

Растительные ресурсы, 2019, T. 55, № 2, стр. 231-245
Состояние ценопопуляции Juglans mandshurica (Juglandaceae) при интродукции на территории Волжско-Камского заповедника (Республика Татарстан)
Д. В. Тишин 1, *, М. Б. Фардеева 1, Р. Р. Ризатдинов 1, Н. А. Чижикова 1
1 Казанский (Приволжский) федеральный университет
г. Казань, Россия
* E-mail: dtishin@kpfu.ru
Поступила в редакцию 11.04.2018
После доработки 01.06.2018
Принята к публикации 16.01.2019
Аннотация
Целью работы была оценка популяционной устойчивости Juglans mandshurica Maxim., интродуцированного в Среднем Поволжье. Изучены четыре ценопопуляции J. mandshurica, произрастающие на территории Волжско-Камского заповедника (Республика Татарстан) на участках чистых и смешанных (с Pinus sylvestris и Fraxinus pennsylvanica) культур, заложенных в 1941–1949 гг. Динамика численности, плотности, онтогенетической и пространственной структуры ценопопуляций J. mandshurica изучалась в течение пяти лет (2012–2017 гг.). Был выполнен анализ морфометрических и дендрохронологических характеристик особей разных онтогенетических состояний. Проведенные исследования радиального роста деревьев и стволового сокодвижения J. mandshurica являются пионерными для этого типа местообитаний; они выявили основные климатические факторы, определяющие рост годичных колец и восходящий поток воды в ксилеме. Полученные результаты указывают на то, что орех натурализовался в лесах заповедника, имеет хорошую всхожесть семян и высокую продуктивность генеративных особей. При этом T. cordata является успешным конкурентом ореха маньчжурского, а процессы естественной динамики сообществ снижают развитие его подроста.
Интродукция растений направлена на обогащение флоры новыми видами и может рассматриваться как деятельность, приводящая к увеличению биологического разнообразия природных и антропогенных экосистем [1, 2]. С одной стороны, процесс интродукции рассматривается как практическая деятельность, направленная на акклиматизацию промысловых животных или особо ценных ресурсных видов растений, с другой – это процесс инвазии чужеродных видов в естественные сообщества. Изучение интродуцентов на охраняемых территориях, где задача сохранения биоразнообразия относится к числу приоритетных, является важной и актуальной задачей.
Раифский участок Волжско-Камского государственного природного биосферного заповедника (ВКГПБЗ) уникален тем, что здесь в силу особенностей рельефа сохраняется большое разнообразие сообществ, представляющих основные варианты таежных, смешанных и широколиственных лесов, в миниатюре отражающих всю широтную зональность лесной растительности европейской части России. Сохранности лесных массивов способствовало то, что с 1674 по 1918 г. эти леса принадлежали Раифскому монастырю, который не допускал здесь сплошных рубок, благодаря чему леса сохранили первозданные естественные черты [3]. С 1922 по 1966 г. в Раифе проводились исследования в области лесоводства и лесоведения, включая создание лесных культур и акклиматизации экзотических видов деревьев и кустарников для установления пригодности их при лесоразведении в Среднем Поволжье [4]. После организации заповедника подобные работы прекратились и натурализация и инвазия культур экзотических видов происходят без вмешательства человека.
Для изучения распространения интродуцентов на территории заповедника были выбраны инвазионные и натурализовавшиеся виды растений. К активно распространяющимся инвазионным видам были отнесены Acer negundо L., Padus maackii Rupr., Padus pennsylvanica (L.) Sok., к натурализовавшимся – хозяйственно ценные виды Juglans mandshurica Maxim., Phellodendron amurense Rupr., Fraxinus pennsylvanica Marshall, F. americana L. [5]. В полосе хвойно-широколиственных лесов многие экзоты прекрасно возобновляются; их семена распространяются с помощью ветра и птиц по отвалам дорог, просекам, полянам и нарушенным участкам. Однако некоторые хозяйственно ценные деревья, хотя и акклиматизировались, но с трудом выживают на зарастающих участках лесных культур. К таким видам деревьев относится Juglans mandshurica.
Ареал J. mandshurica охватывает российский Дальний Восток, Северный Китай и Корею, вид произрастает по долинам рек на аллювиальных почвах, образует второй ярус в лесах широколиственно-кедровой формации и участвует в первом ярусе сообществ ясенево-ильмовой формации. По экологическим характеристикам вид является гигромезофитом, отличается светолюбием, ветроопыляемостью и ветроустойчивостью, обладает высокой морозостойкостью, но довольно чувствителен к засухе [6–8].
С 1936 г. в четырех кварталах Раифского леса закладывались культуры J. mandshurica, как чистые, так и в смеси с другими экзотами. К 1973 г. в двух кварталах культуры погибли; из-за отсутствия ухода здесь сформировался березовый лес. В кв. 75 и 76 (посадки 1941–1949 гг.) на дерново-подзолистых почвах, подстилаемых легким суглинком или супесью, в сосняках липовых с елью вид образовал древостои I и II классов бонитета. В современных условиях это единственные сообщества, в которых ценопопуляции ореха маньчжурского возобновляются и пока еще выживают в условиях демутации естественных лесных сообществ.
Основной целью работы было выявление механизмов популяционной устойчивости J. mandshurica в условиях умерено-континентального климата Среднего Поволжья на основе анализа морфофизиологических характеристик.
МАТЕРИАЛ И МЕТОДЫ
Комплексные исследования четырех ценопопуляций (ЦП) J. mandshurica проводились с 2012 по 2017 г. Это участки первоначально чистых культур (кв. 76; ЦП 1, 2) и смешанных культур с Pinus sylvestris (кв. 75; ЦП 3) и Fraxinus pennsylvanica (кв. 75, ЦП 4). В современных условиях ценопопуляции произрастают на дерново-подзолистых супесчанных почвах в разных по составу древесного яруса фитоценозах, что отражено в формулах древостоя: 5Л3Ор2Б (ЦП 1, в понижении, орехово-липовое снытьево-пролесниковое сообщество); 2С4Ор4Л+Б (ЦП 2, липняк с сосной и орехом волосистоосоково-снытьевый); 4С4Л2Ор+Б (ЦП 3, сосняк с липой и орехом костянично-снытьевый); 4Б2Яс2Ор2Л (ЦП 4, наиболее освещенный участок, березняк с ясенем и орехом снытьево-разнотравный). На этих участках были заложены учетные площадки размером 200–540 м2 в зависимости от количества деревьев изучаемого вида.
Комплекс исследований включал геоботанические описания сообществ, картирование всех деревьев, измерение морфометрических, дендрохронологических характеристик и ксилемного потока воды. Первоначально были выделены онтогенетические группы J. mandshurica [9] и проведены морфометрические и дендрохронологические исследования, что позволило в течение пяти лет изучать динамику их численности, плотности, возрастной и пространственной структуры. На основе динамики численности прегенеративных особей определялся прирост численности по каждой прегенеративной группе в процентах. Оценку условий, благоприятствующих возобновлению J. mandshurica проводили на основе выделения особых микроместообитаний, где встречались скопления подроста ореха маньчжурского (табл. 1). Размер таких участков варьировал от 5 до 20 м2. Одни из них находились под пологом или в прогалинах с боковым освещением, другие в более или менее открытых окнах. Для оценки выживаемости подроста в разных местообитаниях учитывались абиотические факторы (освещенность, влажность, температура воздуха), измерения которых проводились летом c 12 по 16 июня 2016 г.
Таблица 1.
Средняя численность прегенеративных особей Juglans mandshurica (экз./10 м2) в разных микроместообитаниях (n = 21) Table 1. The average number of Juglans mandshurica (ind./10 m2) pre-generative individuals in different microhabitats (n = 21)
| Тип микроместообитания Microhabitat | Группы прегенеративных особей Pre-generative groups of individuals (life stages) |
||||
|---|---|---|---|---|---|
| Все All |
p Seedling |
j Juvenile |
im Immature |
v Virginyl |
|
| Под пологом древостоя Under the stand canopy |
6.0 | 0.8 | 2.8 | 1.9 | 0.5 |
| В окне полога Gap |
4.2 | 0.4 | 2.1 | 1.4 | 0.3 |
Для анализа внутрипопуляционного распределения особей в пространстве нами использовалась парно-корреляционая функция pcf(r) (pair correlation function) [10], которая пропорциональна густоте особей на расстоянии r относительно любой выбранной особи. В случае полной пространственной случайности парная корреляционная функция принимает значение, равное единице. Значения парной корреляционной функции, превышающие единицу, свидетельствуют о том, что на расстоянии r встречается больше особей, чем ожидалось бы в случае полной пространственной случайности. В свою очередь значения парной корреляционной функции менее единицы свидетельствуют о разреженности в размещении особей. Статистическая значимость наблюдаемых отклонений pcf(r) оценивается на основе выхода функции за пределы коридора, охватывающего область значений, характерных для случая полной пространственной случайности. Коридор строится с помощью многократных симуляций случайного размещения особей в соответствии с пуассоновской моделью пространственного размещения. Расчеты функций реализованы в пакете spatstat [11] среды статистического анализа R [12].
Для оценки влияния климатических условий на рост особей древесных растений был проведен дендрохронологический анализ кернов древесины, отобранных возрастным буром [13]. Ширина годичных колец измерялась с помощью полуавтоматической установки Lintab (Rinntech, Германия) и программы TsapWin [14]. Точность измерений составила 0.01 мм. По данным измерений ширины годичных колец деревьев J. mandshurica были построены графики абсолютного радиального прироста, которые использовались для точной датировки. Качество датировки оценивалось с помощью программы Сofecha [15]. Для получения обобщенных хронологий, в которых отсутствует возрастной тренд, вычислялись индексы прироста в программе Arstan [16]. Индекс прироста – относительная величина, показывающая отклонение абсолютного прироста от многолетней тенденции. Вычисляется путем деления толщины годичного слоя каждого года на его среднее значение для данного возраста, тем самым убирается возрастной тренд [17]. Для оценки взаимосвязи между климатическими переменными и радиальным приростом древесины рассчитывались коэффициенты корреляции между индексами прироста и среднемесячными значениями температуры и месячных сумм осадков за период 1960–2015 гг. по данным метеостанции “Садовый” (ВКГПБЗ).
Для оценки физиологических различий в водопотреблении ореха маньчжурского и конкурирующих с ним естественных лесообразующих видов – липы мелколиственной и березы повислой были использованы методы определения сокодвижения в течение вегетации в ЦП 2. Измерения восходящего потока воды (пасоки) по сосудам ксилемы проводилось с помощью одноканального измерителя сокодвижения EMS51A (Environmental Management Systems, Чехия) с интервалом в 10 мин в период с мая по август 2016 г. Принцип работы прибора основан на методе теплового баланса ксилемы [18, 19]. Температура, влажность и солнечная радиация измерялись в лесу с помощью мобильной метеостанции Vantage Pro (Davis, США). Связь сокодвижения с внешними факторами определяли по величине коэффициента ранговой корреляции Спирмена. Статистическая обработка данных проводилась в программе PAST ver. 3.0 [20].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
В научной литературе практически отсутствуют данные об онтогенезе ореха маньчжурского. Для характеристики особей прегенеративных онтогенетических групп (проростков, ювенильных, имматурных) использовались 8 морфометрических параметров – высота побега, порядок ветвления, число годичных приростов, количество сложных листьев, длина рахиса без учета черешка, количество листочков сложного листа, длина и ширина листочков (табл. 2).
Таблица 2.
Морфометрические характеристики прегенеративных особей Juglans mandshurica Table 2. Morphometric characteristics of Juglans mandshurica pre-generative individuals
| Признак Attribute |
Онтогенетические группы Ontogenetic groups (life stages) |
|||
|---|---|---|---|---|
| p Проростки Seedling |
j Ювенильные Juvenile |
im-H Имматурные-H Immature-H |
im-T Имматурные-Т Immature-T |
|
| Высота, м Height, m |
0.35 ± 0.01 | 0.4 ± 0.02 | 1.3 ± 0.06 | 0.8 ± 0.04 |
| Кол-во листьев, шт. Number of leaves, pcs. |
3.1 ± 0.1 | 4.7 ± 0.2 | 11.4 ± 0.5 | 7.9 ± 0.4 |
| Длина рахиса, см Length of rachis, cm |
8.6 ± 0.6 | 24.3 ± 1.5 | 57.1 ± 2.9 | 46.7 ± 2.7 |
| Длина листочка, см Length of leaflet, cm |
7.0 ± 0.4 | 12.4 ± 0.6 | 23.7 ± 0.8 | 20 ± 1.1 |
| Ширина листочка, см Width of leaflet, cm |
3.3 ± 0.3 | 5.2 ± 0.1 | 10.3 ± 0.3 | 8.9 ± 0.5 |
| Кол-во листочков в сложном листе, шт. Number of leaflets in a compound leaf, pcs. |
3 ± 0.2 | 4.7 ± 0.2 | 7.9 ± 0.2 | 7 ± 0.3 |
Онтогенез ореха маньчжурского на протяжении прегенеративного периода в климатических условиях заповедника, имеет ряд особенностей. В связи с тем что у имматурных особей, не покрытых зимой снегом, часто обмерзают верхушечные почки, у них более интенсивно развиваются боковые побеги, а рост замедляется. Мы назвали такие особи имматурные-“торчки” (im-Т). Часть имматурных особей в возрасте 7–8 лет достигает высоты 1.5–2 м – это имматурные-“нормальные” (im-Н), у которых образуется довольно прямой стволик. Ветвление (побеги 2–3-го порядка) отмечается у самой верхушки побега, поэтому по почечным кольцам или числу годичных приростов хорошо определяется календарный возраст. Корректность определения возраста по числу годичных приростов на побеге I порядка подтверждена на примере двух особей, у которых параллельно был определен возраст по числу годичных колец на спиле.
В возрасте 7–8 лет нормальные имматурные особи формируют почти зонтиковидную крону за счет сближенных наверху ветвей II порядка с крупными листьями, при этом диаметр ствола еще небольшой, около 2–2.5 см. Длина листьев у имматурных особей в возрасте 4–5 лет составляет в среднем 36.2 см, в сложном листе отмечено 5–9 листочков; в возрасте 7–8 лет в сложном листе насчитывается 9–13 листочков, длина сложного листа максимально достигает 68 см, а в среднем составляет 57.1 см (табл. 2). Практически по всем морфометрическим параметрам между особями разных возрастных состояний отмечаются достоверные различия. По данным М.Т. Сушко [6], в естественных условиях Дальнего Востока в стадию плодоношения орех маньчжурский вступает на 9–12-м году жизни; в естественных лесах Раифы – несколько позже, в 14–16 лет.
По числу годичных приростов побегов было выяснено, что стадия проростка продолжается 1 год, ювенильное состояние – 2 года, имматурное – 4–5 лет, виргинильное – 5–8 лет. Ранее на основе дендрохронологического и морфометрического анализов были охарактеризованы молодые и зрелые генеративные особи (g1, g2) [9]. К сожалению, данные о происхождении семенного материала и о посадке ореха маньчжурского – неполные и, возможно, сеянцы различались по возрасту. Для сравнения мы опирались на данные проверки Татарской лесной станции (которая не существует с 90-х годов прошлого столетия), содержащие сведения о годах посадки и количестве сеянцев, а также об их численности и состоянии на 1973 г. [4].
На основе дендрохронологического анализа были выявлены особи ореха маньчжурского, имеющие календарный возраст 50–65 лет (g2) (посадка 1949 г.), по ряду морфометрических показателей они были отнесены нами к 1-й генерации: высота ствола – 20–21 м, диаметр – 48–54 см, образуется до 300 плодов. В современных условиях преобладают особи 2-й генерации от 30 до 37 лет – молодые генеративные (g1), высота ствола – 12–18 м, диаметр – 22–39 см, образующие до 100–120 плодов. Предположительно формирование зрелых генеративных особей начинается с 40 лет; и это можно определить по увеличению диаметра и высоты ствола, размера кроны и семенной продуктивности. На более или менее открытых участках или вдоль просеки, где снижена конкуренция с липой, формирование зрелых генеративных особей начинается раньше, в связи с чем единично встреченные 37-летние особи высотой 21 м, диаметром ствола 49 см можно отнести к группе зрелых генеративных (g2). Однако по количеству плодов (около 120 на дерево) такие экземпляры уступают зрелым генеративным деревьям I генерации (возрастом 60–65 лет). Также единично отмечались молодые генеративные особи (g1) ореха маньчжурского возрастом 22–26 лет, высотой 12–16 м, диаметр ствола 22–25 см [21].
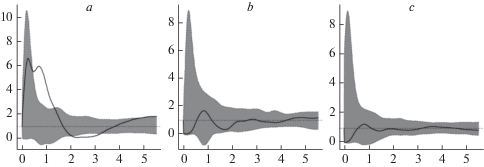
Согласно имеющимся данным, максимальное число особей ореха маньчжурского отмечено в занимающем наибольшую площадь орехово-липовом снытьево-пролесниковом фитоценозе (ЦП 1). Тем не менее средняя плотность особей в ЦП 1 ниже, чем в ЦП 2, произрастающей в липняке с орехом и сосной осоково-снытьевом, хотя первоначально оба этих участка представляли собой чистые культуры ореха маньчжурского. Более низкая плотность ореха маньчжурского отмечена в смешанных культурах: в сосняке с липой (ЦП 3) и в березняке с ясенем пенсильванским (ЦП 4), где зрелые генеративные особи этого вида (g2) отсутствуют (рис. 1а).
Рис. 1.
Характеристика взаимного размещения особей в ЦП J. mandshurica на основе корреляционной функции (PCF). а – размещение прегенеративных особей, b – размещение генеративных особей, c – размещение прегенеративных особей относительно генеративных. По горизонтали – радиус (расстояние от центральной особи), м; по вертикали – значение парной корреляционной функции pcf(r). Fig. 1. Spatial distribution patterns of J. mandshurica described by pair correlation function. a – young individuals (pre-generative life stage), b – adult individuals (generative life stage), c – bivariate pair correlation function of young individuals to adult individuals. X-axis – distance r from any randomly chosen individual of the specified life stage, m; y-axis – pair correlation function pcf(r).
Плотность особей при посадке в 1941 г. в смеси с сосной обыкновенной и ясенем пенсильванским (кв. 75) составляла в среднем 1.9 экз./м2. К 1973 г. особи J. mandshurica имели второй класс бонитета, их плотность снизилась до 1.44 экз./м2. В чистой культуре (1949 г., кв. 76) начальная плотность составляла 0.51 экз./м2; к 1973 г. здесь уже отмечалось 30%-ная примесь подроста из березы и липы, при этом плотность ореха снизилась до 0.36 экз./м2. В динамике за 65-летний период (к 2012 г.) плотность деревьев ореха маньчжурского 1-й генерации снизилась до 0.01–0.014 экз./м2, т.е. в 30 раз по сравнению с 1973 г. В 70-е годы подрост ореха маньчжурского лесники не учитывали, а возможно, и не определяли его.
Онтогенетические спектры в разных эколого-фитоценотических условиях полночленные (рис. 1а), что подтверждает устойчивость популяций. Их поддержанию способствует высокая семенная продуктивность и хорошее прорастание семян. В спектрах преобладают прегенеративные растения, в динамике за пять лет средний прирост численности проростков составляет 73%, ювенильных – 27%. Элиминация J. mandshurica начинается в имматурном и виргинильном состояниях, что обусловлено периодическими морозными зимами, недостатком освещенности, поломкой и, по-видимому, объеданием их зимой лосями. Положительный прирост численности имматурных особей (43.5%) отмечается только в понижении рельефа (ЦП 1), на остальных участках он отрицательный (–24.2%). Прирост численности виргинильных особей везде отрицателен.
В пространственной мозаике проростков и ювенильных особей выделяются скопления, часто приуроченные к подпологовым участкам; скопления имматурных и виргинильных особей отмечаются в основном на более освещенных участках – по прогалинам или в окнах, образованных вывалами старых деревьев ореха маньчжурского. В соответствии с поведением парной корреляционной функции (рис. 1а) тип пространственной структуры прегенеративных особей можно рассматривать как агрегативный: ход кривой указывает на наличие скоплений радиусом от 1.5 до 2.5 м, центры которых удалены друг от друга на расстояние 2.5–4 м. Генеративные особи ореха маньчжурского размещаются в пространстве случайно (рис. 1b). Процесс элиминации, способствующей формированию случайной пространственной структуры, начинается на виргинильной стадии развития. Пространственная мозаика разных возрастных групп обусловлена конкуренцией за свет, которая способствует случайному типу пространственного размещения между собой прегенеративных и генеративных особей (рис. 1c), обеспечивая снижение внутривидовой конкуренции.
В связи с тем, что проростки и ювенильные особи ореха маньчжурского по высоте почти одинаковы (0.3–0.4 м) и находятся в пределах травяного яруса, а виргинильные и имматурные имеют значительно бо́льшую высоту, мы дифференцировали их на две группы и провели раздельно анализ корреляционной связи с факторами среды (табл. 3).
Таблица 3.
Связь численности подроста Juglans mandshurica с абиотическими факторами Table 3. Correlation of Juglans mandshurica undergrowth abundance with abiotic factors
| Фактор Factor |
Микроместообитание Microhabitat |
|||
|---|---|---|---|---|
| Окно в пологе Gap |
Под пологом древостоя Under the stand canopy |
|||
| p + j | im + v | p + j | im + v | |
| Освещенность (макс.) Irradiance (max.) |
0.007 | –0.1 | 0.4* | –0.14 |
| Освещенность (сред.) Irradiance (avg.) |
0.13 | 0.02 | 0.2 | –0.15 |
| Влажность воздуха Air humidity |
0.0 | –0.3* | –0.52* | 0.00 |
| Температура Temperature |
–0.1 | 0.56* | 0.5* | 0.2 |
На Дальнем Востоке для прорастания семян и развития подроста ореха маньчжурского необходимо достаточное освещение и увлажнение [8]. Под пологом липово-ореховых сообществ в Раифе подросту J. mandshurica не хватает освещенности и тепла. Достоверная положительная связь численности с освещенностью и температурой отмечена под пологом для проростков и ювенильных особей. Для имматурных и виргинильных особей температурный фактор является определяющим в “окнах”. Отрицательная корреляция численности, особенно для проростков и ювенильных особей, отмечена с влажностью воздуха под пологом. Однако значения коэффициента корреляции довольно низкие, поэтому четкой зависимости подроста от абиотических факторов, по-видимому, нет. В условиях Раифы более важными факторами являются фитоценотические – разрастание липы в сообществах способствует затенению и соответственно снижает прорастание семян и развитие подроста ореха маньчжурского.
На основе картирования древостоя было выявлено, что коэффициент сопряженности между J. mandshurica и T. cordata высокий, он варьирует от 1.4 до 2.6. Оба вида сходны по своим экологическим оптимумам и часто обитают вместе, что свидетельствует о возможности межвидовой конкуренции. Устойчивости липы способствует ее теневыносливость и морозостойкость, а также стратегия конкурент-толеранта, активно разрастающегося вегетативно [22] в отличие от стресс-толерантного ореха маньчжурского.
Дендрохронологический и физиологический методы исследования для оценки акклиматизации и популяционной устойчивости J. mandshurica применялись в сравнении с оценкой совместно произрастающего с ним вида T. cordata.
В результате дендрохронологического анализа 26 модельных деревьев J. mandshurica можно четко выделить генеративные деревья первого поколения, посаженные в 1941, 1946 и 1949 гг. (кв. 75 и 76), и деревья второй генерации, возникшей после морозной зимы 1978/79 гг., когда температура в Раифе в течение 12 дней держалась около –40 °С. Температурная аномалия привела к гибели 90% особей первой генерации, однако небольшая часть сохранила жизнеспособность, эти особи оказались наиболее устойчивыми и хорошо акклиматизированными. В результате естественного разреживания и осветления участков на вывалах и в окнах появилась путем естественного семенного возобновления вторая генерация J. mandshurica. Особи этой генерации ореха развивались быстрее, чем особи липы, что дало им преимущество в формировании нового сообщества, где орех маньчжурский сейчас формирует 1-й ярус древостоя, а липа в основном, второй.
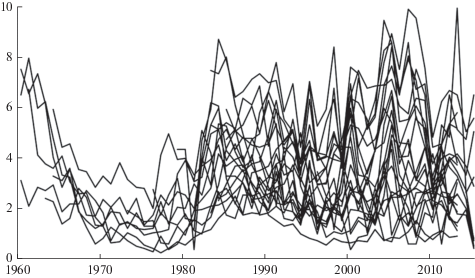
Для каждого модельного дерева была измерена ширина годичных колец и подсчитан календарный возраст. Абсолютные значения радиального прироста ореха маньчжурского разного календарного возраста представлены на графике (рис. 2). Пики приростов древесины у разных деревьев по годам совпадают, что говорит о благоприятности климатических условий этих лет для ореха маньчжурского. Кроме того, по календарному возрасту модельных деревьев можно отметить, что наибольший прирост наблюдался в возрасте от 10 до 20 лет, в пререпродуктивный период орех интенсивно набирает биомассу. Максимальный прирост J. mandshurica отмечен в 1958, 1957 и 2005 гг., минимальный прирост в 1950 и 1951 гг. Периоды подъема радиального прироста наблюдались в 1953–1958, 1981–1984, 2002–2005 гг.; периоды падения: 1958–1961, 1964–1979, 2013–2015 гг.
Рис. 2.
Динамика радиального прироста 26 деревьев J. mandshurica. По горизонтали – календарный год; по вертикали – прирост, мм. Fig. 2. Dynamics of radial increment in 26 J. mandshurica trees. X-axis – year; y-axis – increment, mm.
Периоды подъема и максимальных приростов у J. mandshurica и T. cordata, как правило, не совпадают (рис. 3). У T. cordata максимальный прирост отмечен в 1984, 2000 и 2008 гг., минимальный – в 1973, 1993 и 1995 гг. Периоды подъема радиального прироста наблюдались в 1996–2004, 2005–2007 гг.; периоды падения: 1946–1950, 1990–1995, 2007–2015 гг. То есть увеличение радиальных приростов J. mandshurica отмечается на 5–10 лет раньше, чем у липы. В условиях Раифы увеличение радиальных приростов у липы наблюдается в возрасте 50–60 лет. Как отмечают некоторые авторы, для липы характерны сравнительно медленные темпы роста и развития, сравнительно большая продолжительность жизни [22, 23]. Молодое генеративное состояние в естественных условиях начинается с 30–40 лет, зрелое – с 51–100 лет. Средние значения радиального прироста годичных колец ореха и липы разного возраста представлены в табл. 4.
Рис. 3.
Древесно-кольцевые хронологии J. mandshurica (сплошная линия) и T. cordata (пунктирная линия) в условиях совместного произрастания (76 кв. Раифского участка Волжско-Камского заповедника). По горизонтали – календарный год; по вертикали – индекс прироста, %. Fig. 3. Radial increment indices of J. mandshurica (solid line) and T. cordata (dotted line) in mixed stands of tract No 76 in Raifa part of Volga-Kama nature reserve. X-axis – year; y-axis – radial increment indices, %.
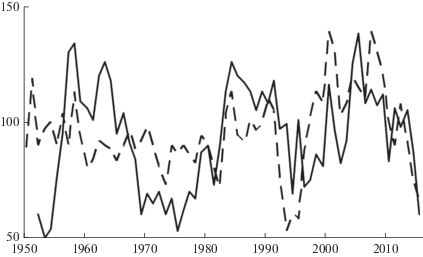
Таблица 4.
Средний возраст и средняя ширина годичного кольца Juglans mandshurica и Tilia cordata Table 4. The average age and tree-ring width of Juglans mandshurica and Tilia cordata
| Вид и число деревьев Species and number of trees |
Зрелые генеративные
особи (g2) Mature adult individuals (g2) |
Молодые генеративные особи (g1) Young adult individuals (g1) |
Особи всех возрастных состояний Individuals of all stages |
|||
|---|---|---|---|---|---|---|
| Ср. возраст Avg. age |
Ср. ширина годичного кольца, мм Average tree-ring width, mm |
Ср. возраст Avg. age |
Ср. ширина годичного кольца, мм Average tree-ring width, mm |
Ср. возраст Avg. age |
Ср. ширина годичного кольца, мм Average tree-ring width, mm |
|
| J. mandshurican = 26 | 54 | 2.5 ± 0.2 | 33 | 4.1 ± 0.2 | 42 | 3.3 ± 0.2 |
| T. cordata n = 12 |
65 | 2.6 ± 0.1 | 49 | 2.4 ± 0.1 | 62 | 2.5 ± 0.1 |
В результате стандартизации абсолютных значений радиального прироста модельных деревьев были получены индексы прироста и построены хронологии для обоих видов (рис. 3). Высокая скорость радиального прироста ореха маньчжурского обусловлена большей, чем у липы мелколистной, площадью листовой поверхности как у виргинильных, так и у генеративных растений. T. cordata уступает ореху и в размерных характеристиках ствола; так, высота ствола у молодых генеративных деревьев (g1) составляет 10–14 м, диаметр ствола 20–22 см; у зрелых деревьев (g2) высота ствола 15–16 м, а диаметр 30–36 см.
Анализ корреляции между индексами прироста и метеофакторами показал (табл. 5), что радиальный прирост ореха маньчжурского положительно связан с температурой января и особенно мая (P < 0.01); была также установлена положительная связь прироста с количеством осадков в июне. Таким образом, радиальный рост J. mandshurica находится под влиянием температуры воздуха зимне-весеннего периода и количества осадков в начале летнего периода.
Таблица 5.
Корреляция индексов прироста J. mandshurica и T. cordata со средней месячной температурой воздуха и месячными суммами осадков Table 5. Correlation of tree growth indices with mean monthly air temperatures and total monthly precipitation
| Месяц Month |
Коэффициент корреляции
с температурой воздуха Mean monthly air temperature correlation coefficient |
Коэффициент корреляции
с суммарными месячными осадками Total monthly precipitation correlation coefficient |
||
|---|---|---|---|---|
| J. mandshurica | T. cordata | J. mandshurica | T. cordata | |
| Январь January |
0.28* | 0.13 | 0.12 | 0.04 |
| Февраль February |
0.24 | 0.01 | 0.10 | 0.00 |
| Март March |
0.03 | 0.17 | 0.10 | 0.08 |
| Апрель April |
–0.09 | 0.14 | 0.14 | –0.17 |
| Май May |
0.32** | 0.12 | –0.23 | 0.23 |
| Июнь June |
0.10 | 0.03 | 0.27* | 0.18 |
| Июль July |
0.05 | –0.25* | 0.17 | 0.26* |
| Август August |
0.10 | 0.23 | 0.20 | 0.00 |
Напротив, для морозостойкой T. сordata более важными оказываются характеристики летнего периода: обнаружена значимая связь радиального прироста с температурой и осадками июля (P < 0.05).
Измерение восходящего потока воды по сосудам ксилемы у ореха маньчжурского показало, что в суточном ходе сокодвижения наблюдается один пик. Максимальные значения приходятся на период с 11.00 до 14.00 ч. Максимальные суточные значения потока воды наблюдались в конце июля и в начале августа (до 0.23 кг ⋅ час–1 ⋅ см–1). Минимальные значения – весной и осенью (0.02 кг ⋅ час–1 ⋅ см–1), а также в дни с пасмурной и дождливой погодой. Наблюдается высокая корреляция сокодвижения с солнечной радиацией (R = 0.8) и температурой воздуха (R = 0.9). С влажностью воздуха наблюдается отрицательная связь (R = –0.7). Наибольшие значения сокодвижения наблюдались, когда солнечная радиация достигала 700 W ⋅ m–2 в июне и августе, при температуре воздуха выше +26 °С и влажности 40% и ниже.
Cравнение мощности сокодвижения светолюбивых видов (ореха маньчжурского и березы повислой) с мощностью сокодвижения теневыносливой липы мелколистной, произрастающих совместно, показало высокую синхронность (рис. 4). Однако у T. сordata наблюдаются более низкие значения восходящих потоков воды по сравнению с J. mandshurica. Например, в жаркие дни c 29 июля по 2 августа 2016 г., в 12–13 ч дня у липы сокодвижение не превышало значений 0.11 кг ⋅ час–1 ⋅ см–1, а у ореха оно достигало значений 0.22 кг ⋅ час–1 ⋅ см–1. Активность сокодвижения у Betula pendula занимает промежуточное положение в сравнении с орехом и липой. Такие различия в сокодвижении у лиственных деревьев связаны не только с различными экологическими требованиями к фактору освещенности, но и с разной площадью листовой поверхности и разной высотой стволов. Являясь самой светолюбивой породой в Восточно-европейских лесах [24], занимая первый ярус на исследуемых участках и поднимаясь чуть выше ореха маньчжурского (до 24 м), береза уступает ему существенно по общей площади листовой поверхности и мощности сокодвижения. Эти параметры у липы также всегда меньше, чем у ореха. Таким образом, в жаркие засушливые дни в результате сильной транспирации ореху маньчжурскому требуется больше воды, чем аборигенным видам, в частности, липе мелколистной и берёзе повислой.
Рис. 4.
Стволовое сокодвижение у лиственных деревьев в период с 29 июля по 02 августа 2016 г. 1 – J. mandshurica, 2 – B. pendula, 3 – T. cordata. По горизонтали – десятиминутки; по вертикали – мощность сокодвижения, кг ⋅ час–1 ⋅ см–1. Fig. 4. Stem sap flow in deciduous tree species from July 29 to August 2, 2016. 1 – J. mandshurica, 2 – B. pendula, 3 – T. cordata. X-axis – 10-minute intervals; y-axis – sаp flow rate, kg ⋅ h–1 ⋅ cm–1.

ЗАКЛЮЧЕНИЕ
Орех маньчжурский Juglans mandshurica Maxim., произрастающий в естественных условиях на богатых почвах в составе долинных широколиственных и кедрово-широколиственных лесов Дальнего Востока имеет предпосылки для достаточно успешного развития в чистых и смешанных культурах при интродукции в условиях Среднего Поволжья (Волжско-Камский государственный природный биосферный заповедник). Вегетационный период ореха маньчжурского не превышает вегетационный период аборигенных видов. Высокая семенная продуктивность и всхожесть семян, выживаемость проростков и ювенильных растений обеспечивает полночленность и устойчивость его ценопопуляций. Для пространственной мозаики ореха маньчжурского характерно групповое распределение прегенеративных особей и случайное распределение генеративных, а также групп прегенеративных и генеративных особей относительно друг друга, что обеспечивает эффективное использование среды и снижает внутривидовую конкуренцию. Большое значение для поддержания вида в условиях более континентального климата имеет значительная масса листвы, обеспечивающая ореху маньчжурскому высокие приросты древесины и хорошее сокодвижение. Вместе с тем более континентальный климат Среднего Поволжья с экстремально низкими температурами зимой и засухой в летний период порой приводит к снижению жизненности отдельных особей ореха в связи с образованием морозобойных трещин на их стволах.
Ведущим лимитирующим фактором развития ореха маньчжурского в естественных фитоценозах является конкуренция с липой мелколистной. Липа мелколистная, как аборигенный вид, обладает большей приспособленностью к природным условиям зоны хвойно-широколиственных лесов европейской части России, чем орех маньчжурский. В исследованных сообществах липа мелколистная, как правило, представлена различными жизненными формами (одноствольное дерево, порослеобразующее дерево, куртинообразующее дерево), позволяющими виду интенсивно внедряться под полог культур ореха маньчжурского и постепенно вытеснять его. Это доказывается высокой численностью и близкой к полночленной возрастной структурой ценопопуляций липы, ее хорошим возобновлением. При надлежащем уходе за культурой ореха маньчжурского его насаждения могут развиваться успешно.
Вопрос о накоплении в биомассе ореха маньчжурского биологически активных веществ при выращивании в условиях интродукции под воздействием стрессовых факторов среды требует специального изучения. Проведенный анализ итогов акклиматизации вида открывает перспективы введения этой древесной породы в культуру.
Список литературы
Лапин П.И. 1972. О терминах, применяемых в исследованиях по интродукции и акклиматизации растений. – Бюл. ГБС. 83: 10–18.
Штильмарк Ф.Р. 1996. Историография российских заповедников (1895–1995). М. 340 с.
Бакин О.В. 2001. Памятник науки. Раифа-Свияжск. Казань. С. 66–76.
Дерюга Е.С., Мурзов А.М. 1977. Состояние культур экзотов и естественное расселение их в Раифском лесном массиве. Тр. Волж.-Камс. гос. зап-ка. Вып. III. Казань. С. 61–79.
Иванов В.Б. 2005. Аннотированный список растений коллекции дендрария Волжско-Камского государственного природного заповедника. Тр. Волж.-Камс.гос. зап-ка. Вып.VI. Казань. С. 343–375.
Сушко М.Т. 1973. Маньчжурский орех и его ресурсы на Дальнем Востоке. – Раст. ресурсы. 9(2): 271–279.
Авеличева С.Н. 2005. Маньчжурский орех как источник сырья для повышения качества непродовольственных товаров. Владивосток. 100 с.
Рейф О.Ю. 2013. Эколого-географические условия местопроизрастания ореха маньчжурского на юге Дальнего Востока. – В сб.: Состояние лесов и актуальные проблемы лесоуправления: Тез. докл. Всероссийской конференции с международным участием. Хабаровск. С. 402–405.
Фардеева М.Б., Исламова Г.Р., Чижикова Н.А. 2014. Пространственно-онтогенетическая структура ценопопуляций Vaccinium myrtillus (Ericaceae) близ южной границы ареала (Республика Татарстан). – Раст. ресурсы. 50(3): 376–397.
Stoyan D., Stoyan H. 1994. Fractals, random shapes and point fields: methods of geometrical statistics. Chichester; New York. 389 p.
Baddeley A., Rubak E., Turner R. 2015. Spatial Point Patterns: Methodology and Applications with R. London. 408 p. https://doi.org/https://doi.org/10.1201/b19708
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2017. https://www.R-project.org/.
Методы дендрохронологии. 2000. Учебно-методическое пособие. Ч. I. Основы дендрохронологии. Сбор и получение древесно-кольцевой информации. Красноярск. 80 с.
Rinn F. 2011. TSAP-Win – Time Series Analysis and Presentation for Dendrochronology and Related Applications, ver. 4.6. Heidelberg. 92 p.
Holmes R.L. 1983. Computer assisted quality control in tree-ring dating and measurement. – Tree-Ring Bulletin. 43: 69–78. https://repository.arizona.edu/handle/10150/261223
Holmes R.L. 1994. Dendrochronological Program Library users manual. Tucson. 51 p. https://www.ltrr.arizona.edu/pub/dpl-mac/68k/dpl.txt
Битвинскас Т.Т. 1974. Дендроклиматические исследования. Л. 172 с.
Kucera J., Cermak J., Penka M. 1977. Improved thermal method of continual recording the transpiration flow rate dynamics. – Biol. Plant. 19(6): 413–420. https://doi.org/https://doi.org/10.1007/BF02922976
Cermak J., Kucera J., Nadezhdina N. 2004 Sap flow measurements with some thermodynamic methods, flow integration within trees and scaling up from sample trees to entire forest stands. – Trees. 18(5): 529–546. https://doi.org/https://doi.org/10.1007/s00468-004-0339-6
Hammer Ø., Harper D.A.T., Ryan P.D. 2001. PAST: Paleontological statistics software package for education and data analysis. – Palaeontologia Electronica. 4(1): 9p. http://folk.uio.no/ohammer/past
Tishin D., Fardeeva M., Chizhikova N., Rizatdinov R. 2018. Acclimation of Juglans mandshurica Maxim. and Phellodendron amurense Rupr. in the Middle Volga region. – IOP Conference Series: Earth and Environmental Science. 107(1): 1–6. https://doi.org/10.1088/1755-1315/107/1/012094
Евстигнеев О.И. 2004. Популяционные стратегии видов деревьев. – В кн.: Восточно-европейские леса (История в голоцене и современность). М. С. 176–204.
Чистякова А.А. 1987. Поливариантность онтогенеза и типы поведения деревьев широколиственных лесов. – В кн.: Популяционная экология растений. М. С. 39–43.
Восточно-европейские леса: история в голоцене и современность. 2004. М. 478 с. http://rjee.ru/wp-content/uploads/2016/03/Kniga_1.pdf.
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы