

Почвоведение, 2020, № 5, стр. 627-638
Изменения сообществ почвообитающих нематод при постагрогенной трансформации торфяных почв и растительности
М. Г. Юркевич a, *, А. А. Сущук a, Е. М. Матвеева a, Д. С. Калинкина a
a Институт биологии КарНЦ РАН
185910 Петрозаводск, ул. Пушкинская, 11, Россия
* E-mail: svirinka@mail.ru
Поступила в редакцию 21.08.2019
После доработки 30.11.2019
Принята к публикации 29.12.2019
Аннотация
Представлены результаты изучения долгосрочных изменений почв, растительности и их влияния на сообщества почвообитающих нематод в ходе постагрогенного развития сеяных лугов, сформированных на мелиорированных торфяных почвах (Eutric Histosols). Исследования проведены на модельном полигоне “Корзинская низина” (Республика Карелия), на опытных площадках с посевом многолетних трав, внесением удобрений, с наблюдением за сработкой торфяной залежи и изменениями растительных сообществ. В почвенных образцах определяли содержание общего углерода, общего азота и подвижного фосфора, рН солевой вытяжки. Описание растительности экспериментального участка проведено с использованием подхода Браун–Бланке; для определения ботанического состава и урожайности травостоя дополнительно использовали весовой метод. Для сообществ нематод оценивали таксономическое разнообразие, плотность популяций и эколого-трофическую структуру. Анализ почвенных профилей показал, что за последние 40 лет (1974–2015 гг.) произошло уменьшение общей мощности торфяной залежи. Установлено, что направление сукцессии фитоценоза в ходе постагрогенного развития сеяных лугов определяется видом вносимых в почву удобрений при сельскохозяйственном использовании земель: происходит становление двух типов растительных сообществ – дернистощучника влажноразнотравного (Deschampsieta caespitosae humidiherbosum) при внесении минеральных удобрений и пырейно-крупнозлаково-разнотравного травостоя (Elytrigeto magnograminosoherbosum) на фоне минеральных и органических удобрений. Это, в свою очередь, оказало влияние на две трофические группы почвобитающих нематод: ассоциированных с растениями, относительное обилие которых снижалось, и паразитов растений, доля которых в сообществе увеличивалась. Выявлено, что видовая структура травостоя определяет экологическую специализацию доминирующих таксонов фитопаразитических нематод.
ВВЕДЕНИЕ
В условиях глобальных изменений природной среды все более актуальны комплексные исследования растительности с учетом ее прошлого, современного состояния и прогнозирования будущего. Составной частью таких исследований является моделирование постагрогенных сукцессий экосистем. Современные построения в теоретической фитоценологии и экологии сводятся к пониманию того, что необходимо создание моделей целостных характеристик любых систем: сообществ, биогеоценозов, экосистем [13]. В нашем понимании это анализ существующих процессов, в частности, в растительном покрове, требующий выявления всех изменений в сообществах для конкретных природных условий на определенный период времени. В последнее время высказываются мнения об усилении процессов экотонизации биоты, в целом, и растительности, в частности. Восстановительные сукцессии в старовозрастных посевах трав играют большую роль в поддержании и восстановлении биологического разнообразия экосистемы [18]. В то же время высокопродуктивные луговые угодья являются основой сельскохозяйственного животноводства. Общая продуктивность луговых экосистем находится в центре внимания большинства исследователей поскольку напрямую связана с проблемами сельскохозяйственного производства. Существуют работы, показывающие связь видового разнообразия и продуктивности лугов [12, 31]. Однако биологическое разнообразие лугов в процессе постагрогенного развития зависит от целого ряда эдафических и других факторов [11]. Например, луговые экосистемы Республики Карелия развиваются в условиях дефицита солнечной энергии, поэтому весь ход восстановительной сукцессии может быть растянут в пространстве и во времени, иметь специфику, связанную с природно-климатическими особенностями территории.
Вопросы оценки влияния режима использования на луговые сообщества рассматриваются в ряде работ [8, 19, 20], где выявлены закономерности формирования и агрогенной динамики лугов с различным гидрологическим режимом в зависимости от характера их использования: сенокосы, пастбища, неиспользуемые угодья. В настоящее время большие площади ранее залуженных многолетними травами сельскохозяйственных земель в агропромышленном секторе не используются. В связи с этим в агрофитоценозах происходит смена растительности, сопровождающаяся изменением биоразнообразия травянистой растительности, в том числе за счет адвентивных видов.
На территории Республики Карелия существует большое количество заброшенных лугов как автоморфных, так и на торфяных мелиорированных почвах, в связи с этим изучение трансформации фитоценозов и экосистем в ходе постагрогенного развития этих ландшафтов имеет высокую актуальность для региона. Несмотря на то, что отдельные территории были изучены, районы Южной Карелии, ранее наиболее активно используемые в сельском хозяйстве, остались неисследованными.
Для реализации подхода целостного анализа состояния экосистемы и более полного понимания процессов, происходящих в ряду долгосрочных изменений, фитоценологические исследования могут быть дополнены почвенно-зоологическими. Так, почвообитающие нематоды являются одной из наиболее многочисленных и разнообразных групп педобионтов в северных экосистемах [15]. Вместе с другими организмами они участвуют в трансформации веществ, включаясь в различные звенья цепей питания; часть видов паразитирует на сельскохозяйственных культурах, оказывая влияние на рост и развитие растений, нанося существенный ущерб урожаю. Повсеместное распространение, высокое разнообразие и разработанный инструментарий (эколого-популяционные индексы, основанные на анализе фауны) придают почвенным нематодам высокую значимость в экологических исследованиях при оценке изменений почвенных условий и функционального состояния почв.
Исследование фауны нематод луговых почв в ряду долгосрочных сукцессионных изменений в условиях Севера ранее не проводилось. Имеются немногочисленные литературные данные об изменениях структуры сообществ почвенных нематод лугов на осушенных торфяниках в Польше [33], о динамике сообществ почвенных нематод сеяных лугов в условиях полевого эксперимента [32], о состоянии почв и почвенной биоты в ходе вторичной сукцессии агроэкосистем после прекращения обработки почвы [21, 27]. Таким образом, сведения об изменениях нематодофауны северных луговых почв в срезе длительных временных периодов ограничены, не раскрывают многие аспекты организации их сообществ.
Целью настоящего исследования являлось изучение долгосрочных изменений торфяных почв, растительности и их влияния на сообщества почвообитающих нематод в ходе постагрогенного развития сеяных лугов Южной Карелии, а также выявление роли внесения органических и минеральных удобрений в установлении направления сукцессионных изменений.
ОБЪЕКТЫ И МЕТОДЫ
Характеристика модельного полигона. Мониторинговые исследования постагрогенных лугов Южной Карелии, ранее мелиорированных закрытой дренажной сетью, проводили на модельном полигоне “Корзинская низина” (Республика Карелия, 61°49′52.25″ N, 33°11′30.98″ E). Первые исследования агрохимических, биохимических свойств и плодородия торфяников, их изменения под влиянием осушения проведены в 1963–1966 гг. В результате этих исследований показано, что наиболее рентабельной культурой для освоенных торфяников являются многолетние травы [1]. Это послужило началом большого блока работ по изучению формирования луговых агрофитоценозов на мелиорированных землях. Исследовали структуру и динамику биогеоценозов сеяных лугов разного возраста и состава, развивающихся в различных экологических и агротехнических условиях, влияние почвенно-мелиоративных и биотических факторов на продуктивность многолетних трав [4].
Исследуемая стационарная площадка находится на полосе третьего глубокооторфованного участка, осушенного в 1962 г. Это старопахотный хорошо окультуренный участок с ежегодным внесением минеральных удобрений. Предшествующие культуры: с 1967 по 1972 г. многолетние травы, 1972 г. – турнепс, 1973 г. – горох-овес на зеленую массу. В 1974 г. здесь была выделена стационарная площадка для биогеоценологических исследований при создании сеяного луга и систематических наблюдений за интенсивностью сработки торфяной залежи. Проведен посев многолетних трав (тимофеевка луговая Phleum pratense L. и овсяница луговая Festuca pratensis Huds.) на фоне ежегодного внесения минеральных удобрений по всему участку и органических удобрений только на одной его половине. Период интенсивного сельскохозяйственного использования продлился до 1995 г., после чего участок был перезалужен моновидовым посевом P. pratense, а затем в 1997 г. он был изъят из сельскохозяйственного оборота и не подвергался антропогенному воздействию.
В 1974 г. на стационарной площадке был заложен почвенный разрез для наблюдений за интенсивностью сработки торфяной залежи. Общая мощность торфяной залежи составляла 250 см с подстиланием озерно-ледниковыми ленточными глинами и грунтовыми водами на глубине 130 см. Определение мощности торфяной залежи проводилось с периодичностью в 20 лет (1974, 1995, 2015 гг.) с помощью торфяного щупа минимум в пяти точках, а описание почвенного разреза – с периодичностью в 40 лет (1974 и 2015 гг.). Таксономическую принадлежность почв устанавливали на основе критериев классификаций [6, 7, 26]. В полученных почвенных образцах выполняли следующие определения: рН водный и солевой потенциометрически, содержание общего азота по Кьельдалю, подвижного фосфора по Кирсанову [17], общего углерода методом высокотемпературного каталитического сжигания на анализаторе TOC-L CPN Shimadzu. Анализ почвенных параметров и описание почвенного разреза выполняли без учета фактора внесения органических и минеральных удобрений.
Описание растительности экспериментального участка проводили с использованием подхода Браун–Бланке [30] для характеристики постагрогенной сукцессии луговой растительности. Дополнительно применяли весовой метод определения ботанического состава и урожайности травостоя, разработанный в ВНИИ кормов [10]. При геоботаническом анализе участка использовали архивные данные лаборатории геоботаники ИБ КарНЦ РАН.
Для изучения фауны почвообитающих нематод в ходе постагрогенного развития сеяных лугов в 2015 г. выполняли полевые исследования на двух участках модельного полигона с различной историей применения удобрений (минеральные удобрения/минеральные и органические удобрения). Отбор образцов почвы проводили с использованием почвенного бура на глубину 0–15 см в девяти повторностях. Выделение нематод из почвы выполняли по модифицированному методу Бермана с экспозицией 48 ч, фиксатор ТАФ (триэтаноламин : формалин : вода в соотношении 2 : 7 : 91) [22]. Идентификацию нематод осуществляли на временных глицериновых препаратах. Эколого-трофическое группирование нематод вели на основе классификации Йейтса с соавт. [36, 37], адаптированной на русский язык авторами статьи с выделением шести трофических групп: бактериотрофы (Б), микотрофы (М), политрофы (П), хищники (Х), нематоды, ассоциированные с растениями (Аср), и паразиты растений (Пр).
Для анализа фаунистического материала проводили сравнение относительного обилия выявленных таксонов, которое представляет собой долю (%) нематод отдельного вида или рода в общем числе особей всех таксонов в данном сообществе. По относительному обилию выделяли пять групп нематод: эудоминанты (10% от всех обнаруженных особей), доминанты (5.1–10.0%), субдоминанты (2.1–5.0%), рецеденты (1.1–2.0%) и субрецеденты (менее 1.1%) [15]. В настоящей статье роды нематод, относительное обилие которых составляло 10% и более, отнесены к доминирующим таксонам.
Для оценки состояния сообществ нематод использовали следующие параметры: таксономическое разнообразие (количество родов), индекс Шеннона H', плотность популяций (численность) нематод (количество экз./100 г сырой почвы), эколого-трофическая структура сообществ. Таксоны нематод с различными экологическими стратегиями распределяли в группы, выраженные в с-р-значениях по шкале Бонгерса: от колонизаторов (с-р = 1), устойчивых к неблагоприятным условиям существования с быстрыми темпами размножения (r-стратеги в широком смысле), до персистеров (с-р = 5) с низкой репродуктивной способностью и повышенной чувствительностью к факторам окружающей среды (K-стратеги в широком смысле) [23]. Также проанализированы эколого-популяционные индексы, характеризующие почвенную трофическую сеть: индексы структурирования (Structure index, SI) и обогащения (Enrichment index, EI) почвенной трофической сети, индекс преобладающего пути разложения органического вещества в почве (Channel index, CI) [24]. Детально расчет экологических индексов и их интерпретация рассмотрены ранее [2, 9].
Для оценки изменений сообществ почвенных нематод лугового биоценоза в ходе восстановительной сукцессии привлечены архивные данные лаборатории паразитологии животных и растений ИБ КарНЦ РАН (нематологические сборы 1974 г.). Статистическую обработку данных проводили с использованием H-критерия Краскела–Уоллиса для малого числа повторностей. Различия между группами считали достоверными при р < 0.05. Числовые данные представлены в форме М ± SE (среднее значение ± стандартная ошибка среднего). Для выявления различий между локальными сообществами почвенных нематод проводили ординацию луговых биоценозов методом главных компонент на основе относительного обилия доминирующих таксонов нематод. Расчеты выполнены при помощи программы PAST 1.68 [25].
Для выполнения исследования почвенных нематод использовали научное оборудование Центра коллективного пользования Федерального исследовательского центра “Карельский научный центр Российской академии наук”.
РЕЗУЛЬТАТЫ
Почва исследованного участка в 1974 г. характеризовалась как перегнойно-торфяная, низинного типа (Eutric Histosols), на торфяной залежи мощностью 250 см, сложенной древесно-осоковым торфом и подстилаемой озерно-ледниковыми ленточными глинами и грунтовыми водами на глубине 130 см. По описанию почвенного разреза 1974 г. пахотный горизонт сложен темно-коричневым хорошо разложившимся торфом со степенью разложения 30–35%. Экспериментальный участок имел следующие почвенные характеристики без учета фактора внесения удобрений: рНKCl 5.06, С 54.99%, N 2.92%, Р2О5 0.66% для глубины 0–15 см.
По результатам обследования 1995 г. усадка торфа составила 29 см (в среднем мощность торфяной залежи уменьшилась с 250 до 221 см). После прекращения использования участка мощность торфяной залежи составила 224 см (2015 г.), наблюдалось небольшое ее увеличение (на 3 см), что связано с процессом торфонакопления, обусловленного изъятием участка из сельскохозяйственного оборота.
По результатам почвенного обследования 2015 г. почва участка характеризуется как торфозем агроминеральный (Eutric Histosols), почвенные параметры изменились следующим образом: рНKCl 5.29; С 33.72%; N 2.21%; Р2О5 0.56% для глубины 0–15 см.
Анализ результатов флористического обследования территории в 70-е годы показал, что при интенсивном использовании травостоев в фитоценозах длительное время сохранялась высокая доля участия сеяных видов (на девятый год пользования доля Phleum pratense – до 60%). При перезалужении участка в 1995 г. трансформация фитоценоза на исследуемом участке во многом зависела от неравномерной усадки торфа и неравномерного разброса удобрений. Из-за данных обстоятельств уже на второй год использования травостой Phleum pratense характеризовался высокой степенью мозаичности (доля сеяного вида не превышала 40%); были отмечены крупные куртины пырея ползучего Elytrigia repens L. и разнотравья. При дефиците минерального питания и появлении понижений микрорельефа отмечены крупные куртины луговика дернистого Deschampsia cespitosa L. Фактический урожай фитоценоза на второй год использования составил 2.90 т/га.
В период последнего десятилетия на территории опытного полигона при отсутствии агрогенного воздействия сформировались два типа растительных сообществ: мелкозлаково-разнотравный луг с высокой степенью мозаичности (на участке, где ранее производилось внесение минеральных удобрений) и крупнозлаково-разнотравный луг (на участке, где ранее производилось совместное внесение органических и минеральных удобрений).
Геоботаническое описание участков показало, что в первом случае – это дернистощучковый влажноразнотравный (Deschampsieta caespitosae humidiherbosum) травостой с явно выраженными куртинами видов различных экологических групп, что указывает на то, что данный участок характеризуется варьированием условий плодородия, кислотности, увлажнения почв. Так, отмечены крупные куртины требовательных к плодородию почв видов: подмаренника мягкого Galium mollugo L., крапивы двудомной Urtica dioica L. и ясколки дернистой Cerastium holosteoides Fr. На части полигона кислотность почвы заметно отличается от сопредельных участков, на что указывает произрастание фитоиндикатора (щавель кислый Rumex acetosa L.). Травостой дополняют как мезофитные кормовые злаки (Phleum pratense, полевица тонкая Agrostis tenuis Sibth.), так и гидрофитный вид двукисточник тростниковый Phalaroides arundinacea L. Урожайность травостоя – крайне низкая (0.9 т/га сухого вещества).
Второй участок – пырейно-крупнозлаково-разнотравный луг является более плодородным из-за достаточного внесения органических удобрений, что подтверждается доминированием в травостое Elytrigia repens с небольшими куртинами Urtica dioica. Участок более выровнен и по увлажненности, в связи с чем куртины влаголюбивого Phalaroides arundinacea незначительны. Урожайность травостоя существенно выше – 3.74 т/га сухого вещества.
Почвообитающие нематоды. В почве исследуемых участков обнаружено 47 видов нематод во все сроки наблюдений и при различных типах луговой растительности (табл. 1). Сравнительный анализ разнообразия фауны почвообитающих нематод по данным 1974 г. и 2015 г. показал увеличение числа выявленных видов (39 против 30) и индекса разнообразия Шеннона в ходе восстановительной сукцессии (табл. 2), а также снижение количества пионерных видов, быстро реагирующих на внесение органического вещества, и появление видов с высокими значениями по с-р-шкале Бонгерса [23]. Так, если в почве сеяного луга доля нематод-бактериотрофов с с-р = 1 составляет 11.8%, то в ходе сукцессии она снижается (3.2–5.5%) и, наоборот, доля высокоспециализированных таксонов бактериотрофов (с-р = 3–4) увеличивается с 3.1 до 9.9–12.9% от общей численности нематод. Кроме того, таксоны, тяготеющие к влажным условиям среды обитания (E-umonhystera, Monhystrella, Prodesmodora, Cylindrolaimus, Prismatolaimus и др.) в почве лугов (2015 г.), представлены в большей степени по сравнению с сеяным лугом.
Таблица 1.
Таксономическое разнообразие и относительное обилие нематод в почве луговых биоценозов, сформированных на торфяных почвах в ходе постагрогенной сукцессии
| Таксон | Значение по с-р шкале | Относительное обилие, % | ||
|---|---|---|---|---|
| тимофеевка и овсяница (1974 г.), n = 6 |
дернистощучник влажноразнотравный (2015 г.), n = 9 | пырейный травостой (2015 г.), n = 9 |
||
| Бактериотрофы | ||||
| Mesorhabditis sp. | 1 | 7.84 ± 4.59 | − | − |
| Panagrobelus sp. | 1 | − | − | 0.45 ± 0.35 |
| Panagrolaimus rigidus | 1 | 3.01 ± 1.53 | 3.62 ± 0.86 | 2.21 ± 0.80 |
| Protorhabditis sp. | 1 | 0.17 ± 0.17 | − | − |
| Rhabditis sp. | 1 | 0.74 ± 0.47 | 1.86 ± 0.57 | 0.54 ± 0.27 |
| Acrobeles sp. | 2 | 0.19 ± 0.19 | 0.39 ± 0.26 | 1.31 ± 0.50 |
| Acrobeloides buetschlii | 2 | 21.72 ± 6.60 | 3.40 ± 0.69 | 2.18 ± 0.79 |
| Anaplectus granulosus | 2 | − | 0.17 ± 0.17 | 10.59 ± 2.57 |
| Cephalobus persegnis | 2 | 10.22 ± 6.21 | − | 0.56 ± 0.41 |
| Ceratoplectus sp. | 2 | 0.19 ± 0.19 | − | − |
| Cervidellus sp. | 2 | − | 0.78 ± 0.45 | 2.94 ± 0.84 |
| Chiloplacus sp. | 2 | 0.49 ± 0.49 | 7.63 ± 1.59 | 3.30 ± 0.66 |
| Eucephalobus sp. | 2 | − | 6.56 ± 1.45 | 12.87 ± 2.11 |
| Eumonhystera sp. | 2 | − | 0.55 ± 0.28 | 0.77 ± 0.31 |
| Heterocephalobus elongatus | 2 | 4.85 ± 2.25 | 0.13 ± 0.13 | − |
| Monhystrella plectoides | 2 | − | 0.85 ± 0.37 | 1.51 ± 0.52 |
| Plectus sp. | 2 | 2.27 ± 0.73 | 5.55 ± 1.44 | 2.32 ± 0.84 |
| Prodesmodora sp. | 2 | − | 5.24 ± 1.86 | 8.64 ± 2.39 |
| Tylocephalus sp. | 2 | − | − | 0.39 ± 0.26 |
| Wilsonema sp. | 2 | 1.07 ± 0.56 | 1.57 ± 0.59 | 0.82 ± 0.55 |
| Cylindrolaimus sp. | 3 | − | 0.77 ± 0.64 | 1.09 ± 0.37 |
| Metateratocephalus sp. | 3 | − | 1.04 ± 0.43 | 0.11 ± 0.11 |
| Odontolaimus sp. | 3 | − | − | 0.64 ± 0.27 |
| Prismatolaimus sp. | 3 | 2.93 ± 1.57 | 6.93 ± 1.94 | 10.15 ± 2.08 |
| Teratocephalus sp. | 3 | − | 0.97 ± 0.45 | 0.94 ± 0.39 |
| Alaimus primitivus | 4 | 0.19 ± 0.19 | 0.21 ± 0.21 | − |
| Микотрофы | ||||
| Aphelenchoides sp. | 2 | 2.54 ± 0.69 | 4.34 ± 1.23 | 1.12 ± 0.49 |
| Aphelenchus avenae | 2 | − | 0.84 ± 0.38 | 1.57 ± 0.56 |
| Ditylenchus sp. | 2 | 2.39 ± 1.05 | 1.49 ± 0.65 | 1.72 ± 0.51 |
| Paraphelenchus sp. | 2 | 0.16 ± 0.16 | − | − |
| Tylencholaimus sp. | 4 | − | − | 0.38 ± 0.25 |
| Политрофы | ||||
| Dorylaimus sp. | 5 | 0.19 ± 0.19 | − | − |
| Eudorylaimus sp. | 5 | 1.96 ± 0.79 | 3.46 ± 1.19 | 2.57 ± 0.54 |
| Laimydorus sp. | 5 | 0.18 ± 0.18 | − | − |
| Mesodorylaimus sp. | 5 | 0.35 ± 0.22 | 0.22 ± 0.22 | 1.84 ± 0.75 |
| Хищники | ||||
| Clarkus papillatus | 4 | 0.55 ± 0.55 | 0.21 ± 0.21 | − |
| Нематоды, ассоциированные с растениями | ||||
| Aglenchus agricola | 2 | 20.06 ± 1.93 | 0.22 ± 0.22 | 0.33 ± 0.22 |
| Coslenchus sp. | 2 | 0.58 ± 0.39 | 0.54 ± 0.27 | 0.38 ± 0.25 |
| Filenchus sp. | 2 | 5.79 ± 1.51 | 0.97 ± 0.64 | 1.89 ± 0.84 |
| Lelenchus leptosoma | 2 | 0.66 ± 0.66 | − | 0.75 ± 0.29 |
| Malenchus sp. | 2 | 3.36 ± 1.42 | 0.68 ± 0.27 | 0.17 ± 0.17 |
| Tylenchus sp. | 2 | 0.56 ± 0.25 | − | − |
| Паразиты растений | ||||
| Paratylenchus | 2 | 4.63 ± 4.63 | 27.39 ± 3.21 | 1.15 ± 0.45 |
| Helicotylenchus sp. | 3 | 0.17 ± 0.17 | − | − |
| Pratylenchus sp. | 3 | − | 5.88 ± 1.03 | 21.83 ± 1.82 |
| Tylenchorhynchus sp. | 3 | − | 5.32 ± 1.48 | − |
| Паразиты беспозвоночных* | ||||
| Steinernema sp. | − | − | 0.22 ± 0.22 | − |
| Всего таксонов | 30 | 33 | 34 | |
Таблица 2.
Характеристики сообществ почвенных нематод луговых биоценозов в ходе постагрогенной сукцессии
| Параметр | Сеяный луг, 1974 г. (тимофеевка и овсяница), n = 6 | Земли, выведенные из сельскохозяйственного оборота, 2015 г. | |
|---|---|---|---|
| дернистощучник влажноразнотравный, n = 9 |
пырейный травостой, n = 9 |
||
| Численность, экз./100 г почвы | 2704 ± 340a | 4978 ± 398b | 2581 ± 252a |
| Количество видов | 30 | 33 | 34 |
| Н ', индекс Шеннона | 3.4 ± 0.2a | 4.2 ± 0.1b | 4.0 ± 0.1b |
| Эколого-трофические группы, % | |||
| Бактериотрофы | 53.5 ± 6.6ab | 46.9 ± 2.8a | 65.6 ± 2.4b |
| Микотрофы | 5.0 ± 1.2a | 7.5 ± 1.5a | 4.7 ± 0.7a |
| Политрофы | 2.9 ± 0.9a | 3.7 ± 1.2a | 4.3 ± 0.9a |
| Хищные нематоды | 0.7 ± 0.6a | 0.2 ± 0.2a | 0 |
| Нематоды, ассоциированные с растениями | 32.2 ± 4.2a | 2.5 ± 0.8b | 3.2 ± 0.7b |
| Паразиты растений | 5.7 ± 4.6a | 39.2 ± 2.2b | 22.2 ± 1.9c |
Современные участки с различным составом растительного покрова (2015 г.) не имели заметных различий в таксономическом разнообразии и относительном обилии нематод. Дернистощучник влажноразнотравный – луг, сформировавшийся на участке с внесением минеральных удобрений – выделялся значимо более высоким показателем общей численности (табл. 2).
По характеру доминирования таксоны, входящие в состав фауны нематод исследованных луговых биоценозов, группируются следующим образом: в почве сеяного луга (1974 г.) эудоминанты и доминанты представлены пятью видами (три из трофической группы бактериотрофов и два – из группы нематод, ассоциированных с растением). В почве современного пырейного травостоя и дернистощучника эудоминантами и доминантами являются представители нематод-бактериотрофов и паразитов растений. На долю субдоминантов в почве сеяного луга приходится около 27% от общего числа таксонов, в ходе восстановительной сукцессии вклад группы уменьшается (12–18%). С другой стороны, рецеденты сеяного луга были представлены одним таксоном, а на землях, выведенных из сельскохозяйственного оборота, разнообразие данной группы возрастает (3–8 таксонов). Субрецеденты – самая многочисленная группа таксонов в исследованных почвах независимо от года наблюдений (15–18 таксонов или 44–54% от общего количества таксонов каждого из сообществ). Так, если общая структура фауны зависит в первую очередь от доминирующих таксонов, то именно массовые группы субрецедентов определяют ее разнообразие.
В целом, рассмотрение характера доминирования таксонов, входящих в состав фауны нематод, показало, что специфичность состава как доминирующих, так и редких видов зависит, в первую очередь, от характера использования луга (интенсивное использование в сельском хозяйстве и длительность заброшенности луга), а затем уже от формирования особенностей условий обитания (тип удобрения и растительности).
В ходе восстановительной сукцессии претерпела изменения и эколого-трофическая структура сообществ нематод. В почве исходного сеяного луга (1974 г.) сообщество представлено всеми шестью трофическими группами с доминированием бактериотрофов и нематод, ассоциированных с растением (Аср). Земли, выведенные из сельскохозяйственного оборота, характеризовались снижением относительного обилия нематод группы Аср и возрастанием популяций паразитов растений (Пр). Сообщество нематод луга, сформировавшегося на участке, где ранее вносилось органическое вещество (пырейный травостой), имеет специфические черты: высокий вклад бактериотрофов в фауну и меньшая доля нематод Пр по сравнению с дернистощучником влажноразнотравным.
Ординация луговых биоценозов методом главных компонент на основе доминирующих таксонов позволила установить роды нематод, количественно преобладающие в фауне и определяющие различия между локальными сообществами почвенных нематод (рис. 1). Сеяный луг, находящийся в сельскохозяйственном использовании (тимофеевка и овсяница, 1974), выделяется по роду Aglenchus (Аср) и двум родам нематод-бактериотрофов (Acrobeloides и Cephalobus). Снятие антропогенного вмешательства имело следствием формирование различного естественного травостоя на исследованных участках, что сказалось на доминирующих в фауне таксонах, в основном из группы фитопаразитов: дернистощучник влажноразнотравный обособляется за счет высокого обилия нематод р. Paratylenchus (эктопаразит), пырейный травостой – р. Pratylenchus (мигрирующий эндопаразит).
Рис. 1.
Ординация луговых биоценозов на основе относительного обилия доминирующих таксонов нематод (с долей в структуре сообщества более 10%) в осях первой и второй главных компонент (ГК1 и ГК2): Phleum Festuca 1974 – сеяный луг (тимофеевка и овсяница), 1974 г.; Deschampsia 2015 – дернистощучник влажноразнотравный, 2015 г.; Elytri-gia 2015 – пырейный травостой, 2015 г.
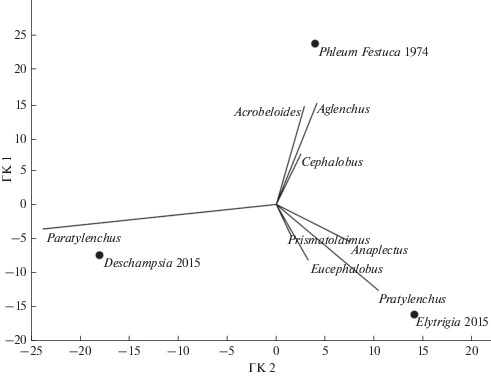
Анализ эколого-популяционных индексов, основанных на фауне нематод и характеризующих состояние почвенной экосистемы, показал уменьшение индекса обогащения почвенной трофической сети (EI) и увеличение индекса преобладающего пути разложения органического вещества в почве (CI) в ходе постагрогенного развития лугов, однако различия статистически незначимы. Индекс структурирования почвенной трофической сети (SI) имел средние значения (табл. 3). Соотношение индексов SI и EI позволяет оценить почвенные экосистемы современных луговых биоценозов (2015 г.) как ненарушенные со сложными трофическими сетями и умеренным уровнем обогащения почв органическим веществом. Сеяный луг (1974 г.), напротив, характеризуется значениями индексов, свидетельствующими об упрощенной и нестабильной трофической сети, приуроченной к высоконарушенной почвенной экосистеме (индекс SI меньше 50, EI больше 50).
Таблица 3.
Эколого-популяционные индексы сообществ почвенных нематод луговых биоценозов в ходе постагрогенной сукцессии
| Параметр | Сеяный луг, 1974 г. (тимофеевка и овсяница), n = 6 |
Земли, выведенные из сельскохозяйственного оборота, 2015 г. | |
|---|---|---|---|
| дернистощучник влажноразнотравный, n = 9 |
пырейный травостой, n = 9 |
||
| SI | 40.0 ± 10.7a | 53.4 ± 3.9a | 53.0 ± 4.1a |
| EI | 51.4 ± 9.6ab | 43.6 ± 3.6b | 25.6 ± 3.6a |
| CI | 10.7 ± 2.2a | 24.8 ± 5.6a | 23.6 ± 12.1a |
Примечание. n – объем выборки; SI – индекс структурирования почвенной трофической сети, EI – индекс обогащения почвенной трофической сети, CI – индекс преобладающего пути разложения органического вещества в почве. Значения в строке с разными буквенными обозначениями статистически различаются (р < 0.05).
Сообщества нематод, сформировавшиеся к 2015 г., имели значимые различия индекса EI, зависящие от состава растительного покрова лугов: на участке с дернистощучником обогащение почвенной трофической сети органическим веществом больше, чем в пырейном травостое.
ОБСУЖДЕНИЕ
Торфяные почвы Севера с легкостью подвергаются деградации при нерациональном использовании. Деградация почв может привести к тяжелым последствиям для биогеоценозов, которые отличаются низким биологическим разнообразием, и потеря каждого вида влечет за собой нарушения функционирования всей экосистемы. Большую роль в обеспечении неоднородности растительности играет почвенная мозаичность. Исследования, проведенные на торфяных почвах, показали повсеместную горизонтальную неоднородность травостоя, возникающую на первоначально хорошо подготовленной и спланированной поверхности полей. Основная причина образования куртин – неравномерная осадка торфяной залежи, происходящая из-за микрокомплексности растительности болот, благодаря чему накапливается торф разной влагоемкости, степени разложения и уплотнения при осушении [14, 28, 29].
На осушенных торфяных почвах, богатых органическим веществом, в процессе их эксплуатации происходит минерализация этого вещества и переход азота из труднодоступных в легкодоступные формы. Экспериментальный участок в целом характеризовался потерей органического углерода в силу минерализации торфа. Запасы общего азота в исследуемых почвах также снизились, но изменения были незначительны. Согласно литературным данным, изменения, происходящие в течение 20-летнего окультуривания торфяных низинных почв, способствуют снижению кислотности почв, увеличению количества обменных оснований, подвижного фосфора, уменьшению содержания аммиачного азота [16]. В результате длительного окультуривания не наблюдается снижения пространственной вариабельности агрохимических показателей торфяных низинных почв.
Об изменении агрохимических показателей почв в луговых биоценозах, сформированных на месте заброшенных экспериментальных участков, свидетельствуют и нематологические данные: снижение в 2015 г. по сравнению с 1974 г. численности нематод-бактериотрофов со значением 1 по с-р-шкале Бонгерса [23]. Данная группа относится к типичным колонизаторам, указывает на всплеск бактериальной активности в почве, быстро увеличивает численность после резких изменений или нарушений среды обитания (внесение удобрений, вспашка и т.д.), а затем замещается нематодами со значением 2 по с-р-шкале, которые обычно преобладают в сообществах, формируя базальную трофическую сеть, и за счет широкой экологической пластичности являются стабильным компонентом почв [24]. Тенденцию подтверждают и значения индекса обогащения EI почвенной трофической сети, связанного с количеством доступного органического вещества в почве, который имел более низкие значения в сборах 2015 г.
Луговые фитоценозы относятся к категории высокоподвижных неустойчивых сообществ. В них обычно наблюдается выраженное пятнистое или мозаичное сложение растительности. Горизонтальная неоднородность травостоя, как правило, является следствием эколого-фитоценотических процессов, протекающих в растительном сообществе, главным образом, в связи с неоднородностью экотопа. Наряду с неравномерной осадкой торфяной залежи отмечены нарушения агротехники, влияющие на урожайность трав. Таким образом, на основе анализа геоботанического описания растительности показано, что виды применяемых удобрений на фоне различных нарушений агротехники (невыровненность полей, неравномерное внесение удобрений, отсутствие интенсивного использования травостоя) существенно влияют на направление растительной сукцессии фитоценоза при прекращении антропогенной нагрузки.
Сравнительное исследование лугов позволило выявить особенности изменений эколого-трофической структуры сообществ почвообитающих нематод в ходе постагрогенной сукцессии биоценоза. Одна из наиболее важных реакций сообществ нематод, показанная в ряду долгосрочных изменений – это уменьшение обилия нематод, ассоциированных с растениями, и увеличение популяций паразитов растений.
Ранее при изучении тенденций восстановления сообществ нематод после полной деградации почвы и растительного покрова было установлено, что на ранних сроках доминируют виды, устойчивые к неблагоприятным условиям среды (бактериотрофы), на более поздних стадиях восстановления почвенных экосистем увеличивается численность хищных и фитопаразитических нематод, что свидетельствует о расширении и усложнении почвенной трофической сети, и является показателем процесса регенерации биоценоза в целом [3]. Известно, что нематоды, ассоциированные с растениями, имеют более широкие трофические возможности: они питаются как за счет эпидермальных клеток корней, так и содержимым гифов грибов. Можно предположить, что нематоды данной группы, благодаря факультативной связи с растениями, получают большие преимущества в сообществах с нестабильной растительной группировкой. По мере формирования сложившегося растительного сообщества возрастает обилие нематод-паразитов растений, более специализированной трофической группы, облигатно связанной с растением-хозяином.
Сходная закономерность была получена в злаково-разнотравных фитоценозах при прекращении использования сеяных лугов и, следовательно, при отсутствии агротехнических мероприятий: без внесения минеральных удобрений сеяные виды злаков на осушенных торфяниках постепенно выпадали, их место занимали дикорастущие травы, что влекло за собой увеличение численности нематод, связанных с растениями [2, 5]. Кроме того, высокая численность и доля в фауне нематод-фитопаразитов наблюдались на осушенных торфяниках при их сравнении с естественными, неосушенными лугами [33], при этом эффект сохранялся в течение 100 лет после мелиорации [34].
При общем высоком обилии фитопаразитических нематод в почве лугов, установившемся к 2015 г., выявлены различия в составе доминирующих родов нематод, связанные с особенностями растительного покрова. Так, дернистощучник характеризовался высоким обилием нематод р. Paratylenchus (идентифицированы два вида – Paratylenchus microdorus (15.6%) и P. nanus (11.8%)), являющихся эктопаразитами корневой системы широкого круга травянистых растений. В пырейном травостое доминировали мигрирующие эндопаразитические нематод р. Pratylenchus (21.8%).
Согласно литературным данным, нематоды рода Paratylenchus имеют значение 2 по с-р-шкале (r-стратеги среди фитопаразитов), обладают высокой экологической пластичностью и могут выступать как супердоминанты при различных нарушениях окружающей среды [33, 35]. В большей степени описанная закономерность свойственна нематодам вида Paratylenchus nanus.
Нематоды рода Pratylenchus, преобладающие в пырейном травостое, имеют значение 3 по с-р-шкале и более требовательны к состоянию почвенной среды. В настоящее время данный фитоценоз сформирован с внедрением Phalaroides arundinacea, высокопродуктивного злака и ценного кормового растения, который расселился после его посева на соседнем участке в 1998 г. В сочетании с пыреем они сформировали быстроразрастающийся и долговечный травостой (10–15 лет), в котором созданы благоприятные условия (мощная корневая система растений) для обитания пратиленхов. В литературе отмечается, что увеличение численности фитопаразитических нематод с высокими (3–5) значениями по с-р-шкале (персистеры или K-стратеги), к которым относится и р. Pratylenchus, указывает на более поздние стадии сукцессии и стабильность среды обитания [33].
Известно, что виды растений, входящие в состав фитоценоза, зачастую являются индикаторами экологических условий. Так, анализ геоботанических описаний растительности позволил охарактеризовать участок с пырейным травостоем как более плодородный и выровненный по уровню увлажнения, отличающийся высокой урожайностью, что согласуется с полученными нематологическими данными.
Таким образом, установлено, что видовая структура травостоя, сформированного в зависимости от характера предшествующих обработок (внесение удобрений), определяет экологическую специализацию доминирующих таксонов фитопаразитических нематод (экто- или эндопаразиты). Показано, что в ходе долгосрочной постагрогенной сукцессии наблюдаются увеличение индекса Шеннона, характеризующего разнообразие фауны нематод, и перестройка структуры сообществ – снижается обилие нематод, ассоциированных с растениями, и возрастает количество паразитов растений, т.е. происходит замещение группы нематод, факультативно связанных с растениями, на более специализированную трофическую группу, имеющую облигатную связь с растением-хозяином, что указывает на поздние стадии сукцессии и сложившееся растительное сообщество, обеспечивающее стабильность среды обитания.
ЗАКЛЮЧЕНИЕ
Проведенное исследование позволило выявить основные тренды динамики сеяных лугов, сформированных на мелиорированных торфяных почвах, в ходе постагрогенной сукцессии. Анализ почвенных профилей показал, что на землях, изъятых из сельскохозяйственного оборота, за последние 40 лет (1974–2015 гг.) произошло уменьшение общей мощности торфяной залежи. Прекращение сельскохозяйственного использования лугов привело к смене видового состава фитоценоза. На основе анализа геоботанического описания растительности показано, что виды применяемых удобрений на фоне различных нарушений агротехники существенно влияют на направление растительной сукцессии фитоценоза; происходит становление двух типов растительных сообществ в зависимости от типа удобрений: дернистощучника влажноразнотравного при внесении минеральных удобрений и пырейно-крупнозлаково-разнотравного на фоне минеральных и органических удобрений травостоя. Это, в свою очередь, оказало влияние на сообщества почвообитающих нематод. Наибольшие изменения структуры сообществ почвенных нематод связаны с относительным обилием трофических групп нематод, связанных с растениями: уменьшением доли нематод, ассоциированных с растениями, и увеличением доли паразитов растений. Установлено, что видовая структура травостоя определяет экологическую специализацию фитопаразитических нематод; выявлена связь между составом растительного покрова и доминированием отдельных таксонов. Так, в дернистощучнике преобладали эктопаразиты р. Paratylenchus, в пырейном травостое – эндопаразиты р. Pratylenchus. Такие особенности трофической структуры сообществ почвообитающих нематод указывают на поздние стадии сукцессии фитоценозов и сложившееся растительное сообщество, обеспечивающее стабильность среды обитания.
Список литературы
Бухман В.А., Цыба М.М. Агрохимические свойства и плодородие торфяных почв Карелии. Петрозаводск, 1967. 105 с.
Груздева Л.И., Матвеева Е.М., Коваленко Т.Е. Изменения в комплексах почвенных нематод под влиянием удобрений // Почвоведение. 2007. № 6. С. 756–768.
Груздева Л.И., Сущук А.А. Тенденции восстановления сообществ нематод после деградации почвенного покрова // Известия РАН. Сер. Биологическая. 2010. № 6. С. 750–755.
Дубровина И.А., Богданова Т.В. Корзинский научный стационар – полвека на службе науке // Труды Карельского научного центра РАН. 2016. № 6. С. 3–15.
Елисеева Т.С., Козлов Л.Г. Видовой состав травостоя сеяных лугов на осушенных торфяника // Строение и продуктивность природных и сеяных лугов. Петрозаводск: Карельский филиал АН СССР, 1980. С. 19–28.
Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
Классификация и диагностика почв СССР. М.: Колос, 1977. 224 с.
Кокорина Н.В. Оценка биоразнообразия пойменных фитоценозов // Вестник Югорского гос. ун-та. 2009. № 3(14). С. 44–51.
Матвеева Е.М., Сущук А.А. Особенности сообществ почвенных нематод в различных типах естественных биоценозов: информативность параметров оценки // Известия РАН. Сер. Биологическая. 2016. № 5. С. 551–560. https://doi.org/10.7868/S0002332916040093
Методика опытных работ на сенокосах и пастбищах. М.: ВНИИ кормов им. В.Р. Вильямса, 1961. 287 с.
Миркин Б.М., Хасанова Г.Р., Абрамова Л.М., Суюндуков Я.Т. Сукцессии в посевах многолетних трав: закономерности, вклад в биоразнообразие и устойчивость агроэкосистем // Бюллетень Московского общества испытателей природы. Отд. биол. 2002. Т. 107. Вып. 4. С. 39–45.
Панченкова И.А. История изучения и современное состояние лугов Приволжской возвышенности // Вестник Оренбургского гос. ун-та. 2010. № 6(112). С. 16–18.
Сизых А.П., Воронин В.И. Почвенно-геоботаническое профилирование в изучении сообществ контакта “лес-экстразональная степь” и зональной лесостепи (Байкальский регион) // Экология. 2013. № 2. С. 83–89.
Синькевич Е.И. Эволюция и плодородие осушенных торфяных почв Европейского Севера России. Автореф. дис. … д. с.-х. наук. СПб., 1997. 73 с.
Соловьева Г.И. Экология почвенных нематод. Л.: Наука, 1986. С. 5–14.
Стрелкова А.А., Марченкова Н.Е., Перевозчикова Е.М., Кябелева Г.К., Амозова М.П., Павлова Н.Н. Пространственное варьирование агрохимических свойств пахотных торфяных низинных почв // Почвы Карелии и вопросы их мелиорации. Петрозаводск: Карельский филиал АН СССР, 1982. С. 49–62.
Теория и практика химического анализа почв / Под ред. Воробьевой Л.А. М.: ГЕОС, 2006. 400 с.
Шварц Е.А. Сохранение биоразнообразия: сообщества и экосистемы. М.: Товарищество научных изданий КМК, 2004. 111 с.
Шушпанникова Г.С. Формирование и деградация лугов под влиянием сенокошения и выпаса в поймах рек Вычегды и Печоры // Экология. 2014. № 1. С. 40–44.
Юркевич М.Г. Горизонтальная структура агроценозов южной Карелии // Экология и география почв. Петрозаводск: КарНЦ РАН, 2009. С. 105–116.
Adl S.M., Coleman D.C., Read F. Slow recovery of soil biodiversity in sandy loam soils of Georgia after 25 years of no-tillage management // Agriculture, Ecosystems and Environment. 2006. V. 114. P. 323–334. https://doi.org/10.1016/j.agee.2005.11.019
van Bezooijen J. Methods and techniques for nematology. Wageningen: The Netherlands, Wageningen University Press, 2006. 112 p.
Bongers T. The maturity index: an ecological measure of environmental disturbance based on nematode species composition // Oecologia. 1990. V. 83. P. 14–19. https://doi.org/10.1007/BF00324627
Ferris H., Bongers T., de Goede R.G.M. A framework for soil food web diagnostics: extension of the nematode faunal analysis concept // Appl. Soil Ecology. 2001. V. 18. P. 13–29. https://doi.org/10.1016/S0929-1393(01)00152-4
Hammer Ø., Harper D.A.T., Ryan P.D. Past: paleontological statistics software package for education and data analysis // Paleontological Electronica. 2001. 4(1). P. 1–9. http://palaeo-electronica.org/2001_1/past/issue1_01.htm
IUSS Working Group WRB. 2014. World Reference Base for Soil Resources 2014. International soil classification system for naming soils and creating legends for soil maps. World Soil Resources Reports No. 106. FAO, Rome.
Kardol P., Bezemer T.M., van der Wal A., van der Putten W.H. Successional trajectories of soil nematode and plant communities in a chronosequence of ex-arable lands // Biological Conservation. 2005. V. 126. P. 317–327. https://doi.org/10.1016/j.biocon.2005.06.005
Kempen B., Brus D.J., Stoorvogel J.J. Three-dimensional mapping of soil organic matter content using soil type-specific depth functions // Geoderma. 2011. V. 162. P. 107–123.
Kõlli R., Ellermäe O., Köster T., Lemetti I., Asi E., Kauer K. Stocks of organic carbon in Estonian soils // Estonian J. Earth Sci. 2009. V. 58. P. 95–108.
Raabe U., Brandes D. Flora und Vegetation der Dörfer im nordöstlichen Burgenland // Phytocoenologia. 1988. V. 16. № 2. P. 225–258.
Robertson T., Rosenzweig C., Benson V., Williams J.R. Projected impacts of carbon dioxide and climate change on agriculture in the Great Plains // Challenges in Dryland Agriculture. 1993. № l. P. 675–677.
Viketoft M., Sohlenius B., Boström S., Palmborg C., Bengtsson J., Berg M.P., Huss-Danell K. Temporal dynamics of soil nematode communities in a grassland plant diversity experiment // Soil Biol. Biochem. 2011. V. 43. P. 1063–1070. https://doi.org/10.1016/j.soilbio.2011.01.027
Wasilewska L. Changes in the structure of the soil nematode community over long-term secondary grassland succession in drained fen peat // Appl. Soil Ecology. 2006. V. 32. № 2. P. 165–179.
Wasilewska L. Impact of human activities on nematode communities in terrestrial ecosystems // Ecology of arable land – perspectives and challenges. Kluwer Academic Publishers, 1989. P. 123–132.
Wasilewska L. Soil invertebrates as bioindicators, with special reference to soil-inhabiting nematodes // Russ. J. Nematology. 1997. V. 5. № 2. P. 113–126.
Yeates G.W., Bongers T., de Goede R.G.M., Freckman D.W., Georgieva S.S. Feeding habits in soil nematode families and genera: An outlain for soil ecologists // J. Nematology. 1993. V. 25. № 3. P. 315–331.
Yeates G.W., Wardle D.A., Watson R.N. Relationships between nematodes, soil microbial biomass and weed-management strategies in maize and asparagus cropping systems // Soil Biol. Biochem. 1993. V. 25. № 7. P. 869–876. https://doi.org/10.1016/0038-0717(93)90089-t
Дополнительные материалы отсутствуют.