

Растительные ресурсы, 2021, T. 57, № 2, стр. 145-157
Масса тонких корней в почвах лесных сообществ на постагрогенных землях в условиях средней тайги (на примере Республики Карелия)
А. Ю. Карпечко 1, *, А. В. Туюнен 1, М. В. Медведева 1, Е. В. Мошкина 1, И. А. Дубровина 2, Н. В. Геникова 1, В. А. Сидорова 2, А. В. Мамай 1, О. В. Толстогузов 3, Л. М. Кулакова 3
1 Институт леса – обособленное подразделение Федерального государственного бюджетного учреждения науки Федерального исследовательского центра “Карельский научный центр Российской академии наук”
г. Петрозаводск, Россия
2 Институт биологии – обособленное подразделение Федерального государственного бюджетного учреждения науки Федерального исследовательского центра “Карельский научный центр Российской академии наук”
г. Петрозаводск, Россия
3 Институт экономики – обособленное подразделение Федерального государственного бюджетного учреждения науки Федерального исследовательского центра “Карельский научный центр Российской академии наук”
г. Петрозаводск, Россия
* E-mail: anna.karpechko@gmail.com
Поступила в редакцию 07.05.2020
После доработки 24.09.2020
Принята к публикации 10.12.2020
Аннотация
Определена и проанализирована масса тонких корней (диаметром до 2 мм) в лесных сообществах, которые сформировались на участках, использовавшихся в прошлом с сельскохозяйственными целями. Объекты изучения располагаются в среднетаежной подзоне, вблизи г. Петрозаводска (южный агроклиматический район Республики Карелия). Пробные площади были заложены в смешанном мелколиственном сообществе возрастом 20 лет и в 65- и 110-летних еловых лесах. Установлено, что наиболее высокая масса тонких корней характерна для лиственного древостоя, наиболее низкая – для 65-летнего елового древостоя с примесью лиственных пород. С увеличением возраста древостоя процентная доля тонких корней в общей подземной фитомассе снижается. Показано, что соотношение массы тонких корней березы и ели европейской не соответствует их доле в общем запасе стволовой древесины. Наибольший запас подстилки характерен для 110-летнего ельника, наименьший – для 20-летнего березняка. В еловых лесах при увеличении возраста древостоя от 65 до 110 лет доля тонких корней в общем запасе подстилки не изменяется.
Леса, возобновляющиеся на землях постагрогенного использования, являются эффективным поглотителем углерода [1–4]. В настоящее время Россия занимает лидирующие позиции по площади земель, выведенных из сельскохозяйственного оборота [5, 6]. Лесовосстановление в одних и тех же физико-географических условиях может проходить по разным сценариям [7–9]. Площадь зарастающих земель, тип почвы, таксационные показатели окружающих насаждений, а также вид сельскохозяйственного использования участков до момента его прекращения – все это отражается на показателях восстанавливающихся экосистем [10–12].
Основная функция тонких корней – снабжение дерева питательными веществами и водой [13–16]. Они играют важную роль в круговороте углерода в связи с более высокой скоростью разложения, по сравнению с надземными частями деревьев [17]. Согласно некоторым исследованиям [16, 18], доля тонких корней в ежегодной чистой первичной продукции бореальных лесов составляет 32%, при этом их участие в общей древесной биомассе составляет менее 5%. Тонкие корни прямо и опосредованно влияют на почвенные биогеохимические циклы [16]. По данным K. Vogt et al. [19], Clemmensen et al. [20] корни и микроорганизмы ризосферы являются источником 50–70% содержащегося в почве углерода. В некоторых экосистемах тонкие корни являются основным источником поступления органического вещества в почву [21].
Существуют разные градации деления корней на фракции по толщине. Чаще всего к тонким относят корни с диаметром в диапазоне от 0.6 до 5 мм. А.Я. Орлов [22] выделял фракцию корней диаметром до 0.6 мм. М.И. Калинин [14], L. Finer et al. [23], A. Lehtonen et al. [24] к тонким корням относят корни с диаметром до 2-х мм. В других работах [25, 26] корни делились на две группы: с диаметром менее 2 мм и от 2 до 5 мм. Иногда выделяют фракцию тонких корней с диаметром менее 1 мм [27, 28].
Накопление элементов минерального питания в корнях зависит от их размеров. Тонкие корни (диаметром до 2 мм) содержат большее количество зольных элементов и азота, чем более крупные корни. Согласно оценкам Н.И. Казимирова и Р.М. Морозовой [29], в мелких корнях количество зольных элементов и азота на единицу веса в 1.4 раза больше, чем в корнях диаметром до 1 см и в 2 раза больше, чем в корнях диаметром более 1 см. Кроме того, количество азота в корнях возрастает с повышением плодородия почвы [29]. Эти факты подчеркивают важность оценки массы тонких корней и изучение ее динамики.
Свойства почв после прекращения сельскохозяйственного использования постепенно изменяются по кислотно-щелочным показателям, пулу микро- и макроэлементов, ферментативной активности, характеристикам биотического компонента. Изменяется количество и качество опада, который интенсивнее вовлекается в природный круговорот [30–32]. Трудно минерализуемые остатки хвойных растений медленно разлагаются в почве, что является причиной формирования мощных грубогумусных лесных подстилок [33], в то время как опад лиственных растений положительно влияет на биологическую активность почв, процессы его микробиологической трансформации происходят быстрее [34]. Взаимосвязь и взаимообусловленность свойств почв и развития корней определяют необходимость их комплексного изучения.
Связующим звеном различных компонентов биогеоценоза являются лесные подстилки. На фоне антропогенного воздействия происходит изменение состава органического вещества почв, ризосферных процессов, микробиологических и экофизиологических показателей (рис. 1).
Анализ динамики накопления фитомассы на бывших сельхозземлях представляется актуальным, особенно это касается ее подземной части, как наименее изученной в связи с трудоемкостью работ и сложностью определения содержания корней в почве.
Целью настоящего исследования является оценка изменения массы и распределения в почве тонких корней древесных растений в период от 20 до 110 лет после прекращения сельскохозяйственного использования земель в условиях средней тайги.
МАТЕРИАЛ И МЕТОДЫ
Изучаемая территория располагается в среднетаежной подзоне Республики Карелия. Климат – континентальный с чертами морского [36]. Среднегодовая температура воздуха по многолетним наблюдениям составляет 2.0…2.5 °С, средняя температура января –9…–10 °С, июля +16…+17 °С. Для региона характерен режим избыточного увлажнения, сумма осадков составляет 550–750 мм в год [37]. По теплообеспеченности почв район относится к теплому, количество дней с температурой выше +10 °С составляет 115–125 [38].
На территории преобладают еловые (Picea abies (L.) Karst.) леса, которые могут сменяться мелколиственными после антропогенных нарушений. Почвенный покров характеризуется комплексностью. Широко распространены подзолистые почвы различной степени агрогенной трансформации. На фоне антропогенного воздействия (пашня, вырубки, урбанизация и пр.) происходит существенное изменение свойств почв. В целом для почв характерна природная ограниченность использования для нужд сельскохозяйственного производства, так как благоприятный период для вегетации растений относительно короткий [39].
Работа велась на участках, вовлеченных в прошлом в сельскохозяйственное использование. Они расположены в окрестностях г. Петрозаводска (61°44′53″ С 34°20′35″ В). Было заложено 3 пробные площади (ПП).
ПП 1 представлена 20-летним смешанным мелколиственным (Salix caprea L., Betula pubescens Ehrh., Alnus incana (L.) Moench) разнотравным сообществом. Почва агродерново-подзолистая типичная [40] легкосуглинистая на суглинистой морене. В почве наблюдаются признаки первоначального восстановления текстурной дифференциации. Общее проективное покрытие напочвенного покрова составляет 40%. Доминантом является вейник лесной Calamagrostis arundinaciae (L.) Roth., покрытие других видов незначительно. Моховой покров развит слабо, представлены Brachythecium salebrosum (F. Weber & D. Mohr) Bruch et al. – 10%, Climacium dendroides (Hedw.) F.Weber & D.Mohr – 1%.
ПП 2 заложена в 65-летнем ельнике черничном. Почва дерново-подзолистая постагрогенная [40] легкосуглинистая на суглинистой морене. По морфологическому строению изучаемая почва близка к естественным зональным, однако в ней еще наблюдаются признаки агрогенной трансформации. Общее проективное покрытие травяно-кустарничкового яруса составляет 20%, преобладают кислица Oxalis acetosella L., черника Vaccinium myrtillis L. и щитовник картузианский Dryopteris carthusiana (Vill.) H.P. Fuchs. Проективное покрытие мохово-лишайникового яруса составляет 5%, представлены Pleurozium shreberi (Willd. ex Brid.) Mitt., Dicranum scuparium Hedw., Rhodobrium roseum (Hedw.) Limpr.).
ПП 3 заложена в 110-летнем ельнике черничном. Почва подзолистая типичная [40] легкосуглинистая на суглинистой морене. Для нее характерна типичная текстурная дифференциация профиля, отсутствие гумусово-аккумулятивного горизонта. Напочвенный покров отличается высоким проективным покрытием Vaccinium myrtillis (40%), покрытие Oxalis acetosella составляет 15%, Vaccinium vitis-idaea L. – 10%. В моховом ярусе преобладают Pleurozium shreberi (20%) и Sphagnum girgensohnii Russow (15%).
Закладка пробных площадей и описание древостоев (табл. 1) проводилось в соответствии с общепринятыми методиками [41]. Были выполнены геоботанические описания с оценкой общего проективного покрытия и покрытия доминантных видов напочвенного покрова. На основании полученных таксационных показателей и плотности древесины [42] рассчитывалась общая фитомасса древостоя, в составе которой, исходя из известных соотношений [29, 43], выделялась фракция подземной фитомассы.
Таблица 1.
Таксационная характеристика древостоев Table 1. Forest stand characteristics
| ПП Sample plot |
Древесная порода Tree species |
Запас, м3 га–1 Standing stock, m3 ha–1 |
Состав Stand composition |
Общая фитомасса,
т га–1 Total phytomass, t ha–1 |
Число стволов,
экз. га–1 Number of stems, ind. ha–1 |
Средние Average |
Сумма площадей сечений, м2 га–1 Basal area, m2 ha–1 |
|
|---|---|---|---|---|---|---|---|---|
| диаметр, см diameter, cm |
высота, м height, m |
|||||||
| 1 | Ива Salix caprea |
77 | 38B52S10Al | 105 | 11 167 | 4.4 | 6.4 | 16.6 |
| Береза Betula pubescens |
56.1 | 8750 | 3.9 | 8.1 | 10.2 | |||
| Ольха Alnus incana |
15.6 | 667 | 7.7 | 8.9 | 3.05 | |||
| 2 | Ель Picea abies |
193 | 67Pic16Al 12Pop5B | 200 | 1444 | 14.6 | 13.9 | 24.1 |
| Ольха Alnus incana |
37.7 | 222 | 17.6 | 14.8 | 5.42 | |||
| Осина Populus tremula |
29.2 | 44 | 31.4 | 18.3 | 3.44 | |||
| Береза Betula pubescens |
15.6 | 44 | 23.7 | 17.2 | 1.96 | |||
| Рябина Sorbus aucuparia |
9.3 | 267 | 8.6 | 10.4 | 1.6 | |||
| Ива Salix caprea |
4.1 | 44 | 13.8 | 13.5 | 0.62 | |||
| 3 | Ель Picea abies |
459 | 95Pic4B1Pop | 319 | 575 | 28.5 | 27.3 | 36.8 |
| Береза Betula pubescens |
17.5 | 71 | 17.3 | 23.3 | 1.69 | |||
| Осина Populus tremula |
3.27 | 4 | 31.9 | 25.0 | 0.29 | |||
Массу тонких корней определяли методом монолитов [22]. На каждой ПП отбиралось по 10–15 монолитов, размер которых составлял 10 × 10 × 20 см. Из образцов извлекались корни, которые сортировались по размерам: до 1 и до 2 мм по диаметру. Далее корни высушивались до абсолютно сухого состояния при температуре 105 °С и взвешивались. Запас подстилок определяли термостатно-весовым методом, с пересчетом абсолютно сухого веса в т га–1 [44].
Достоверность различий величин исследованных параметров оценивалась с использованием критерия Стьюдента.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Для начальной стадии восстановления лесного покрова после прекращения использования участков в сельскохозяйственных целях в условиях среднетаежной подзоны характерно формирование лиственных древостоев с участием в составе березы пушистой, ивы козьей, ольхи серой (ПП 1). Этому способствуют благоприятные эдафические условия (повышенное содержание органического вещества), ежегодный высокий урожай семян в лиственных насаждениях, произрастающих на прилегающих территориях, а также высокая энергия роста надземных и подземных органов лиственных пород по сравнению с хвойными.
Масса тонких корней (диаметр менее 2 мм) древесных пород является более высокой в мелколиственном сообществе (ПП 1) по сравнению с другими изучаемыми сообществами (ПП 2, 3) (табл. 2). Это свидетельствует о характерной для постагрогенной экосистемы активной аккумуляции фитомассы. Полученный результат согласуется с данными других исследователей [45], согласно которым масса тонких корней диаметром до 2 мм достигает более высокого значения на участке, выведенном из сельскохозяйственного оборота 20 лет назад, по сравнению с контрольным лесным участком.
Таблица 2.
Масса тонких корней древесных пород в верхнем слое почвы Table 2. Tree species fine roots mass in the upper soil layer
| Общая подземная фитомасса, т га–1 Total underground phytomass, t ha–1 |
Масса корней древесных пород по фракциям диаметра Root mass of tree species by diameter |
|||
|---|---|---|---|---|
| т га–1 t ha–1 |
% от общей подземной фитомассы % of total underground phytomass |
|||
| 0–1 мм 0–1 mm |
0–2 мм 0–2 mm |
0–1 мм 0–1 mm |
0–2 мм 0–2 mm |
|
| ПП1 Березняк разнотравный 20 лет | ||||
| SP1 Betuletum herbosum 20 years | ||||
| 12.60 | 1.95 ± 0.23 | 3.21 ± 0.37 | 15.5 | 25.5 |
| ПП2 Ельник черничный 65 лет | ||||
| SP2 Piceetum myrtillosum 65 years | ||||
| 31.28 | 0.89 ± 0.08 | 1.78 ± 0.17 | 2.8 | 5.7 |
| ПП3 Ельник черничный 110 лет | ||||
| SP3 Piceetum myrtillosum 110 years | ||||
| 59.40 | 1.09 ± 0.09 | 2.58 ± 0.21 | 1.8 | 4.3 |
В березняке с ивой разнотравном (ПП 1) общая масса корней составляет 12.6 т га–1, из которой на долю тонких корней диаметром до 2 мм приходится 25.5% (табл. 2). Полученная величина значительно отличается от соответствующих величин в других изученных сообществах. Это объясняется тем, что мелколиственный древостой, отличающийся высокой густотой (более 20 тыс. экз. га–1), находится на этапе активного роста и нуждается в большом количестве минеральных веществ. Согласно имеющимся обзорным работам, в биоме бореальных лесов доля тонких корней в общей корневой массе составляет в среднем 16% [18].
Общая подземная фитомасса 65-летнего елового древостоя на ПП 2 составляет 31.28 т га–1 (табл. 2). На долю корней диаметром до 2 мм приходится около 6% от этой величины. В 110-летнем еловом древостое (ПП 3) общая подземная фитомасса равна 59.4 т га–1, тонкие корни диаметром до 2 мм составляют 4.3% (табл. 2). Полученные результаты согласуются с данными T. Sun et al. [16], согласно которым с возрастом доля тонких корней деревьев имела тенденцию к уменьшению в ряду 36-, 57- и 82-летних древостоев. В соответствии с более высокой общей подземной фитомассой в 110-летнем сообществе, масса корней древесных пород менее 2 мм в диаметре на ~30% больше, чем в 65-летнем ельнике.
При сравнении массы корней отдельных древесных пород (табл. 3) установлено, что в 65-летнем сообществе, несмотря на примерно 2-кратное преобладание по запасу ели европейской над лиственными породами, масса корней лиственных пород диаметром до 1 и до 2 мм на 40% больше (p < 0.05). Согласно некоторым данным [46] при одновременном или почти одновременном возобновлении ели и лиственных пород, корни ели при контакте с корнями лиственных пород нередко частично отмирают или сильно деформируются. Б.С. Мартинович [47] указывает, что корни березы обладают большей энергией роста по сравнению с корнями хвойных пород. Полученные результаты демонстрируют более высокую активность роста корней березы пушистой по сравнению с корнями ели европейской при освоении свободного пространства. Это было также отмечено и в других работах, посвященных изучению корневых систем [48]. В тоже время, согласно Г.А. Чибисову [46] при разложении корней березы в процессе ее естественного изреживания, условия для роста корневых систем ели улучшаются.
Таблица 3.
Масса тонких корней в верхнем слое почвы по породам Table 3. Fine root mass in the upper soil layer by tree species
| Корни лиственных пород, т га–1 Roots of deciduous species, t ha–1 |
Корни ели европейской, т га–1 Roots of Picea abies, t ha–1 |
||
|---|---|---|---|
| 0–1 мм 0–1 mm |
0–2 мм 0–2 mm |
0–1 мм 0–1 mm |
0–2 мм 0–2 mm |
| ПП1 Березняк разнотравный 20 лет | |||
| SP1 Betuletum herbosum 20 years | |||
| 1.95 ± 0.23 | 3.21 ± 0.37 | – | – |
| ПП2 Ельник черничный 65 лет | |||
| SP2 Piceetum myrtillosum 65 years | |||
| 0.56 ± 0.07 | 1.10 ± 0.14 | 0.33 ± 0.06 | 0.68 ± 0.10 |
| ПП3 Ельник черничный 110 лет | |||
| SP3 Piceetum myrtillosum 110 years | |||
| 0.09 ± 0.01 | 0.19 ± 0.04 | 1.00 ± 0.09 | 2.39 ± 0.21 |
В 110-летнем ельнике доля лиственных пород в составе древостоя составляет лишь 5%. К этому возрасту в значительной степени происходит их отпад. Доля корней лиственных пород в общей массе подземных органов (менее 10%), примерно соответствует их доле общей фитомассе древостоя.
Анализ распределения массы корней древесных растений по почвенным горизонтам (табл. 4) показал, что в 65-летнем сообществе в минеральном слое почвы масса корней диаметром до 1 и до 2 мм больше, чем в горизонте лесной подстилки. Для фракции до 2 мм отмеченные различия достоверны при p < 0.05. В 110-летнем древостое картина обратная, лесная подстилка содержит больше корней по сравнению с минеральным слоем почвы. Для фракции до 1 мм различия достоверны при p < 0.05. Полученный результат можно объяснить наличием на ПП 3 более мощной подстилки, слой которой имеет толщину в среднем 5 см, тогда как на ПП 2 он составляет 3 см. Кроме того, известно, что после окончания использования участка в сельскохозяйственном обороте, характерные для географических условий почвообразовательные процессы начинают играть ведущую роль в формировании почвы [49]. Для изучаемой территории характерен зональный подзолообразовательный процесс. Почвенный профиль 110-летнего древостоя характеризуется наличием обедненного элементами питания подзолистого горизонта и соответственно концентрация тонких корней происходит в лесной подстилке более богатой органическими веществами. В 20-летнем лиственном древостое анализ распределения корней по генетическим почвенным горизонтам выполнить не представлялось возможным вследствие недифференцированности почвенного профиля.
Таблица 4.
Масса тонких корней в верхних почвенных горизонтах Table 4. Fine root mass in the upper soil layer
| Масса корней в лесной подстилке, т га–1 Root mass in forest litter, t ha–1 |
Масса корней в минеральном слое, т га–1 Root mass in mineral layer, t ha–1 |
||
|---|---|---|---|
| 0–1 мм 0–1 mm |
0–2 мм 0–2 mm |
0–1 мм 0–1 mm |
0–2 мм 0–2 mm |
| ПП2 Ельник черничный 65 лет | |||
| SP2 Piceetum myrtillosum 65 years | |||
| 0.41 ± 0.05 | 0.71 ± 0.08 | 0.48 ± 0.07 | 1.07 ± 0.13 |
| ПП3 Ельник черничный 110 лет | |||
| SP3 Piceetum myrtillosum 110 years | |||
| 0.71 ± 0.09 | 1.34 ± 0.16 | 0.38 ± 0.04 | 1.24 ± 0.14 |
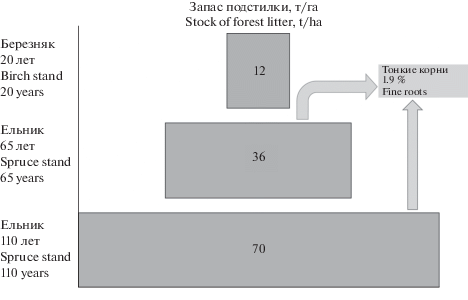
Запас подстилки в 20-летнем лиственном древостое по сравнению с хвойными относительно низкий (рис. 2). Это обусловлено более быстрым разложением опада в связи с высокой микробиологической активностью почв и более интенсивным круговоротом элементов питания [50]. Для хвойных древостоев характерно замедление темпа трансформации органического вещества и усиление процессов его консервации, эфемерные трофические связи, характерные для молодых сообществ, становятся устойчивыми [51]. В результате происходит формирование более мощных лесных подстилок, дифференциация почв на отдельные генетические горизонты становится заметнее. Депонирование N,C,P,K и других элементов питания растений в лесной подстилке приводит к перераспределению массы тонких корней с концентрацией их в органогенном горизонте. Независимо от возраста еловых древостоев масса тонких корней составляет около 2% от запаса подстилок. Можно предположить, что в почвах мелкие корни занимают определенный объем межпорового пространства, заполняют благоприятные для их развития локусы, создавая континуум живой части подземного пространства.
Рис. 2.
Содержание тонких корней в лесной подстилке. Fig. 2. Content of fine roots in forest litter.
ЗАКЛЮЧЕНИЕ
В условиях среднетаежной подзоны, земли, исключенные из сельскохозяйственного использования, относительно быстро зарастают древесной растительностью, чему способствует повышенное содержание органического вещества в почвенном профиле, характерное для постагрогенной сукцессии.
На территории южного агроклиматического района Республики Карелия в 20-летнем березняке разнотравном доля тонких корней в общей подземной фитомассе значительно (в 4.5–6 раз) выше, чем в мелколиственно-еловом и еловом сообществах. С увеличением времени, прошедшего с момента прекращения сельскохозяйственного использования территории, общая фитомасса древостоя и ее подземная часть, закономерно возрастают. При этом доля тонких корней в подземной фитомассе снижается. Это объясняется уменьшением величины прироста фитомассы и, соответственно, количества корней, необходимого для его обеспечения.
Доля древесной породы в общей фитомассе древостоя или запасе стволовой древесины не всегда соответствует доле тонких корней в подземной части древостоя. В 65-летнем сообществе при значительном участии в составе древостоя ели европейской (67% по запасу), масса ее корней диаметром до 2 мм на 40% меньше по сравнению с массой корней лиственных пород. Этот факт демонстрирует более активный рост корней лиственных.
Для лиственного древостоя характерен наиболее низкий запас подстилки. При восстановлении елового древостоя этот показатель возрастает и приходит в соответствие с величинами, характерными для зональных почв. Доля тонких корней в запасе подстилок исследуемых еловых лесов возрастом 65 и 110 лет одинакова.
Проведенное исследование позволило выявить специфику изменения массы и распределения в почве тонких корней древесных растений в лесных фитоценозах, находящихся в условиях постагрогенной трансформации среды. Представляется целесообразным активизировать изучение тонких корней, как компонента, активно участвующего в цикле углерода, в почвах лесных сообществ, формирующихся на бывших сельхозземлях. Это необходимо для более точной оценки потоков углерода и лучшего понимания роли постагрогенных экосистем в глобальных климатических процессах.
Список литературы
Fan S., Gloor M., Mahlman J., Pacala S., Sarmiento J., Takahashi T., Tans P. 1998. A large terrestrial carbon sink in North America implied by atmospheric and oceanic carbon dioxide data and models. – Science. 282: 442–6. https://doi.org/10.1126/science.282.5388.442
Silver W.L., Osterlag R., Lugo A.E. 2000. The potential for carbon sequestration through reforestation of abandoned tropical agricultural and pasture lands. – Restor. Ecol. 8(4): 394–407. https://doi.org/10.1046/j.1526-100x.2000.80054.x
Pacala S.W., Hurtt G.C., Baker D., Peylin P., Houghton R.A., Birdsey R.A., Heath L., Sundquist E.T., Stallard R.F., Ciais P., Moorcroft P., Caspersen J.P., Shevliakova E., Moore B. nine others. 2001. Consistent land- and atmosphere-based U.S. carbon sink estimates. – Science. 292: 2316 –20. https://doi.org/10.1126/science.1057320
Hooker T.D., Compton J.E. 2003. Forest ecosystem carbon and nitrogen accumulation during the first century after agricultural abandonment. – Ecol. Applicat. 13(2): 299–313. https://doi.org/10.1890/1051-0761(2003)013[0299:FECANA]2.0.CO;2
Люри Д.И., Горячкин С.В., Караева Н.А., Денисенко Е.А., Нефедова Т.Г. 2010 Динамика сельскохозяйственных земель России в XX веке и постагрогенное восстановление растительности и почв. М. 416 с.
Рыжова И.М., Ерохова А.А., Подвезенная М.А. 2015. Изменение запасов углерода в постагрогенных экосистемах в результате естественного восстановления лесов в Костромской области. – Лесоведение. 4: 307–317. https://www.elibrary.ru/item.asp?id=23906221&
Уткин А.И., Гульбе Т.А, Гульбе Я.И., Ермолова Л.С. 2002. О наступлении лесной растительности на сельскохозяйственные земли в Верхнем Поволжье. – Лесоведение. 5: 44–52.
Залесов С.В., Магасумова А.Г., Юровских Е.В. 2009. Зарастание бывших сельскохозяйственных угодий в Слободо-Туринском районе Свердловской области. – Леса России и хозяйство в них. 3(34): 14–23. http://usfeu.ru/media/filer_public/08/aa/08aacbee-61f0-4fd0-97b1-c4787d5c9b61/lesa_rossii_i_khoziaistvo_v_nikh__4_342009.pdf
Мошкина Е.В., Медведева М.В., Туюнен А.В., Карпечко А.Ю., Геникова Н.В., Дубровина И.А., Мамай А.В., Сидорова В.А., Толстогузов О.В., Кулакова Л.М. 2019. Особенности естественного восстановления лесных экосистем на бывших сельскохозяйственных землях (на примере южного агроклиматического района Карелии). – Биосфера. 11(3): 134–145. https://doi.org/10.24855/biosfera.v11i3.506
Houghton R.A., Hackler J.L., Lawrence K.T. 1999. The U.S. carbon budget: contributions from land-use change. – Science. 285: 574–578. https://doi.org/10.1126/science.285.5427.574
Caspersen J.P., Pacala S.W., Jenkins J.C., Hurtt G.C., Moorcroft P.R., Birdsey R.A. 2000. Contributions of land-use history to carbon accumulation in U.S. forests. – Science. 290: 1148–1151. https://doi.org/10.1126/science.290.5494.1148
Goodale C.L., Aber J.D., McDowell W.H. 2000. The long-term effects of disturbance on organic and inorganic nitrogen export in the White Mountains, New Hampshire. – Ecosystems. 3: 433–450. https://doi.org/10.1007/s100210000039
Рахтеенко И.Н. 1952. Корневые системы древесных и кустарниковых пород. М., Л. 107 с.
Калинин М.И. 1983. Формирование корневой системы деревьев. М. 152 с.
Ehrenfeld J.G., Parsons W.F.J., Han X., Parmelee R.W., Zhu W. 1997. Live and dead roots in forest soil horizons: contrasting effects on nitrogen dynamics – Ecology. 78(2): 348–362. https://doi.org/10.1890/0012-9658(1997)078[0348:LADRIF]2.0.CO;2
Sun T., Dong L.L., Mao Z.J., Li Y.Y. 2015. Fine root dynamics of trees and understory vegetation in a chronosequence of Betula platyphylla stands. – Forest Ecology and Management. 346: 1–9. https://doi.org/10.1016/j.foreco.2015.02.035
Ruess R., Hendrick R., Burton A., Pregitzer K., Sveinbjornsson B., Allen M., Maurer G. 2003. Coupling fine root dynamics with ecosystem carbon cycling in black spruce forests of interior Alaska. – Ecol. Monogr. 73(4): 643–662. https://doi.org/10.1890/02-4032
Yuan Z.Y., Chen Y.H. 2010. Fine root biomass, production, turnover rates, and nutrient contents in boreal forest ecosystems in relation to species, climate, fertility, and stand age: Literature review and meta-analyses. – CRC Crit. Rev. Plant Sci. 29(4): 204–221. https://doi.org/10.1080/07352689.2010.483579
Vogt K.A., Vogt D.J., Palmiotto P.A., Boon P., O’Hara J., Asbjornsen H. 1995. Review of root dynamics in forest ecosystems grouped by climate, climatic forest type and species. – Plant Soil. 187(2): 159–219. https://doi.org/10.1007/BF00017088
Clemmensen K.E., Bahr A., Ovaskainen O., Dahlberg A., Ekblad A., Wallander H., Stenlid J., Finlay R.D., Wardle D.A., Lindahl B.D. 2013. Roots and associated fungi drive long term carbon sequestration in boreal forest. – Science. 339: 1615–1618. https://doi.org/10.1126/science.1231923
Jackson R.B., Mooney H.A., Schulze E.D. 1997. A global budget for fine root biomass, surface area, and nutrient contents. – PNAS. 94(14): 7362–7366. https://doi.org/10.1073/pnas.94.14.7362
Орлов А.Я. 1967. Метод определения массы корней деревьев в лесу и возможность учета годичного прироста органической массы в толще лесной почвы. – Лесоведение. 1: 64–70.
Finér L., Ohashi M., Noguchi K., Hirano Y. 2011. Fine root production and turnover in forest ecosystems in relation to stand and environmental characteristics. – Forest Ecol. Manag. 262(11): 2008–2023. https://doi.org/10.1016/j.foreco.2011.08.042
Lehtonen A., Palviainen M., Ojanen P., Kalliokoski T., Nöjd P., Kukkola M., Penttilä T., Mäkipää R., Leppälammi-Kujansuu J., Helmisaari H.-S. 2016. Modelling fine root biomass of boreal tree stands using site and stand variables. – Forest Ecol. Manag. 359: 361–369. https://doi.org/10.1016/j.foreco.2015.06.023
Vanninen P., Mäkelä A. 1999. Fine root biomass of Scots pine stands differing in age and soil fertility in southern Finland. – Tree Physiology. 19(12): 823–830. https://doi.org/10.1093/treephys/19.12.823
Helmisaari H.-S., Derome J., Nöjd P., Kukkola M. 2007. Fine root biomass in relation to site and stand characteristics in Norway spruce and Scots pine stands. – Tree Physiology. 27(10): 1493–1504. https://doi.org/10.1093/treephys/27.10.1493
Усольцев В.А. 1988. Рост и структура фитомассы древостоев. Новосибирск. 253
Усольцев В.А., Залесов С.В. 2005. Методы определения биологической продуктивности насаждений. Екатеринбург. 147 с.
Казимиров Н.И., Морозова Р.М. 1973. Биологический круговорот веществ в ельниках Карелии. Л. 175 с.
Владыченский А.С., Телеснина В.М., Чалая Т.А. 2012. Влияние растительного опада на химические свойства и биологическую активность постагрогенных почв южной тайги. – Вестник Московского университета. Сер. 17. Почвоведение. 1: 3–10. https://www.elibrary.ru/item.asp?id=17637057
Телеснина В.М. 2015. Постагрогенная динамика растительности и свойств почвы в ходе демутационной сукцессии в южной тайге. – Лесоведение. 4: 293-306. https://www.elibrary.ru/item.asp?id=23906220
Телеснина В.М., Ваганов И.Е., Карлсен А.А., Иванова А.Е., Жуков М.А., Лебедев С.М. 2016. Особенности морфологии и химических свойств постагрогенных почв южной тайги на легких отложениях (Костромская область) – Почвоведение. 1: 115–129. https://doi.org/10.7868/S0032180X16010111
Прокушкин С.Г. 1992. Динамика минерализации соснового опада. – Лесоведение. 4: 28–34.
Кузнецов М.А. 2010. Влияние условий разложения и состава опада на характеристики и запас подстилки в среднетаежном чернично-сфагновом ельнике. – Лесоведение. 6: 54–60. https://www.elibrary.ru/item.asp?id=15510017
Neina D. 2019. The role of soil pH in plant nutrition and soil remediation. – Appl. Environ. Soil Sci.9: 1–9. https://doi.org/10.1155/2019/5794869
Научно-прикладной справочник по климату СССР. 1988. 3 (1-6). Л. 693 с.
Романов А.А. 1961. О климате Карелии. Петрозаводск. 139 с.
Атлас Карельской АССР. Москва. 1989. 40 с.
Федорец Н.Г., Морозова Р.М., Солодовников А.Н. 2003. Лесные почвы Карелии и оценка их продуктивности. – Труды Карельского научного центра РАН. 5: 108–120. http://resources.krc.karelia.ru/transactions/doc/trudy2003/trudy_2003_5_108-120.pdf
Шишов Л.Л., Тонконогов В.Д., Лебедева И.И., Герасимова М.И. 2004. Классификация и диагностика почв России. Смоленск. 342 с.
ОСТ 56-69-83. Площади пробные лесоустроительные. Метод закладки.
Полубояринов О.И. 1976. Плотность древесины. М. 160 с.
Казимиров Н.И., Морозова Р.М., Куликова В.К. 1979. Органическая масса и потоки веществ в березняках средней тайги. Л. 216 с.
Методические рекомендации по определению запасов лесной подстилки и ее зольности при лесоводственных исследованиях. М. 1979. 38 с.
Кондратова А.В., Абрамова Е.Р. 2018. Особенности формирования тонких корней на различных стадиях восстановления постагрогенных экосистем в зоне южной тайги. – Успехи современного естествознания. 9: 18–22. https://www.natural-sciences.ru/pdf/2018/9/36860.pdf
Чибисов Г.А. 1971. Рост корневых систем в березово-еловых насаждениях и влияние на них рубок ухода. – Сб. работ по лесному хозяйству и лесохимии. Архангельск. С. 121–130.
Мартинович Б.С., Кабашникова Г.И., Крот Л.А. 1982. Особенности роста и жизнедеятельности ели и березы в смешанных елово-березовых насаждениях. – В сб.: Регулирование роста, развития и питания растений в фитоценозах. Минск. С. 49–78.
Schmid I. 2002. The influence of soil type and interspecific competition on the fine root system of Norway spruce and European beech. – Basic Appl. Ecol. 3(4): 339–346. https://doi.org/10.1078/1439-1791-00116
Почвообразовательные процессы. 2006. М. 510 с.
Германова Н.И., Медведева М.В., Мамай А.В. 2012. Динамика разложения листового опада в среднетаежных насаждениях Карелии. – Известия вузов. Лесной журнал. 1: 24–32.
Зябченко С.С., Дьяконов В.В., Федорец Н.Г., Синькевич Т.А. 1994. Лесные экосистемы заповедника “Кивач”. – В сб.: Структурно-функциональная организация лесных почв среднетаежной подзоны Карелии. Петрозаводск. С. 5–37.
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы