

Почвоведение, 2022, № 7, стр. 825-842
Изменение запасов углерода, микробной и ферментативной активности агродерново-подзолов южной тайги в ходе постагрогенной эволюции
И. Н. Курганова a, *, В. М. Телеснина b, В. О. Лопес де Гереню a, В. И. Личко a, Л. А. Овсепян a, c
a Институт физико-химических и биологических проблем почвоведения РАН
142290 Московская обл., Пущино, ул. Институтская, 2, Россия
b МГУ им. М.В. Ломоносова
119991 Москва, Ленинские горы, 1-12, Россия
c Тюменский государственный университет
625003 Тюмень, ул. Володарского, 6, Россия
* E-mail: ikurg@mail.ru
Поступила в редакцию 01.12.2021
После доработки 18.01.2022
Принята к публикации 26.01.2022
- EDN: JEOIDS
- DOI: 10.31857/S0032180X22070073
Аннотация
Органическое вещество (ОВ) почвы в значительной степени контролирует экосистемные функции почвы и является источником энергии и питательным субстратом для почвенного микробного сообщества. На примере хроноряда агродерново-подзолов (Retic Albic Podzol, южно-таежная зона, Костромская область), выведенных из сельскохозяйственного использования 11, 16 и 40 лет назад, проанализирована постагрогенная динамика содержания органического углерода (Сорг), общего азота (Nобщ) и их отношения C/N, значений рН, показателей микробиологической и ферментативной активности почв. Для почв хроноряда были также оценены стратификационные отношения (СтО) (0–5 : 5–10; 0–10 : 10–20 и 0–10 : 20–30) для содержания Сорг и Nобщ. Показано, что в ходе постагрогенной сукцессии в слое 0–5 см наблюдается достоверное увеличение содержания Сорг и Nобщ, а также усиление стратификации их распределения в пределах старопахотной толщи, особенно после образования сомкнутого древостоя на бывших пахотных почвах. Постагрогенная сукцессия также способствует росту скорости базального дыхания и содержания микробной биомассы, максимальные значения которых в пределах рассматриваемого хроноряда были характерны для почвы 40-летнего мелколиственного леса. По мере естественного лесовосстановления на бывших пахотных почвах выявлено достоверное усиление фосфатазной активности в пределах всего старопахотного горизонта. Пики активности оксиредуктаз соответствуют стадиям сукцессии с максимальным поступлением легкоразлагаемого растительного опада. За 40 лет постагрогенной сукцессии наблюдается статистически значимое увеличение всех анализируемых показателей (за исключением значений рН) в слое 0–5 см, который более других обогащен органическим веществом.
ВВЕДЕНИЕ
Органическое вещество (ОВ) почвы является ее жизненно важной составляющей, которая в значительной степени контролирует ее экосистемные функции и является источником энергии и питательным субстратом для почвенного микробного сообщества [18, 33, 35]. Количество и качественный состав почвенного ОВ определяются приуроченностью почв к той или иной природно-климатической зоне, в пределах которой эти характеристики будут зависеть от типа ценоза, вида землепользования и сельскохозяйственной нагрузки [64, 67, 79]. На запасы почвенного ОВ существенно влияет и смена типа землепользования [39, 59, 60]. Так, например, при вовлечении целинных почв в сельскохозяйственное использование, происходят неминуемые потери почвенного ОВ вследствие процессов минерализации и почвенной эрозии [38, 65, 74, 75]. Обратный процесс – прекращение использования почв в сельском хозяйстве, прежде всего, под пашню, ведет к восстановлению естественной растительности и накоплению ОВ в бывшем пахотном слое почв [44, 47, 50, 51, 55, 58].
В зоне южной тайги в ходе постагрогенной сукцессии на бывших сельскохозяйственных угодьях происходят процессы естественного лесовосстановления [43, 45, 46, 53], а по мере роста древостоя изменяется соотношение основных пулов почвенного ОВ [48, 62, 71, 72]. За счет формирования лесной подстилки на стадии образования сомкнутого древостоя в почвах образуется существенный пул слаборазложившегося ОВ [17, 44, 62], который составляет значительную часть общего пула почвенного углерода [43, 45, 76]. Кроме того, в ходе лесовосстановительной сукцессии возрастает сложность строения подстилки, увеличивается степень пространственного варьирования морфологических показателей органогенного горизонта [72, 73], и кардинальные изменения претерпевает биохимический состав растительного опада [56]. В целом при постагрогенном лесовосстановлении происходит изменение баланса между процессами минерализации и гумификации ОВ почв [9]. В легких почвах перераспределение ОВ в профиле особенно выражено и проявляется в уменьшении содержания и запасов органического углерода в минеральной части профиля за счет их увеличения в подстилке [43].
Изменение содержания и запасов ОВ в почвах залежей в ходе естественного лесовосстановления в подзоне южной тайги достаточно хорошо изучено [10, 11, 45, 55, 61]. Показано, что в зависимости от степени окультуренности бывших пахотных почв, содержание углерода в них может возрастать [17, 51, 55], уменьшаться [12, 16, 58] или оставаться на прежнем уровне [42, 62]. Прекращение сельскохозяйственного использования почв в лесной зоне ведет к усилению их дыхательной активности и увеличению пула микробного углерода [40, 54, 55, 57, 69, 78].
Несмотря на то, что микробная активность и ферментный пул почвы играют важную роль в круговороте почвенного ОВ, изучению ферментного статуса почв в ходе их постагрогенной эволюции до настоящего времени уделялось достаточно мало внимания. Хотя известно, что изменение микробной и ферментативной активности обнаруживается задолго до изменения других показателей качества почвы [81]. Внеклеточные почвенные ферменты отвечают за процессы окисления и гидролиза полимерных и олигомерных органических соединений, осуществляя баланс между разложением поступающих в почву растительных остатков и образованием устойчивых гумусовых соединений [31, 79]. Считается, что гидролитические ферменты запускают начальные стадии деструкции свежих растительных остатков и являются ответственными за расщепление гидролизуемых соединений – углеводов (целлюлозы, хитина) и белков [20, 31, 37, 77]. Внеклеточные гидролитические ферменты катализируют большинство биохимических реакций в почве [70], расщепляя высокомолекулярные соединения на мономеры, которые могут напрямую использоваться микроорганизмами [34]. Ферменты группы оксидоредуктаз (фенолоксидазы и пероксидазы) участвуют в процессах минерализации фенольных соединений, включая окисление сложных ароматических структур лигнина и гумусовых кислот [23, 36, 41, 80]. При окислении и спонтанной конденсации фенольных субстратов различной молекулярной массы параллельно могут происходить и реакции синтеза специфических гумусовых кислот [30, 80]. Таким образом, активность внеклеточных ферментов определяет как скорость образования растворимых органических соединений, доступных почвенным микроорганизмам, так и интенсивность процессов трансформации ОВ почв.
Активность разных групп ферментов является важной характеристикой функционирования почвенного микробного сообщества [6, 14, 28]. Из-за быстрой реакции ферментного пула на многие изменения окружающей среды и землепользования [32, 78], активность ферментов широко используется в качестве индикатора микробной активности и плодородия почвы [27, 29]. Немногочисленные литературные данные свидетельствуют о том, что в дерново-подзолах Костромской области за 40 лет постагрогенного развития происходило восстановление инвертазной активности почти до фонового уровня, тогда как уровень целлюлазной активности за этот период не восстановился [72]. Активность каталазы в почвах хроноряда возрастала в течение всей сукцессии, что, возможно, было связано с уменьшением плотности сложения почв и улучшением их воздушного режима. При исследовании постагрогенных серых лесных почв [14] выявлена низкая активность ферментов гидролитической группы для молодых залежей, в то время как на залежах старше 30 лет активность гидролаз сравнима с таковой в почве вторичного леса. Активность ферментов группы оксидаз была наиболее высокой в почвах “крайних” временных стадий сукцессии – на пашне и во вторичном лесном ценозе. Причины подобной динамики и многие аспекты взаимосвязи ферментативной активности с другими почвенными свойствами в настоящее время остаются открытыми. Исследование структурных изменений в составе ферментного пула позволит понять особенности процессов трансформации почвенного ОВ в ходе самовосстановления почв после прекращения их сельскохозяйственного использования.
Задачи настоящей работы включали: 1 – изучение постагрогенной динамики содержания Сорг, Nобщ, отношения C/N, значений рН, показателей микробиологической и ферментативной активности в дерново-подзолах Костромской области, 2 – оценку стратификационных отношений для содержания Сорг и Nобщ в пределах старопахотного горизонта и 3 – определение взаимосвязей микробного и ферментного пула почв с другими почвенными характеристиками. Мы предполагаем, что восстановление естественной растительности на бывших пахотных почвах, которое влечет за собой изменение объема, доступности и биохимического разнообразия растительного опада, будет приводить к увеличению запасов ОВ в почвах и сопряженному росту микробного и ферментного пулов. Скорость постагрогенных изменений анализируемых показателей в пределах старопахотного горизонта будет различной.
ОБЪЕКТЫ И МЕТОДЫ
Природные условия территории исследования. Исследование проводили в Мантуровском районе Костромской области в хроноряду залежей разного возраста (табл. 1). Климат региона умеренно-континентальный, за последние 20 лет среднегодовая температура воздуха составила 3.6°С, среднегодовое количество осадков – 644 мм [1]. Территория относится к северо-восточной подобласти атлантико-континентальной лесной области [4]. Условия почвообразования в районе исследований характерны для южной тайги, где естественные растительные сообщества представлены еловыми травяно-кустарничковыми лесами [15], а преобладающими типами почв являются дерново-подзолистые (Albic Retisol) и подзолы (Retic Albic Podzol) [7]. Почвообразующие породы представляют собой разнообразные ледниковые и водно-ледниковые отложения. Чаще всего это двучленные породы, или еще более литологически сложные и неоднородные отложения, хотя верхние 40–45 см, включая старопахотную толщу, довольно однородны и имеют супесчаный гранулометрический состав [72].
Таблица 1.
Местоположение, общая характеристика растительности и почв в залежном хроноряду в Костромской области
| Стадия зарастания, координаты | Растительность | Почва [7], формула профиля |
|---|---|---|
| 1. Пашня N 58°10′55.0″ E 44°28′27.8″ |
Посевы овса | Агродерново-подзол литобарьерный среднепахотный супесчаный на ледниковых отложениях: P(0–30)– Ehi(30–46)–Ef(46–66)–BF(66–72)–BC(72–100) |
| 2. Залежь, 11 лет N 58°10′52.7″ E 44°28′23.1″ |
Луг с преобладанием овсяницы красной (Festuca rubra L.), ежи сборной (Dactylis glomerata L.) и ястребинки зонтичной (Hieracium umbellatum L.). Единичные низкорослые деревья ивы козьей (Salix caprea L.) | Агродерново-подзол реградированный литобарьерный среднепахотный супесчаный на ледниковых отложениях: AYpa(0–30)–Ehi(30–43)–Ef(43–67)–BF(67–105) |
| 3. Залежь, 16 лет N 58°10′52.8″ E 44°28′21.0″ |
Почти сомкнутый древостой из ивы козьей, в травостое преобладает овсяница красная, ситник нитевидный (Juncus filiformis L.) и грушанка круглолистная (Pyrola rotundifolia L.) | Агродерново-подзол реградированный литобарьерный глееватый среднепахотный супесчаный на ледниковых отложениях: AYpa(0–30)–Ehi(30–42)–Еfg (42–55)–BFg(55–92) |
| 4. Лес вторичный, 40 лет N 58°10′54.0″ E 44°28′21.6″ |
Лес осиново-березовый с преобладанием звездчатки жестколистной (Stellaria holostea L.) и черники (Vaccinium myrtillus L.) в напочвенном покрове | Дерново-подзол постагрогенный литобарьерный супесчаный на ледниковых отложениях: O(0–2)–AYpa(2–25)– Ehi(25–43)–Ef(43–64)–BF(64–110) |
| 5. Лес старовозрастный смешанный, 100 лет N 58°10′56.1″ E 44°28′29.0″ |
Лес елово-березовый с преобладанием черники в напочвенном покрове | Подзол грубогумусированный литобарьерный супесчаный на ледниковых отложениях: O(0–4)–AH(4–6)–Ehi(6–22)–E(22–48)–Ef(48–60)–BF(60–97) |
Хроноряд зарастающих пахотных почв состоял из следующих стадий (табл. 1): 1) пашня (0-стадия), засеянная овсом или многолетними травами; почва слабо окультурена, в последние 25–30 лет проводилась распашка без внесения удобрений; 2) 11-летняя залежь с луговой растительностью; 3) 16-летняя залежь с древостоем из ивы козьей и богатым разнотравьем; 4) мелколиственный осиново-березовый вторичный лес 40-летнего возраста; 5) березово-еловый лес, возраст которого около 100 лет, выступающий в качестве контроля.
Данные об истории и продолжительности освоения земель получены от сотрудников экологической станции ИПЭЭ РАН им. А.Н. Северцова, расположенной в Мантуровском районе Костромской области. Возраст древостоя определяли методом кернения (бур Пресслера). Начиная с 2008 г. на площадках ведутся регулярные наблюдения за изменением состава растительности и свойств почвы [72, 73, 76]. Описание растительности и отбор биомассы травяного (травяно-кустарничкового) яруса проводили практически ежегодно, массу древостоя и его фракций определяли аллометрически [5], надземную массу травяно-кустарничкового яруса оценивали методом укосов, корневую – методом монолитов (пятикратная повторность).
Отбор почвенных проб и определение общих химических свойств почв. На каждом из пяти объектов, составляющих хроноряд, методом конверта были отобраны смешанные почвенные пробы со следующих глубин: 0–5, 5–10, 10–20 и 20–30 см. В прикопках, на тех же глубинах, используя бур Качинского, определяли плотность почвы (BD, г/см3) весовым методом [22]. Образцы почв высушивали до воздушно-сухого состояния и просеивали через сито с диаметром ячеек 2 мм. Корни и грубодисперсный органический материал удаляли вручную. Содержание органического углерода (Cорг) и общего азота (Nобщ) определяли на автоматическом CHNS анализаторе (Leco, США) в трехкратной повторности. Запасы Сорг (Зап-Сорг, т С/га) в определяемом слое рассчитывали с учетом содержания Сорг (г С/кг почвы), плотности почвы (г/см3) и мощности слоя (H, см) по формуле:
(1)
${\text{Зап - }}{{{\text{С}}}_{{{\text{орг}}}}} = {{({{{\text{С}}}_{{{\text{орг}}}}} \times BD \times H)} \mathord{\left/ {\vphantom {{({{{\text{С}}}_{{{\text{орг}}}}} \times BD \times H)} {10}}} \right. \kern-0em} {10}}.$Запасы Сорг в 20-сантиметровой толще находили простым суммированием Зап-Сорг в каждом из составляющих ее слоев (0–5, 5–10 и 10–20 см). Определение величины рН проводили в 1 М растворе КСl (соотношение почвы и раствора 1 : 2.5) на рН-метре Metler-Toledo (Швейцария) в двухкратной повторности.
Анализ микробной активности почв. Базальное дыхание почв (БД) определяли в трехкратной повторности по интенсивности выделения СО2 из почвы после семи дней предварительной инкубации при увлажнении, соответствующем 70–75% их наименьшей влагоемкости (НВ), и температуре 22–24°С [52]:
(2)
$\begin{gathered} {\text{БД}}\,\,\left( {{{{\text{мг С}}} \mathord{\left/ {\vphantom {{{\text{мг С}}} {\left( {{\text{кг}}\,\,{\text{почвы}}\,\,{\text{сут}}} \right)}}} \right. \kern-0em} {\left( {{\text{кг}}\,\,{\text{почвы}}\,\,{\text{сут}}} \right)}}} \right) = \\ = \,\,\left( {{{С}_{1}}--{\text{ }}{{С}_{0}}} \right) \times 12 \times {{V}_{{{\text{флак}}}}} \times {{1000} \mathord{\left/ {\vphantom {{1000} {\left( {m \times 22.4 \times t \times 100} \right)}}} \right. \kern-0em} {\left( {m \times 22.4 \times t \times 100} \right)}}, \\ \end{gathered} $Содержание углерода микробной биомассы (Смик) определяли методом субстрат-индуцированного дыхания [24] в почвенных образцах после измерения БД и внесения 1 мл раствора глюкозы из расчета 10 мг глюкозы на 1 г почвы. Скорость субстрат-индуцированного дыхания (VСИД) рассчитывали по формуле (2), выражая в мкл CО2/(г почвы час). Содержание Смик (мг С/кг почвы) рассчитывали по формуле [24]:
На основе полученных микробных характеристик (БД и Смик) и содержания Cорг для почв хроноряда были оценены относительные индексы функционирования микробного сообщества почвы и доступности ОВ разложению [40, 49, 66, 68]:
1. Микробный метаболический коэффициент (qCO2) или удельное дыхание микробной биомассы, представляющий собой отношение БД/Cмик (мг СО2-С/г С мик/сут) и являющийся показателем экофизиологического состояния микробного сообщества почвы [26];
2. Отношение БД/Cорг как индикатор доступности органического углерода почвы разложению [52, 53].
Анализ ферментативной активности почв. Активность ферментов группы оксидоредуктаз (полифенолоксидазы и пероксидазы) определяли методом Карягиной-Михайловой [20]. С этой целью брали навеску почвы массой 1 г, добавляли к ней 10 мл свежеприготовленного 1% раствора гидрохинона и 1 мл 0.05% раствора перекиси водорода. Затем суспензию тщательно перемешивали и помещали в термостат на 30 мин при +30°C. В качестве контроля использовали смесь растворов гидрохинона и перекиси водорода без почвы. После проведения инкубации в колбы добавляли 10 мл этилового спирта, суспензию снова тщательно перемешивали и фильтровали через двойной фильтр “синяя лента”. Оптическую плотность спиртовых вытяжек, имеющих желтую окраску, измеряли при длине волны 450 нм на спектрофотометре (UNICO 2080, США). Активность пероксидазы (ПРО) выражали в условных единицах, соответствующих преобразованию 1 мг бензохинона на 1 г почвы за 30 мин инкубирования (мг БЗХ/г почвы/30 мин) при температуре 30°С [20].
Определение полифенолоксидазной (ПФО) активности проводили аналогично вышеописанному алгоритму определения пероксидазы, но перекись водорода не вносили, поскольку для окисления фенолов используется кислород воздуха [20]. Навеска почвы составляла 2 г.
Активность фосфатазы (ФСФ) определяли методом Галстян-Арутюнян [20]. Для этого брали навеску почвы массой 1 г, предварительно увлажняли до 90% их НВ, добавляли к ней 1 мл свежеприготовленного 1% раствора фенолфталеинфосфата натрия и 1–2 капли толуола, затем суспензию тщательно перемешивали и помещали в термостат на 1 ч при 30°С. После проведения инкубации в колбы добавляли 45 мл дистиллированной воды, 1 мл алюмокалиевых квасцов и 2 мл 10% раствора аммиака. Суспензию снова тщательно перемешивали и фильтровали через фильтр “синяя лента”. Оптическую плотность получившихся вытяжек, имеющих фиолетовую окраску, определяли при длине волны 553 нм на спектрофотометре (UNICO 2080, США) и выражали в мг Р2О5/100 г почвы/час [21].
Обработка данных. Оценку статистической значимости постагрогенных изменений для всех свойств почвы за 40 лет выполняли с помощью моделей линейной регрессии вида у = aх + b, выражающих зависимости свойств почвы (у, средние арифметические значения) от возраста залежи (х). Возраст пахотной почвы принимали равным 0; лесной ценоз из анализа исключали. Степень дифференциации старопахотного горизонта по содержанию Cорг и Nобщ при зарастании пашни лесом оценивали с помощью стратификационного индекса (СтО), которое представляет отношение между содержанием Сорг (Nобщ) в различных слоях почвы: СтО(0–5 : 5–10), СтО(0–10 : 10–20) и СтО(0–10 : 20–30).
Для определения связи между изучаемыми почвенными свойствами (средние арифметические значения) в пределах полного хроноряда с включением старовозрастного леса использовали коэффициент корреляции Пирсона (F-критерий). Визуализация данных выполнялась в программе MS Excel 2013. В таблицах и на графиках представлены средние значения и стандартная ошибка (SE). Статистический анализ проводили с использованием пакета Анализ данных (MS Excel 2013) и программы Statistica 6 при уровне значимости α = 0.05.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Изменение экосистемных пулов углерода. В течение демутационной сукцессии происходит существенное изменение состава и структуры растительных сообществ. Так, примерно через 7–8 лет после прекращения распашки появляются отдельные деревья [76], а через 15 лет наблюдается слабо сомкнутый древостой из ивы козьей. Спустя 35–40 лет демутационной сукцессии во вторичном мелколиственном лесу появляется подрост из ели, который, впоследствии сможет сформировать древостой. Таким образом, в ходе постагрогенной восстановительной сукцессии возрастает биомасса древостоя (табл. 2), которая в 40-летнем лесу достигает 138 т/га. Биомасса травяно-кустарничкового яруса, напротив, уменьшается с 3.4 т/га в 11-летней залежи до 0.50 т/га в залежи под 40-летним лесом и затем снова возрастает до 0.92 т/га в 100-летнем лесу за счет возрастания доли кустарничков. В лесу 100-летнего возраста в древостое преобладает ель, в связи с чем в его опаде доля хвои и шишек достигает 40% [19, 73], что не может не отражаться на скорости разложения органического вещества.
Таблица 2.
Динамика некоторых показателей биологического круговорота в ходе постагрогенной сукцессии
| Показатели круговорота | Пашня | Залежь | Лес | |||
|---|---|---|---|---|---|---|
| 11 лет | 16 лет | 40 лет | 100 лет | |||
| Биомасса древостоя, т/га | надземная | 0 | 0 | 23.8 | 138.2 | 155.8 |
| подземная | 0 | 0 | 4.2 | 38.8 | 43.8 | |
| Биомасса травяно-кустарничкового яруса, т/га | надземная | 0 | 3.4 | 0.62 | 0.50 | 0.92 |
| подземная | 3.9 | 5.1 | 3.3 | 4.5 | 4.8 | |
| Трудноразлагаемый опад т/га | ветки, шишки, хвоя | 0 | 0 | 2.6 | 3.7 | 3.6 |
| Легкоразлагаемый опад (ЛРО), т/га, в том числе |
листва деревьев | 0 | 0 | 1.2 | 3.4 | 2.5 |
| травяно-кустарничковый ярус, надземный | 0 | 3.4 | 0.21 | 0.23 | 0.04 | |
| травяно-кустарничковый ярус, корневой | 1.3 | 1.7 | 1.1 | 1.5 | 1.6 | |
| ВСЕГО | 1.3 | 5.1 | 2.5 | 5.1 | 4.1 | |
| Запасы подстилки, т/га | 0 | 0 | 1.00 | 2.16 | 18.9 | |
| Зольность подстилки (горизонт L), % | – | – | 8.7 | 6.6 | 4.5 | |
| Запасы С в почве (слой 0–20 см), т С/га | 31.8 | 37.8 | 47.8 | 46.4 | 128.6 | |
В течение демутационной сукцессии, запасы ежегодно поступающего в почву легкоразлагаемого опада, включающего в себя листву деревьев, надземную часть травяно-кустарничкового яруса и часть корневой биомассы травяно-кустарничкового яруса, имеют два максимума. Первый наблюдается на стадии луга (за счет высокопродуктивного травостоя), второй характерен для стадии мелколиственного леса (за счет листового опада). По мере развития древостоя увеличивается запасы и мощность подстилки, а ее масса в старовозрастном лесу почти в 10 раз больше, чем в молодом осиново-березовом древостое. Величина зольности верхнего горизонта подстилки, которая может служить косвенной характеристикой скорости минерализации поступающего опада, уменьшается в ходе сукцессии почти в 2 раза. Запасы Сорг в слое 0–20 см уже на стадии 16-летней залежи в 1.5 раза превосходят запасы Сорг в пахотной почве и остаются на том же уровне в почве молодого леса, что свидетельствует о восстановлении углерод-секвестрирующей функции почв на более поздних стадиях постагрогенного развития, благодаря постепенному возобновлению естественной растительности. В почве старовозрастного леса (слой 0–20 см) запасы Сорг уже в 4 раза превышают таковые на пахотном участке. Таким образом, по мере восстановления естественной растительности емкость биологического круговорота, косвенно определяемая объемом растительной биомассы, увеличивается, а интенсивность биологического круговорота, которую можно косвенно охарактеризовать зольностью подстилки, уменьшается.
Постагрогенная динамика общих свойств почв. В течение постагрогенной сукцессии в верхней части профиля увеличивается потенциальная кислотность почвы, что согласуется с данными, полученными другими авторами для лесных почв [11, 13]. В пределах старопахотного горизонта (0–30 см), наибольший градиент постагрогенного подкисления выявлен для верхнего (0–5 см) слоя (рис. 1a), в котором уже через 10–11 лет после прекращения распашки величина рНKCl уменьшается почти на единицу. В почвах лесных стадий сукцессии, когда образован сомкнутый древостой, значения рНKCl практически не превышают 4.0 ед., что связано с резким уменьшением поступления травяного опада и формированием лесной подстилки, продукты разложения которой оказывают подкисляющее действие на почву. Уменьшение величины рНKCl в ходе 40-летней постагрогенной сукцессии показывает значимый линейный тренд (р < 0.05) на всех глубинах, кроме слоя 0–5 см (табл. 3). Скорость снижения значений рНKCl максимальна в слое 0–5 см (–0.032 ед./год), а во всех других слоях она примерно одинакова и составляет –0.023…–0.025 ед./год.
Рис. 1.
Постагрогенная динамика величины рНКСL (a), содержания Сорг (b), Nобщ (c) и соотношения С/N (d) на разных глубинах старопахотного горизонта агродерново-подзолов: 1 – 0–5 см, 2 – 5–10 см, 3 – 10–20 см, 4 – 20–30 см.

Таблица 3.
Характеристики регрессионных линейных моделей, описывающих изменение величины рНКСl, содержания Сорг, Nобщ и отношения С/N в течение 40 лет постагрогенной эволюции агродерново-подзолов. R2 – коэффициент детерминации, p – уровень значимости, а – коэффициент регрессионной модели у = aх + b (единицы измерения анализируемого свойства/год)
| Свойство | Характеристика модели | Глубина слоя, см | |||
|---|---|---|---|---|---|
| 0–5 | 5–10 | 10–20 | 20–30 | ||
| рНКСl | R2 | 0.77 | 0.91 | 0.87 | 0.83 |
| р | 0.12 | 0.04 | 0.07 | 0.09 | |
| а | –0.032 | –0.023 | –0.025 | –0.025 | |
| Сорг, г С/кг почвы | R2 | 0.99 | 0.97 | 0.47 | 0.92 |
| р | 0.00 | 0.02 | 0.31 | 0.04 | |
| а | 0.64 | 0.25 | 0.10 | 0.32 | |
| Nобщ, г N/кг почвы | R2 | 0.97 | 0.78 | 0.26 | 0.94 |
| р | 0.02 | 0.12 | 0.49 | 0.03 | |
| а | 0.038 | 0.012 | 0.003 | 0.019 | |
| C/N | R2 | 0.91 | 0.74 | 0.60 | 0.39 |
| р | 0.05 | 0.14 | 0.23 | 0.37 | |
| а | 0.065 | 0.060 | 0.053 | 0.062 | |
Содержание Сорг в ходе постагрогенной сукцессии демонстрирует существенный рост, особенно выраженный в самой верхней части старопахотной толщи. Так, в слое 0–5 см за 11 лет залежного режима содержание Сорг по сравнению с пахотной почвой возрастает в 2 раза, на стадии мелколиственного леса – в 4 раза, а в березово-еловом лесу – в 7 раз. В слое 5–10 см содержание Сорг наиболее значительно изменяется при переходе от мелколиственного 40-летнего леса к смешанному старовозрастному (рис. 1b). В более глубоких слоях старопахотного горизонта (10–20 и 20–30 см) увеличение содержание Сорг происходит только до стадии 40-летнего мелколиственного леса, а в почве старовозрастного леса оно либо не изменяется (слой 10–20 см), либо уменьшается (слой 20–30 см). Постагрогенная динамика содержания Nобщ показывает тенденции, сходные с теми, что были описаны выше для содержания Сорг (рис. 1c). Соотношение С/N в ходе восстановительной постагрогенной сукцессии увеличивается почти на всех анализируемых глубинах, что говорит об уменьшении интенсивности разложения ОВ почв, особенно на более поздних стадиях сукцессии, соответствующих лесным сообществам.
Оценка линейных трендов изменения содержания Сорг показывает, что за 40 лет постагрогенной сукцессии наблюдается его статистически значимое увеличение во всей старопахотной толще (за исключением слоя 20–30 см) с наибольшей скоростью прироста 0.64 г С/(кг почвы год) в верхнем слое (0–5 см). Значимый рост содержания Nобщ наблюдается в слоях 0–5 и 20–30 см с коэффициентами линейной регрессии 0.038 и 0.019 г N/(кг почвы год), соответственно. Как правило, в ходе восстановительной сукцессии прирост содержания Сорг и Nобщ в более глубоких слоях становится менее выраженным. Для почв хроноряда выявлена отрицательная корреляция между содержанием Сорг, Nобщ и отношением С/N с величиной рНKCl в слоях 0–5 и 5–10 см (r = –0.75…–0.90, p < < 0.05) (табл. 4), что отмечалось и более ранних работах для почв изучаемого хроноряда [55].
Таблица 4.
Коэффициенты корреляции Пирсона между почвенными свойствами в слоях 0–5 и 5–10 см в исследованном хроноряду дерново-подзолов (корреляции, значимые при р < 0.05, выделены цветом: красным – положительные, синим – отрицательные)

[i] * Свойства: Сорг – содержание органического углерода (г С/кг почвы), Nобщ – содержание общего азота (г N/кг почвы), C/N отношение Сорг к Nобщ, БД – скорость базального дыхания (мг С/(кг почвы cут)), Смик – содержание микробной биомассы (мг С/кг почвы), ПФО – активность полифенолоксидазы (мг БЗХ/(г почвы 30 мин)), ПРО – активность пероксидазы (мг БЗХ/(г почвы 30 мин)), ФСФ – активность фосфатазы (мг Р2О5/(100 г почвы ч)).
Изменение стратификационных отношений Сорг и Nобщ в ходе постагрогенного развития. В пахотной почве распределение Сорг и Nобщ в изучаемом слое 0–30 см достаточно равномерно и с глубиной изменяется незначительно (рис. 1b, 1c). По мере восстановления растительности наблюдается выраженное перераспределение ОВ в пределах старопахотного горизонта. Так, в почве под 11-летним лугом содержание Сорг на глубине 0–5 и 20–30 см различается в 2 раза, а в почве под старовозрастным лесом – более чем в 4 раза. На ранних стадиях постагрогенной сукцессии это обусловлено обособлением дернового горизонта, а на более поздних – формированием органопрофиля, типичного для лесных почв, в котором грубогумусная верхняя часть профиля почти сразу сменяется минеральными горизонтами, обедненными органическим веществом.
Используя стратификационные отношения (СтО), мы количественно оценили особенности перераспределения содержания Сорг и Nобщ в пределах старопахотного горизонта (рис. 2). Их постепенная стратификация начинается в слое 0–10 см: в пашне значения СтО(0 : 5–5 : 10) не превышают 1.2, а в почве мелколиственного леса составляют 1.8. В почве старовозрастного березово-елового леса, когда в самую верхнюю часть минерального профиля проникают органические вещества из подстилки, значение СтО(0 : 5–5 : 10) достигает 2.6. Величины СтО(0 : 10–10 : 20) и СтО(0 : 10–20 : 30) содержания Сорг и Nобщ изменяются сходным образом до стадии формирования 40-летнего мелколиственного леса. В почвах старовозрастного леса значения СтО между слоем 0–10 см и более глубокими слоями достигают 6.0–8.1 и 6.3–9.5 для содержания Сорг и Nобщ соответственно. Такое изменение профильного распределения ОВ в ходе сукцессии свидетельствует о постепенной дифференциации старопахотного горизонта и формировании подстилки в почвах под лесными насаждениями, сформировавшимися в ходе восстановительной сукцессии.
Рис. 2.
Изменение стратификационных отношений (СтО) содержания Сорг (a) и Nобщ (b) в различных слоях старопахотного горизонта в ходе постагрогенного развития агродерново-подзолов: 1 – СтО(0 : 5–5 : 10), 2 – СтО(0 : 10–10 : 20), 3 – СтО(0 : 10– 0 : 30).

Под естественной растительностью ОВ почв, как правило, более стратифицировано по глубине по сравнению с пахотными почвами, поэтому при естественном лесовосстановлении величины СтО для Сорг и Nобщ увеличивается, что свидетельствует об улучшении экологического качества почв [49, 61]. Стратификационные отношения часто используют для оценки секвестрации углерода в почвах [63, 82]. Ранее, на примере двух залежных хронорядов Костромской области с использованием СтО (0–10 : 20–30) продемонстрирована постепенная дифференциация старопахотной толщи по содержанию и запасам Сорг, которая была максимально выражена в почвах субклимаксных смешанных лесов 90–100 лет [61]. Было также показано, при естественном лесовосстановлении наблюдается заметная стратификация верхней части старопахотного горизонта для всех почвенных характеристик, положительно связанных с содержанием Сорг [55].
Постагрогенная динамика характеристик микробной активности почв. Показатели биологической активности почв, такие как базальное дыхание и содержание микробной биомассы наиболее быстро реагируют на изменение локальных экологических условий [25], в том числе характера растительности в процессе естественного постагрогенного лесовосстановления [51, 53, 56]. Вслед за увеличением запасов ОВ в почвах в ходе постагрогенной сукцессии, содержание Смик демонстрирует наиболее заметный рост в слое 0–5 см (рис. 3a). Пахотная почва характеризуется минимальным содержанием Смик во всех слоях старопахотного горизонта (140–180 мг C/кг почвы), где единственным источником ОВ является корневая биомасса, оставшаяся после уборки урожая сельскохозяйственных культур. Следовательно, в почве постоянно присутствует дефицит свежего легкоразлагаемого растительного материала, что не может не отражаться на состоянии микробного сообщества. Через 11 лет после прекращения распашки и формирования лугового ценоза с богатым разнотравьем содержание Смик в слое 0–5 см увеличивается более чем вдвое, а через 40 лет, на стадии мелколиственного леса содержание Смик достигает максимального значения – 936 мг C/кг почвы. Возможно, это обусловлено тем, что на этой стадии происходит существенное возрастание запасов поступающего в почву легко разлагаемого опада, характеризующегося относительно высоким содержанием зольных элементов (табл. 2). Вместе с тем кислотность опада и верхнего слоя почвы еще не настолько повышена, чтобы ингибировать активность почвенной микробиоты. Именно кислая реакция среды верхнего почвенного слоя и хвойно-лиственного опада в старовозрастном смешанном лесу послужила причиной резкого снижения содержания Смик в слое 0–5 см (до 245 мг C/кг почвы). Сходный характер постагрогенной динамики содержания Смик наблюдается и в других слоях старопахотного горизонта, но выраженность различий с глубиной сглаживается.
Рис. 3.
Постагрогенная динамика содержания углерода микробной биомассы, Смик (a), скорости базального дыхания, БД (b), отношения БД/Сорг (c) и метаболического коэффициента qCO2 (d) на разных глубинах старопахотного горизонта агродерново-подзолов: 1 – 0–5 см, 2 – 5–10 см, 3 – 10–20 см, 4 – 20–30 см.
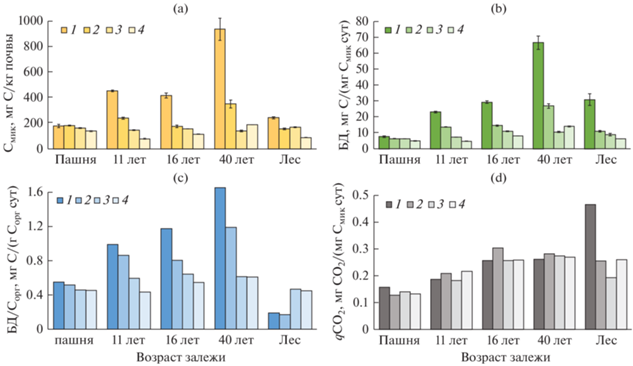
Постагрогенная динамика скорости БД во всех слоях старопахотного горизонта в общих чертах повторяет динамику содержания Смик (рис. 3b): минимальные значения БД в пахотной почве, затем трехкратный рост скорости БД на луговой стадии и максимальная величина БД на стадии мелколиственного леса. Наиболее заметные изменения скорости БД в почвах разных стадий сукцессии проявляются в верхнем (0–10 см) почвенном слое.
Оценка линейных трендов изменения микробных характеристик показывает, что за 40 лет постагрогенной сукцессии наблюдается статистически значимое увеличение содержания Смик в слое 0–5 см, а скорости БД – на всех глубинах старопахотного слоя за исключением глубины 10–20 см. Наиболее выраженный рост этих характеристик отмечается в слое 0–5 см и составляет 18.51 мг С/кг почвы/год для Смик и 1.48 мг С/кг почвы/сут/год для скорости БД (табл. 5), а длительность постагрогенного восстановления объясняет 97–99% дисперсии содержания Смик и скорости БД, соответственно. В слое 5–10 см величина прироста содержания Смик и величины БД уменьшается в 4.4 и 3 раза соответственно. Между скоростью БД и содержанием Смик (слои 0–5 и 5–10 см) выявлена значимая положительная корреляция (r = 0.94, p < 0.05) (табл. 4), что отмечается в ходе постагрогенной эволюции почв и в других биоклиматических зонах [51, 57].
Таблица 5.
Характеристики регрессионных линейных моделей, описывающих изменение микробной и ферментативной активности в течение 40 лет постагрогенной эволюции агродерново-подзолов. R2 – коэффициент детерминации, p – уровень значимости, а – коэффициент регрессионной модели у = aх + b (единицы измерения анализируемого свойства/год)
| Свойство | Характеристики модели | Глубина слоя, см | |||
|---|---|---|---|---|---|
| 0–5 | 5–10 | 10–20 | 20–30 | ||
| Смик, мг С/кг почвы | R2 | 0.97 | 0.76 | 0.70 | 0.43 |
| р | 0.01 | 0.13 | 0.16 | 0.34 | |
| а | 18.51 | 4.19 | –0.51 | 1.81 | |
| БД, мг С/(кг почвы сут) | R2 | 1.00 | 0.99 | 0.57 | 0.91 |
| р | 0.00 | 0.00 | 0.25 | 0.05 | |
| а | 1.48 | 0.50 | 0.11 | 0.24 | |
| БД/Сорг, мг С/(г Сорг сут) | R2 | 0.96 | 0.93 | 0.42 | 0.77 |
| р | 0.02 | 0.04 | 0.35 | 0.12 | |
| а | 0.027 | 0.016 | 0.003 | 0.004 | |
| ФСФ, мг P2O5/(100 г почвы ч) | R2 | 0.98 | 0.90 | 0.95 | 0.96 |
| р | 0.01 | 0.05 | 0.02 | 0.02 | |
| а | 8.37 | 4.68 | 4.87 | 3.61 | |
| ПРО, мг БЗХ/(г почвы 30 мин) | R2 | 0.90 | 0.56 | 0.92 | 0.79 |
| р | 0.05 | 0.25 | 0.04 | 0.11 | |
| а | 1.27 | 0.98 | 0.79 | 0.82 | |
| ПФО, мг БЗХ/(г почвы 30 мин) | R2 | 0.98 | 0.93 | 0.29 | 0.97 |
| р | 0.01 | 0.04 | 0.46 | 0.01 | |
| а | 0.70 | 0.72 | 0.52 | 0.81 | |
Отношение БД/Сорг может служить индикатором доступности ОВ почв разложению [51, 52]. Анализ постагрогенной динамики БД/Сорг показывает, что в течение первых 40 лет восстановительной сукцессии в верхней части старопахотной толщи (слои 0–5 и 5–10 см) этот показатель увеличивается в 2–4 раза (рис. 3c). Это свидетельствует о том, что доступность разложению вновь образованного в ходе постагрогенной сукцессии ОВ увеличивается за счет поступления легкоразлагаемого растительного опада. Следовательно, стабильность ОВ при прекращении распашки уменьшается, что отмечалось для почв изучаемого хроноряда и ранее [55]. В почве смешанного старовозрастного леса отношение БД/Сорг (слой 0–10 см) в 3 раза меньше, чем в пахотной почве, и, по-видимому, объясняется тем, что ОВ, сформированное за счет хвойно-лиственного опада, существенно труднее поддается разложению. Отмеченные закономерности в целом соответствуют данным, полученным ранее для хроноряда постагрогенных серых лесных почв [51, 53].
Оценка линейных трендов изменения отношения БД/Сорг за 40 лет постагрогенной сукцессии выявила статистически значимый рост этого показателя только в слоях 0–5 и 5–10 см, а длительность постагрогенного восстановления объясняла 93–96% дисперсии этого показателя (табл. 5). Для слоев 0–5 и 5–10 см выявлена значимая взаимосвязь между отношением БД/Сорг и содержанием Смик (r = 0.82, p < 0.05) (табл. 4), что вполне ожидаемо.
Микробный метаболический коэффициент qCO2, представляющий отношение скорости БД к содержанию Cмик служит индикатором обогащенности среды субстратом, необходимым для жизнедеятельности микробиоты, то есть показывает, насколько доступным является ОВ почвенным микроорганизмам [3, 40]. Наименьшие значения qCO2 (0.13–0.16 мг СО2/(мг Смик сут)) характерны для пахотной почвы, в которой микробное сообщество испытывает недостаток питательного субстрата, поскольку поступление растительного опада здесь чрезвычайно мало (табл. 3). В течение 40-летней восстановительной сукцессии бывших пахотных почв до стадии мелколиственного леса значения qCO2 постепенно возрастают во всех слоях старопахотного горизонта (рис. 3d). И хотя в пределах слоя 0–30 см значения qCO2, как правило, близки между собой, более высокие значения микробного метаболического коэффициента наблюдаются на глубине 5–10 см в почвах переходной к лесу 16-летней залежи и во вторичном 40-летнем лесу (0.28–0.30 мг СО2/(мг Смик сут)). Это связано не только с возрастанием запасов поступающего опада, доступного к быстрому разложению микроорганизмами, но и его высоким биохимическим разнообразием (листва деревьев, травянистые растения разных биоморфологических групп). Более высокие значения qCO2 в слое 5–10 см и глубже на стадиях зарастания с богатым травянистым покровом могут объясняться также наличием в этих слоях легкодоступной корневой мортмассы. Максимальное значение qCO2 (0.46 мг СО2/(мг Смик сут)) выявлено в слое 0–5 см в почве под смешанным старовозрастным лесом, по-видимому, благодаря высокому поступлению опада, в составе которого присутствуют соединения разной степени доступности. Для слоя 0–10 см выявлена тесная положительная корреляция (r = 0.71–0.84, р < 0.05) значений qCO2 с содержанием Сорг, Nобщ и отношением С/N и отрицательная – с величиной рНKCl (r = –0.74, р < 0.05) (табл. 4). Таким образом, относительно высокое содержание ОВ и достаточное поступление доступного для разложения растительного материала на средних стадиях сукцессии способствуют благоприятному экофизиологическому состоянию микробных сообществ и их активному функционированию в ходе постагрогенной сукцессии.
Постагрогенная динамика ферментативной активности почв. Ферментативная активность также является чувствительным индикатором постагрогенных изменений, обусловленных снятием сельскохозяйственной нагрузки [14, 56]. Активность ферментов гидролитической группы, в том числе и фосфатазы, обусловлена в значительной степени запасами ОВ [20], поэтому в пахотной почве и в почвах молодых залежей ФСФ-активность не высока (рис. 4a). Наиболее существенный рост активности ФСФ в ходе постагрогенной эволюции наблюдается в слое 0–5 см на стадии мелколиственного леса, а в старовозрастном лесу ФСФ-активность достигает своего максимума. В слое 5–10 см выявленная закономерность изменения активности ФСФ сохраняется, но при более низких ее значениях. В нижней части старопахотного слоя (10–30 см), где содержание Сорг невысоко, активность фосфатазы заметно снижена. Для слоя 0–10 см выявлена тесная положительная корреляция между ФСФ-активностью и содержанием Сорг и Nобщ (r = 0.99, р < 0.05), а также с величиной метаболического коэффициента (r = 0.83, р < 0.05) (табл. 4). Вместе с тем активность ФСФ отрицательно коррелирует с величиной рН (r = –0.73, р < < 0.05). Оценка линейных трендов изменения ФСФ активности за 40 лет постагрогенной сукцессии выявила статистически значимый рост этого показателя на всех глубинах старопахотного слоя (табл. 5). В слое 0–5 см ежегодный прирост активности ФСФ был более, чем в 2 раза интенсивнее, чем в других слоях старопахотного слоя, а длительность постагрогенного восстановления объясняла 90–98% дисперсии этого показателя.
Рис. 4.
Постагрогенная динамика активности ферментов (a – фосфатаза, ФСФ; b – полифенолоксидаза, ПФО; c – пероксидаза, ПРО; d – отношение ПФО/ПРО).

Постагрогенная динамика оксиредуктаз отличается от динамики фосфатазы, так как активность ферментов этой группы связана прежде всего с трансформацией ОВ, а именно с соотношением процессов его минерализации и гумификации. По мере восстановления растительности пероксидазная активность в слоях 0–5 см демонстрирует постепенный рост, достигая своего максимума в почвах молодого мелколиственного и старовозрастного смешанного леса (рис. 4c). В более глубоких слоях старопахотного слоя активность ПРО в ходе постагрогенной эволюции также демонстрирует общую тенденцию роста, но в почве старовозрастного леса резко уменьшается. Это может быть связано с тем, что в более глубоких слоях подзола, сформированного под старовозрастным лесом, ОВ практически не накапливается. Более низкая активность ПРО в почвах пашни по сравнению залежами объясняется тем, что того пула поступающих с пожнивными остатками органических веществ явно не хватает для протекания активных минерализационных процессов. И только при переходе к 40-летнему мелколиственному лесу, по-видимому, накапливается достаточный запас ОВ и ПРО-активность резко увеличивается. Для слоев 0–5 и 5–10 см обнаружена положительная корреляция между ПРО-активностью со скоростью БД, величиной qCO2 и ПФО-активностью (r = 0.78, р < 0.05) (табл. 4). Выявленные взаимосвязи, по-видимому, объясняются тем, что изменение вышеперечисленных параметров определяется общим фактором, которым скорее всего является количество поступающего в почву легкоразлагаемого субстрата.
Постагрогенная динамика активности ПФО более сложная, чем ПРО, при том что корреляция между ними достаточно тесная (r = 0.78, р < 0.05) (табл. 4). В ходе восстановительной сукцессии в слоях 0–5 и 5–10 см наблюдаются два существенных пика ПФО активности – на стадии молодого высокопродуктивного луга (11 лет) и на стадии мелколиственного леса (40 лет). Спад полифенолоксидазной активности в почве на залежи 16 лет обусловлен, по-видимому, неким временным разрывом между высокопродуктивным травяным фитоценозом и лесным фитоценозом, что создает некоторый дефицит поступления с опадом легко доступных разложению веществ и снижает активность их трансформации. Кроме того, фенольные соединения, содержащиеся в листьях ивы, могут ингибировать процессы разложения опада [55].
Оценка линейных трендов изменения активности ПРО и ПФО за 40 лет постагрогенной сукцессии выявила статистически значимый рост этих показателей во всех слоях старопахотного горизонта за исключением глубины 10–20 см для ПФО-активности и 5–10 и 20–30 см для активности ПРО (табл. 5). Скорость увеличения ПРО-активности была более высокой в слое 0–5 см, в то время как ПРО активность демонстрировала сходную скорость прироста во всей старопахотной толще.
По мнению некоторых исследователей [2, 8], отношение активностей полифенолоксидазы и пероксидазы (ПФО/ПРО) может служить индикатором степени преобладания процессов гумификации над минерализацией, то есть использоваться как коэффициент гумификации. В изучаемом хроноряду четко выраженного тренда данного показателя не выявлено (рис. 4d), однако коэффициент ПФО/ПРО на глубине 0–5 см имеет минимальное значение (0.51) на стадии 16-летней залежи, а максимальное (1.31) – на стадии 40-летнего леса. На глубине 5–10 см минимальное значение ПФО/ПРО соответствует пашне, а максимальное (2.05) – залежи 11 лет. Для почвы старовозрастного леса на обеих глубинах отношение ПФО/ПРО составляет около 1.0. Это говорит о том, что при недостаточном поступлении опада процессы гумификации выражены сильнее, чем процесс минерализации, что может иметь место на пашне, где субстратом для разложения являются лишь пожнивные остатки. Максимальные величины отношение ПФО/ПРО показывает в слоях 5–10 и 10–20 см на стадии высокопродуктивного луга, что можно объяснить более активной гумификацией подземной фракции биомассы травяных растений, которая на этой стадии максимальна (табл. 2). В почвах 16-летней залежи с древостоем из ивы козьей и довольно хорошо выраженным травяным ярусом, отношение ПФО/ПРО демонстрирует одни из самых низких значений (рис. 4d). Ранее, используя в качестве индекса гумификации величину HIX1, оцененную с помощью спектров флуоресценции водных вытяжек из почв, мы получили в некоторой степени сходные закономерности [55]. Было показано, что индекс гумификации водорастворимого почвенного вещества в ходе естественного лесовосстановления снижается, принимая самые низкие значения на стадии, где сформировался древостой из ивы козьей, опад которой богат фенольными соединениями, способными ингибировать процессы активного преобразования органических субстратов. Наши результаты показали проблематичность и дискуссионность применения отношения ПФО/ПРО в качестве индекса гумификации. Необходимо применение других инструментальных методов, таких как твердофазная 13С-ЯМР-спектроскопия, позволяющих более адекватно судить о степени преобразованности ОВ почв в ходе постагрогенного развития.
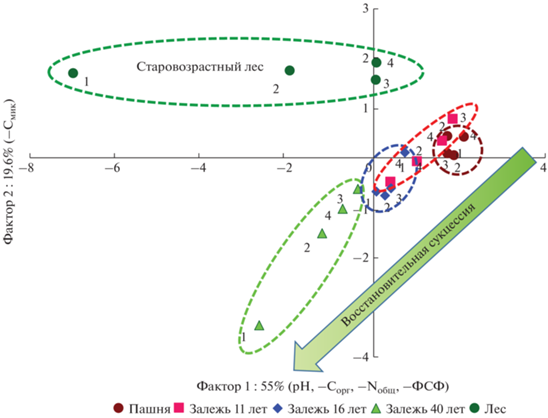
Чтобы интегрировать и визуализировать полученную в работе информацию по изменению широкого спектра свойств на четырех различных глубинах старопахотного горизонта в пяти объектах, составляющих хроноряд залежных почв, мы использовали метод главных компонент. С учетом всей совокупности изученных свойств отчетливо разделяется старовозрастный смешанный и молодой мелколиственный лес, а также пахотная почва и залежь 16 лет (рис. 5). Самая молодая 11-летняя залежь занимает промежуточное положение между пашней и 16-летней залежью. Фактор 1, который наиболее сильно коррелирует с содержанием Сорг (r = –0.91) и Nобщ (r = –0.90), активностью фосфатазы (r = –0.93) и величиной рН (r = = 0.76), описывает 55.0% общей информации, а ось ординат четко отделяет почвы лесов от почв пашни и более молодых залежей, на которых преобладает луговая растительность. Фактор 2, который наиболее тесно коррелирует с содержанием Смик (r = –0.79), ответственен только за 19.6% выявленных закономерностей. Таким образом в зоне южной тайги в ходе восстановительной залежной сукцессии в пределах всего старопахотного горизонта происходит последовательное увеличение содержания ОВ, и как следствие – усиление их микробной и ферментативной активности, которое наиболее выражено в верхнем (0–10 см) слое почвы старовозрастного леса, представляющего субклимаксную стадию постагрогенного лесовосстановления, находятся в состоянии динамического равновесия, для достижения которого постагрогенным почвам может потребоваться еще не один десяток лет.
Рис. 5.
Распределение объектов исследования по методу главных компонент с использованием значений различных почвенных характеристик на разных глубинах: 1 – 0–5 см, 2 – 5–10 см, 3 – 10–20 см, 4 – 20–30 см. Свойства: рН – потенциальная кислотность почв; Сорг – содержание органического углерода, Nобщ – содержание общего азота, Смик – содержание микробной биомассы, ФСФ – активность фосфатазы (мг Р2О5/(100 г почвы ч)).
ЗАКЛЮЧЕНИЕ
Естественное лесовосстановление на бывших пахотных почвах в подзоне южной тайги сопровождается увеличением поступления растительного опада, что приводит к значительному росту содержания органического углерода и общего азота в верхней части старопахотного слоя с одновременным уменьшением значений pHKCl. Наряду с этим, в ходе постагрогенной сукцессии наблюдается заметное усиление стратификации содержания Сорг и Nобщ в слое 0–30 см, особенно выраженное на стадии образования сомкнутого древостоя и свидетельствующее о восстановлении углерод-секвестрирующей функции почв на более поздних стадиях постагрогенного развития. Самые высокие значения СтО (0–5 : 5–10, 0–5 : 10–20 и 0–5 : 20–30) наблюдаются в почве старовозрастного леса, принципиально отличающегося строением органопрофиля.
Обогащение почв органическим веществом индуцирует заметное усиление микробной активности, которое выражается в увеличении скорости базального дыхания и содержания микробной биомассы. Наиболее заметные изменения этих показателей в ходе восстановительной сукцессии проявляются в верхнем (0–10 см) почвенном слое, а их максимальные значения наблюдается в почве 40-летней залежи, соответствующей мелколиственному лесу, где легкоразлагаемый опад обеспечивает наличие большого запаса доступного субстрата для микробного сообщества. Микробный метаболический коэффициент также постепенно возрастает в течение 40-летней восстановительной сукцессии бывших пахотных почв, что связано не только с возрастанием запасов поступающего опада, доступного к быстрому разложению микроорганизмами, но и его высоким биохимическим разнообразием. Отношение базального дыхания к содержанию Сорг, характеризующее доступность ОВ разложению микроорганизмами, в верхнем (0–10 см) слое бывшего пахотного горизонта в течение первых 40 лет восстановительной сукцессии увеличивается в 2–4 раза и тесно коррелирует с содержанием Смик.
Большую роль в трансформации органического вещества играют почвенные ферменты, как гидролитической группы, так и оксидоредуктазы. Активность фосфатазы в ходе постагрогенного восстановления возрастает многократно, особенно в верхнем (0–10 см) слое пахотного горизонта, что в первую очередь связано с возрастанием содержания органического вещества и подтверждается тесной положительной корреляцией между этими показателями. Активность оксиредуктаз в ходе постагрогенной сукцессии не имеет четко выраженного тренда, однако ее пики, соответствующие стадии высокопродуктивного луга (пероксидаза) и молодого мелколиственного леса (фенолоксидазы), могут быть связаны с повышенным поступлением легкоразлагаемого растительного опада с разным биохимическим составом – травяного либо мелколиственного.
За 40 лет постагрогенной сукцессии наблюдается наиболее значительное увеличение практически всех рассматриваемых показателей в слое 0–5 см. В более глубоких слоях изменение анализируемых почвенных свойств в ходе восстановительной сукцессии, как правило, становится менее выраженным. Таким образом, предположения, которые мы выдвигали в начале работы полностью подтвердились.
Список литературы
Архив климатических данных [Электронный ресурс]. URL: http:// climatbase.ru/ дата обращения 04.06.2020.
Аюпов З.З., Сидорова Л.В., Анохина Н.С., Миннебаева И.Ф., Гареев Н.И. Органическое вещество и ферментативная активность чернозема выщелоченного в зависимости от приемов основной обработки почвы и удобрения // Вестник БГАУ. 2010. № 2. С. 11–16.
Благодатская Е.В., Ананьева Н.Д., Мякшина Т.Н. Характеристика состояния микробного сообщества по величине метаболического коэффициента // Почвоведение. 1995. № 2. С. 205–210.
Болысов С.И., Фузеина Ю.Н. Физико-географические условия Костромского Заволжья. Геолого-геоморфологическое устройство // Костромское Заволжье: природа и человек. М., 2001. С. 36–60.
Замолодчиков Д.Г., Уткин А.И., Коровин Г.Н. Определение запасов углерода по зависимым от возраста насаждений конверсионно-объемным коэффициентам // Лесоведение. 1998. № 3. С. 84–93.
Иванова А.Е., Николаева В.В., Марфенина О.Е. Изменение целлюлозолитической активности городских почв в связи с изъятием растительного опада (на примере Москвы) // Почвоведение. 2015. № 5. С. 562–570.
Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 341 с.
Коваленко М.В., Марковская Г.К. Влияние способов основной обработки почвы на ее ферментативную активность // Вестник Казанского ГАУ. 2013. № 1(27). С. 108–111.
Литвинович А.В., Павлова О.Ю., Плылова И.А. Изменение состава растительного покрова, содержания и запасов гумуса окультуренной дерново-подзолистой глееватой суглинистой почвы в процессе постагрогенной эволюции // Изв. С.-Пб. гос. аграр. ун-та. 2011. № 22. С. 74–77.
Литвинович А.В., Павлова О.Ю., Чернов Д.В. Изменение гумусового состояния дерново-подзолистой почвы при прекращении антропогенного воздействия // Доклады РАСХН. 2002. № 6. С. 26–28.
Литвинович А.В., Плылова И.А. Изменение кислотно-основных свойств дерново-подзолистой суглинистой почвы в процессе постагрогенной эволюции // Научное обеспечение развития АПК в условиях реформирования. СПб., 2009. С. 160–164.
Люри Д.И., Горячкин С.В., Караваева Н.А., Денисенко Е.А., Нефедова Т.Г. Динамика сельскохозяйственных земель России в ХХ веке и постагрогенное восстановление растительности и почв. М.: ГЕОС, 2010. 416 с.
Матинян Н.Н., Бахматова К.А., Алексеев С.С. Постагрогенная трансформация почв, сформированных на контрастных по гранулометрическому составу породах // Гумус и почвообразование. СПб–Пушкин, 2007. С. 52–60.
Овсепян Л.А., Курганова И.Н., Мостовая А.С., Лопес де Гереню В.О., Личко В.И., Благодатская Е.В., Кузяков Я.В. Ферментативная активность постагрогенных серых лесных почв нагорной дубравы “Лес на Ворскле” // Известия Самарского научного центра Российской академии наук. 2017. Т. 19. № 2. С. 151–158.
Огуреева Г.Н. Ботанико-географическое районирование СССР. М.: Изд-во Моск. ун-та, 1991. 76 с.
Романовская А.А., Коротков В.Н., Карабанов Р.Т., Смирнов Н.С. Динамика элементов баланса углерода на неиспользуемых пахотных угодьях Валдайской возвышенности // Экология. 2012. № 5. С. 347–352. https://doi.org/10.1134/S106741361204011X
Рыжова И.М., Ерохова А.А., Подвезенная М.А. Изменение запасов углерода в постагрогенных экосистемах в результате естественного восстановления лесов в Костромской области // Лесоведение. 2015. № 4. С. 307–317.
Семенов В.М., Когут Б.М. Почвенное органическое вещество. М.: ГЕОС, 2015. 233 с.
Телеснина В.М., Богатырев Л.Г., Бенедиктова А.И., Земсков Ф.И., Маслов М.Н. Динамика поступления растительного опада и некоторых свойств лесных подстилок при постагрогенном лесовосстановлении в условиях южной тайги // Вестник. Моск. ун-та. Сер. 17. Почвоведение. 2019. № 4. С. 3–10.
Хазиев Ф.Х. Методы почвенной энзимологии. М.: Наука, 2005. 251 с.
Хомутова Т.Э., Демкина Т.С., Каширская Н.Н., Демкин Фосфатазная активность современных и погребeнных каштановых почв Волго-Донского междуречья // Почвоведение. 2012. № 4. С. 478–483.
Шеин Е.В., Карпачевский Л.О. Теории и методы физики почв. М.: Гриф и К, 2007. 616 с.
Якушев А.В., Кузнецова И.Н., Благодатская Е.В., Благодатский С.А. Зависимость активности полифенолпероксидаз и полифенолоксидаз в современных и погребенных почвах от температуры // Почвоведение. 2014. № 5. С. 1–7.
Anderson J.P.E., Domsch K.H. A phisiological method for the quantitative measurement of microbial biomass in soils // Soil Biol. Biochem. 1978. V. 10. № 3. P. 215–221.
Anderson T.-H. Physiological analysis of microbial communities in soil: Applications and limitations // Beyond the Biomass. London: Wiley-Sayce Publication, 1994. P. 67–76.
Anderson T.-H., Domsch K.H. The metabolic quotient for CO2 (qCO2) as a specific activity parameter to assess the effects of environmental conditions, such as pH, on the microbial biomass of forest soils // Soils Biol. Biochem. 1993. V. 25. P. 393–395.https://doi.org/10.1016/0038-0717(93)90140-7
Aponte H., Medina J., Butler B., Meier S., Cornejo P., Kuzyakov Y. Soil quality indices for metal(loid) contamination: An enzymatic perspective // Land Degradation & Development. 2020. V. 31. P. 2700–2719.
Azarenko M.A., Kazeev K.Sh., Yermolaeva O.Yu., Kolesnikov S.I. Changes in the plant cover and biological properties of Chernozems in the postagrogenic period // Eurasian Soil Science. 2020. № 11. P. 1645–1654. https://doi.org/10.1134/S1064229320110034
Badiane N.N.Y., Chotte J.L., Pate E., Masse D., Rouland C. Use of soil enzyme activities to monitor soil quality in natural and improved fallows in semiarid tropical regions // Applied Soil Ecology. 2001. V. 18. P. 229–238.
Bollag J.M., Dec J., Huang P.M. 1998. Formation mechanisms of complex organic structures in soil habitats // Adv. Agron. V. 63. P. 237–265.
Burns R.G., DeForest J.L., Marxsen J., Sinsabaugh R.L., Stromberger M.E., Wallenstein M.D., Weintraub M.N., Zoppini A. Soil enzymes in a changing environment: Current knowledge and future directions // Soil Biology and Biochemistry. 2013. V. 58. P. 216–234.
Dick R.P., Myrold D.D., Kerle E.A. Microbial Biomass and Soil Enzyme Activities in Compacted and Rehabilitated Skid Trail Soils // Soil Sci. Soc. Am. J. 1988. V. 52. P. 512–516. https://doi.org/10.2136/sssaj1988.03615995005200020038x
Duarte-Guardia S., Peri P., Amelung W., Thomas E., Borchard N., Baldi G., Cowie A., Ladd B. Biophysical and socioeconomic factors influencing soil carbon stocks: a global assessment // Mitigation and Adaptation Strategies for Global Change. 2020. V. 25(6). https://doi.org/:10.1007/s11027-020-09926-1
Eivazi F., Tabatabai M.A. Factors affecting glucosidase and galactosidase activities in soils // Soil Biology and Biochemistry. 1990. V. 22. P. 891–897. https://doi.org/10.1016/0038-0717(90)90126-K
Falkengren-Grerup U., ten Brink D.-J., Brunet J. Land use effects on soil N, P, C and pH persist over 40–80 years of forest growth on agricultural soils // Forest Ecol. Manag. 2006. V. 225. № 1–3. P. 74–81.https://doi.org/10.1016/j.foreco.2005.12.027
Floch C., Alarcon-Gutiérrez E., Criquet S. ABTS assay of phenol oxidase activity in soil // J. Microbiological Methods. 2007. V. 71. P. 319–324.
German D.P., Marcelo K.R.B., Stone M.M., Allison S.D. The Michaelis-Menten kinetics of soil extracellular enzymes in response to temperature: a cross-latitudinal study // Global Change Biology. 2012. V. 18. P. 1468–1469.
Guillaume T., Muhammad D., Kuzyakov Y. Losses of soil carbon by converting tropical forest to plantations: Erosion and decomposition estimated by δ13C // Global Change Biology 2015. V. 21. P. 3548–3560.
Guo L.B., Gifford R.M. Soil carbon stocks and land use change: a metaanalysis // Global Change Biology. 2002. V. 8. P. 345–360. https://doi.org/10.1046/j.1354-1013.2002.00486x
Ivashchenko K.V., Ananyeva N.D., Vasenev V.I., Kudeyarov V.N., Valentini R. Biomass and respiration activity of soil microorganisms in anthropogenically transformed ecosystems (Moscow region) // Eurasian Soil Science. 2014. V. 47. № 9. P. 892–903. https://doi.org/10.1134/S1064229314090051
Johnsen A.R., Jacobsen O.S. A quick and sensitive method for the quantification of peroxidase activity of organic surface soil from forests // Soil Biology & Biochemistry. 2008. V. 40. P. 814–821.
Kalinina O., Chertov O., Dolgikh A.V., Goryachkin S.V., Lyuri D.I., Vormstein S., Giani L. Self-restoration of post-agrogenic Albeluvisols: Soil development, carbon stocks and dynamics of carbon pools // Geoderma. 2013. V. 207–208. P. 221–233. https://doi.org/10.1016/j.geoderma.2013.05.019
Kalinina O., Goryachkin S.V., Karavaeva N.A., Lyuri D.I., Giani L. Dynamics of carbon pools in post-agrogenic sandy soils of southern taiga of Russia // Carbon Balance and Management. 2010. V. 5. № 1. P. 1–9. https://doi.org/10.1186/1750-0680-5-1
Kalinina O., Goryachkin S.V., Lyuri D.I., Giani L. Post-agrogenic development of vegetation, soils, and carbon stocks under self-restoration in different climatic zones of European Russia // Catena. 2015. V. 129. P. 18–29. https://doi.org/10.1016/j.catena.2015.02.016
Kalinina O., Goryachkin S.V., Karavaeva N.A., Lyuri D.I., Najdenko L., Giani L. Self-restoration of post-agrogenic sandy soils in the southern Taiga of Russia: Soil development, nutrient status, and carbon dynamics // Geoderma. 2009. V. 152. P. 35–42. https://doi.org/10.1016/j.geoderma.2009.05.014
Kalinina O.Yu., Chertov O., Frolov P., Goryachkin S., Kuner P., Küper J., Giani L. Alteration process during the post-agricultural restoration of Luvisols of the temperate broad-leaved forest in Russia // Catena. 2018. V. 171. P. 602–612. https://doi.org/10.1016/j.catena.2018.08.004
Kämpf I., Hölzel N., Störrle M., Broll G., Kiehl K. Potential of temperate agricultural soils for carbon sequestration: A meta-analysis of land-use effects // Science of the Total Environment. 2016. V. 566–567. P. 428–435. https://doi.org/10.1016/j.scitotenv.2016.05.067
Kechaikina I.O., Ryumin A.G., Chukov S.N. Postagrogenic transformation of organic matter in soddy-podzolic soils // Eurasian Soil Science. 2011. V. 44. № 10. P. 1077–1089. https://doi.org/10.1134/S1064229311100061
Kooch Y., Ehsanib S., Akbariniac M. Stratification of soil organic matter and biota dynamics in natural andanthropogenic ecosystems // Soil and Tillage Research. 2020. V. 200. P.104621. https://doi.org/10.1016/j.still.2020.104621
Kurganova I., Lopes de Gerenyu V., Kuzyakov Y., Six J. Carbon cost of collective farming collapse in Russia // Global Change Biology. 2014. V. 20. № 3. P. 938–947. https://doi.org/10.1111/gcb.12379
Kurganova I., Merino A., Lopes de Gerenyu V., Barros N., Kalinina O., Giani L., Kuzyakov Y. Climate dependent mechanisms of carbon sequestration and stabilization by restoration of arable soils after abandonment // Geoderma. 2019. V. 354. P. 113882. https://doi.org/10.1016/j.geoderma.2019.113882
Kurganova I.N., Lopes de Gerenyu V.O., Gallardo Lancho J.F., Oehm C.T. Evaluation of the rates of soil organic matter mineralization in forest ecosystems of temperate continental, Mediterranean, and tropical monsoon climates // Eurasian Soil Sci. 2012. V. 45. № 1. P. 68–79. https://doi.org/10.1134/S1064229312010085
Kurganova I.N., Lopes de Gerenyu V.O., Mostovaya A.S., Ovsepyan L.A., Telesnina V.M., Lichko V.I., Baeva Y.I. The Effect of Reforestation on Microbial Activity in Post-Agrogenic Soils in European Part of Russia // Contemporary Problem of Ecology. 2018. № 7. P. 7–17. https://doi.org/10.1134/S1995425518070089
Kurganova I.N., Semenov V.M., Kudeyarov V.N. Climate and Land Use as Key Factors of the Stability of Organic Matter in Soils // Doklady Biological Sciences. 2019. V. 489. P. 189–192.
Kurganova I.N., Telesnina V.M., Lopes de Gerenyu V.O., Lichko V.I., Karavanova E.I. The Dynamics of Carbon Pools and Biological Activity of Retic Albic Podzols in Southern Taiga during the Postagrogenic Evolution // Eurasian Soil Science. 2021. V. 54. № 3. P. 337–351.https://doi.org/10.1134/S1064229321030108
Ovsepyan L., Kurganova I., Lopes de Gerenyu V., Kuzyakov Ya. Conversion of cropland to natural vegetation boosts microbial and enzyme activities in soil // Science of the Total Environment. 2020. V. 743. 140829 https://doi.org/10.1016/j.scitotenv.2020.140829
Ovsepyan L., Kurganova I., Lopes de Gerenyu V., Kuzyakov Ya. Recovery of organic matter and microbial biomass after abandonment of degraded agricultural soils: the influence of climate // Land Degr. Dev. 2019. V. 30. P. 1861–1874. https://doi.org/10.1002/ldr.3387
Ovsepyan L.A., Kurganova I.N., Lopes de Gerenyu V.O., Rusakov A.V., Kuzyakov Ya.V. Changes in the Fractional Composition of Organic Matter in the Soils of the Forest–Steppe Zone during Their Postagrogenic Evolution // Eurasian Soil Science. 2020. V. 53. № 1. P. 50–61. https://doi.org/10.1134/S1064229320010123
Poeplau C., Don A., Vesterdal L., Leifeld J., Van Wesemael B., Schumacher J., Gensior A. Temporal dynamics of soil organic carbon after land-use change in the temperate zone – carbon response functions as a model approach: soil organic carbon and land-use change // Glob. Change Biol. 2011. V. 17. P. 2415–2427. https://doi.org/10.1111/j. 1365-2486.2011.02408.x
Post W.M., Kwon K.C. Soil carbon sequestration and land-use change: processes and potential // Global change biology. 2000. V. 6. P. 317–328. https://doi.org/10.1046/j.1365-2486.2000.00308.x
Ryzhova I.M., Telesnina V.M., Sitnikova A.A. Dynamic of soil properties and carbon stock structure in postagrogenic ecosystems of Southern taiga during natural reforestation // Eurasian Soil Science. 2020. V. 53. № 2. P. 230–243. https://doi.org/10.1134/S0032180X20020100
Ryzhova I.M., Erokhova A.A., Podvezennaya M.A. Dynamics and Structure of Carbon Storage in the Postagrogenic Ecosystems of the Southern Taiga // Eurasian Soil Science. 2014. V. 47. № 12. P. 1207–1215. https://doi.org/10.1134/S1064229314090117
Sá J.C.M., Lal R. Stratification ratio of soil organic matter pools as an indicator of carbon sequestration in a tillage chronosequence on a Brazilian Oxisol // Soil and Tillage Research. 2009. V. 103. № 1. P. 46–56. https://doi.org/10.1016/j.still.2008.09.003
Smith P. Land use change and soil organic carbon dynamics // Nutr Cycl Agroecosyst. 2008. V. 81. P. 169–178. https://doi.org/10.1007/s10705-007-9138-y
Soussana J.-F., Loiseau P., Vuichard N., Ceschia E., Balesdent J., Chevallier T., Arrouays D. Carbon cycling and sequestration opportunities in temperate grasslands // Soil Use and Management. 2004. V. 20. P. 219–230.
Spohn M., Novák T.J., Incze J., Giani L. Dynamics of soil carbon, nitrogen, and phosphorus in calcareous soils after land-use abandonment – A chronosequence study // Plant and Soil. 2016. V. 401. № 1–2. P. 185–196. https://doi.org/10.1007/s11104-015-2513-6
Stockmann U., Padarian J., McBratney A., Minasny B., de Brogniez D., Montanarella L., Hong S.Y., Rawlins B.G., Field D.J. Global soil organic carbon assessment // Glob Food Sec. 2015. V. 6. P. 9–16. https://doi.org/10.1016/j.gfs.2015.07.001
Stolnikova E.V., Ananyeva N.D., Chernova O.V. The Microbial Biomass and Its Activity and Structure in the Soils of Old Forests In the European Russia // Eurasian Soil Science. 2011. V. 44. № 4. P. 437–452. https://doi.org/10.1134/S1064229311040107
Susyan E.A., Wirth S., Ananyeva N.D., Stolnikova E.V. Forest succession on abandoned arable soils in European Russia – Impacts on microbial biomass, fungal-bacterial ratio, and basal CO2 respiration activity // Eur. J. Soil Biol. 2011. V. 47. P. 169–174. https://doi.org/10.1016/j.ejsobi.2011.04.002
Swift M.J., Heal O.W., Anderson J.M. Decomposition in terrestrial ecosystems, Studies in ecology. University of California Press, Berkeley, 1979.
Telesnina V.M., Kurganova I.N., Lopes de Gerenyu V.O., Ovsepyan L.A., Lichko V.I., Yermolaev A.M., Mirin D.M. Dynamics of soil properties and plant composition during postagrogenic evolution in different bioclimatic zones // Eurasian Soil Science. 2017. V. 50. № 12. P. 1515–1534. https://doi.org/10.1134/S1064229317120109
Telesnina V.M., Vaganov I.E., Karlsen A.A., Ivanova A.E., Zhukov M.A., Lebedev S.M. Specific features of the morphology and chemical properties of coarse-textured postagrogenic soils of the southern taiga, Kostroma oblast // Eurasian Soil Science. 2016. V. 49. № 1. P. 102–115. https://doi.org/10.1134/S1064229316010117
Telesnina V.M., Zhukov M.A. The Influence of Agricultural Land Use on the Dynamics of Biological Cycling and Soil Properties in the Course of Postagrogenic Succession (Kostroma Oblast) // Eurasian Soil Science. 2019. V. 52. № 9. P. 1114–1129. https://doi.org/10.1134/S1064229319070135
Titlyanova A.A., Bulavko G.I., Kudryashova S.Ya., Naumov A.V., Smirnov V.V., Tanasienko A.A. Stocks and losses of organic carbon in soils of Siberia // Eurasian Soil Sci. 1998. V. 1. P. 51–59.
Titlyanova A.A., Naumov A.V. Carbon losses from soils of Western Siberia under agriculture // Eurasian Soil Sci. 1995. V. 11. P. 1357–1362.
Vladychenskii A.S., Telesnina V.M., Rumyantseva K.A., Chalaya T.A. Organic Matter and Biological Activity of Postagrogenic Soils in the Southern Taiga Using the Example of Kostroma Oblast // Eurasian Soil Science. 2013. V. 46. № 5. P. 518–529. https://doi.org/10.1134/S1064229313050141
Wallenstein M.D., McMahon S.K., Schimel J.P. Seasonal variation in enzyme activities and temperature sensitivities in Arctic tundra soils // Global Change Biol. 2009. V. 15. P. 1631–1639. https://doi.org/10.1111/j.1365-2486.2008.01819.x
Wang B., Liu G.B., Xue S., Zhu B. Changes in soil physicochemical and microbiological properties during natural succession on abandoned farmland in the Loess Plateau // Environmental Earth Sciences. 2011. V. 62. № 5. P. 915–925. https://doi.org/10.1007/s12665-010-0577-4
Wiesmeier M., Urbanski L., Hobley E., Lang B., von Lützow M., Marin-Spiotta E., van Wesemael B., Rabot E., Ließ M., Garcia-Franco N., Wollschläger U., Vogel H.J., Kögel-Knabner I. Soil organic carbon storage as a key function of soils – a review of drivers and indicators at various scales // Geoderma. 2019. V. 333. P. 149–162. https://doi.org/10.1016/j.geoderma.2018.07.026
Zavarzina A.G. Heterophase synthesis of humic acids in soils by immobilized phenol oxidases // Soil Enzymology. 2011. Springer Verlag Berlin Heidelberg. P. 207–228.
Zhang L., Chen W., Burger M., Yang L., Gong P., Wu Z. Changes in Soil Carbon and Enzyme Activity As a Result of Different Long-Term Fertilization Regimes in a Greenhouse Field // PLoS ONE. 2015. V. 10. e0118371. https://doi.org/10.1371/journal.pone.0118371
Zhao F., Yang G., Han X., Feng Y., Ren G. Stratification of Carbon Fractions and Carbon Management Index in Deep Soil Affected by the Grain-to-Green Program in China // PLoS ONE. 2014. V. 9. № 6. https://doi.org/10.1371/journal.pone.0099657
Дополнительные материалы отсутствуют.