у более молодых позднепермских и триасовых архизигоптер (в том числе, кеннедиид), скорее всего, связано с уменьшением размеров крыла и коррелятивным преобразованием его жилкования (Притыкина, 1981). Уникальный по строению узелок мезозойских Protomyrmeleontidae, в котором отсутствуют n и sn, видимо, также связан с усиленной специализацией жилкования в сторону уменьшения размеров и компактизации формы крыла.
Список литературы
Добрускина И.А. Триасовые флоры Евразии. М.: Наука, 1982. 182 с.
Копылов Д.С. Новые пилильщики подсемейства Madygellinae (Hymenoptera, Xyelidae) из среднего-верхнего триаса Kиргизии / Палеонтол. журн. 2014. № 6. С. 37–45. https://doi.org/10.7868/S0031031X14060087
Притыкина Л.Н. Новые триасовые стрекозы Средней Азии // Новые ископаемые насекомые с территории СССР / Ред. Вишнякова В.Н., Длусский Г.М., Притыкина Л.Н. М.: Наука, 1981. С. 5–42.
Сикстель Т.А. Стратиграфия континентальных отложений верхней перми и триаса Средней Азии. Ташкент: Изд-во СамГУ, 1960. 147 с. (Тр. Ташкентского гос. ун-та. Т. 176. Кн. 3).
Фелькер А.С. Первые стрекозы семейства Kennedyidae (Odonata: Protozygoptera) из пограничных пермо-триасовых отложений Кузнецкого бассейна России // Палеонтол. журн. 2021а. № 2. С. 52–59. https://doi.org/10.31857/S0031031X21020057
Фелькер А.С. Новые стрекозы семейства Kennedyidae (Odonata: Protozygoptera) из верхнепермских отложений Вологодской области // Палеонтол. журн. 2021б. № 4. С. 41–49. https://doi.org/10.31857/S0031031X21040061
Фелькер А.С., Василенко Д.В. Древнейшие стрекозы семейства Triadophlebiidae из нового местонахождения Петропавловка (граница нижнего и среднего триаса Приуралья) // Палеострат-2019. Годичн. собр. (науч. конф.) секции палеонтологии МОИП и Моск. отд. Палеонтол. об-ва при РАН. Прогр. и тез. докл. / Ред. Алексеев А.С., Назарова В.М. М.: ПИН РАН, 2019. С. 66.
Carpenter F.M. The Lower Permian insects of Kansas. Part 2. The orders Paleodictyoptera, Protodonta and Odonata // Amer. J. Sci. Ser. 5. 1931. V. 21. P. 97–139. https://doi.org/10.2475/ajs.s5-21.122.97
Carpenter F.M. The Lower Permian insects of Kansas. Part 6: Delopteridae, Protelytroptera, Plectoptera and a new collection of Protodonata, Odonata, Megasecoptera, Homoptera and Psocoptera // Proc. Amer. Acad. Arts & Sci. 1933. V. 68. № 11. P. 411–505. https://doi.org/10.2307/20022959
Carpenter F.M. The Lower Permian insects of Kansas. Part 8. Additional Megasecoptera, Protodonata, Odonata, Homoptera, Psocoptera, Protelytroptera, Plectoptera, and Protoperlaria // Proc. Amer. Acad. Arts & Sci. 1939. V. 73. № 3. P. 29–70. https://doi.org/10.2307/25130151
Carpenter F.M. Lower Permian insects from Oklahoma. Part 1. Introduction and the orders Megasecoptera, Protodonata, and Odonata // Proc. Amer. Acad. Arts & Sci. 1947. V. 76. № 2. P. 25–54. https://doi.org/10.2307/20023497
Dobruskina I.A. Keuper (Triassic) flora from Middle Asia (Madygen, southern Fergana) // New Mexico Mus. Natur. Hist. Sci. Bull. 1995. V. 5. P. 1–49.
Felker A.S. New damselflies of the family Kennedyidae from Permian deposits of European Russia // Paleontol. J. 2020. V. 54. № 7. P. 734–742. https://doi.org/10.1134/S0031030120070047
Nel A., Bethoux O., Bechly G. et al. The Permo-Triassic Odonatoptera of the “Protodonate” grade (Insecta: Odonatoptera) // Ann. Soc. Entomol. Fr. (N.S.). 2001. V. 37. № 4. P. 501–525.
Nel A., Bechly G., Prokop J. et al. Systematics and evolution of Paleozoic and Mesozoic damselfly like Odonatoptera of the Protozygopteran grade // J. Paleontol. 2012. V. 86. № 1. P. 81–104. https://doi.org/10.1666/11-020.1
Shcherbakov D.E. Madygen, Triassic Lagerstätte number one, before and after Sharov // Alavesia. 2008. V. 2. P. 113–124.
Shcherbakov D.E., Timm T., Tzetlin A.B. A probable oligochaete from an Early Triassic Lagerstätte of the southern Cis-Urals and its evolutionary implications // Acta Palaeontol. Pol. 2020. V. 65. № 2. P. 219–233. https://doi.org/10.4202/app.00704.2019
Tillyard R.J. Kansas Permian insects. Part 5. The orders Protodonata and Odonata // Amer. J. Sci. Ser. 5. 1925. V. 10. P. 41–73. https://doi.org/10.2475/ajs.s5-10.55.41
Tillyard R.J. A Permian fossil damselfly wing from the Falkland Islands // Trans. R. Entomol. Soc. Lond. 1928. V. 76. № 1. P. 55–63. https://doi.org/10.1111/j.1365-2311.1928.tb01187.x
Tillyard R.J., Fraser F.C. A reclassification of the order Odonata based on some new interpretations of the venation of the dragonfly wing // Austral. Zool. 1938. V. 9. P. 125–169.
Voigt S., Buchwitz M., Fischer J. et al. Triassic life in an inland lake basin of the warm-temperate biome – the Madygen Lagerstätte (Southwest Kyrgyzstan, Central Asia) // Terrestrial Conservation Lagerstätten: Windows into the Evolution of Life on Land / Eds. Fraser N.C., Sues H.-D. Edinburgh: Dunedin Acad. Press, 2017. P. 65–104.
Zheng D.R., Nel A., Wang B. et al. The first Triassic ‘Protodonatan’ (Zygophlebiidae) from China: stratigraphical implications // Geol. Mag. 2017a. V. 154. № 1. P. 169–174. https://doi.org/10.1017/S0016756816000625
Zheng D., Nel A., Wang H. et al. The first Late Triassic Chinese triadophlebiomorphan (Insecta: Odonatoptera): biogeographic implications // Sci. Rep. 2017b. V. 7. № 1. P. 1–7. https://doi.org/10.1038/s41598-017-01710-7
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал