

Палеонтологический журнал, 2023, № 4, стр. 26-36
Новые наутилиды из ассельско-сакмарского рифового комплекса Шахтау (Башкортостан)
Т. Б. Леонова a, *, А. Ю. Щедухин a, **
a Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
* E-mail: tleon@paleo.ru
** E-mail: d_alsch2017@mail.ru
Поступила в редакцию 06.02.2023
После доработки 13.02.2023
Принята к публикации 13.02.2023
- EDN: OHDXXG
- DOI: 10.31857/S0031031X23040074
Аннотация
В статье описаны новые наутилиды из ассельско-сакмарского рифового комплекса Шахтау. Обсуждается их место в системе наутилид, пока еще очень далекой от совершенства. Два вида двух родов: Omorphoceras igori gen. et sp. nov. и Stenopoceras (Leptodomatoceras) bashkiricum subgen. et sp. nov. отнесены к подсемейству Domatoceratinae Miller et Youngquist семейства Grypoceratidae Hyatt. Третий вид, Foveroceras magnum gen. et sp. nov. со сложной перегородочной линией, отнесен к новому роду. Он включен в состав экзотического семейства Permoceratidae Miller et Collinson, которое ранее считалось монотипическим и было известно только из ранней перми о-ва Тимор. Рассмотрена классификация семейства Grypoceratidae. Поддержана гипотеза о родственных связях между семействами Grypoceratidae и Permoceratidae.
ВВЕДЕНИЕ
Во время полевых работ на карьере Шахтау (Башкортостан), существующем в настоящее время на месте бывшего шихана Шахтау, в рифовых известняках позднеассельского–сакмарского возраста совместно с многочисленными находками самых разнообразных наутилид (Leonova, Shchedukhin, 2019, 2020; Shchedukhin, Leonova, 2020; Щедухин, 2022а, б) была собрана коллекция крупных раковин необычной формы. Эти раковины характеризуются высокими оборотами, широкими плоскими боковыми сторонами и довольно сложной перегородочной линией. Обломки раковин такого типа были обнаружены еще во время первых экспедиций А.В. Мазаева на Шахтау (2015–2017 гг.) и сразу обратили на себя внимание, но сохранность первых находок была столь неудовлетворительной, что мы не сочли возможным описать материал. В последние годы, благодаря новым сборам сотрудников лаборатории моллюсков Палеонтологического ин-та им. А.А. Борисяка РАН (ПИН РАН), коллекция позволяет с достаточной степенью детальности охарактеризовать новые формы и найти их место в системе наутилид.
Основной трудностью при изучении наутилид (особенно палеозойских) является крайний дефицит фундаментальных исследований этой группы. В отличие от аммоноидей, изучением которых занимались сотни палеонтологов, исследования наутилоидей, разработку принципов их систематики, анализ особенностей их морфологии и филогении проводило не так много ученых. И сейчас основным источником информации по системе палеозойских наутилид остаются работы, написанные в середине прошлого века и даже ранее (Круглов, 1928; Miller et al., 1933, 1949; Flower, 1952; Руженцев, Шиманский, 1954; Шиманский, 1962, 1967; Kummel, 1964 и некоторые другие). Самыми современными можно назвать работы В.Н. Шиманского (1979) и Е.С. Соболева (1989) и несколько работ зарубежных авторов (Niko, Mapes, 2015, 2016; King, Evans, 2019). А.А. Шевырев (2006) очень детально разобрал историю построения основных вариантов системы наутилоидей, предложенных разными авторами в течение ХХ в., их достоинства и недостатки. До сих пор нет согласия в определении четких критериев для создания стройной системы подкласса Nautiloidea, которая была бы принята большинством специалистов. Недаром И.С. Барсков (2018) назвал “хаосом” состояние системы высших таксонов головоногих, в которой в разные годы было выделено больше подклассов, чем отрядов. В своей работе мы постарались не увеличивать число таксонов семейственного ранга, а найти место впервые найденным и весьма необычным представителям этой группы в системе отряда Nautilida Agassiz, 1847, разработанной Шиманским (1967, 1979). Эта система представляется нам наиболее приемлемой из всех предложенных до настоящего времени. Близкими к ней по основным принципам являются более ранняя система Шиманского (1962), опубликованная в “Основах палеонтологии”, а также система Б. Каммела (Kummel, 1964), опубликованная в американской сводке “Treatise on Invertebrate Paleontology”. По свидетельству самого Шиманского (1979), различия систем, предложенных им и Каммелом, состоят, главным образом, в выделении таксонов высокого ранга: подотрядов и надсемейств, а на уровне семейств и, особенно, родов они становятся незначительными. Как указывает сам Шиманский (1979, с. 7): “различие в их [родов] числе у этих двух исследователей не превышает 10% от их общего числа”.
РЕЗУЛЬТАТЫ
В нашей коллекции в настоящее время имеются 19 экз. очень крупных наутилид, резко отличающихся по форме раковины от других представителей этого отряда из одновозрастных отложений. На первом этапе изучения, имея только несколько фрагментов оборотов без лопастных линий, мы вместе с Барсковым полагали, что, возможно, эти экзотические формы относятся к неизвестной группе семейственного ранга. Во время полевых работ 2019–2022 гг. одному из авторов статьи, А.Ю. Щедухину, удалось найти дополнительно несколько лучше сохраннившихся экземпляров с перегородочной линией. Самый полный экземпляр, описанный здесь как голотип типового вида нового рода Omorphoceras, был найден и подарен генеральным директором АО “Сырьевая компания” А.В. Шкурко. Детально изучив весь материал, мы пришли к выводу, что этих наутилид можно отнести к двум родственным семействам надсемейства Centroceratoidea Hyatt, 1900: сем. Grypoceratidae Hyatt, 1900 и Permoceratidae Miller et Collinson, 1953. Основанием для такого заключения явились характерные комплексы основных признаков: форма раковины, скульптура (точнее, ее отсутствие), строение и форма элементов перегородочной линии, высота газово-жидкостных камер, положение сифона. К сожалению, сохранность имеющегося материала не позволила изучить эмбриональную раковину новых таксонов.
Изученная коллекция, зарегистрированная под № 5668, хранится в лаб. моллюсков ПИН РАН.
ПОДОТРЯД CENTROCERATINA
НАДСЕМЕЙСТВО CENTROCERATOIDEA HYATT, 1900
СЕМЕЙСТВО GRYPOCERATIDAE HYATT, 1900
ПОДСЕМЕЙСТВО DOMATOCERATINAE MILLER ET YOUNGQUIST, 1949
Род Omorphoceras Leonova et Shchedukhin, gen. nov.
Название рода от όμορφο κέρατο (omorpho kerato) греч.– красивый рог.
Типовой вид – Omorphoceras igori sp. nov. из ассельско–сакмарских известняков карьера Шахтау, Башкортостан.
Диагноз. Раковина дискоконовая, полуинволютная, с высоким угловато-овальным сечением оборота, гладкая. Вентральная сторона узкая, уплощенная, в центральной части слегка вогнутая, с резкими вентральными краями, образующими кили. Боковые стороны широкие, слабовыпуклые, почти плоские. Умбиликальный край округленный. Умбилик средних размеров, ступенчатый. Сифон занимает субцентральное положение. Камеры низкие, на высоту оборота их приходится пять.
Поверхность раковины практически гладкая. На ядре также не наблюдается никаких скульптурных элементов.
Перегородочная линия с неширокой и глубокой вентральной лопастью, узким высоким наружным седлом, расположенным на вентральном крае. Латеральная лопасть очень широкая, занимает всю боковую сторону, и очень глубокая. На умбиликальном перегибе расположено невысокое умбиликальное седло, на дорсальной стороне – неглубокая дорсальная лопасть, возможно, с аннулярным отростком.
Видовой состав. Типовой вид.
Сравнение. От наиболее близкого рода Domatoceras Hyatt, 1891 отличается формой поперечного сечения со значительно большей высотой и меньшей шириной оборота, а также сильно уплощенными боковыми сторонами и слегка вогнутой очень узкой вентральной стороной. Обороты не прилегающие, а объемлющие. Все элементы перегородочной линии выражены гораздо контрастнее, чем у Domatoceras: вентральная лопасть более узкая и гораздо более глубокая, наружное седло более высокое и латеральная лопасть значительно глубже. От другого близкого рода, Stenopoceras Hyatt, 1893, отличается формой сечения оборота с более плоскими и почти параллельными боковыми сторонами, значительно более широким умбиликом (Ду/Д 0.33–0.34 против 0.04–0.07), полуинволютной, а не инволютной раковиной и наличием глубокой вентральной лопасти в перегородочной линии.
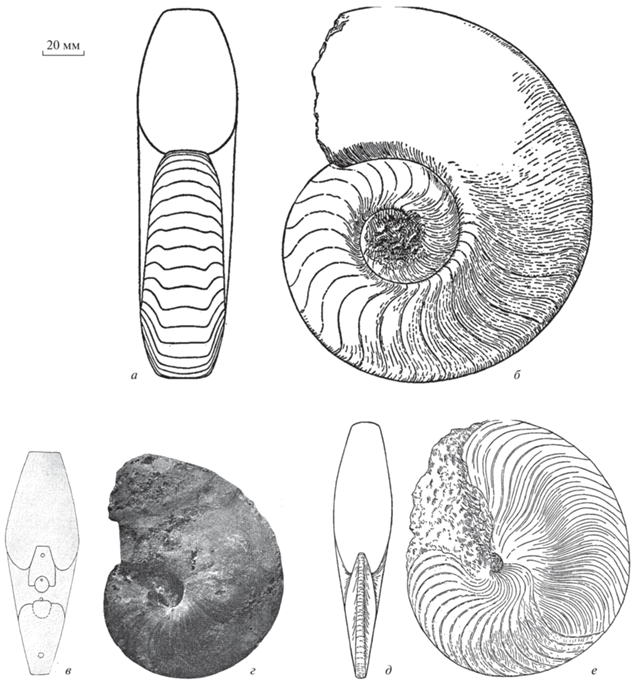
Замечания. Недавно Ш. Нико и Р. Мэйпс (Nico, Mapes, 2016) описали нового позднекаменноугольного представителя доматоцератин из штата Оклахома (США) под видовым названием Domatoceras collinsvillense Nico et Mapes, 2016. Этот вид по форме раковины напоминает описываемый нами род, но отличается от него перегородочной линией с неглубокой вентральной лопастью и менее глубокой латеральной лопастью, а также менее широким умбиликом. Включение этого вида в состав рода Domatoceras вызывает большие сомнения, поскольку его раковина имеет в значительной мере объемлющие обороты, а для Domatoceras характерны практически не объемлющие, а прилегающие обороты (рис. 1, а, б). К сожалению, описание вида дано авторами не в полном объеме, также отсутствует рисунок перегородочной линии. Не исключено, что дальнейшие исследования позволят отнести вид D. collinsvillense к роду Omorphoceras.
Рис. 1.
Семейство Grypoceratidae, подсемейство Domatoceratinae, типовые виды: а, б – рода Domatoceras (D. umbilicatum Hyatt); в, г – подрода Domatoceras (Stenodomatoceras) – [D. (S.) moorei (Miller, Dunbar et Condra]; д, е – рода Stenopoceras [S. dumbli (Hyatt)]. Во всех случаях ×0.5 [по: Miller, Youngquist, 1949 (а–г) и Miller et al., 1933 (д, е)].
Omorphoceras igori Leonova et Shchedukhin, sp. nov.
Табл. V , фиг. 3 (см. вклейку)
Название вида – в честь проф. Игоря Сергеевича Барскова (1937–2019).
Голотип – ПИН, № 5668/211; Южный Урал, Башкортостан, карьер Шахтау; нижняя пермь, пограничные ассельско-сакмарские рифовые известняки.
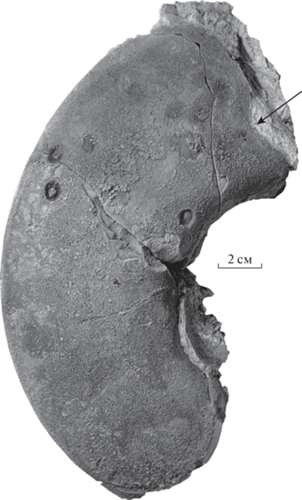
Описание (рис. 2; 3, а). Раковина крупная, дисковидной формы, с высокими и узкими оборотами угловато-овального сечения, умеренно инволютная. Высота оборота превышает его ширину в 1.5 раза у взрослых экземпляров. Вентральная сторона узкая (в 2.5 раза уже соответствующей ширины оборота), уплощенная, на взрослых оборотах в центральной части слегка вогнутая с резкими вентральными краями, образующими кили. Боковые стороны очень широкие, слабовыпуклые, почти плоские. Умбиликальный край округленный. Умбилик средних размеров (Ду/Д = 0.33–0.34), неглубокий, ступенчатый. Умбиликальное отверстие около 4 мм. На одном из образцов видно очертание устья (рис. 2), на вентральной стороне оно образует широкий и глубокий синус, на внешней части боковой стороны – плавный изгиб вперед, а в приумбиликальной зоне – небольшой синус; возможно, в этой месте помещался глаз животного. Сифон занимает субцентральное положение. Камеры низкие, на высоту оборота их приходится 5–5.5.
Рис. 2.
Omorphoceras igori gen. et sp. nov., экз. ПИН, № 5668/213, фрагмент жилой камеры с остатками устья; Башкортостан, карьер Шахтау; пограничные ассельско-сакмарские рифовые известняки.
На ядре не наблюдается никаких скульптурных элементов. Поверхность раковины гладкая, за исключением очень тонких линий нарастания. Последние едва заметны на сохранившихся участках раковины и образуют глубокий и широкий синус на вентральной стороне.
Перегородочная линия (рис. 3, а) с неширокой и глубокой вентральной лопастью (ее глубина немного превышает высоту камеры), узким высоким наружным седлом, приостренная вершина которого расположена на вентральном крае. Латеральная лопасть очень широкая, более или менее симметричная, с округлым основанием; занимает всю боковую сторону, глубокая – вдвое превышает высоту камеры. На умбиликальном перегибе и стенке расположено невысокое и довольно широкое умбиликальное седло, на дорсальной стороне – неглубокая дорсальная лопасть, возможно, с аннулярным отростком.
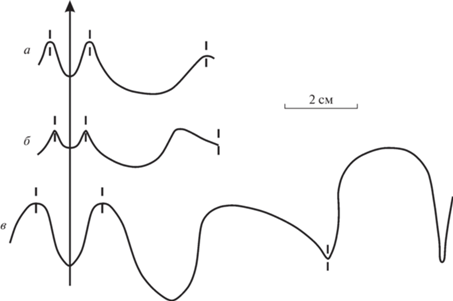
Рис. 3.
Перегородочные линии: а – Omorphoceras igori gen. et sp. nov., голотип ПИН, № 5668/211; б – Stenopoceras (Leptodomatoceras) bashkiricum sp. nov., голотип ПИН, № 5668/117; в – Foveroceras magnum gen. et sp. nov., голотип ПИН, № 5668/200; Башкортостан, карьер Шахтау; пограничные ассельско-сакмарские рифовые известняки.
Размеры в мм и отношения
Материал. Семь экз., два хорошей и пять плохой сохранности из пограничных ассельско–сакмарских рифовых известняков карьера Шахтау, Башкортостан.
Род Stenopoceras Hyatt, 1893
Подрод Leptodomatoceras Leonova et Shchedukhin, subgen. nov.
Название подрода от λεπτός (leptós) греч. – стройный, и рода Domatoceras.
Типовой вид – Stenopoceras (Leptodomatoceras) bashkiricum sp. nov.; карьер Шахтау, Башкортостан, Южный Урал; пограничные ассельско-сакмарские рифовые известняки, нижняя пермь.
Диагноз. Дискоконовая инволютная раковина средних размеров с высоко-трапециевидным сечением оборота и очень узким умбиликом. На средних и поздних стадиях вентральная сторона плоская, неширокая. Боковые стороны широкие, уплощенные, на взрослых стадиях отделены от вентральной резким перегибом. Умбилик очень узкий, глубокий, с выраженным умбиликальным перегибом. Скульптура отсутствует, за исключением очень тонких струек роста, которые образуют на боковой стороне плавный изгиб вперед, а на вентральной – синус. Сифон занимает субцентральное положение. Камеры средней высоты, на высоту оборота их приходится четыре. Перегородочная линия с глубокой и умеренно широкой вентральной лопастью, занимающей всю вентральную сторону, и узким вентро-латеральным седлом, расположенным на вентральном перегибе. На боковой стороне находится глубокая и широкая латеральная лопасть, немного асимметричная, с округленным основанием; в умбиликальной зоне расположено высокое и широкое латерально-умбиликальное седло округло-треугольной формы. Центр широкой умбиликальной лопасти фиксируется на умбиликальном шве.
Видовой состав. Типовой вид.
Сравнение. От номинативного подрода Stenopoceras (Stenopoceras) отличается очертаниями поперечного сечения оборота: более широкой вентральной стороной (отношение ширины оборота к ширине вентральной стороны 2–2.5 против 4.5–6) и более параллельными (менее расходящимися к умбилику) боковыми сторонами. Перегородочная линия нового подрода отличается наличием хорошо сформированной вентральной лопасти вместо уплощенной вершины вентрального седла.
Stenopoceras (Leptodomatoceras) bashkiricum Leonova et Shchedukhin, sp. nov.
Табл. V , фиг. 1, 2
Название вида – от Башкирии.
Голотип – ПИН, № 5668/217; Южный Урал, Башкортостан, карьер Шахтау; нижняя пермь, пограничные ассельско-сакмарские рифовые известняки.
Описание Раковина средних размеров, дисковидная, инволютная с высоким трапециевидным сечением оборота. Угловатость наблюдается за счет хорошо выраженных вентральных перегибов, ограничивающих узкую плоскую вентральную сторону. Ширина вентральной стороны в 2–2.5 раза меньше соответствующей ширины оборота. Боковые стенки очень широкие, почти плоские, под небольшим углом (12–15 градусов) расходятся от вентрального перегиба к зоне умбилика, где раковина имеет максимальную ширину. Умбилик очень узкий, с крутыми умбиликальными стенками. Умбиликального отверстия наблюдать не удалось. Сифон занимает субцентральное положение. Камеры средней высоты, на высоту оборота их приходится четыре.
Поверхность раковины покрыта едва различимыми тонкими струйками роста, которые образуют на боковой стороне широкий изгиб вперед, а на вентральной – глубокий синус.
Перегородочная линия (рис. 3, б). Вентральная лопасть глубокая, с округленным основанием, занимает всю вентральную сторону, ограничена двумя узкими наружными седлами, приостренные вершины которых расположены на вентральных перегибах. Латеральная лопасть очень глубокая и широкая, немного асимметричная, занимает больше половины боковой стороны. За ней следует высокое и широкое седло, которое в умбиликальной зоне переходит в широкую умбиликальную лопасть, центр которой расположен на умбиликальном шве. Дорсальную часть перегородочной линии изучить не удалось.
Размеры в мм и отношения
Сравнение. В подроде Leptodomatoceras один вид.
Материал. Два экз. удовлетворительной сохранности из пограничных ассельско-сакмарских рифовых известняков карьера Шахтау, Башкортостан.
СЕМЕЙСТВО PERMOCERATIDAE MILLER ET COLLINSON, 1953
Foveroceras Leonova et Shchedukhin, gen. nov.
Название рода от φοβερο κέρατο (fovero kerato) греч. – изумительный рог.
Типовой вид – Foveroceras magnum sp. nov.; карьер Шахтау, Башкортостан, Южный Урал; пограничные ассельско-сакмарские рифовые известняки, нижняя пермь.
Диагноз. Очень крупная инволютная раковина с параболическим сечением высокого оборота (рис. 4). На ранних и средних стадиях вентральная сторона округленная, без выраженных перегибов, затем она уплощается и на третьем обороте становится совсем плоской, а к концу его – даже немного вогнутой в центральной части. Боковые стороны широкие, уплощенные, на взрослых стадиях отделены от вентральной резким перегибом. Умбилик очень узкий, глубокий, воронкообразный, без выраженного умбиликального перегиба. Сифон занимает субцентральное положение. Камеры средней высоты, в три раза меньше высоты оборота. Скульптура отсутствует, раковина гладкая. Перегородочная линия с очень глубокой и узкой вентральной лопастью, высоким приостренным наружным седлом, расположенным на вентральном перегибе, и очень глубокой асимметричной латеральной лопастью с приостренным основанием, занимающей половину боковой стороны; на приумбиликальной части боковой стороны расположено резко асимметричное высокое и широкое седло. Умбиликальная лопасть асимметричная, узкая, на внутренней стороне оборота находится еще одно очень высокое и широкое седло с округленной вершиной. Дорсальная лопасть очень узкая и глубокая, с аннулярным отростком.
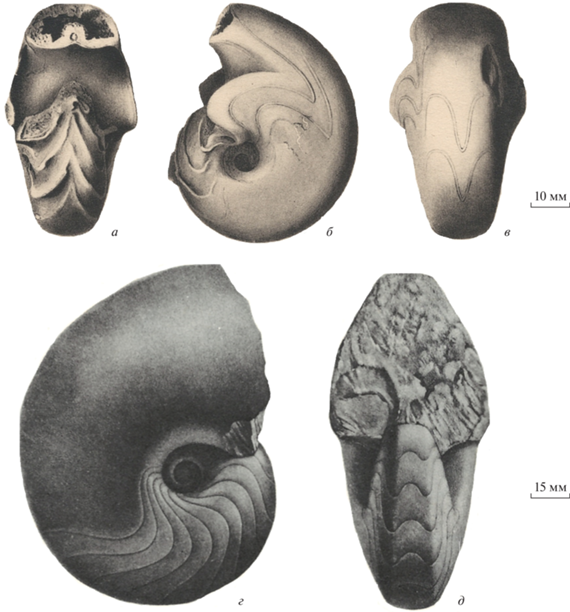
Рис. 4.
Foveroceras magnum gen. et sp. nov., экз. ПИН, № 5668/202, вид с боковой стороны; Башкортостан, карьер Шахтау; пограничные ассельско-сакмарские рифовые известняки.

Видовой состав. Типовой вид.
Сравнение. От типового рода этого семейства – Permoceras Miller et Collinson (Haniel, 1915, табл. LV (10), фиг. 3) – отличается более узкой раковиной (Ш/Д 0.37–0.41 против 0.57–0.63) с плоской вентральной стороной и угловатыми вентро-латеральными перегибами на взрослых оборотах. К. Ганиэль (Haniel, 1915) при описании вида, впоследствии принятого как типовой для рода Permoceras, указывает, что сначала ширина превышает высоту оборота, потом они становятся равными или даже высота превышает ширину, соотношение Ш/В = 0.94–1.02, а у уральской формы на всех стадиях высота больше ширины – 0.63–0.72. Также отличается значительно более узким умбиликом: Ду/Д 0.05–0.07 против 0.16–0.22. Перегородочная линия нового рода отличается более широкой вентральной, более симметричной, а не клювовидной латеральной и более глубокой дорсальной лопастями. Основания всех лопастей нового рода (кроме дорсальной) имеют менее заостренную форму.
Foveroceras magnum Leonova et Shchedukhin, sp. nov.
Табл. VI , фиг. 1, 2 (см. вклейку)
Название вида от magnus лат. – крупный.
Голотип – ПИН, № 5668/200; Южный Урал, Башкортостан, карьер Шахтау; нижняя пермь, пограничные ассельско-сакмарские рифовые известняки.
Описание (рис. 4). Взрослая раковина состоит из трех оборотов, очень крупная, диаметром до 200 мм и более, инволютная, с высоким параболическим сечением оборота. На первом и половине второго оборота вентральная сторона округленная, затем она уплощается и на третьем обороте становится совершенно плоской, а к концу его – даже немного вогнутой. Боковые стороны сходятся к вентральной стороне, они широкие, слегка выпуклые, почти плоские, на взрослых стадиях отделены от вентральной резким перегибом. Умбилик очень узкий, глубокий, воронкообразный, без выраженного умбиликального перегиба. Сифон занимает субцентральное положение. Камеры средней высоты, на высоту оборота их приходится три. Скульптуру наблюдать не удалось, раковина гладкая.
Перегородочная линия (рис. 3, в) с очень глубокой и довольно узкой вентральной лопастью, высоким приостренным наружным седлом, расположенным на вентральном перегибе, и очень глубокой (2.5 камеры) асимметричной латеральной лопастью с приостренным основанием, занимающей половину боковой стороны. На приумбиликальной части боковой стороны расположено высокое и широкое, резко асимметричное седло. Умбиликальная лопасть узкая, клювовидная. На внутренней стороне оборота находится дорсо-умбиликальное седло, очень высокое и широкое, с округленной вершиной. Дорсальная лопасть очень узкая и глубокая, с длинным аннулярным отростком.
Размеры в мм и отношения
| Экз. ПИН № | Д | В | Ш | Ду | В/Д | Ш/Д | Ду/Д | Ш/В |
|---|---|---|---|---|---|---|---|---|
| Голотип 5668/200 | 190* | 108 | 78 | 10 | 0.57 | 0.41 | 0.05 | 0.72 |
| 104 | 69 | 0.66 | ||||||
| 5668/203 | 132 | 76 | 48 | 8 | 0.58 | 0.36 | 0.06 | 0.63 |
| 5668/202 | 170* | 100* | 64* | 12 | 0.59 | 0.38 | 0.07 | 0.64? |
| 5668/201(дорс) | 59 |
Сравнение. В роде Foveroceras один вид.
Материал. 10 экз., представленных крупными фрагментами раковины средней и плохой сохранности из пограничных ассельско-сакмарских рифовых известняков карьера Шахтау, Башкортостан.
ОБСУЖДЕНИЕ
При изучении описанного материала самым сложным был вопрос о систематическом положении найденных форм. По основным параметрам раковины и строению перегородочной линии они ближе всего к представителям семейства Grypoceratidae Hyatt, 1900, подсемейства Domatoceratinae Miller et Youngquist, 1949. История этого таксона довольно сложная, состав его неоднократно менялся. Американские палеонтологи (Miller, Youngquist, 1949) выделили семейство Domatoceratidae. Руженцев и Шиманский (1954) в своей работе приняли Domatoceratidae в этом качестве. Впоследствии ранг семейства был понижен до подсемейства, оно вошло в состав семейства Grypoceratidae. Палеозойские формы были объединены в подсемейство Domatoceratinae, а триасовые роды – в подсемейство Grypoceratinae Hyatt, 1900. В отечественной сводке “Основы палеонтологии” (Шиманский, 1962, с. 133) опубликована схема с делением грипоцератид на четыре подсемейства, два из них названы выше; кроме них, сюда же были отнесены Syringonautilinae Mojsisovics, 1902 и Clymenonautilinae Shimansky, 1962. Уже в следующей монографии Шиманский (1967, с. 46) отказался от обособления Grypoceratinae от Domatoceratinae, все роды этих двух подсемейств он рассматривал в составе единого семейства грипоцератид. Два других подсемейства были объединены в семейство Syringonautilidae. В “Treatise …” (Kummel, 1964) семейство Grypoceratidae также рассматривалось без деления на подсемейства, в него включались 13 родов с очень разнообразной морфологией. По нашему мнению, деление семейства Grypoceratidae Hyatt, 1900 на два подсемейства было вполне оправданным. В своей работе мы возвращаемся к такой классификации этой группы. В качестве подсемейства грипоцератин обособляются два–три триасовых рода со своеобразной морфологией (инволютные раковины с трапециевидной или даже треугольной формой поперечного сечения). Во второе подсемейство Domatoceratinae попадают более многочисленные и широко распространенные по всему миру каменноугольно–пермские роды. Это, главным образом, формы с полуэволютной или эволютной раковиной с субпрямоугольным или субквадратным сечением, с четырех- или шестилопастной перегородочной линией.
Два из трех новых видов, описанных выше, несомненно, должны войти в состав подсемейства Domatoceratinae. Один из них относится к новому роду и описан как Omorphoceras igori gen. et sp. nov. Вторая форма обладает морфологическими признаками, часть которых характерна для Stenopoceras Hyatt, а другая – для Stenodomatoceras Ruzhencev et Shimansky. В отношении таксономичеcкой принадлежности родов Stenopoceras и Stenodomatoceras cуществует полная неясность. Руженцев и Шиманский (1954, с. 94) выделили Stenodomatoceras в качестве самостоятельного рода с типовым видом Domatoceras moorei Miller, Dunbar et Condra, 1933 в семействе Domatoceratidae (в таком ранге они тогда принимали этот таксон). Уже в “Основах палеонтологии” (Шиманский, 1962) Stenodomatoceras рассматривался в качестве подрода рода Stenopoceras, а немного позднее (Шиманский, 1967) этот же род был введен снова в качестве подрода, но уже в состав рода Domatoceras. В “Treatise…” (Kummel, 1964) Stenodomatoceras обозначен как синоним рода Domatoceras. По-видимому, такая нестабильность в понимании положения Stenodomatoceras связана с тем, что его типовой вид D. moorei, по определению авторов, занимает промежуточное положение между двумя родами, Domatoceras и Stenopoceras, поскольку оба рода “не описаны Хайеттом адекватно” (Miller et al., 1933, с. 222). Все три рода определенно генетически связаны, что следует из наличия общих морфологических черт (рис. 1), находки их типовых видов известны из пенсильвания и нижней перми Канзаса. Вместе с тем, можно наблюдать много отличительных признаков, как в форме раковины (разная степень инволютности: у Domatoceras обороты почти не объемлют друг друга, раковины двух других родов инволютны), так и в строении перегородочной линии (у Domatoceras и Stenodomatoceras имеется вентральная лопасть, а у Stenopoceras – вентральное седло). Существенных различий в строении эмбриональной раковины не найдено. По свидетельству американских исследователей, и у Domatoceras (типовой вид D. umbilicatum Hyatt, пенсильваний Канзаса), и у Stenopoceras [типовой вид Stenopoceras dumbli (Hyatt), нижняя пермь Канзаса], а также у Stenodomatoceras (типовой вид Domatoceras moorei, пенсильваний Канзаса) поперечное сечение первого оборота округлое (Miller et al., 1933). Отличаются эмбриональные раковины этих таксонов размерами первого оборота и умбиликального отверстия: у Domatoceras они крупнее, чем у Stenopoceras (Miller et al., 1933). К сожалению, сохранность изученного материала не позволила нам исследовать эмбриональные раковины. После скрупулезного анализа всех имеющихся данных мы приняли решение включить новый вид в состав рода Stenopoceras в ранге самостоятельного подрода.
Третий род по форме раковины близок к грипоцератинам, особенно к триасовому Grypoceras Hyatt, 1884, но резко отличается от них более сложной формой перегородочной линии. Этот признак сближает башкирскую форму с монотипическим раннепермским семейством Permoceratidae Miller et Collinson, 1953 [Permoceras bitauniense (Haniel) из слоев Битауни о-ва Тимор], по предположению Шиманского (1967), связанным своим происхождением с грипоцератидами. Такая трактовка представляется вполне обоснованной, т.к. те и другие близки по форме раковины (рис. 5). Что касается перегородочной линии, то, кроме Permoceras, похожая линия наблюдается у некоторых представителей позднетриасового рода Proclydonautilus Moisisovics, 1902 (подотряд Liroceratina, надсемейство Clydonautiloidea Hyatt, 1900, семействo Clydonautilidae Hyatt, 1900). По мнению Соболева (1989), детально изучавшего представителей этого семейства на богатом материале, все Clydonautilidae достоверно известны только из позднего триаса. Указание на находку древнейшего рода клидонаутилид Styrionautilus Mojsisovics, 1902 (Шиманский, 1965, с. 164, табл. 16 , фиг. 8 ) из верхней перми Закавказья он считает сомнительным, так как “Stirionautilus sp.” был “описан по одному экземпляру недостаточной сохранности” (Соболев, 1989, с. 92). Относить башкирский вид к триасовым клидонаутилидам не представляется возможным – последние отличаются как формой раковины, так и отсутствием глубокой вентральной лопасти и аннулярного отростка на дорсальной лопасти. Поэтому более обоснованным решением нам представляется включение нового рода в состав пермоцератид. Род Permoceras Miller et Collinson, 1953 был описан по очень скудному материалу с о-ва Тимор и первоначально определен Ганиэлем (Haniel, 1915) как Nautilus (Aganides) bitauniensis Haniel, 1915. Впоследствии американские авторы (Miller, Collinson, 1953) выделили для него монотипическое семейство и род с единственным видом Permoceras bitauniense (Haniel). Наиболее яркой особенностью этого рода является наличие очень сложной перегородочной линии с узкой глубокой вентральной лопастью, очень глубокой асимметричной клювообразной боковой лопастью и неглубокой воронковидной дорсальной. Такая форма перегородочной линии также известна у юрского Pseudonautilus geinitzi (Oppel). Как образец гомеоморфии эти два не связанные между собой рода приводятся в “Treatise…” (Kummel, 1964, с. K436, рис. 315 ), а также Шиманским (1967; Богословский, Шиманский, 1990). Как пишет Шиманский (1967, с. 58), “Большинство форм с сильно дифференцированной перегородочной линией существовало короткий отрезок времени и, как правило, оставалось изолированными родами (Aturia), монотипическими (Permoceratidae, Gonionautilidae, Siberionautilidae) или очень небольшими семействами (Pseudonautilidae, Clydonautilidae)”. Находка в Шахтау наутилид из семейства Permoceratidae позволяет укрепить аргументацию родственных связей между Permoceratidae и Grypoceratidae, поскольку новый род Foveroceras обладает некоторыми “промежуточными” признаками: его перегородочная линия сложнее, чем у грипоцератид, но немного примитивнее, чем у пермоцератид. С учетом того, что уральский род существовал на два века раньше тиморского, он вполне может быть принят за исходную форму семейства Permoceratidae, считавшегося до настоящего времени тиморским эндемиком. Кроме этого, несомненная близость Foveroceras magnum и Permoceras bitauniense подтверждает наличие миграционных путей между этими бассейнами.
ВЫВОДЫ
Изучение новых материалов из ассельско-сакмарских рифовых известняков Шахтау позволило не только установить новые таксоны наутилид с необычной морфологией, но также расширить наши знания о представителях двух филогенетически связанных семейств, Grypoceratidae и Permoceratidae, и внести некоторые уточнения в систему этой непростой и недостаточно изученной группы.
* * *
Авторы выражают свою искреннюю благодарность А.В. Мазаеву, организатору полевых работ на Шахтау на протяжении последних восьми лет. Мы благодарим генерального директора АО “Сырьевая компания” А.В. Шкурко, создавшего благоприятную атмосферу для работы нашего коллектива на карьере. Кроме этого, мы очень признательны нашим рецензентам С.В. Николаевой и В.В. Митта, их замечания и предложения позволили улучшить качество статьи.
Работа выполнена при финансовой поддержке гранта РНФ, проект № 22-24-00099 “Эволюция сообществ моллюсков раннепермского рифа Шахтау”.
Список литературы
Барсков И.С. Хаос в системе высших таксонов цефалопод // Современные проблемы изучения головоногих моллюсков: морфология, систематика, эволюция, экология и биостратиграфия. Вып. 5. Матер. совещ. (Москва, 29–31 октября 2018 г.). М.: ПИН РАН, 2018. С. 20–24.
Богословский Б.И., Шиманский В.Н. Роль конвергенции в историческом развитии наутилоидей и аммоноидей // Ископаемые цефалоподы. М.: Наука, 1990. С. 5–15 (Тр. ПИН АН СССР. Т. 243).
Круглов М.В. Верхнекаменноугольные и пермские наутилиды Урала // Тр. Геол. музея АН СССР. 1928. Т. 3. С. 63–206.
Руженцев В.Е., Шиманский В.Н. Нижнепермские свернутые и согнутые наутилоидеи Южного Урала. М.: Наука, 1954. С. 1–150 (Тр. ПИН АН СССР. Т. 50).
Соболев Е.С. Триасовые наутилиды Северо-Восточной Азии. Новосибирск: Наука, СО АН СССР, 1989. 193 с.
Шевырев А.А. Макросистема цефалопод: исторический обзор, современное состояние и основные проблемы. 2. Классификация наутилоидных цефалопод // Палеонтол. журн. 2006. № 1. С. 43–52.
Шиманский В.Н. Отряд Nautilida // Основы палеонтологии. Моллюски-головоногие. I / Pед. В.Е. Руженцев. М.: Изд-во АН СССР, 1962. С. 115–154.
Шиманский В.Н. Надотряд Nautiloidea // Развитие и смена морских организмов на рубеже палеозоя и мезозоя. М.: Наука, 1965. С. 157–165 (Тр. ПИН АН СССР. Т. 108).
Шиманский В.Н. Каменноугольные Nautilida. М.: Наука, 1967. С. 1–258 (Тр. ПИН АН СССР. Т. 115).
Шиманский В.Н. Наутилида (изученность, стратиграфическое и географическое распространение, этапы развития). М.: Наука, 1979. С. 1–68 (Тр. ПИН АН СССР. Т. 170).
Щедухин А.Ю. Новые Rhiphaeoceratidae (Nautilida, Cephalopoda) из нижнепермских отложений Шахтау (Башкортостан) // Палеонтол. журн. 2022а. № 6. С. 14–19.
Щедухин А.Ю. Неаммоноидные цефалоподы ассельско-сакмарских отложений рифа Шахтау (республика Башкортостан) // Учен. зап. Казан. ун-та. Сер. Естеств. науки. 2022б. Т. 164. Кн. 2. С. 318–327.
Flower R.H. The ontogeny of Centroceras with remarks of the phylogeny of the Centroceratidae // J. Paleontol. 1952. V. 26. № 3. P. 519–528.
Haniel C.A. Die Cephalopoden der Dyas von Timor // Paläontol. von Timor. 1915. Lief. 3. S. 1–153.
King A.H., Evans D.H. High-level classification of the nautiloid cephalopods: a proposal for the revision of the Treatise Part K //Swiss J. Palaeontol. 2019. V. 138. P. 65–85.
Kummel B. Nautiloidea-Nautilida // Treatise on Invertebrate Paleontology. Pt K. Mollusca 3. Lawrence: Univ. Kansas Press, 1964. P. K383–K457.
Leonova T.B., Shchedukhin A.Yu. Cephalopods of the Early Permian Shakh-Tau reef // Proc. of Kazan Golovkinsky Stratigraphic Meeting, 2019. Late Palaeozoic Sedimentary Earth Systems: Stratigraphy, Environments, Climate and Resources / Ed. Nurgaliev D. Bologna: Filodiritto Editore – Proceeding, 2019. P. 155–158.
Leonova T.B., Shchedukhin A.Yu. Asselian-Sakmarian nautiloids of the Shakh-Tau Reef (Bashkortostan) // Paleontol. J. 2020. V. 54. № 10. P. 1113–1134.
Miller A.K., Collinson C. An abberant nautiloids of the Timor Permian // J. Paleontol. 1953. V. 27. № 2. P. 293–295.
Miller A.K., Dunbar C.O., Condra G.E. Nautiloid Cephalopods of the Pennsylvanian System in Mid-Continent Region // Nebraska Geol. Surv. 2nd Ser. 1933. Bull. 9. 293 p.
Miller A.K., Youngquist W.L. American Permian nautiloids // Mem. Geol. Soc. Amer. 1949. V. 41. P. 1–59.
Niko S., Mapes R.H. Early Carboniferous nautiloids from the Ruddell Shale Member in Arkansas, Midcontinent North America // Paleontol. Res. 2015. V. 19. № 1. P. 52–60.
Niko S., Mapes R.H. Late Carboniferous coiled nautiloids from the Lost Branch Formation of Oklahoma, Midcontinent North America // Paleontol. Res. 2016. V. 20. № 2. P. 75–79.
Shchedukhin A.Y., Leonova T.B. Non-ammonoid cephalopod assemblages of the Early Permian Shakh-Tau Reef // Proc. of Kazan Golovkinsky Young Scientists’ Stratigraphic Meeting, 2020 / Ed. Nurgaliev D. Bologna: Filodiritto Editore – Proceeding, 2020. P. 246–249.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал