

Почвоведение, 2021, № 7, стр. 849-861
Биологическая память почв об изменениях условий почвообразования и антропогенной деятельности в прошлом: микробная и ферментная составляющие
А. В. Борисов a, Т. С. Демкина a, Н. Н. Каширская a, Т. Э. Хомутова a, Е. В. Чернышева a, *
a Институт физико-химических и биологических проблем почвоведения РАН –
обособленное подразделение ФИЦ ПНЦБ РАН
142290 Пущино, Московской области, ул. Институтская, 2, Россия
* E-mail: chernysheva1988@gmail.com
Поступила в редакцию 20.06.2020
После доработки 23.12.2020
Принята к публикации 28.12.2020
Аннотация
Развивается концепция биологической памяти почв и культурных слоев археологических памятников. Под этим термином предлагается понимать информацию об условиях почвообразования в прошлом, носителями которой являются живые организмы, их генеративные и покоящиеся формы, отмершие и минерализованные организмы и ткани, биоорганические соединения, низко- и супрамолекулярные продукты микробной трансформации органического вещества, а также следы и продукты жизнедеятельности живых организмов в почвенном профиле. В общем виде рассмотрена структура биологической памяти и более детально показаны механизмы функционирования микробной памяти почв как способности микробного сообщества почв сохранять изменения своей структуры, функционального разнообразия и биологической активности, возникшие в результате действия природных или антропогенных факторов в прошлом. Показано, что изменения климатических условий отражаются в микробной памяти погребенных почв в виде изменения биомассы и эколого-трофической структуры почвенного микробного сообщества. Применительно к культурным слоям поселений и почвам со следами древнего антропогенного преобразования, наряду с микробной памятью, реализуется ферментная память, позволяющая реконструировать поступление в почву неспецифичных для нее субстратов, в первую очередь субстратов антропогенной природы. В микробной памяти их поступление в почву сохраняется в виде увеличения численности микроорганизмов, специализирующихся на разложении данного субстрата. В ферментативной памяти – в виде увеличения активности экзо- и эндоферментов, которые участвуют в утилизации этих субстратов почвенным микробными микроорганизмами. Установленные на сегодняшний день временные масштабы функционирования микробной и ферментной памяти почв составляют до нескольких тысяч лет.
ВВЕДЕНИЕ
Концепция памяти почв была сформулирована в конце 70-х–начале 80-х годов прошлого века [44] и является одной из наиболее ярких и продуктивных идей современного этапа отечественного почвоведения. Память почв рассматривается как “…способность почвенной системы запоминать, записывать в своих устойчивых свойствах информацию об условиях (факторах) и процессах своего формирования и дальнейшего изменения во времени” [41]. Следует отметить, что эта мысль, так или иначе, присутствовала на всех этапах развития почвоведения, начиная от известного выражения В.В. Докучаева “почва – зеркало ландшафта” до почвенно-генетической триады факторы–процессы–свойства. Но только в последние десятилетия XX в. идея о сохранении в почве информации об условиях предыдущего этапа развития почвы была оформлена на понятийном и терминологическом уровнях [41, 42, 45].
В качестве носителей на самом низком уровне почвенной памяти выступают отдельные аморфные соединения, кристаллические минералы, наночастицы. Затем следуют уровни органо-минералов, агрегатов, горизонтов, почвенного профиля и почвенного покрова отдельных ландшафтов. Завершает этот перечень носителей почвенной памяти педосфера Земли [42].
При анализе потенциальных носителей почвенной памяти биологическая составляющая почвы долгое время не получала заслуженного внимания. Это относится как к первым работам автора концепции памяти почв, так и к более поздним публикациям. Так, в работе Таргульяна и Бронниковой при глубоком анализе почвенной памяти о физико-химических процессах почвообразования, биогенные носители почвенной памяти упоминаются в числе признаков, генетически не связанных напрямую с почвенными процессами [43]. В их числе авторы упомянули углистые частицы, семена, раковины моллюсков, пыльцу, фитолиты, спикулы губок, микроводоросли, споры грибов и бактерий, н-алканы, лигнины, липиды, жирные кислоты и изотопный состав почвенного органического вещества. Какой-либо попытки систематизации этих весьма разнообразных носителей почвенной памяти и определения их иерархической позиций авторы не предпринимают.
В то же время есть все основания полагать, что биологические носители почвенной памяти потенциально могут функционировать на всех иерархических уровнях организации почвы, начиная с внутриагрегатного уровня. Именно на этом иерархическом уровне зерна почвенного скелета и органо-минеральные соединения ассоциируются в сложные и динамические соединения, в которых глинистые минералы выступают в роли средообразующего фактора для биологической составляющей – почвенных микроорганизмов [25]. Поэтому можно говорить о том, что на всех иерархических уровнях выше внутриагрегатного потенциально существует возможность записи информации об условиях почвообразования на биологических носителях почвенной памяти. Соответственно, информационный потенциал этого типа памяти практически неограничен.
К настоящему времени из всех потенциально многообразных биогенных носителей информации об условиях почвообразования в прошлом достаточно полно исследованы лишь “микробиоморфная”, “гумусовая” и “микологическая” памяти почв. В первом случае в качестве носителей почвенной памяти выступают фитолиты, споры, пыльца, угли, детрит и иные остатки растительного происхождения [14–16]. В основе “гумусовой” памяти почв лежит его способность сохранять ряд свойств в геологических масштабах времени и адекватно отражать особенности природной среды периода своего формирования. Информация о природных условиях прошлых эпох сохраняется на уровне состава, структурных особенностей и свойств гуминовых кислот и фульвокислот, синтез которых зависит от условий влажности и температуры [21, 60]. Исследование системы гумусовых веществ в почвах привело к формированию концепции ее супрамолекулярности, которая в настоящее время активно развивается и обсуждается [47].
Первым шагом в раскрытии информационной емкости микробного сообщества почвы как носителя почвенной памяти оказались исследования микобиоты почв и введение в научный оборот понятия “микологической” памяти почв [27, 32, 33]. Для микологической памяти почв характерны два аспекта: наряду со способностью хранить информацию о смене климатических условий, данный пласт почвенной памяти несет сведения об особенностях антропогенного влияния на природную среду в прошлом. Свидетельство о вековой динамике климата проявляется в том, что грибное сообщество представляет собой один из основных компонентов большинства наземных экосистем; он отражает свойства почв и растительного покрова на каждом участке ландшафта, благодаря закономерной изменчивости собственных характеристик, чувствительных к изменениям остальных компонентов экосистемы [32]. Здесь нельзя не упомянуть многолетние работы Демкиной, Хомутовой, Каширской и ряда других микробиологов, в которых микробное сообщество погребенных почв рассматривалось с этих же позиций, при этом были задействованы другие носители почвенной памяти, такие как микробная биомасса и структура микробного сообщества [18, 29, 49, 50].
Второй аспект микологической памяти, освещенный в работах Марфениной и Ивановой, открыл принципиально новый пласт почвенной памяти, а именно – реконструкцию поступления в почву органических субстратов антропогенной природы в древности и средневековье [66]. Так как при разложении разных субстратов могут участвовать специфические виды грибов, становится возможным по изменению структуры почвенной микобиоты и увеличению доли отдельных видов грибов реконструировать особенности поступления в почву того или иного органического материала [27, 34, 35, 70]. В частности, исходное поступление в почву шерсти, пера, кожи удалось реконструировать по обилию кератинолитических грибов [27, 32].
На основе этих двух аспектов микологической памяти почв, Марфенина предложила термин более высокого иерархического уровня – “биотическая память почв” [33]. Впоследствии весьма плодотворная тема биотической памяти почв не получила достаточно глубокого развития. В этой связи целью данной работы является развитие идей О.Е. Марфениной о биологическом компоненте феномена почво-памяти и обобщение авторских данных о двух ее составляющих – микробной и ферментной памяти почв.
“БИОТИЧЕСКАЯ” ИЛИ “БИОЛОГИЧЕСКАЯ” ПАМЯТЬ
Развитие в науке предполагает не только открытие чего-то нового, но и уточнение, детализацию и усовершенствование уже имеющегося, известного ранее. Именно в этом ключе следует рассматривать наше предложение по введению в научный оборот термина “биологическая память”. При общей и очевидной близости терминов “биологический” и “биотический” в строгом смысле их нельзя рассматривать как синонимы. Например, трудно заменить устоявшиеся понятия биологическая активность, биологическая емкость и др. новыми терминами с корнем “биота”. Кроме того, термин “биотическая память почв” можно интерпретировать как память почвенной биоты, что в значительной мере сужает многообразие биогенных носителей почвенной памяти и оставляет за пределами рассмотрения следы их жизнедеятельности, сохранившиеся в минеральной массе. На наш взгляд, термин “биологический”, будучи более высокого иерархического ранга, более широк и нейтрален. Да и применительно к минерализованным организмам и их дериватам как носителям почвенной памяти термин “биотический” подходит в меньшей степени.
Поэтому нам представляется, что термин “биологическая” память почв будет в большей степени соответствовать действительности и охватывать все потенциальные биогенные носители почвенной памяти.
СТРУКТУРА БИОЛОГИЧЕСКОЙ ПАМЯТИ
В работах Марфениной были заданы общие контуры биологической памяти, внутри которой было предложено выделять: “…1 – “абсолютную память” – палеоданные о флоре и фауне прошлого, аналогичные документальным историческим фактам; 2 – “модифицированную” память, которая может быть несколько искажена со временем … и 3 – факты, стертые из памяти, то есть отмершие и разложившиеся организмы…” [33]. Учитывая новые данные, эта структурная схема может быть расширена и дополнена.
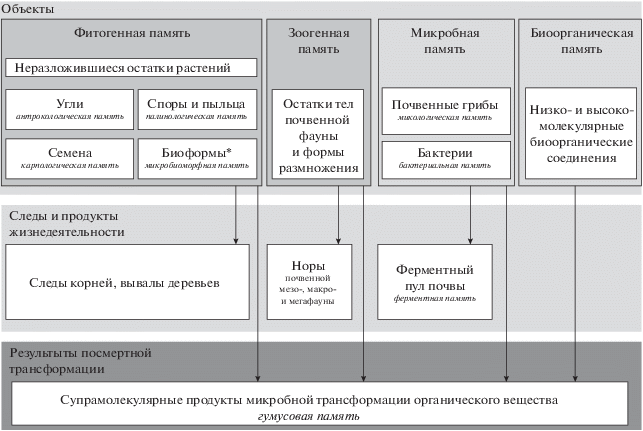
Развивая идеи О.Е. Марфениной и сохраняя предлагаемый ею принцип разделения носителей биологической памяти, мы предлагаем несколько иной механизм разделения, в основу которого будет взят тип объекта – носителя биологической памяти, а также результаты его жизнедеятельности и последующих преобразований. В этой связи первый уровень носителей биологической памяти будет логичнее назвать “объектным”.
Объектный уровень биологической памяти почвы можно разделить на фитогенную, зоогенную и микробную память в соответствие с жизненными формами ее носителей, которые могут присутствовать в почве и нести информацию об условиях почвообразования в прошлом.
В блок фитогенной памяти входят следующие носители информации о природно-антропогенных событиях прошлых эпох:
– неразложившиеся остатки растений, торфяные горизонты (собственно фитогенная память);
– обугленные остатки высших растений (антрокологическая память) [9, 54, 55];
– споры и пыльца растений (палинологическая память) [63, 74, 76, 77, 83, 84];
– семена растений (карпологическая память) [40];
– биоморфы и непалиноморфы (включая водоросли) (микробиоморфная память) [15, 46, 62, 65, 81, 87].
Остатки почвенной мезо-, макро- и мегафауны образуют блок зоогенной памяти [1, 22, 24].
Почвенные бактерии и грибы формируют блок микробной памяти. В некоторых случаях для этой группы уместно ввести более дробное подразделение на эукариоты (“микологическая” память) [27, 32, 33] и прокариоты (“бактериальная” память). Этот блок микробной памяти почв представлен в работах Демкиной, Хомутовой, Каширской и других исследователей [20, 29, 48].
Закрывает объектный уровень биологической памяти блок низко- и высокомолекулярных биоорганических соединений, в который входят алканы, лигнин, жирные кислоты и другие соединения, которые, попадая в почву из живого организма, не изменяются вовсе, либо изменяются незначительно (деградируют в соответствии с известными химическими законами).
Живые организмы, жизненный цикл которых связан с почвой, являются агентами, способными преобразовывать почву и верхний слой почвообразующей породы, изменяя их морфологические и химические свойства. В этом проявляется еще один уровень “результатов деятельности организмов”, который объединяет сохранившиеся в почвенном профиле следы и продукты жизнедеятельности носителей объектной памяти. В зависимости от их жизненной формы на этом уровне можно рассматривать ходы корней [5], нарушенное залегание почвенных горизонтов или их отсутствие под вывалами деревьев [9], норы почвенной мезо-, макро- и мегафауны [22, 23], а также изменение ферментного пула почвы [28, 53, 59].
Третьим уровнем биологической памяти является уровень супрамолекулярных продуктов микробного разложения отмерших организмов и продуктов их жизнедеятельности – “гумусовая память” (по Дергачевой [21]), отражающая условия почвообразования, существовавшие во время жизни биогенных носителей почвенной памяти и процессы их последующего преобразования.
Схема структуры биологической памяти приведена на рис. 1. Раскрыть в деталях каждый блок этой схемы – задача, выходящая за пределы данной работы. Подобные вопросы мы оставляем компетенции специалистов в каждой области исследований, представленной на схеме. Есть основания надеяться, что настоящая статья откроет дискуссию по этой весьма перспективной, сложной и многогранной теме. Мы же представили схему в первую очередь для того, чтобы показать место в общей системе биологической памяти одной из ее составляющих – микробной памяти и ее производной – ферментной памяти почв. При этом под термином “микробная память” мы предлагаем понимать способность микробного сообщества почв сохранять изменения своей структуры, функционального разнообразия и биологической активности, возникшие в результате действия природных или антропогенных факторов в прошлом.
ОБЪЕКТЫ И МЕТОДЫ
В качестве объектов для изучения микробной памяти почв безусловно лучшими являются почвы археологических памятников – естественные ненарушенные подкурганные погребенные почвы [18, 51, 56, 69, 73], с одной стороны, и культурные слои древних поселений, антропогенно-преобразованные почвы возле археологических памятников и специфические почвоподобные тела как результат максимальной антропогенной трансформации исходной почвы [57, 58, 64, 72, 93].
В первом случае в распоряжении исследователей оказывается микробное сообщество, отражающее состояние природных условий на момент сооружения кургана. После создания курганной насыпи поступление растительных остатков в погребенную почву прекращалось практически полностью, за исключением единичных магистральных корней кустарничковых растений при высоте насыпи менее 1 м. Влагообмен с насыпью осуществлялся только в испарительном режиме [17], годовой ход температуры и влажности минимальный. Это, с учетом диагенетических изменений, дает возможность получить весьма репрезентативную картину состояния микробного сообщества на момент сооружения кургана. При этом диагенетические изменения отражаются, в первую очередь, на количественных показателях почвенного микробного сообщества и в меньшей степени затрагивают его структуру. К настоящему времени наиболее полно исследован информационный потенциал микробных сообществ подкурганных погребенных почв сухостепной и пустынно-степной зоны в пределах Ергенинской и Приволжской возвышенностей, Прикаспийской низменности и Сальско-Манычской гряды [19, 29, 48, 49].
Если в случае с подкурганными почвами в распоряжении исследователя оказывается почвенное тело с минимальными антропогенными нарушениями, то культурные слои древних поселений представляют собой обратную ситуацию – это результат максимального антропогенного преобразования почвы вследствие хозяйственной деятельности человека в прошлом. Культурный слой можно рассматривать как многолетний эксперимент, заложенный сотни и тысячи лет назад, в котором в точно известный момент времени, на точно известной территории в почву поступали определенные органические субстраты. Набор их довольно ограничен – остатки пищи, строительные материалы и бытовой мусор растительной и животной природы, фекалии, навоз. Результат их минерализации обусловил сегодняшнее состояние микробного сообщества культурных слоев. Антропогенные изменения микробного сообщества были изучены в культурных слоях поселений эпохи бронзы и раннего средневековья на территории Кисловодской котловины [52, 53, 57–59, 80], в лесостепной зоне Подонья [13, 37], в центральной части Восточно-Европейской равнины [29] и степном Предкавказье [67].
При изучении эколого-трофической структуры почвенного микробного сообщества и численности отдельных групп микроорганизмов использовали метод посева и учета КОЕ бактерий и грибов на селективных питательных средах [18, 58]; биомассу живых клеток определяли по содержанию фосфолипидов в почве [49]; активность ферментов по общепринятым методикам [29, 59].
Рассмотрим несколько примеров функционирования микробной памяти почв и культурных слоев древних поселений.
ОТРАЖЕНИЕ В МИКРОБНОЙ ПАМЯТИ ПОЧВ ИЗМЕНЕНИЙ ПАЛЕОКЛИМАТИЧЕСКИХ УСЛОВИЙ
Микробные сообщества подкурганных почв попали в поле зрения исследователей довольно давно. Уже в середине 1970-х появились первые работы, где исследователи обращают внимание на микробные характеристики погребенных почв [26, 31, 39]. Было показано, что в погребенных почвах обнаружены живые микроорганизмы, и что их численность заметно меньше по сравнению с современными фоновыми аналогами. Одной из первых работ в этом направлении стала публикация Демкиной с соавт. [18], в которой было установлено, что структура и свойства микробного сообщества палепочвы в целом отражают климатические условия, существовавшие в период незадолго до сооружения кургана.
Сохранению почвенных микроорганизмов на протяжении неопределенно долгого времени способствовали их адаптационные механизмы выживания (например, анабиоз, переход бактерий в наноформы и др.) [11, 12]. В частности, было выявлено, что в горизонте А1 каштановой почвы эпохи бронзы, погребенной около 5000 лет назад, до 80% клеток относятся к наноформам, с объемом клеток на уровне 0.09 мкм3. В аналогичной современной почве таких клеток насчитывалось не более 60% [29]. Это подтверждает гипотезу, что формирование наноформ представляет собой универсальную ответную реакцию организма на стресс и неблагоприятные внешние условия [12].
В результате сопряженного анализа химических свойств погребенных степных почв и их биологической активности разработаны микробиологические индикаторы аридных и гумидных условий почвообразования в прошлом. Это биомасса микроорганизмов и эколого-трофическая структура микробного сообщества (соотношение микроорганизмов, растущих на почвенном агаре (ПА) и использующих элементы питания из рассеянного состояния: микроорганизмов, растущих на нитритном агаре (НА) и потребляющие гумус, и микроорганизмов, растущих на богатой среде (БС) и разлагающих растительные остатки), а также индекс олиготрофности (ПА : БС × 100), отражающий способность микробного сообщества потреблять зольные элементы питания из рассеянного состояния, является индикатором неблагоприятных природных условий [19, 20].
Почвенно-микробиологический подход к реконструкции годового хода осадков. Сопоставление характеристик микробных сообществ с химическими свойствами погребенных почв позволило дать оценку внутригодового хода осадков [68]. Известно, что для аридных периодов характерны низкая биомасса живых клеток (рассчитанная по содержанию фосфолипидов в почве [49]), высокий индекс олиготрофности микробных сообществ и уменьшение доли микроорганизмов, растущих на БС. При этом палеопочвы характеризовались засоленностью почвенного профиля, высоким содержанием карбонатов и пр. И наоборот, в гумидные периоды возрастала микробная биомасса, уменьшался индекс олиготрофности на фоне рассоления почвенного профиля. Подобного рода ситуация с синхронным уменьшением количества зимних и летних осадков, установлена для раннего железного века в сухостепной зоне [18, 20].
При изучении палеопочв Волго-Донского междуречья, погребенных в середине III тыс. до н. э., была вскрыта необычная закономерность асинхронного изменения физико-химических и микробиологических параметров. Микробные характеристики указывали на наличие гумидного периода – количество живой биомассы значительно превышало фоновые показатели (в 2 раза и более), доля живой биомассы в органическом углероде погребенных почв была в 3 раза выше современного уровня. Такие же закономерности были выявлены для соотношения групп микроорганизмов, растущих на богатой и бедной среде (БС : НА). При этом индекс олиготрофности в погребенных почвах был меньше такового в их фоновых аналогах [48]. А химические свойства демонстрировали резкую аридизацию (близкое к поверхности залегание аккумуляции легкорастворимых солей и гипса, отсутствие сегрегационных форм карбонатных новообразований, низкая магнитная восприимчивость и др.). В результате комплексного анализа этой ситуации впервые в практике палеоэкологических исследований был поставлен вопрос о возможности реконструкции годового хода осадков.
Как известно, в степной зоне химические свойства почв определяются условиями влагообеспеченности зимнего периода. Уменьшение глубины вскипания, вымывание водорастворимых солей, формирование сегрегационных форм карбонатных новообразований, накопление гумуса, увеличение мощности гумусового горизонта, увеличение магнитной восприимчивости в верхних горизонтах – иными словами, все признаки гумидных условий почвообразования проявляются при увеличении влагообеспеченности холодного времени года. Влияние зимних осадков на водный режим степных почв ранее неоднократно был показано многими исследователями [7, 8, 30, 47, 88].
Осадки, выпадающие в теплое время года, традиционно принято считать неэффективными [38], так как в условиях сухостепной зоны они промачивают почву на глубину не более 30 см, а в пустынно-степной зоне глубина промачивания светло-каштановых почв и солонцов в летний период еще меньше [7, 30]. В этом случае можно говорить о том, что влияние летних осадков на химические свойства почв незначительно [7].
В то же время для микробного сообщества почв летние осадки имеют большое значение, вызывая увеличение массы отавы и фитомассы в целом [2], особенно молодых растений [6]. Таким образом, в степной зоне частые летние осадки, практически не оказывающие влияния на солевой режим почвы, способны вызывать существенное увеличение поступления в почву растительных остатков, что находит свое отражение в структуре микробного сообществ почв.
При анализе полученных закономерностей был сформирован новый методический подход для реконструкции годового хода осадков. Суть его заключается в сопряженном анализе химических свойств почв и их биологической активности. При этом исходили из того, что динамика химических свойств (в первую очередь, профильное содержание солей) в большей степени обусловлена влагообеспеченностью холодного времени года, в то время как биомасса и трофическая структура микробных сообществ почв отражают условия для вегетации растений и развития почвенной микробиоты, и в большей степени зависят от влагообеспеченности теплого периода года [68].
Благодаря этому подходу удалось раскрыть феномен суббореальной аридизации в степи. Ранее на основе анализа химических свойств погребенных почв неоднократно указывалось на резко выраженные аридные условия в середине – второй половине III тыс. до н. э., с пиком аридизации на рубеже III–II тыс. до н. э. [4, 10], но огромное количество курганов, созданных в тот период, указывает на довольно плотное заселение степи. Этот парадокс долгое время оставался без ответа. Ситуация прояснилась лишь после проведения анализа микробиологических свойств почв и реконструкции влагообеспеченности теплого времени года. Полученный вывод позволил иначе взглянуть на аридизацию – ключевое понятие в палеоэкологии степной зоны. Стало очевидно, что в условиях степной зоны аридизация климата, диагностируемая по комплексу химических свойств погребенных почв, это прежде всего уменьшение нормы зимних осадков как следствие усиление зимнего антициклона. То есть в зимний период устанавливалась холодная сухая погода при малой мощности снегового покрова, либо его полном отсутствии. В этих условиях все химические и морфологические свойства почв показывали резкое усиление засушливости.
Если же в аридных почвах возрастала биомасса почвенных микроорганизмов, а в структуре микробного сообщества заметно увеличивалась доля клеток, использующих легкодоступное органическое вещество, то эти признаки указывали на периодические летние осадки, которые предотвращали выгорание степи и способствовали большему поступлению в почву растительных остатков [68].
Эта гипотеза позволила согласовать химические и микробиологические характеристики погребенных в середине III тыс. до н. э. палеопочв и объяснить высокую насыщенность региона памятниками этого времени. Холодные сухие зимы обеспечивали благоприятные условия для зимнего выпаса скота, при этом осадки в весенне-летний период поддерживали продуктивность фитоценозов и делали пустынные степи весьма привлекательными для древних скотоводов [68].
Таким образом, влагообеспеченность летнего периода выступает как “фактор” в известной триаде фактор → процесс → свойство, указывая на возрастание поступления фитомассы (“процесс”), масштабы которого можно установить с помощью анализа эколого-трофической структуры микробного сообщества почв (“свойство”).
ОТРАЖЕНИЕ В БИОЛОГИЧЕСКОЙ ПАМЯТИ ПОЧВ ПОСЛЕДСТВИЙ АНТРОПОГЕННОЙ ДЕЯТЕЛЬНОСТИ
В микробной памяти наиболее четко сохраняется информация о поступлении в почву органических материалов антропогенной природы, которые обычно не характерны для почв (например, остатки пищи белковой и липидной природы, мочевина, кератин и др.), либо поступление которых в естественных условиях было незначительным. Как только тот или иной участок почвы становится местом длительного и компактного проживания крупного коллектива людей, в почву начинают поступать органические субстраты, неспецифические для данных почв. Такого рода неспецифичные органические материалы, оказавшись в почве, разлагаются с участием почвенных микроорганизмов, что приводит к существенной перестройке структуры и функционального разнообразия почвенного микробного сообщества. Здесь следует отметить признанный приоритет российских исследователей в этой области [71]. Развивая идеи об отражении в почвенном микробном сообществе антропогенного фактора, мы предлагаем выделить 2 аспекта биологической памяти – микробную и ферментативную память почв. Остановимся более подробно на этих вопросах.
Микробная память. При поступлении в почву неспецифического для нее органического субстрата происходит увеличение численности микроорганизмов, специализирующихся на его разложении. При наступлении неблагоприятных условий почвенные микроорганизмы переходят в неактивное состояние, в котором они могут находиться неопределенно длительное время, что возможно благодаря различным формам выживания микроорганизмов [78, 82, 89]. Часть этого микробного пула может сохранять способность образовывать колонии при посеве на чашках Петри при стандартных условиях, что делает возможным их видовую идентификацию [85]. Таким образом, повышенная численность КОЕ определенных групп микроорганизмов, может указывать на поступление в прошлом определенного субстрата, что послужило причиной всплеска их численности в почве. В настоящие время этот механизм биологической памяти почв исследован лишь на примере сообщества почвенных микроскопических грибов, но уже очевидно, что по такому же принципу будет работать и бактериальная память почвы.
Различные формы антропогенной деятельности приводят к появлению в почве различных неспецифических для нее микроорганизмов. В частности, носителями информации о древнем сельскохозяйственном использовании почв оказались термофильные микроорганизмы. В почвах численность этой группы микроорганизмов может существенно изменяться лишь в результате сельскохозяйственного освоения территории [36]. Показано, что наиболее вероятные источники поступления этих микроорганизмов в почву – это навоз и компост, которые вносятся на поля в качестве органических удобрений. Разложение органических материалов сопровождается экзотермическими реакциями. На этой стадии получают преимущества термофильные микроорганизмы, адаптированные к размножению и развитию при высоких температурах (более 60°С). По мере исчерпания субстрата активность термофильных микроорганизмов уменьшается, и температура компоста начинает снижаться [67]. В этот момент начинается активный процесс спорообразования. Споры термофильных микроорганизмов в дальнейшем могут попадать в почву и сохраняться в ней неопределенно долгий период времени. Выявлена высокая численность КОЕ термофильных микроорганизмов в почвах вблизи средневековых поселений (V–VIII вв. н. э.), что указывает на длительное использование органических удобрений, что хорошо подтверждается обилием керамики в почвах [91]. По мере удаления от памятника происходило существенное уменьшение их численности [58]. Кроме того, высокая численность термофилов была выявлена в почвах внутри древних каменных загонов для скота, что указывает на длительную аккумуляцию навоза.
Впервые нами был проведен молекулярно-генетический анализ выделенных чистых культур термофильных микроорганизмов из почв земледельческих террас и загонов для скота [52]. Анализ последовательности гена 16S рРНК показал, что 2 штамма, выделенных из слоя почвы земледельческой террасы V–VII века, относятся к спорообразующим, умеренно термофильным бактериям рода Anoxybacillus. Из средневекового слоя почвы загона выделен штамм, относящийся к роду Parageobacillus облигатных термофильных спорообразующих бактерий. Из этого же образца выделен штамм, относящийся к роду термофильных актиномицетов Thermoactinomyces.
В рассмотренном примере функционирования микробной памяти триаду фактор → процесс → → свойство можно представить в следующем виде – антропогенная деятельность → поступление в почву компостированных материалов → высокая численность термофильных микроорганизмов.
Аналогичный алгоритм функционирования микробной памяти был разработан для изучения особенностей погребального обряда. В этом случае объектом исследований явился почвенно-грунтовый материал, содержащий следы микробной трансформации одежды погребенных, деталей убранства погребального ложа и иных составляющих погребального комплекса, которые могли содержать кератин [90]. При реконструкции одежды погребенных из захоронений эпохи бронзы и античного времени показана перспективность использования такого показателя, как обилие кератинолитических грибов [67]. Этот подход был апробирован на грунтах из погребений эпохи бронзы могильника “Бейсужек-35” (Краснодарский край). Так, в частности, высокие значения численности КОЕ грибов на среде с кератином при посеве суспензии из образцов грунта, отобранного под ступнями погребенных, являются свидетельством исходного присутствия обуви из шерсти [67].
Ферментная память. Поступление в почву любого органического субстрата приводит к изменению количественных и качественных показателей микробного сообщества почв, в частности к изменению ферментативной активности. В процессе разложения микроорганизмами органических остатков происходит выделение в почву ферментов, которые могут сохраняться в почве без потери их активности на протяжении неопределенно длительного периода времени [58, 75, 86]. Идеальным объектом исследования этого вида почвенной памяти являются культурные слои и почвы археологических памятников. Они могут рассматриваться в качестве уникального почвенного образования, в которое на протяжении длительного времени в прошлом поступали определенные органические материалы (мочевина, белки, жиры, крахмал, целлюлоза и др.), связанные с хозяйственной деятельностью человека. Попадая в почву, эти органические остатки вызывали рост численности микроорганизмов, участвующих в их разложении, которые, в свою очередь, выделяли специфические ферменты, ответственные за гидролиз того или иного органического субстрата. В результате определенные группы ферментов накапливались в почвах и культурных слоях. Такие изменения энзиматического пула, имевшие место в древности, могут сохраняться до наших дней, образую ферментную память почв.
Данный вид почвенной памяти практически не исследован, хотя его потенциал представляется весьма внушительным. Известна работа, в которой показано, что в почве древней земледельческой террасы (около 1500 л. н.) сохранился высокий уровень фосфатазной и амидазной активности, превышающий уровень ферментативной активности современных целинных и пахотных почв [61]. Изучение информационного потенциала ферментативной памяти почв было начато с определения уреазной активности культурных слоев древних поселений [11, 80]. Установлено существенное увеличение уреазной активности в почвах в окрестностях средневековых поселений (V–VIII вв. н. э.) в зоне предполагаемой распашки с внесением органических удобрений [53, 59]. Использование показателя уреазной активности оказалось удачным и для выявления зоны интенсивной хозяйственной деятельности, а также для уточнения ее характера в связи с расположением конкретных сооружений и общей планировкой археологического памятника [13].
Еще одним ферментом, перспективность использования которого при реконструкциях изменений условий почвообразования хорошо доказана, является фосфатаза. На настоящий момент проведены исследования фосфатазной активности культурных слоев древних поселений и почв, погребенных под курганами эпохи бронзы и раннего железного века в степной зоне юга России. Показано, что высокий уровень фосфатазной активности, соответствующий показателям современных почв, наблюдается в культурных слоях возрастом более 4000 лет, причем отмечены пики фосфатазной активности, связанные с особенностями формирования культурного слоя в различные периоды функционирования поселения [28, 37].
Длительность функционирования ферментной памяти для разных групп ферментов неодинакова. Ранее было выявлено, что в погребенных почвах и культурных слоях поселений активность фосфатаз сохраняется в образцах возрастом более 5000 лет на уровне, превышающем фоновый в современных почвах [28]. Активность уреазы в образцах такого возраста заметно меньше фоновых показателей; можно говорить о том, что этот фермент сохраняет активность до 2–3 тыс. лет [57].
Механизм функционирования ферментной памяти следующий: признак (высокая активность фермента) является следствием процесса (накопления фермента при разложении больших объемов специфических субстратов антропогенной природы) и представляет собой результат влияния на почву антропогенной деятельности как фактора почвообразования.
ЗАКЛЮЧЕНИЕ
Биологическая память почв – это информация об условиях почвообразования и антропогенной деятельности в прошлом, носителями которой являются живые организмы, их генеративные и покоящиеся формы, отмершие и минерализованные организмы и ткани, биоорганические соединения, низко- и супрамолекулярные продукты микробной трансформации органического вещества, а также следы и продукты жизнедеятельности живых организмов в почвенном профиле.
Биологическая память реализуется на трех информационных уровнях: 1) на уровне собственно объектов – носителей почвенной памяти; 2) на уровне следов и продуктов их жизнедеятельности в почве; 3) на уровне продуктов посмертной микробной трансформации их тел. В зависимости от жизненной формы носителей биологической памяти объектный уровень объединяет фитогенную, зоогенную и микробную память, а также информацию в виде низко- и высоко молекулярных биоорганических соединений, ранее входивших в состав живых организмов и поступивших в почву после их отмирания без изменений.
Второй уровень объединяет результаты жизнедеятельности бактерий и грибов, оставленные ими в минеральной составляющей почвы и органо-минеральных комплексах. Следы их функционирования сохраняются в виде ходов корней, вывалов деревьев, нор почвенной мезо- и мегафауны, а также в виде эндо- и экзоферментов, которые были синтезированы почвенными микроорганизмами в прошлом.
Третьим уровнем биологической памяти является уровень супрамолекулярных продуктов микробной трансформации самих организмов и продуктов их жизнедеятельности – “гумусовая память”, отражающая условия почвообразования во время жизни биогенных носителей почвенной памяти и процессы их последующего преобразования.
Микробная память почв является важной составляющей феномена почвенной биологической памяти. Под термином “микробная память” мы предлагаем понимать способность микробного сообщества почв сохранять изменения своей структуры, функционального разнообразия и биологической активности, возникшие в результате действия природных или антропогенных факторов в прошлом.
Наибольший информационный потенциал микробной памяти почв раскрывается при изучении процессов поступления и преобразования в почве органических субстратов естественной и антропогенной природы. Это позволяет реконструировать динамику природных условий и разные формы бытовой и производственной деятельности человека в прошлом.
Отражение в микробной почвенной памяти информации о климатических флуктуациях происходит в виде изменения биомассы почвенного микробного сообщества и его эколого-трофической структуры. Этот аспект почвенной памяти наиболее полно реализуется в случае с подкурганными погребенными почвами. В этом случае триаду фактор → процесс → свойство можно представить следующим образом: климатические изменения → поступление в почву отмершей фитомассы → изменения биомассы и эколого-трофической структуры микробного сообщества. Использование микробиологических методов изучения погребенных почв позволило вывести палеоэкологические реконструкции на качественно новый уровень и перейти к реконструкциям годового хода осадков.
Информация о поступлении органических субстратов антропогенной природы сохраняется в биологической почвенной памяти в виде увеличения численности микроорганизмов, специализирующихся на разложении данного субстрата (микробная память), а также в увеличении активности соответствующих ферментов, участвующих в процессе минерализации (ферментная память почв). И в том, и в другом случаях резкое возрастание численности определенной группы почвенных микроорганизмов или увеличение активности фермента является следствием поступления и разложения в почве специфических субстратов антропогенной природы и отражает влияние на почву антропогенной деятельности как фактора почвообразования.
Микробная и ферментная память как составляющие биологической памяти почв позволяют реконструировать почвенные процессы, связанные с антропогенной деятельностью, которые не могут быть реконструированы с помощью других методов.
Список литературы
Абатуров Б.Д. Млекопитающие как компонент экосистем. М.: Наука, 1984. 288 с.
Абрамова М.М. К вопросу об эффективности летних осадков в условиях в условиях засушливого климата // Почвоведение. 1962. № 9. С. 4–53.
Александровский А.Л., Александровская Е.И. Эволюция почв и географическая среда. М.: Наука, 2005. 223 с.
Александровский А.Л., Бронникова М.А., Водяницкий Ю.Н., Герасимова М.И. и др. Память почв. Почва как память биосферно-геосферно-антропосферных взаимодействий. М.: Изд-во ЛКИ, 2008. 690 с.
Алексеева Т.В., Алексеев А.О., Калинин П.И. Палеопочвы нижнего карбона в карьере “Бронцы” (Калужская область) // Почвоведение. 2018. № 7. С. 787–800.
Алпатьев А.М. Почвоувлажнительный и биологический эффект атмосферных осадков // Почвоведение. 1959. № 2. С. 12–14.
Афанасьева Е.А. Водно-солевой режим обыкновенных и южных черноземов Юго-Востока европейской части СССР. М.: Наука, 1980. 216 с.
Базыкина Г.С. Водный режим и баланс мелиорируемых в культурных биогеоценозах // Биогеоценотические основы освоения полупустыни Северного Прикаспия. М.: Наука, 1974. 359 с.
Бобровский М.В. Лесные почвы Европейской России: биотические и антропогенные факторы формирования. М.: КМК, 2010. 359 с.
Борисов А.В., Мимоход Р.А. Аридизация: формы проявления и влияние на население стеной зоны в бронзовом веке // Российская археология. 2017. № 2. С. 48–60.
Бухарин О.В., Гинцбург А.Л., Романова Ю.М., Эль-Регистан Г.И. Механизмы выживания бактерий. М.: Медицина, 2005. 367 с.
Вайнштейн М.Б., Кудряшова Е.Б. О наннобактериях // Микробиология. 2000. Т. 69. № 2. С. 163–174.
Гак Е.И., Чернышева Е.В., Ходжаева А.К., Борисов А.В. Опыт выявления и систематизации инфраструктурных признаков поселения катакомбной культуры Рыкань-3 в лесостепном Подонье // Российская археология. 2014. № 4. С. 19–28.
Гольева А.А. Микробиоморфная память почв // Память почв: Почва как память биосферно-антропосферных взаимодействий. М.: Изд-во ЛКИ, 2008. С. 500–529.
Гольева А.А. Микробиоморфные комплексы природных и антропогенных ландшафтов: генезис, география, информационная роль. М.: Изд-во ЛКИ, 2008. 238 с.
Гольева А.А. Фитолиты и их информационная роль при исследовании природных и археологических объектов. М.–Сыктывкар–Элиста, 2001. 200 с.
Демкин В.А. Почвоведение и археология. Пущино: ОНТИ, 1997. 196 с.
Демкина Т.С., Борисов А.В., Демкин В.А. Характеристика микробных сообществ палеопочв археологических памятников пустынно-степной зоны // Почвоведение. 2000. № 9. С. 1117–1126.
Демкина Т.С., Борисов А.В., Демкин В.А., Хомутова Т.Э., Кузнецова Т.В., Ельцов М.В., Удальцов С.Н. Палеоэкологический кризис в степях Нижнего Поволжья в эпоху средней бронзы (рубеж III–II тыс. до н. э.) // Почвоведение. 2017. № 7. С. 799–813.
Демкина Т.С., Борисов А.В., Хомутова Т.Э. Сравнительная характеристика современных и погребенных почвенных комплексов в пустынно-степной зоне Волго-Донского междуречья // Почвоведение. 2019. № 11. С. 1295–1306.
Дергачева М.И. Гумусовая память почв // Память почв: Почва как память биосферно-антропосферных взаимодействий. М.: Изд-во ЛКИ, 2008. С. 530–560.
Динесман Л.Г. Биогеоценозы степей в голоцене. М.: Наука, 1977. 160 с.
Динесман Л.Г. Изучение истории биогеоценозов по норам животных. М.: Наука, 1968. 100 с.
Занина О.Г. Ископаемые норы грызунов из мёрзлых позднеплейстоценовых отложений Колымской низменности // Зоологический журн. 2005. Т. 84. № 6. С. 728–736.
Звягинцев Д.Г. Почва и микроорганизмы. М.: Изд-во Моск. ун-та, 1987. 256 с.
Золотун В.П. Развитие почв юга Украины за последние 50–45 веков. Автореф. дис. … докт. с.-х. н. Киев, 1974. 74 с.
Иванова А.Е., Марфенина О.Е., Кислова Е.Е., Зазовская Э.П. Микологические характеристики культурного слоя средневекового поселения на дерново-карбонатных почвах // Почвоведение. 2006. № 1. С. 62–71.
Каширская Н.Н., Плеханова Л.Н., Чернышева Е.В., Ельцов М.В., Удальцов С.Н., Борисов А.В. Пространственно-временные особенности фосфатазной активности естественных и антропогенно-преобразованных почв // Почвоведение. 2020. № 1. С. 89–101.
Каширская Н.Н., Хомутова Т.Э., Демкина Т.С., Демкин В.А. Микробная биомасса подкурганных и современных почв степной зоны Нижнего Поволжья // Почвоведение. 2009. № 5. С. 581–587.
Ковда В.А., Большаков А.Ф. Водно-солевой режим почв центральной части Каспийской равнины. Труды конференции по почвоведению и физиологии растений. Саратов, 1937. Т. 1. С. 144–166.
Крупеников И.А. Почвенный покров Молдовы: прошлое, настоящее, управление, прогноз. К.: Штиинца, 1992. 265 с.
Марфенина О.Е. Антропогенная экология почвенных грибов. М.: Медицина для всех, 2005. 196 с.
Марфенина О.Е. Свойства почвенной микробиоты как одна из форм биотической памяти почв // Память почв: почва как память биосферно-антропосферных взаимодействий. М.: Изд-во ЛКИ, 2008. С. 561–577.
Марфенина О.Е., Горбатовская Е.В., Горленко М.В. Микологические характеристики культурных слоев почв древнерусских поселений // Микробиология. 2001. Т. 70. № 6. С. 855–859.
Марфенина О.Е., Иванова А.Е., Кислова Е.Е., Зазовская Э.П., Чернов И.Ю. Грибные сообщества почв раннесредневековых поселений таежно-лесной зоны // Почвоведение. 2008. № 7. С. 850–860.
Мишустин Е.Н. Термофильные микроорганизмы в природе и практике. М.–Л.: Изд-во АН СССР, 1950. 391 с.
Потапова А.В., Пинской В.Н., Гак Е.И., Каширская Н.Н., Борисов А.В. Изменчивость свойств культурного слоя поселения эпохи бронзы Ксизово-1 в лесостепной зоне // Российская археология. 2020. № 1. С. 60–75.
Роде А.А. Водный режим и баланс целинных почв полупустынного комплекса // Водный режим почв полупустыни. М.: Изд-во АН СССР, 1963. С. 5–83.
Савостьянова З.А., Нащокин В.Д. К истории почвенного покрова степной зоны Хакасии // Почвенные условия выращивания защитных насаждений. Красноярск, 1974. С. 7–35.
Сергеев А.Ю., Лебедева Е.Ю. К вопросу о раннеаланском земледелии: находки культурных и сорных растений на укреплении Подкумское-2 (II–IV вв. н. э.) // Краткие сообщения института археологии. 2017. № 249-II. С. 311–328.
Таргульян В.О. Некоторые теоретические проблемы почвоведения как науки о Земле // Почвоведение. 1986. № 12. С. 107–116.
Таргульян В.О. Память почв: формирование, носители, пространственно-временное разнообразие // Память почв: почва как память биосферно-антропосферных взаимодействий. М.: Изд-во ЛКИ, 2008. С. 23–57.
Таргульян В.О., Бронникова М.А. Память почв: теоретические основы концепции, современное состояние и перспективы развития // Почвоведение. 2019. № 3. С. 259–275.
Таргульян В.О., Соколов И.А. Структурный и функциональный подход к почве: почва-память и почва-момент // Математическое моделирование в экологии. М.: Наука, 1978. С. 17–33.
Таргульян В.О., Соколова Т.А. Почва как биокосная природная система: реактор, память и регулятор биосферных взаимодействий // Почвоведение. 1996. № 1. С. 34–47.
Темралеева А.Д., Москаленко С.В., Ельцов М.В., Вагапов И.М., Овчинников А.Ю., Гугалинская Л.А., Алифанов В.М., Пинский Д.Л. Устойчивость, морфологическая и молекулярно-генетическая идентификация водорослей погребенных почв // Почвоведение. 2017. № 8. С. 983–991.
Федотов Г.Н., Шалаев В.С. О природе гумусовых веществ //Лесной вестник. 2013. № 7. С. 105–110.
Хомутова Т.Э., Демкин В.А. Оценка биомассы микробных сообществ почв сухих степей по содержанию в них фосфолипидов // Почвоведение. 2011. № 6. С. 748–754.
Хомутова Т.Э., Демкина Т.С., Борисов А.В., Шишлина Н.И. Состояние микробных сообществ подкурганных палеопочв степной зоны эпохи средней бронзы (XXVII–XXVI вв. до н. э.) в связи с динамикой увлажненности климата // Почвоведение. 2017. № 2. С. 239–248.
Хомутова Т.Э., Демкина Т.С., Демкин В.А. Оценка суммарной и активной микробной биомассы разновозрастных подкурганных палеопочв // Микробиология. 2004. Т. 73. С. 241–247.
Чернов Т.И., Железова А.Д., Кутовая О.В., Макеев А.О., Тхакахова А.К., Бгажбa Н.А., Курбанова Ф.Г., Русаков А.В., Пузанова Т.А., Хохлова О.С. Сравнительная оценка структуры микробиомов погребенных и современных почв при помощи анализа микробной ДНК // Микробиология. 2018. Т. 87. № 6. С. 737–746.
Чернышева Е.В., Каширская Н.Н., Демкина Е.В., Коробов Д.С., Борисов А.В. Термофильные микроорганизмы в почвах как результат хозяйственной деятельности человека в древности // Микробиология. 2019. Т. 88. № 5. С. 624–626.
Чернышева Е.В., Каширская Н.Н., Коробов Д.С., Борисов А.В. Биологическая активность дерново-карбонатных почв и культурных слоев аланских поселений Кисловодской котловины // Почвоведение. 2014. № 9. С. 1068–1076.
Bobrovsky M., Kupriaynov D., Khanina L. Anthracological and morphological analysis of soils for the reconstruction of the forest ecosystem history (Meshchera Lowlands, Russia) // Quat. Int. 2019. V. 516. P. 70–82. https://doi.org/10.1016/j.quaint.2018.06.033
Carcaillet C., Thinon M. Pedoanthracological contribution to the study of the evolution of the upper treeline in the Maurienne Valley (North French Alps): methodology and preliminary data // Review of palaeobotany and palynology. 1996. V. 91(1–4). P. 399–416.
Chandler D.P., Brockman F.J., Bailey T.J., Fredrickson J.K. Phylogenetic diversity of archaea and bacteria in a deep subsurface paleosol // Microbial Ecology. 1998. V. 36. P. 37–50.
Chernysheva E., Khomutova T., Fornasier F., Kuznetsova T., Borisov A. Effects of long-term medieval agriculture on soil properties: A case study from the Kislovodsk basin, Northern Caucasus, Russia // J. Mounting Sci. 2018. V. 15(6). P. 1171–1185. https://doi.org/10.1007/s11629-017-4666-7
Chernysheva E., Korobov D., Borisov A. Thermophilic microorganisms in arable land around medieval archaeological sites in Northern Caucasus, Russia: novel evidence of past manuring practices // Geoarchaeology. 2017. V. 32. P. 494–501. https://doi.org/10.1002/gea.21613
Chernysheva E.V., Korobov D.S., Khomutova T.E., Borisov A.V. Urease activity in cultural layers at archaeological sites // J. Archaeological Sci. 2015. V. 57. P. 24–31. https://doi.org/10.1016/j.jas.2015.01.022
Dergacheva M., Bazhina N., Nekrasova O., Zenin V. Shestakovo site of Western Siberia (Russia): Pedogenic features, humic substances and paleoenvironment reconstructions for last 20–25 ka // Quat. Int. 2016. V. 420. P. 199–207. https://doi.org/10.1016/j.quaint.2015.10.087
Dick R.P., Sandor J.A., Eash N.S. Soil enzyme activities after 1500 years of terrace agriculture in the Colca Valley, Peru // Agriculture, Ecosystems & Environment. 1994. V. 50. P. 123–131. https://doi.org/10.1016/0167-8809(94)90131-7
Enevold R., Rasmussen P., Løvschal M., Olsen J., Vad Odgaard B. Circumstantial evidence of non-pollen palynomorph palaeoecology: a 5500 year NPP record from forest hollow sediments compared to pollen and macrofossil inferred palaeoenvironments // Vegetation History and Archaeobotany. 2019. V. 28. P. 105–121. https://doi.org/10.1007/s00334-018-0687-6
Ershova E., Alexandrovskiy A., Krenke N. Paleosols, paleovegetation and neolithic occupation of the moskva river floodplain, Central Russia // Quat. Int. 2014. V. 324. P. 134–145. https://doi.org/10.1016/j.quaint.2014.01.031
Evershed R.P. Organic residue analysis in archaeology: the archaeological biomarker revolution // Archaeometry. 2008. V. 50(6). P. 895–924. https://doi.org/10.1111/j.1475-4754.2008.00446.x
Golyeva A., Svirida N. Quantitative distribution of phytoliths as reliable diagnostical criteria of ancient arable lands // Quat. Int. 2017. V. 434. P. 51–57. https://doi.org/10.1016/j.quaint.2015.12.062
Ivanova A., Marfenina O. Soil fungal communities as bioindicators of ancient human impacts in medieval settlements in different geographic regions of Russia and southwestern Kazakhstan // Quat. Int. 2015. V. 365. P. 212–222. https://doi.org/10.1016/j.quaint.2014.10.016
Kashirskaya N., Kleshchenko A., Mimokhod R., Borisov A. Microbiological approach for identification of wool clothes in ancient burials // J. Archaeological Science: Reports. 2020. V. 31. https://doi.org/10.1016/j.jasrep.2020.102296
Khomutova T.E., Kashirskaya N.N., Demkina T.S., Kuznetsova T.V., Borisov A.V., Fornasier F., Shishlina N.I. Precipitation pattern during warm and cold periods in the Bronze Age (around 4.5–3.8 ka BP) in the desert steppes of Russia: soil-microbiological approach for palaeoenvironmental reconstruction // Quat. Int. 2019. V. 507. P. 84–94. https://doi.org/10.1016/j.quaint.2019.02.013
Makeev A., Aseeva E., Rusakov A., Sorokina K., Puzanova T., Khokhlova O., Kust P., Kurbanova F., Chernov T., Kutovaya O., Lebedeva M., Mihailov E. The environment of the Early Iron Age at the southern fringe of the forest zone of the Russian Plain // Quat. Int. 2018. V. 502B. P. 218–237. https://doi.org/10.1016/j.quaint.2018.04.002
Marfenina O.E., Ivanova A.E., Kislova E.E., Sacharov D.S. The mycological properties of medieval culture layers as a form of soil ‘biological memory’ about urbanization // J. Soils and Sediments. 2008. V. 8. P. 340–348. https://doi.org/10.1007/s11368-008-0032-2
Margesin R., Siles J., Cajthaml T., Öhlinger B., Kistler E. Microbiology Meets Archaeology: Soil Microbial Communities Reveal Different Human Activities at Archaic Monte Iato (Sixth Century BC) // Microbial Ecology. 2016. V. 73(4). P. 925–938. https://doi.org/10.1007/s00248-016-0904-8
Marinova E., Linseele V., Kühn M. Bioarchaeological research on animal dung – possibilities and limitations // J. Environmental Archaeology. 2013. V. 18(1). P. 1–3. https://doi.org/10.1179/1461410313Z.00000000023
Mitusov A.V., Mitusova O.E., Pustovoytov K., Lubos C.M., Dreibrodt S., Bork H.R. Palaeoclimatic indicators in soils buried under archaeological monuments in the Eurasian steppe: a review // Holocene. 2009. V. 19. P. 1153–1160.
Moore P., Webb J., Collison M. Pollen analysis. Blackwell, Oxford, 1991. 216 p.
Nannipieri P., Kandeler E., Ruggiero P. Enzyme activities and microbiological and biochemical processes in soil // Enzymes in the environment. Activity, ecology and applications. N.Y.: Marcel Dekker, 2002. P. 1–33.
Novenko E., Tsyganov A., Volkova E., Kupriyanov D., Mironenko I., Babeshko K., Utkina A., Popov V., Mazei Yu. Mid- and Late Holocene vegetation dynamics and fire history in the boreal forest zone of European Russia: a case study from the south-eastern part of Meschera Lowlands // Palaeogeography, Palaeoclimatology, Palaeoecology. 2016. V. 459. P. 570–584. https://doi.org/10.1016/j.palaeo.2016.08.004
Novenko T., Tsyganov A., Mazei N., Kupriyanov D., Rudenko O., Bobrovsky M., Erman N., Nizovtsev V. Palaeoecological evidence for climatic and human impacts on vegetation in the temperate deciduous forest zone of European Russia during the last 4200 years: A case study from the KaluzhskiyeZaseki Nature Reserve // Quat. Int. 2019. V. 516. P. 58–69. https://doi.org/10.1016/j.quaint.2018.06.028
Nyström T. Not quite dead enough: on bacterial life, culturability, senescence, and death // Archives of Microbiology. 2001. V. 176. P. 159–164. https://doi.org/10.1007/s002030100314
Patchaye M., Sundarkrishnan B., Tamilselvan S., Sakthivel N. Microbial management of organic waste in agroecosystem // Microorganisms for Green Revolution. V. 2: Microbes for Sustainable Agro-Ecosystem. Singapore: Springer, 2018. P. 45–74.
Peters S., Borisov A., Reinhold S., Korobov D., Thiemeyer H. Microbial characteristics of soils depending on the human impact on archaeological sites in the Northern Caucasus // Quat. Int. 2014. V. 324(4). P. 162–171. https://doi.org/10.1016/j.quaint.2013.11.020
Rosen A. Phytolith analysis // Encyclopedia of Archaeology. 2008. P. 1818–1822. https://doi.org/10.1016/B978-012373962-9.00247-8
Roszak D.B., Colwell R.R. Survival strategies of bacteria in the natural environment // Microbiological Reviews. 1987. V. 51. P. 365–379.
Ryabogina N., Borisov A., Idrisov I., Bakushev M. Holocene environmental history and populating of mountainous Dagestan (Eastern Caucasus, Russia) // Quat. Int. 2019. V. 516. P. 111–126. https://doi.org/10.1016/j.quaint.2018.06.020
Sadori L. Postglacial Pollen Records from Southern Europe //Reference Module in Earth Systems and Environmental Sciences. 2014. https://doi.org/10.1016/B978-0-12-409548-9.09389-1
Shi T., Reeves R.H., Gilichinsky D.A., Friedmann E.I. Characterization of viable bacteria in Siberian permafrost by 16S rDNA sequencing // Microbiolal Ecology. 1997. V. 33. P. 169–179. https://doi.org/10.1007/s002489900019
Skujins J.J. Extracellular enzymes in soil // Critical Reviews in Microbiology. 1976. V. 4. P. 383–421.
Trunova V., Zvereva V., Polosmak N., Kochubey D., Kriventsov V., Kuper K. Investigation of Organic Materials From the 'Royal' Burials of Xiongnu (Noin-Ula, Mongolia) by SRXRF and XAFS Methods // Archaeometry. 2015. V. 57(6). P. 1060–1077.https://doi.org/10.1111/arcm.12109
Tunno I., Mensing S. The value of non-pollen palynomorphs in interpreting paleoecological change in the Great Basin (Nevada, USA) // Quat. Res. 2017. V. 87. P. 529–543. https://doi.org/10.1017/qua.2017.8
van der Wal A., van Veen J., Smant W., Boschker H., Bloem J., Kardol P., van der Putten W., de Boer W. Fungal biomass development in a chronosequence of land abandonment // Soil Biol. Biochem. 2006. V. 38. P. 51–60.
Von Holstein I.C.C., Penkman K.E.H., Peacock E.E., Collins M.J. Wet degradation of keratin proteins: linking amino acid, elemental and isotopic composition // Rapid Commun. Mass Spectrom. 2014. V. 28(19). P. 2121–2133. https://doi.org/10.1002/rcm.6999
Wilkinson T. The definition of ancient manured zones by means of extensive shred-sampling techniques // J. Field Archaeol. 1982. V. 9. P. 323–333.
Williams C.A., Hills L.V., Krause F.F. Preserved organic matter and miospores in buried Middle Devonian (Givetian) paleosols: Indicators of weathering, oxidation and maturity // Catena. 1996. V. 28. Iss. 1–2. P. 1–19. https://doi.org/10.1016/S0341-8162(96)00027-6
Zarnoza R., Guerrero C., Mataix-Solera J., Scow K., Arcenegui V., Mataix-Beneyto J. Changes in soil microbial community structure following the abandonment of agricultural terraces in mountainous areas of Eastern Spain // Applied Soil Ecology. 2009. V. 42. P. 315–323.
Дополнительные материалы отсутствуют.