

Растительные ресурсы, 2019, T. 55, № 3, стр. 362-376
Реакция Pinus sylvestris засухоустойчивого сорта ‘Острогожская’ на гидротермический стресс в условиях ex situ и ex vivo
Е. С. Клушевская 1, *, Н. Ф. Кузнецова 1
1 Всероссийский научно-исследовательский институт лесной генетики,
селекции и биотехнологии
г. Воронеж, Россия
* E-mail: ekogenlab@gmail.com
Поступила в редакцию 10.03.2018
После доработки 10.12.2018
Принята к публикации 17.04.2019
Аннотация
На основе ряда физиолого-биохимических параметров хвои проведено сравнительное изучение реакции деревьев сосны обыкновенной (Pinus sylvestris L.) засухоустойчивого сорта ‘Острогожская’ и деревьев сосны обыкновенной в культурах, типичных по засухоустойчивости и продуктивности для Центрально-Черноземного района России (контроль), на гидротермический стресс. Изучена норма реакции показателей водного режима (общее количество влаги, количество коллоидно-связанной воды, дефицит влаги) и содержания аминокислоты пролин в свежесобранной хвое модельных деревьев и ее изменение при имитации сильного и умеренного стресса в лабораторных условиях. Показано, что при сильном стрессе адаптивные возможности растений контрольного и опытного объектов одинаковы. Действие умеренного стресса расширяет границы нормы реакции растений контрольного насаждения. Деревья сосны обыкновенной сорта ‘Острогожская’ реагируют на стресс однотипно, стресс-реакция не зависит от силы и продолжительности воздействия. Обсуждаются механизмы устойчивости сорта сосна ‘Острогожская’ к засухе.
В России засухам и процессам опустынивания подвержена значительная часть территории. Начавшиеся в XIX в. в лесостепной зоне процессы остепнения в последние десятилетия ускоряются в связи с глобальными изменениями климата. Наиболее уязвимыми в данной ситуации оказываются лесные древесные растения [1]. Аридизация климата ведет к изменению и перераспределению водного баланса, снижению продуктивности и устойчивости лесов, обеднению их видового и генетического разнообразия [2–5].
Сосна обыкновенная Pinus sylvestris L. одна из основных лесообразующих пород, обладает высокой экологической пластичностью и стресс-толерантностью, что позволяет ей выживать в широком диапазоне условий температуры, влажности, аэрации, минерального состава почв [6]. В последнее столетие антропогенное и климатогенное воздействие на сосновые леса было настолько велико, что негативно сказалось на состоянии общего генофонда вида, его межвидовой конкурентоспособности при естественном возобновлении. На южной границе и в центре зоны экологического оптимума вида в Европейской части России появились первые признаки локальной популяционной неустойчивости, произошла дестабилизация генетического материала. Снижение устойчивости и конкурентоспособности сосны ведет к потере генетических ресурсов и сужению ареала. Процессы дестабилизации на юге бореальной зоны обусловлены резким падением уровня грунтовых вод (в 2–4 раза) и произошедшими здесь сильными засухами 2007, 2010, 2012, 2014 гг., что определяет актуальность физиолого-биохимических исследований засухоустойчивости.
Установлено, что адаптивным морфологическим изменениям предшествуют биохимические и физиологические перестройки, вызванные неблагоприятными биотическими и абиотическими факторами среды [7]. Внутриклеточные механизмы адаптации, определяющие устойчивость растений к гидротермическому стрессу, коррелируют с генетической природой засухоустойчивости, которая обусловлена как наследственными признаками, так и вырабатывается в процессе онтогенеза [8].
Норма реакции является универсальным понятием, применяемым ко всем экологически зависимым признакам для характеристики генотипов, популяций и экосистем в целом. Она характеризует долю участия среды в их реализации, и имеет свои генетически обусловленные пределы. Норма реакции относится к важнейшим показателям оценки текущего состояния генотипа и популяции, выступает как фактор отбора при формировании лесных экосистем по принципу взаимодействия “генотип–среда”, обеспечивает структурно-функциональную устойчивость лесных древесных растений к флуктуациям регионального климата [9–11]. Поэтому крайне важен вопрос изучения широты нормы реакции у древесных растений в оптимальные по погодным условиям годы и ее трансформации при воздействии гидротермического стресса.
Цель данной работы – сравнительное изучение физиолого-биохимических параметров хвои (содержание влаги и коллоидно-связанной воды, водный дефицит, содержание пролина) деревьев сосны обыкновенной в разных по засухоустойчивости лесных культурах в оптимальных и стрессовых гидротермических условиях.
МАТЕРИАЛЫ И МЕТОДЫ
Объект исследования – 38-летние лесные культуры сосны обыкновенной (Pinus sylvestris L.) сорта ‘Острогожская’, II класса бонитета, произрастающие на юге лесостепной зоны (Воронежская область, Острогожский район) на слабо- и среднеподзоленных влажных песчаных почвах. Сосна сорта ‘Острогожская’ характеризуется повышенной засухоустойчивостью и семенной продуктивностью (патент на селекционное достижение № 9187). Насаждение было отобрано Н.Ф. Кузнецовой и О.С. Машкиной в засуху 2007 г., и его селекционная ценность подтверждена в ходе сортоиспытания на фоне оптимальных и трех засушливых лет [12]. Сформировавшийся рядом с насаждением 2–5-летний подрост естественного возобновления нормально перенес сильнейшую за всю историю метеонаблюдений засуху 2010 г. После вступления в фазу семеношения проведено испытание сорта по потомству и установлена достаточно высокая его репродуктивная воспроизводимость по семенной продуктивности и засухоустойчивости.
Контролем служили 50–55-летние лесные культуры сосны обыкновенной II класса бонитета Ступинского тест-объекта (Воронежская обл., Рамонской р-н), произрастающие на слабо- и среднеподзоленных влажных песчаных почвах. Насаждение с восточной стороны прилегает к Воронежскому биосферному заповеднику. Все деревья по фенотипу и состоянию относятся к категории региональной нормы: по высоте лучшие, хорошие и средние. Ступинский тест-объект – это объект длительного мониторинга систем семенного размножения в центральной части лесостепной зоны (1987–2018 гг.), типичное для ЦЧР насаждение по вегетативной и семенной продуктивности, относится к числу эталонных территорий с ненарушенным почвенным покровом [13]. Оба насаждения характеризуются сходными, отвечающими биологии вида, лесорастительными условиями (ТУМ А2, сосняк травяной). Физиолого-биохимические исследования проводятся с 2013 года.
Физиолого-биохимические показатели хвои (дефицит влаги, общее количество влаги, количество коллоидно-связанной воды, содержание свободного пролина) определялись в трех повторностях для каждого из 10 модельных деревьев в насаждениях сосны обыкновенной сорта ‘Острогожская’ (средняя высота 15.9 м, диаметр 25.1 см) и 10 модельных деревьев сосны обыкновенной в культурах Ступинского тест-объекта (средняя высота 17.4 м, диаметр 32.8 см) с последующим усреднением данных. В мае с модельных деревьев были отобраны побеги второго года жизни и проведен анализ физиолого-биохимических параметров хвои в трех вариантах опыта: 1) свежесобранные пробы; 2) воздействие сильного гидротермического стресса “засуха + температура” (7 сут при 45 °С); и 3) воздействие умеренного стресса “засуха” (10 сут при комнатной температуре). Для имитации засухи использовали 0.6 М раствор маннита. Концентрация маннита, температурный режим и время экспозиции устанавливались на основании кривой отклика, построенной по предварительным экспериментам [14]. Исследования водного режима растений проводили по общепринятой методике Х.Н. Починка [15]. Содержание свободного пролина определяли в воздушно-сухих пробах методом Бейтса [16]. Статистическую обработку проводили с помощью программы Statistica 6. Для сравнения выборок и оценки влияния исследуемых климатических факторов использовали дисперсионный анализ (ANOVA).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Как известно, основными факторами, влияющими на рост и развитие растений, являются температура, влажность, освещение, минеральный состав почв. Все анализируемые физиолого-биохимические показатели (общее количество влаги, количество коллоидно-связанной воды, дефицит влаги, содержание пролина) являются экологически зависимыми и их изменение тесно связано с колебаниями условий внешней среды. Исследование влияния индуцированных стрессовых факторов (сильная и умеренная засуха) позволило в лабораторных условиях выявлять особенности стресс-реакции хвои деревьев разных по засухоустойчивости насаждений сосны обыкновенной.
Общее количество влаги. Сравнение общего количества влаги в хвое деревьев сосны в двух изученных насаждениях выявило достоверные различия (F = 32.07; p < 0.05) по данному показателю. Для деревьев сосны сорта ‘Острогожская’ средние значения признака составили 48.9%, для деревьев Ступинского насаждения – 55.3%, (рис. 1а), что в целом соответствует литературным данным [17]. Возможно, одной из причин различий является более южное (на 150 км) расположение насаждения сосны сорта ‘Острогожская’. Но очевидно существуют и другие причины, так как растения по-разному реагируют на индуцированный стресс.
Рис. 1.
Содержание влаги (a–c) и его амплитуда (d) в хвое деревьев культур сосны обыкновенной: засухоустойчивого сорта сосна ‘Острогожская’ и Ступинского тест-объекта (контроль) в нативном состоянии (a), при сильном (b) и умеренном (c) гидротермическом стрессе. По вертикали: a, b, c – содержание влаги, %; d – амплитуда изменения признака, %. По горизонтали: d – 1 – нативное состояние, 2 – сильный гидротермический стресс, 3 – умеренный гидротермический стресс. Светлые столбики – культуры сосны Ступинского тест-объекта; темные столбики – культуры сосны сорта ‘Острогожская’. Fig. 1. Moisture content (a–c) and its range (d) in Scots pine needles of: drought-tolerant variety ‘Ostrogozhskaya’ and trees of Stupinsky cultivated forest (control) in the native state (a), under severe (b) and moderate (c) hydrothermal stress. X-axis: a, b, c – moisture content, %; d – range of moisture content change, %. Y-axis: d – 1 – native state, 2 – severe hydrothermal stress, 3 – moderate hydrothermal stress. Light columns – Scots pine of Stupinsky cultivated forest (control); dark columns – plantations of drought-tolerant Scots pine ‘Ostrogozhskaya’.
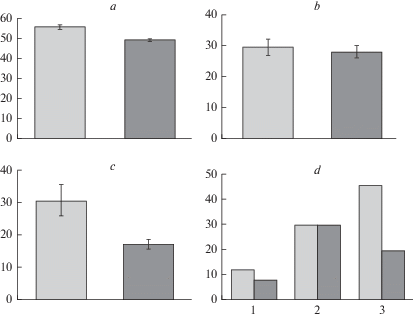
По данным настоящего и предшествующих исследований деревья сосны Ступинского тест-объекта в силу индивидуальных особенностей очень разнородны по содержанию влаги [18]. Диапазон изменчивости признака в нативном состоянии достигает 11.6% (50.7–62.3%) что в полтора раза больше, чем у сосны сорта ‘Острогожская’ – 7.7% (43.8–51.5%). По-видимому, это обусловлено разной ответной реакцией деревьев на гидротермический стресс во время последней тепловой волны, которая в районе исследований продолжалась в течение 9 лет (2006–2014 гг.) и включала 4 сильные засухи (2007, 2010, 2012 и 2014 гг.). Это привело к резкому понижению уровня грунтовых вод (с 3 до 9.8 м в 2010 г.). Значительно более узкая норма реакции сосны сорта ‘Острогожская’ может свидетельствовать о наличии у нее механизмов засухоустойчивости.
При индуцированном стрессе между контрольным и опытным насаждениями наблюдаются еще более существенные различия (рис. 1b, c). У деревьев Ступинского тест-объекта уровень влаги в хвое понижается почти вдвое (F = 81.1; p < 0.05, F = 23.3; p < 0.05), но не зависит от силы и времени действия стрессового фактора (F = 0.04; p > 0.05). Отклик сосны сорта ‘Острогожская’, напротив, зависит от вида стресса (F = 16.2; p < 0.05). При сильном гидротермическом стрессе (рис. 1b), уровень влаги в хвое понижается в 1.8 раза (F = 91.3; p < 0.05), при действии умеренного стресса (рис. 1c) – в 2.8 раза (F = 335.2; p < 0.05).
Сильный стресс уравнивает показатели содержания влаги (рис. 1b) и границы колебания значений признака (рис. 1d-2) в двух насаждениях (F = 0.21, р > 0.05); значения совпадают приблизительно на 80%. По-видимому, это указывает на сходные механизмы выживания растений в экстремальных условиях.
При умеренном стрессе, когда появляется возможность адаптации, у контрольных деревьев амплитуда изменчивости признака (рис. 1d-3) повышается примерно в 4 раза (45.6%), а у опытных только в 2.5 раза (19.4%). При этом, если минимальные значения содержания влаги в хвое сосны в контрольном и опытном насаждениях сопоставимы (соответственно 9.8 и 10.8%), то максимальные значения только у отдельных растений контрольного объекта остаются в пределах диапазона изменчивости признака в нативных образцах хвои – 55.4%, что значительно увеличивает амплитуду изменения признака. У деревьев опытного насаждения области изменчивости признака в нативном состоянии и при индуцированном стрессе не перекрываются (максимум при стрессе составляет 30.2%). Более узкий по сравнению с контролем диапазон изменчивости признака (рис. 1), а так же более выровненный отклик деревьев сосны сорта ‘Острогожская’ на стресс, могут свидетельствовать о лучшей приспособленности растений к данной гидротермической нагрузке.
Количество коллоидно-связанной воды. Известно, что для сохранения постоянства внутренней среды требуется определенный уровень влажности. П.А. Генкель [8] указывал на значимость определенных минимальных значений содержания влаги (гомеостатической воды), присущих каждому генотипу растений, и обязательных для сохранения его жизнеспособности. Существуют внутриклеточные механизмы, направленные на поддержание необходимого и достаточного уровня оводнения, за счет увеличения количества связанной воды, которое зависит от растворенных в клеточном соке веществ. Вода, связанная с биополимерами клетки, и в первую очередь с белками, является основным компонентом цитоплазмы, средой протекания внутриклеточных процессов. Благодаря наличию гидратной оболочки, белки, с одной стороны, сохраняют свою архитектонику, необходимую для нормального функционирования биомолекул, с другой – поддерживают определенный уровень влагосодержания клетки [19]. В фазе адаптации к неблагоприятным условиям усиливается синтез стрессовых белков, таких как осмотины [20], дегидрины [21, 22] и др., способствующих удержанию воды. Существует мнение, что даже в пределах вида, устойчивость растений к негативным факторам во многом определяется их генетически детерминированной способностью к синтезу определенных видов стрессовых белков [23].
В свежесобранных пробах хвои деревьев Ступинского насаждения и насаждения сосны сорта ‘Острогожская’ средний уровень коллоидно-связанной воды (рис. 2а) идентичен и относительно невысок: соответственно 19.1% (10.8–27.7%) и 22.2% (15.1–32.8%) (рис. 2а). Диапазоны изменчивости полученных значений совпадают на 57% (F = 2.1; p > 0.05). Как и содержание влаги, количество коллоидно-связанной воды при сильном стрессе (рис. 2b) соизмеримо в образцах хвои обоих насаждений (значения перекрываются почти на 84% (F = 0.02; p > 0.05)). Однако, если в контрольном варианте происходит повышение максимальных значений в 1.5 раза (F = 11.2; p < 0.05), что почти на 80% больше, чем в состоянии ex situ, то в опытном – только в 1.2 раза (F = 5.0; p < 0.05), т.е. на 42%.
Рис. 2.
Содержание коллоидно-связанной воды (а–c) и его амплитуда (d) в хвое деревьев культур сосны обыкновенной: засухоустойчивого сорта сосна ‘Острогожская’ и Ступинского тест-объекта (контроль) в нативном состоянии (а), при сильном (b) и умеренном (c) гидротермическом стрессе. По вертикали: a, b, c – содержание коллоидно-связанной воды, %; d – амплитуда изменения признака, %. По горизонтали: d – 1 – нативное состояние, 2 – сильный гидротермический стресс, 3 – умеренный гидротермический стресс. Светлые столбики – культуры сосны Ступинского тест-объекта; темные столбики – культуры сосны сорта ‘Острогожская’. Fig. 2. Amount of colloid bound water (a–c) and its range (d) in Scots pine needles of: drought-tolerant variety ‘Ostrogozhskaya’ and trees of Stupinsky cultivated forest (control) in the native state (a), under severe (b) and moderate (c) hydrothermal stress. X-axis: a, b, c – amount of colloid bound water, %; d – range of amount of colloid bound water change, %. Y-axis: d – 1 – native state, 2 – severe hydrothermal stress, 3 – moderate hydrothermal stress. Light columns – Scots pine of Stupinsky cultivated forest (control); dark columns – plantations of drought-tolerant Scots pine ‘Ostrogozhskaya’.
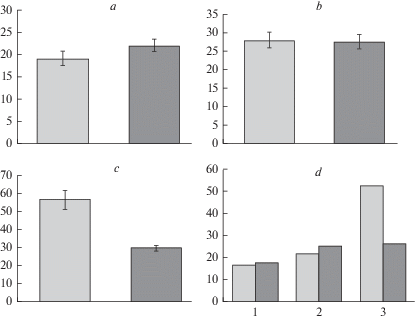
При действии умеренного стресса количество коллоидно-связанной воды (рис. 2c) в хвое деревьев сосны Ступинского насаждения увеличивается почти втрое относительно свежесобранных проб (F = 50.8; p < 0.05). Признак варьирует в очень широких пределах (36.7–89.5%), амплитуда составляет около 53% (рис. 2d-3), при этом минимальные показатели выше значений, полученных в условиях ex situ почти на 9%. Данные свидетельствуют, что деревья Ступинского насаждения по-разному реагируют на действие сильного и умеренного индуцированного стресса (F = 28.2; p < 0.05). Несмотря на то, что путь сохранения гомеостаза внутренней среды один и тот же – повышение содержания коллоидно-связанной воды, механизмы его реализации, по-видимому, различные. Между контрольным и опытным насаждениями по отклику деревьев на разную стрессовую нагрузку также выявлены существенные различия. В хвое деревьев сосны Ступинского насаждения содержание коллоидно-связанной воды при умеренном стрессе в 1.9 раз больше (F = 35.24; p < 0.05), чем в хвое деревьев сосны сорта ‘Острогожская’ (рис. 2c), области распределения признака перекрывается лишь на 11.4%.
Для сосны сорта ‘Острогожская’ продолжительность и сила индуцированного стресса не имеют существенного значения (F = 0.8; p > 0.05). Количество коллоидно-связанной воды при умеренном стрессе (рис. 2c) увеличивается в 1.3 раза (F = 13.3; p < 0.05). Область распределения значений признака (18.0–44.8%) на 50.0% перекрывается с областью значений в свежесобранных пробах (15.1–32.8%) и на 87.4% с областью значений при сильном гидротермическом стрессе (16.9–42.4%). При этом амплитуда изменения признака при сильном и умеренном стрессе приблизительно одинаковая (рис 2d-2, 3). Возможно, отсутствие у сосны сорта ‘Острогожская’ принципиальных различий по вариантам опыта подтверждает повышенную ее устойчивость к засухе, наличие у деревьев внутриклеточных приспособительных механизмов, обеспечивающих гомеостаз внутренней среды в ответ на гидротермическую нагрузку в засушливые годы в условиях ex situ и при лабораторной ее имитации ex vivo.
Дефицит влаги. Данный признак является одной из важнейших характеристик водного режима, позволяющей судить как о силе стрессового воздействия, так и о текущем жизненном состоянии растений. Показано, что даже незначительные колебания признака отражаются на протекании физиолого-биохимических процессов, а долговременный водный дефицит может приводить к анатомическим и морфологическим изменениям [24, 25]. В Архангельской области уровень дефицита влаги у сосны обыкновенной составляет 12–14% [17]. В Воронежской области его уровень в обоих насаждениях более чем на треть меньше: 87% значений этого показателя у опытных деревьев и 80% – у контрольных ниже представленных в литературе. Вероятно, это обусловлено зональной спецификой района исследований, сформированной в чреде поколений адаптацией сосны к произрастанию на влагодефицитной территории лесостепной зоны.
По величине дефицита влаги у сосны сорта ‘Острогожская’ (в среднем 9.4%) и у деревьев Ступинского тест-объекта (8.7%) достоверных различий не обнаружено (F = 0.4; p > 0.05) (рис. 3а). Область распределения значений признака у деревьев опытного насаждения полностью включает в себя область значений признака у деревьев контрольного насаждения. Минимальные показатели дефицита влаги (5–6%) в двух насаждениях также сопоставимы. На 30% более высокие максимальные значения у сосны сорта ‘Острогожская’, вероятно, вызваны расположением опытного объекта на южной границе лесостепной зоны, а контрольного в центральной ее части.
Рис. 3.
Дефицит влаги (а–c) и его амплитуда (d) в хвое деревьев культур сосны обыкновенной: засухоустойчивого сорта сосна ‘Острогожская’ и Ступинского тест-объекта (контроль) в нативном состоянии (а), при сильном (b) и умеренном (c) гидротермическом стрессе. По вертикали: а, b, c – дефицит влаги, %; d – амплитуда изменения признака, %. По горизонтали: d – 1 – нативное состояние, 2 – сильный гидротермический стресс, 3 – умеренный гидротермический стресс. Светлые столбики – культуры сосны Ступинского тест-объекта; темные столбики – культуры сосны сорта ‘Острогожская’. Fig. 3. Water deficit (a–c) and its range (d) in Scots pine needles of: drought-tolerant variety ‘Ostrogozhskaya’ and trees of Stupinsky cultivated forest (control) in the native state (a), under severe (b) and moderate (c) hydrothermal stress. X-axis: а, b, c – water deficit, %; d – range of water deficit change, %. Y-axis: d – 1 – native state, 2 – severe hydrothermal stress, 3 – moderate hydrothermal stress. Light columns – Scots pine of Stupinsky cultivated forest (control); dark columns – plantations of drought-tolerant Scots pine ‘Ostrogozhskaya’.
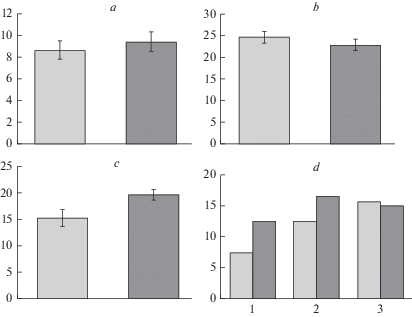
При имитации сильной засухи в лабораторных условиях более интенсивный отклик показателя дефицита влаги в хвое зафиксирован у деревьев Ступинского насаждения. Его средняя величина повысилась в 2.8 раза (F = 106.8; p < 0.05) (рис. 3b), диапазон изменчивости признака увеличился на 41.2% (рис. 3d-1, 2), показатели вышли за пределы нормы реакции. Минимальное значение дефицита влаги составило 18%, что в 1.4 раза выше его максимальных показателей ex situ.
У сосны сорта ‘Острогожская’ в этом варианте опыта дефицит влаги в среднем повысился в 2.4 раза (F = 68.0; p < 0.05), за пределы нормы реакции вышло примерно 87% значений. Область распределения значений признака шире, чем у сосны Ступинского насаждения на 25% и полностью включает в себя контрольные значения (рис. 3b, d-2). Cредние значения и амплитуды дефицита влаги контрольного и опытного насаждений достоверно не различаются (F = 0.8; p > 0.05).
У деревьев Ступинского тест-объекта обнаружены достоверные различия (F = 20.2; p < 0.05) величины дефицита влаги в двух вариантах опыта ex vivo. При умеренном стрессе дефицит влаги увеличился в 1.8 раза (F = 13.8; p < 0.05), (рис. 3c), при сильном стрессе – в 2.8 раза (F = 106.8; p < 0.05) (рис. 3b). Амплитуда значений признака при умеренном стрессе увеличилась соответственно в 2.2 и 1.3 раза относительно величин, установленных в свежесобранных пробах и при сильном стрессе (рис. 3d-1–3). У деревьев сосны сорта ‘Острогожская’ также происходит рост дефицита влаги, одинаковый (F = 3.7; p > 0.05), как при умеренном стрессе – в 2 раза (F = 58.0; p < 0.05), так и при сильном – в 2.4 раза (F = 68.0; p < 0.05). Область значений признака в двух вариантах опыта совпадает.
Содержание пролина. Поддержание уровня метаболизма максимально близкого к гомеостатическому при стрессовой нагрузке (насколько это возможно в условиях лабораторного эксперимента) достигается во многом за счет повышения осмотически и коллоидно активных веществ, в том числе и аминокислот (пролин, аланин и др.) [26]. Аминокислота пролин, образующаяся при негативном воздействии на растение в значительных количествах, связывается с белками, защищая их от денатурации и, таким образом, сохраняя их функциональную активность [27, 28]. Содержание этого стрессового лабильного метаболита сильно возрастает в засуху. Доказана его многофункциональная роль в поддержании клеточного гомеостаза при адаптации растений к стрессу [29].
При изучении концентрации свободного пролина по вариантам опыта оказалось, что она изменяется аналогично показателю водного режима растений, который направлен на поддержание гомеостатического состояния – коллоидно-связанной воды. Рядом исследователей отмечалась зависимость между дефицитом влаги и содержанием свободного пролина [30, 31, 32].
Для свежесобранной хвои деревьев контрольного Ступинского насаждения характерны невысокие значения содержания свободного пролина, которые варьируют в достаточно узких (1.8–4.5 мкМ/г) пределах (рис. 4а). При сильном стрессе концентрация свободного пролина достоверно не изменяется (F = 4.2; p > 0.05) (рис. 4b), верхняя граница распределения признака повышается на 31% (1.8–6.6 мкМ/г), но 60% значений остаются в пределах нормы. Это свидетельствует о том, что не все растения способны адаптироваться к данной стрессовой нагрузке, в силу ее чрезмерности. Существенные изменения содержания пролина (F = 17.1; p < 0.05) происходят при умеренном, постепенно нарастающем стрессе (рис. 4c). В этом случае отдельные значения повышаются в 9 и более раз, область распределения признака (3.8–26.9 мкМ/г) значительно расширяется, амплитуда возрастает в 8.5 раз (рис. 4d-3), среднее значение – почти в 4 раза. В пределах нормы остается только 10% значений.
Рис. 4.
Содержание свободного пролина (а–c) и его амплитуда (d) в хвое деревьев культур сосны обыкновенной: засухоустойчивого сорта сосна ‘Острогожская’ и Ступинского тест-объекта (контроль) в нативном состоянии (а), при сильном (b) и умеренном (c) гидротермическом стрессе. По вертикали: а, b, c – содержание свободного пролина, мкМ/г; d – амплитуда изменения признака, мкМ/г. По горизонтали: d – 1 – нативное состояние, 2 – сильный гидротермический стресс, 3 – умеренный гидротермический стресс. Светлые столбики – культуры сосны Ступинского тест-объекта; темные столбики – культуры сосны сорта ‘Острогожская’. Fig. 4. Free proline content (a–c) and its range (d) in Scots pine needles of: drought-tolerant variety ‘Ostrogozhskaya’ and trees of Stupinsky cultivated forest (control) in the native state (a), under severe (b) and moderate (c) hydrothermal stress. X-axis: а, b, c – free proline content, μM/g; d – range of free proline content change, μM/g. Y-axis: d – 1 – native state, 2 – severe hydrothermal stress, 3 – moderate hydrothermal stress. Light columns – Scots pine of Stupinsky cultivated forest (control); dark columns – plantations of drought-tolerant Scots pine ‘Ostrogozhskaya’.
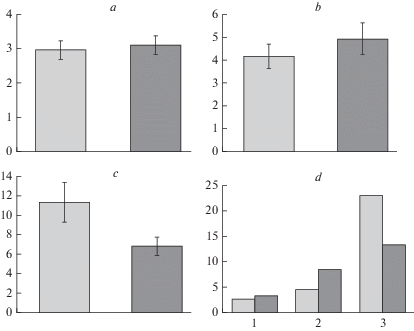
Аналогичные изменения, но значительно менее выраженные, зафиксированы и у сосны сорта ‘Острогожская’. Модельные деревья опытного насаждения по сравнению с деревьями контрольного насаждения, как было показано и при анализе водного режима, сходным образом реагируют на сильный стресс и менее остро на условия умеренного стресса. Кроме того установлено, что исходно низкие значения признака, близкие к значениям, наблюдаемым у сосны Ступинского насаждения (1.5–5.0 мкМ/г), возрастают в обоих вариантах опыта. При сильном стрессе содержание свободного пролина у деревьев опытного насаждения увеличивается в среднем в 1.6 раза (рис. 4b), что по интенсивности отклика аналогично контролю (F = 0.7; p > 0.05). Область распределения значений признака становится существенно шире за счет сдвига верхней границы (1.4–9.7 мкМ/г) относительно нативных образцов (F = 6.2; p < 0.05), однако 53% значений не превышают норму.
В варианте умеренного воздействия (рис. 4c) стресс-реакция оказалась при сравнении с контрольным насаждением менее выраженной (F = 5.2; p < 0.05). Среднее значение возрастает только в 2.2 раза (у сосны в Ступино в 9 и более раз), область распределения признака увеличивается примерно в 4 раза (F = 14.7; p < 0.05), но 47% значений (3.0–16.2 мкМ/г) не выходят за пределы нормы реакции. Зафиксированы повышения отдельных значений максимума в 4 раза. Статистически достоверные различия между двумя вариантами лабораторного опыта отсутствуют (F = 2.7; p > 0.05).
Различия реакций сосны сорта ‘Острогожская’ и сосны Ступинского насаждения хорошо демонстрируют обобщенные данные физиолого-биохимического анализа, представленные на примере изменения по сравнению с нативным состоянием трех параметров: содержания влаги, количества коллоидно-связанной воды и содержания пролина после сильной и умеренной стрессовой нагрузки (рис. 5а, b).
Рис. 5.
Изменение физиолого-биохимических параметров в опытных (сосна ‘Острогожская’) и контрольных (Ступинский тест-объект) образцах хвои сосны обыкновенной при действии стресса в процентах от значений ex situ: а – сильный, b – умеренный стресс. По вертикали: величина изменения признака, %. По горизонтали: 1 – общее содержание влаги, 2 – количество коллоидно-связанной воды, 3 – содержание свободного пролина. Светлые столбики – культуры сосны Ступинского тест-объекта; темные столбики – культуры сосны сорта ‘Острогожская’. Fig. 5. Changes in physiological and biochemical parameters of Scots pine needles in the experimental (‘Ostrogozhskaya’) and control (Stupinsky cultivated forest) samples under the stress as a percentage of ex situ values: a – severe, b – moderate stress. X-axis: rate of traist change, %. Y-axis: 1 – total moisture content, 2 – amount of colloid bound water, 3 – free proline content. Light columns – Scots pine of Stupinsky cultivated forest (control); dark columns – plantations of drought-tolerant Scots pine ‘Ostrogozhskaya’.
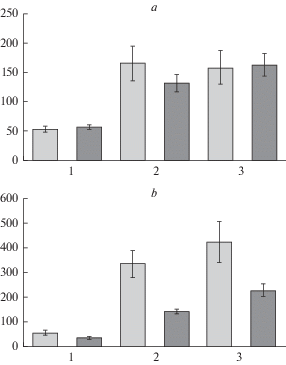
Эти признаки связаны с устойчивостью растений к засухе и показывают, что отклик на стресс, присущий деревьям в типичных для ЦЧР культурах сосны обыкновенной (Ступинский тест-объект) и деревьям сосны засухоустойчивого сорта ‘Острогожская’, является однотипным по характеру, но разным по интенсивности. В обоих вариантах лабораторного эксперимента имеет место снижение общего количества влаги и увеличение водного дефицита, повышение содержания коллоидно-связанной воды и пролина. Модальные значения изученных параметров у деревьев обоих насаждений в пробах свежесобранной хвои сопоставимы. Хорошо видно, что при высокой гидротермической нагрузке (рис. 5а) различия также находятся в пределах статистической ошибки, благодаря одинаковой реакции растений на сильный стресс. Важно отметить, что различия между объектами наиболее отчетливо выявляются во время умеренного стресса (рис. 5b). Несмотря на однотипность реакции (уменьшение содержания влаги, увеличение коллоидно-связанной воды, водного дефицита и свободного пролина) деревья Ступинского тест-объекта более остро отреагировали на его воздействие, чем деревья сосны засухоустойчивого сорта ‘Острогожская’. Отклик по содержанию коллоидно-связанной воды и свободного пролина у них был почти в два раза интенсивнее.
Известно, что все виды стресса ведут к расширению изменчивости признаков [33, 34]. Существует мнение, что в экстремальной ситуации, растение мобилизует все энергетические ресурсы (резервы), пытаясь выжить [35]. При умеренной стрессовой нагрузке реализуются потенциальные возможности, связанные с перестройкой метаболизма. Когда раздражитель действует в течение длительного времени, у растений появляется возможность приспособиться к нему [36]. Некоторые авторы предполагают, что данный механизм устойчивости связан в первую очередь с изменением экспрессии генов и синтезом стрессовых белков [37]. При сильном или усиливающемся стрессе наступает фаза истощения. В этом случае могут быть задействованы иные механизмы выживания растений [38]. Полученные нами результаты по однотипности преобразования норм реагирования сосны на сильный стресс полностью согласуются с литературными данными [7, 33, 36, 38].
В экспериментах ex vivo установлено, что при умеренном и сильном стрессе норма реакции экологически зависимых физиолого-биохимических признаков по сравнению с нативными образцами существенно расширяется. Несмотря на присущие исследованным насаждениям особенности, направленность изменений физиолого-биохимических параметров при воздействии стресса в целом совпадает. Причем, при умеренной нагрузке диапазон изменчивости признаков максимален у растений контрольного Ступинского тест-объекта; при более сильной нагрузке происходит сужение нормы реакции. Сильный гидротермический стресс в значительной степени “уравнивает” возможности деревьев к адаптации вне зависимости от генотипа и места произрастания объекта. Возможно, при такой повреждающей нагрузке растения не пытаются приспособиться, и выбирают наиболее приемлемый и наименее энергозатратный индивидуальный путь защиты и выживания. К тому же жесткие условия требуют немедленной ответной реакции. Можно предположить, что способов выживания на пределе возможностей не так уж много, и они должны быть общими для всех растительных организмов.
Очевидно, что лабораторные эксперименты не могут однозначно приравниваться к процессам, происходящим в естественных условиях. Отклик физиолого-биохимических параметров вегетативной сферы сосны обыкновенной в определенной степени согласуется с характером изменения нормы реакции параметров семенной продуктивности (полнозернистость семян, число семян на шишку) при сильной (1991 и 2007 гг.) и умеренной (1995 г.) засухе в условиях Ступинского тест-объекта [39]. Полученные данные подтвердили, что вегетативная сфера сосны обыкновенной достаточно устойчива к основному региональному стрессору – засухе. Аналогичный вывод относится и к генеративной сфере, о чем свидетельствует сохранение ее репродуктивного потенциала в период заложения генеративных почек [4] во время самой сильной и длительной за всю историю наблюдений засухи 2010 г.
ЗАКЛЮЧЕНИЕ
В результате сравнительного исследования ряда физиолого-биохимических параметров хвои (общее количество и дефицит влаги, количество коллоидно-связанной воды, содержание свободного пролина) у деревьев сосны обыкновенной засухоустойчивого сорта ‘Острогожская’ и деревьев сосны в типичных для ЦЧР культурах Ступинского тест-объекта (контроль) на территории Воронежской области установлено, что их модальные значения и норма реакции существенно не различаются в оптимальные по погодным условиям годы (за исключением общего количества влаги). Для деревьев опытного и контрольного насаждений характерны достаточно низкие значения анализируемых признаков, и минимальный размах их изменчивости. Это свидетельствует о том, что в районе исследований сосна обыкновенная характеризуется достаточно высоким адаптивным потенциалом и устойчивостью к региональным флуктуациям климата.
Полученные данные показывают, что вид обладает достаточным запасом экологической емкости к дополнительной гидротермической нагрузке. При имитации засухи в лабораторных условиях ex vivo индуцированный стресс выводит физиолого-биохимические показатели хвои сосны в обоих насаждениях из состояния равновесия, в значительной степени расширяя диапазон их изменчивости. При сильном стрессе адаптивные возможности деревьев обоих насаждений являются сходными. Умеренный стресс сильно расширяет границы нормы реакции признаков у деревьев сосны в культурах Ступинского тест-объекта. У сосны обыкновенной сорта ‘Острогожская’ существенных различий исследованных параметров при сильном и умеренном индуцированном стрессе не обнаружено (за исключением общего содержания влаги).
По многим проанализированным физиолого-биохимическим признакам у деревьев контрольного насаждения выявлен значительно более интенсивный отклик на индуцированную засуху. Для сосны сорта ‘Острогожская’ характерна менее острая стресс-реакция, что, вероятно, является следствием наличия у нее определенных адаптаций к условиям засухи. Это позволяет деревьям с наименьшими затратами переживать стрессовые ситуации и быстрее возвращаться к нормальному равновесному состоянию, что, по-видимому, можно рассматривать как один из физиологических механизмов засухоустойчивости.
Список литературы
Hamanishi E.T., Campbell M.M. 2011. Genome-wide responses to drought in forest trees. – Forestry. 84 (3): 273–283. https://doi.org/10.1093/forestry/cpr012
Переведенцев Ю.П., Гоголь Ф.В., Наумов К.М., Шаталинский К.М. 2007. Глобальные и региональные изменения климата на рубеже ХХ и ХХI столетий. – Вестник ВГУ. Сер. Геогафия. Геоэкология. 2: 5–12. http://www.vestnik.vsu.ru/pdf/geograph/2007/02/2007-02-01.pdf
Глобальные и региональные изменения климата и природной среды позднего кайнозоя в Сибири. 2008. Новосибирск. 511 с.
Свинцова В.С., Кузнецова В.Ф., Пардаева Е.С. 2014. Влияние засухи на генеративную сферу и жизнеспособность пыльцы сосны обыкновенной. – Лесоведение. 3: 49–57.
Онучин А.А., Буренина Т.А., Зирюкина Н.В., Фарбер С.К. 2014. Лесогидрологические последствия рубок в условиях Средней Сибири. – Сиб. лесной журн. 1: 110–118. http://сибирскийлеснойжурнал.рф/articles/archive/arkhiv-2014/lesogidrologicheskie-posledstviya-rubok-v-usloviyakh-sredney-sibiri-/
Орлов А.Я., Кошельков С.П. 1977. Почвенная экология сосны. М. 323 с.
Судачкова Е.А. 1998. Состояние и перспективы изучения влияния стрессов на древесные растения. – Лесоведение. 2: 3–8.
Генкель П.А. 1982. Физиология жаро- и засухоустойчивости растений. М. 280 с.
Шварц С.С. 1980. Экологические закономерности эволюции. М. 378 с.
Глотов Н.В., Тараканов В.В. 1985. Норма реакции генотипа и взаимодействие генотип-среды в природных популяциях. – Журн. общ. биол. 46 (6): 760–770.
Кузнецова Н.Ф. 2003. Направления отбора в жизненном цикле сосны обыкновенной. – Лесоведение. 4: 9–16.
Кузнецова Н.Ф. 2017. Засухоустойчивая сорт-популяция сосна Острогожская. – Современная лесная наука: проблемы и перспективы. Материалы Всероссийской научно-практической конференции 20-22 декабря 2017 года. – Воронеж. С. 93–98. http://www.vniilgisbiotech.org/pdf/sbornik_sovr_les_nauk.pdf
Середа Л.О., Яблонских Л.А., Куролап С.А. 2015. Оценка эколого-геохимического состояния почвенного покрова городского округа город Воронеж. – Вестник ВГУ. Серия: География. Геоэкология. 4: 59–65. http://www.vestnik.vsu.ru/pdf/geograph/2015/04/2015-04-11.pdf
Доспехов Б.А. 1973. Методика полевого опыта. М. 306 с.
Починок Х.Н. 1976. Методы биохимического анализа растений. Киев. 334 с.
Bates L.S., Waldren R.P., Teare I.D. 1973. Rapid determination of free proline for water stress studies. – Plant Soil. 39 (1): 205–207. https://doi.org/10.1007/BF00018060
Тарханов С.Н. 2011. Индивидуальная изменчивость биохимических признаков и состояние форм сосны обыкновенной в условиях аэротехногенного загрязнения. – Лесоведение. 2: 62–69.
Клушевская Е.С., Кузнецова Н.Ф. 2016. Оценка устойчивости сосны обыкновенной к засухе по физиологическим характеристикам хвои. – Лесоведение. 3: 216–222.
Кузнецов В.В., Дмитриева Г.А. 2006. Физиология растений: учебник. М. 742 с.
Jami S.K., Anuradha T.S., Guruprasad L., Kirti P.B. 2007. Molecular, biochemical and structural characterization of osmotin-like protein from black nightshade (Solanum nigrum). – J. Plant Physiol. 164 (3): 238–252. https://doi.org/10.1016/j.jplph.2006.01.006
Velasco-Conde T., Yakovlev I., Majada J.P., Aranda I., Johnsen Ø. 2012. Dehydrins in maritime pine (Pinus pinaster) and their expression related to drought stress response. – Tree Genetics & Genomes. 8(5): 957–973. https://doi.org/10.1007/s11295-012-0476-9
Arumingtyas E.L., Savitri E.S., Purwoningrahayu R.D. 2013. Protein Profiles and Dehydrin Accumulation in Some Soybean Varieties (Glycine max L. Merr) in Drought Stress Conditions. – Amer. J. Plant Scien.. 4 (1): 134–141. https://doi.org/10.4236/ajps.2013.41018
Ristic Z., Gifford D.J., Gass D.D. 1991. Heat shock protein in two lines of Zea mays L. that differ in drought and heat resistance. – Plant Physiol. 97 (4): 1430–1434. https://doi.org/10.1104/pp.97.4.1430
Пономарева Т.В., Кузьмина Н.А., Кузьмин С.Р. 2009. Влияние влагозапасов почвы на рост видов хвойных в условиях эксперимента. – Вестн. Красноярского Гос. Аграрного ун-та. 12: 45–49.
Urli M., Porté A.J., Cochard H., Guengant Y., BurlettR., Delzon S. 2013. Xylem embolism threshold for catastrophic hydraulic failure in angiosperm trees. – Tree Physiology. 33 (7): 672–683. https://doi.org/10.1093/treephys/tpt030
Чиркова Т.В. 2002. Физиологические основы устойчивости растений: Учебное пособие студентов биологических факультетов вузов. СПб. 244 с.
Samuel D., Ganesh G., Yang P.-W., Chang M.-M., Wang S.-L., Hwang K.-C., Yu C., Jayaraman G., Kumar T.K.S., Trivedi V.D., Chang D.-K. 2000. Proline inhibits aggregation during protein refolding. – ProteinScience. 9 (2): 344–352. https://doi.org/10.1110/ps.9.2.344
Mohammadkhani N., Heidari R. 2008. Drought-induced Accumulation of Soluble Sugars and Proline in Two Maize Varieties. – World Applied Sciences Journal. 3 (3): 448–453.
Кузнецов В.В., Шевякова Н.И. 1999. Пролин при стрессе: биологическая роль, метаболизм, регуляция. – Физиол. раст. 46 (2): 321–336.
Бирюкова З.П. 1986.Свободный пролин как показатель физиологического состояния сосны обыкновенной. – Физиол. раст. 33 (5): 1027–1030.
Gao D., Gao Q., Xu H.-Y., Ma F., Zhao C.-M., Liu J.-Q. 2009. Physiological responses to gradual drought stress in the diploid hybrid Pinus densata and its two parental species. – Trees. 23: 717–728. https://doi.org/10.1007/s00468-009-0314-3
Corcuera L., Gil-Pelegrin E., Notivol E. 2012. Aridity promotes differences in proline and phytohormone levels in Pinus pinaster populations from contrasting environments. – Trees. 26: 799–80. https://doi.org/10.1007/s00468-011-0651-x
Levitt J. 1980. Responses of plants to environmental stresses. Vol. 1. Chilling, freezing and high temperature stresses. New York. 497 p. https://doi.org/10.2307/3899731
Котов М.М. 1997. Изменчивость сосны обыкновенной по адаптивным признакам в связи с условиями произрастаний. – Лесоведение. 3: 51–60.
Семихатова О.А. 1995. Дыхание поддержания и адаптация растений. – Физиол. раст. 42 (2): 312–319.
Пахомова В.М. 1995. Основные положения современной теории стресса и неспецифический адаптационный синдром у растений. – Цитология. 1 (2): 66–91.
Arnholdt-Schmitt B. 2004. Stress-induced cell reprogramming. A role for global genome regulation? – Plant Physiol. 136: 2579–2586. https://doi.org/10.1104/pp.104.042531
Титов А.Ф., Акимова Т.В., Таланова А.А., Топчиева Л.В. 2006. Устойчивость растений в начальный период действия неблагоприятных температур. М. 143 с.
Кузнецова Н.Ф. 2015. Развитие неспецифической и специфической реакций у Pinus sylvestris L. на популяционном уровне в стрессовом градиенте засушливых лет. – Экология. 5: 332–338.
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы