

Поверхность. Рентгеновские, синхротронные и нейтронные исследования, 2021, № 11, стр. 101-103
Исследование изменения вязкости постсинаптической мембраны при изменении ее состава методом молекулярной динамики
a Институт кристаллографии им. А.В. Шубникова ФНИЦ “Кристаллография и фотоника” РАН
119333 Москва, Россия
b Национальный исследовательский центр “Курчатовский институт”
123182 Москва, Россия
* E-mail: tostars@mail.ru
Поступила в редакцию 12.01.2021
После доработки 04.03.2021
Принята к публикации 07.03.2021
Аннотация
Методом молекулярного моделирования построены четыре модели постсинаптической мембраны с различным содержанием холестерина. Исследована молекулярная динамика построенных моделей. Прослежена зависимость подвижности атомов липидов от изменения концентрации холестерина. Показано, что увеличение концентрации холестерина приводит к увеличению вязкости липидного бислоя. Проведено сравнение с опубликованными экспериментальными данными и показано, что полученные результаты соответствуют им. Таким образом, полученные модели могут быть использованы для изучения структурных основ влияния липидного состава постсинаптической мембраны на работу постсинаптических рецепторов.
ВВЕДЕНИЕ
Передача сигналов между нейронами центральной нервной системы осуществляется путем активации рецепторов постсинаптических мембран молекулами различных медиаторов [1]. Основными рецепторами такой мембраны являются глутаматные AMPA- и NMDA-рецепторы, никотиновые и мускариновые рецепторы ацетилхолина, катехоламиновые рецепторы, серотониновые и дофаминовые рецепторы, рецепторы γ-аминомасляной кислоты. Известно, что на функцию белков-рецепторов существенным образом влияет вязкость постсинаптической мембраны [2]. Постсинаптическая мембрана состоит главным образом из липидов и белков. Липидная составляющая мембраны представлена прежде всего фосфатидилэтаноламином, фосфатидилхолином, фосфaтидилсерином и фосфатидилинозитолом в определенной концентрации [3]. Также существенную роль в функционировании постсинаптической мембраны играет холестерин, от которого главным образом зависит ее вязкость [4]. Известно, что ряд патологий и синдромов, связанных с нарушением функции рецепторов постсинаптической мембраны, связан с изменением вязкости самой мембраны. В качестве примеров можно привести возрастное снижение когнитивных способностей [5], болезнь Паркинсона [6], разного рода нарушения сна [7]. Изучению причин изменения вязкости постсинаптической мембраны посвящен ряд работ [2, 8, 9], однако на структурном уровне данное явление не было хорошо изучено. Настоящая работа посвящена получению моделей постсинаптической мембраны с различным содержанием холестерина (18, 20, 22 и 24%) и исследованию динамики полученных моделей. В работе прослежено влияние изменения концентрации холестерина на подвижность атомов липидов, составляющих мембрану, и показано, что увеличение концентрации холестерина приводит к уменьшению подвижности атомов липидов и, следовательно, к увеличению вязкости липидного бислоя. Полученная модель может быть использована для изучения структурных основ влияния липидного состава постсинаптической мембраны на работу постсинаптических рецепторов.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Начальные модели постсинаптической мембраны, содержащие 100 молекул с каждой из сторон липидного бислоя, построены с использованием интернет-сервиса CHARMM-GUI [21]. Согласно [3] основными составляющими постсинаптической мембраны являются фосфатидилэтаноламин (30%), фосфатидилхолин (34%), фосфaтидилсерин (14%) и фосфатидилинозитол (4%) и холестерин (18%). Были построены четыре модели мембраны с увеличенным содержанием холестерина. Состав моделей приведен в табл. 1. Кроме того, каждая из моделей была помещена в водный раствор (20 Å с каждой стороны бислоя), а также к системе было добавлено 0.15 M KCl. Моделирование методом молекулярной динамики проводилось с использованием программного пакета AMBER [10] c CUDA-версией pmemd [11]. В процессе моделирования использовали силовое поле CHARMM36 [12]. В качестве модели воды брали модель TIP3P. Для релаксации структуры и во избежание стерических препятствий при дальнейшей симуляции была проведена минимизация энергии системы. Затем система была уравновешена в шесть стадий по 125–500 пс каждая при температуре 310 К и давлении 1 атм в ансамблях NVT и NPT с ограничениями на подвижность атомов. Температуру и давление в системе контролировали с помощью термостата Ланжевена [13] и полуизотропного баростата Монте-Карло [14] с двумя поверхностями. В результате было промоделировано 100 нс с шагом 2 фс для каждой из четырех систем. Для получения достаточных статистических данных каждый расчет был повторен три раза.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ И ВЫВОДЫ
В результате работы построены четыре модели бислоя, сходного по липидному составу с постсинаптической мембраной. Модели отличаются различным содержанием холестерина, которое составляет 18, 20, 22 и 24%. Для того чтобы оценить влияние изменения концентрации холестерина на вязкость липидного бислоя, были рассчитаны среднеквадратичные флуктуации атомов липидов, которые удобно использовать для оценки их подвижности при разной концентрации холестерина в бислое. В каждом из вычислительных экспериментов среднеквадратичные флуктуации рассчитывали для всех атомов, после чего вычисляли среднее значение для систем с каждой из четырех различных концентраций холестерина (рис. 1). Из рисунка видно, что присутствует четкая тенденция снижения подвижности атомов липидов при повышении концентрации холестерина в составе бислоя.
Рис. 1.
Зависимость подвижности атомов липидного бислоя (RMSF – Root Mean Square Fluctuation), имитирующего постсинаптическую мембрану, от концентрации холестерина.
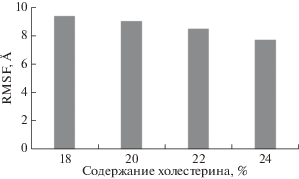
Полученные результаты показывают, что даже относительно небольшое увеличение концентрации холестерина в составе бислоя приводит к повышению его вязкости.
Список литературы
Белоконева О.С., Зайцев С.В. // Биохимия. 1992. Т. 58. С. 1685.
Dotti C.G., Esteban J.A., Ledesma M.D. // Front. Neuroanat. 2014. V. 8. P. 76. https://doi.org/10.3389/fnana.2014.00076
Cotman C.W., Blank M.L., Moehl A., Snyder F. // Biochemistry. 1969. V. 8. P. 4606. https://doi.org/10.1021/bi00839a056
Renner M., Choquet D., Triller. A. // J. Neurosci. 2009. V. 29. P. 2926. https://doi.org/10.1523/JNEUROSCI.4445-08.2009
Lynch M.A. // Physiol. Rev. 2004. V. 84. P. 87. https://doi.org/10.1152/physrev.00014.2003
Samii A., Nutt J.G., Ransom B.R. // Lancet. 2004. V. 363. P. 1783. https://doi.org/10.1016/S0140-6736(04)16305-8
Van Maanen A., Meijer A.M, van der Heijden K.B., Oort F.J. // Sleep Med. Rev. 2016. V. 29. P. 52 https://doi.org/10.1016/j.smrv.2015.08.009
Unwin N. // IUCrJ. 2020. V. 7. P. 852. https://doi.org/10.1107/S2052252520009446
Tulodziecka K., Diaz-Rohrer B.B., Farley M.M., Chan R.B., Di Paolo G., Levental K.R., Waxham M.N., Levental I. // Mol. Biol. Cell. 2016. V. 27. P. 3480. https://doi.org/10.1091/mbc.E16-06-0420
Case D.A., Cheatham III T.E., Darden T. et al. // J. Comput. Chem. 2005. V. 26. № 16. P. 1668.
Salomon-Ferrer R., Götz A. W., Poole D. et al. // J. Chem. Theory Comput. 2013. V. 9. № 9. P. 3878.
Klauda J.B., Venable R.M., Freites J.A. et al. // J. Phys. Chem. B. 2010. V. 114. № 23. P. 7830.
Allen M.P., Tildesley D.J. Computer Simulation of Liquids. New York: Oxford University Press, 1991. 385 p.
Steinbach P.J. // Proteins: Structure, Function, and Bioinformatics. 2004. V. 57. № 4. P. 665.
Дополнительные материалы отсутствуют.
Инструменты
Поверхность. Рентгеновские, синхротронные и нейтронные исследования