

Растительные ресурсы, 2021, T. 57, № 4, стр. 320-332
Морфология и прорастание семян Nepeta formosa и N. bucharica (Lamiaceae)
Т. В. Елисафенко 1, *, А. Ю. Асташенков 1, Н. Ю. Курочкина 1
1 ФГБУН Центральный сибирский ботанический сад СО РАН
г. Новосибирск, Россия
* E-mail: tveli@ngs.ru
Поступила в редакцию 02.02.2021
После доработки 11.02.2021
Принята к публикации 15.09.2021
Аннотация
Изучены особенности морфологии эремов и семян и динамика прорастания семян центральноазиатских видов Nepeta formosa Kudrjasch. и N. bucharica Lipsky. Анализ семян из природных популяций (in situ) и собранных в условиях ex situ показал, что коэффициент вариации по всем выбранным признакам не превышает 20%. Возраст растений и условия произрастания больше влияют на размеры эремов и семян у N. bucharica, чем у N. formosa. Большое число эремов N. formosa (86%) из природных популяций не имели зародыша, или он был деформирован. В условиях ex situ у растений второго года жизни, у обоих видов, 80–90% семян имели сформированный зародыш. Грунтовая всхожесть, определенная для семян N. bucharica, собранных in situ, составила 7.7%, лабораторная всхожесть достигала 20.0% (после 8 мес. хранения). Установлено, что у семян двух видов в условиях in situ и ex situ динамика прорастания сходна. Различия наблюдались в зависимости от срока хранения и происхождения семян (ex situ и in situ). Лабораторная всхожесть выше у семян, собранных в условиях культуры. У свежесобранных семян всхожесть после 15 дней опыта составляла около 60% у обоих видов. Репродуктивная способность N. bucharica оказалась более адаптивна к изменению экологических и погодных условий, по сравнению с N. formosa.
В сем. Lamiaceae род Nepeta L. один из самых крупных, его представители распространены главным образом в горных областях Центральной Азии. Виды Nepeta относятся к числу важных в практическом и экономическом отношении растений, многие из них известны как пищевые, пряно-ароматические и лекарственные [1, 2]. Метаболиты видов рода Nepeta находятся в центре внимания исследователей всего мира в связи с широким спектром их биологической активности [3–8]. Большой группой вторичных метаболитов, непосредственно участвующих в механизме адаптации, являются фенольные соединения [9–11]. Их высокая физиологическая активность коррелирует с целебными свойствами в организме человека. Фенольные соединения способствуют противокашлевому, мочегонному, антиастматическому, антисептическому, спазмолитическому и жаропонижающему действию видов Nepeta [12]. Некоторые виды используются для лечения ушибов, ревматических болей, лихорадки, кожных высыпаний [13, 14]. Сообщалось об антимикробных, в том числе антикандидных, противоопухолевых и противовоспалительных свойствах некоторых видов Nepeta [15–17]. Помимо этих ценных качеств некоторые виды Nepeta обладают естественными репеллентными свойствами в отношении домашних насекомых [18]. В настоящее время проводятся исследования на цитотоксичность малатиона и эфирного масла N. crispa Willd. в отношении клеточных линий позвоночных и беспозвоночных животных для борьбы с вредителями сельского хозяйства [19].
Под действием антропогенной нагрузки вследствие перевыпаса скота и заготовки сырья, а также изменения экологических условий природные популяции видов рода Nepeta находятся под угрозой. В связи с этим, необходимы обоснованные рекомендации и планомерная работа по сохранению естественных и созданию интродукционных популяций ценных и перспективных видов, в том числе исчезающих и эндемичных. Начальным этапом такой работы является изучение морфологии семян и особенностей их прорастания. Для исследования мы выбрали два вида Nepeta (N. formosa Kudrjasch., N. bucharica Lipsky), которые перспективны для использования. Сведения о морфологии эремов и семян, а также биологии прорастания этих видов в литературе отрывочные [20]. Ранее нами был изучен онтогенез особей N. formosa, N. bucharica в природных популяциях Центральной Азии [21, 22].
Цель исследования состояла в изучение биологии прорастания и морфологии семян и эремов Nepeta formosa, N. bucharica в зависимости от их происхождения.
МАТЕРИАЛЫ И МЕТОДЫ
Nepeta formosa – эндемик Центральной Азии. Вид произрастает от среднегорий до высокогорий в пределах от 1700 до 3000 м над уровнем моря на северных, северо-восточных и восточных макросклонах и на соответственно ориентированных экспозициях гор. Предпочитает тенистые мезофильные местообитания, растет по речным водоразделам, каменистым берегам речек и ручьев, реже по саям ущелий. Вид обычен в поясах чернолесья, крупнотравных полусаванн, арчовников, степей и субальпийских лугов. Встречается в составе разнотравных и высокотравных кленовников (Acer regelii Pax), ореховых лесов (Juglans regia L.), березняков (Betula tianschanica Rupr.), экзохордников (Exochorda racemosa (Lindl.) Rehder) и юганников (Prangos pabularia Lindl.), зарослей Rosa kokanica (Regel) Regel ex Juz., торонников (Polygonum coriarium Grig.) и разнотравных лугов [23, 24]. Материал по изучению семян N. formosa собран из естественной популяции (in situ), расположенной в ущелье р. Кугарт (Республика Кыргызстан) Ферганского хребта (правый борт безымянного второстепенного ущелья, травянистая группировка на каменистых выходах крутого склона на верхней границе пояса редколесий (Crataegus spp.).
N. bucharica – эндемик среднеазиатских горных систем Тянь-Шаня и Памиро-Алая. В пределах Центральной Азии этот вид произрастает от пояса чернолесья до субальпийских лугов. Он приурочен к увлаженным местообитаниям, обитает на альпийских лужайках, по берегам рек, ручьев, родников и водопадов на щебнисто-каменистых субстратах, конусах выноса склонов различных экспозиций. Обычен в составе люцерновых костровников (Medicago lupulina L., Bromus oxyodon Schrenk), шульховников (Rumex paulsenianus Rech. f.), кленовников (Acer regelii), розариев (Rosa kokanica), юганников (Prangos pabularia) и полынников (Artemisia dracunculus L.). Нередко встречается как сорное растение в посевах [23, 24]. Материал для изучения семян N. bucharica собран in situ в Республике Кыргызстан, Таласский Алатау, перевал Алабель, верховья р. Чичкан. Крупнообломочное русло реки, замшелый галечник, альпийский мезопсихрофитный луг.
Кроме эремов из природных популяций сбора 5–11 августа 2017 г., изучали эремы особей первого и второго года жизни, выращенных в условиях ex situ, собранные в 2018 г. (конец июля–начало августа) и 22 июля 2019 г. на экспериментальном участке Центрального сибирского ботанического сада СО РАН.
Изучение морфологии эремов и семян
Массу эремов определяли путем трехкратного взвешивания 100 шт., среднее арифметическое значение пересчитывали на 1000 шт. При изучении макроморфологии эремы замачивали на сутки, измеряли под бинокуляром в 20-кратной повторности. Семя извлекали с помощью продольного разреза лезвием по стенке эрема и помещали в каплю воды. Зародыш освобождали от семенной кожуры препаровальной иглой. Определяли метрические показатели (мм): длину, ширину, толщину эрема, длину и ширину цикатрикса и семени, длину зародыша, семядолей, оси зародыша (зародышевый корешок и гипокотиль). Вычисляли аллометрические признаки: отношение ширины эрема к его длине, отношение ширины цикатрикса к его длине, отношение ширины семядолей к их длине, отношение длины семени к длине эрема, отношение длины зародыша к длине эрема, отношение длины оси зародыша к длине зародыша, отношение длины семядолей к длине зародыша. Исследования проводили в центре микроскопического оборудования ЦСБС СО РАН на стереомикроскопе Carl Zeiss Stereo Discovery V12 с цветной цифровой камерой высокого разрешения AxioCam MRс-5 с программой AxioVision 4.8 для получения, обработки и анализа изображений. При описании эремов применяли терминологию З.М. Артюшенко и А.А. Федорова [25, 26], Э.С. Терехина [27]
Изучение прорастания семян
Для определения полевой (грунтовой) всхожести семян эремы высевали 21 мая 2018 г. по 100 шт. на 1 погонный метр в трех повторностях на экспериментальном участке. Всходы отмечали в течение месяца с интервалом 5–7 дней.
Для определения лабораторной всхожести эремы проращивали при комнатной температуре (20–22 °С) в чашках Петри на слое бумажного фильтра с ватной подложкой в 3-кратной повторности по 100 эремов. Семя считалось проросшим при формировании корешка размером с семя. Проросшие семена учитывали от начала прорастания ежедневно в течение недели, далее – через день. Использовали семена свежесобранные (начало опыта – 30 сентября) и те, которые хранились 8 месяцев в комнатных условиях (начало опыта 12–27 марта). Всхожесть определяли как процент проросших семян в течение опыта.
По результатам опыта устанавливали длительность периода от начала опыта до прорастания семян, продолжительность периода прорастания, всхожесть семян (%), энергию прорастания (%). Энергию прорастания определяли как процент семян, проросших в первые пять дней от начала прорастания [28]. В работе использован показатель “интенсивность энергии прорастания”, который рассчитывается как отношение всхожести к энергии прорастания (%) [29].
Результаты обрабатывали методами математической статистики. Определяли среднее арифметическое значение (М), его ошибку (m), коэффициент вариации (V). Различия средних оценивали по критерию Стьюдента при 95%-ном уровне вероятности [30]. Для построения графика использовали данные варианта с максимальной всхожестью семян.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
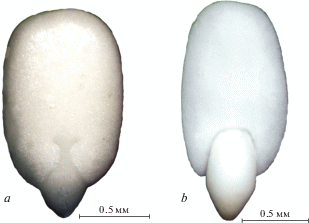
Эремы обоих видов сплюснутые в дорсивентральном направлении. У N. formosa эремы овальной формы, темно-коричневого цвета, N. bucharica – обратнояйцевидной формы, коричневого цвета (рис. 1а, b; 4а, b). Цикатрикс эремов изучаемых видов V-образной формы, у N. formosa он имеет тупой угол в основании, длину 0.31–0.34 мм и ширину 0.56–0.62 мм. У N. bucharica угол в основании острый, длина 0.47–0.50 мм, ширина 0.55–0.59 мм (табл. 1). Вся поверхность цикатрикса имеет слой, несущий белые сферические образования.
Рис. 1.
Общий вид эрема Nеpeta bucharica. а – вентральная сторона, b – латеральная сторона. Fig. 1. Nepeta bucharica erem. a – ventral side, b – lateral side.

Таблица 1.
Биометрическая характеристика эремов, семян и зародышей Nepeta formosa и Nepeta bucharica в условиях культуры (ЦСБС, СО РАН, г. Новосибирск) Table 1. Biometric characteristics of erems, seeds and embryos of Nepeta formosa and Nepeta bucharica under cultivation in Central Siberian Botanical Garden, Novosibirsk
| Признак Feature |
M ± m | V, % | lim | M ± m | V, % | lim | M ± m | V, % | lim | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N. formosa | |||||||||||
| ex situ | in situ | ||||||||||
| 1 года жизни one-year-old plants |
2 года жизни two-year-old plants |
||||||||||
| Эрем Erem |
Длина, мм Length, mm |
1.76 ± 0.03 | 5.04 | 1.59–1.9 | 1.71 ± 0.03 | 7.41 | 1.53–2.1 | 1.92 ± 0.04 | 7.36 | 1.55–2.07 | |
| Ширина, мм Width, mm |
1.09 ± 0.02 | 7.37 | 0.96–1.2 | 1.01 ± 0.02 | 7.27 | 0.83–1.16 | 1.06 ± 0.03 | 9.81 | 0.93–1.31 | ||
| Ширина/длина Width/length |
0.62 ± 0.01 | 5.39 | 0.56–0.68 | 0.59 ± 0.01 | 8.06 | 0.48–0.67 | 0.56 ± 0.02 | 10.67 | 0.48–0.72 | ||
| Толщина, мм Thickness, mm |
0.76 ± 0.02 | 8.92 | 0.63–0.83 | 0.74 ± 0.01 | 7.16 | 0.63–0.85 | 0.72 ± 0.02 | 8.45 | 0.6–0.83 | ||
| Цикатрикс Cicatrix |
Длина, мм Length, mm |
0.34 ± 0.01 | 11.91 | 0.29–0.41 | 0.31 ± 0.01 | 14.34 | 0.24–0.41 | 0.33 ± 0.01 | 11.75 | 0.25–0.4 | |
| Ширина, мм Width, mm |
0.62 ± 0.01 | 5.79 | 0.56–0.67 | 0.06 ± 0.01 | 5.94 | 0.52–0.66 | 0.56 ± 0.02 | 12.08 | 0.47–0.69 | ||
| Ширина/длина Width/length |
1.87 ± 0.07 | 12.05 | 1.54–2.31 | 2.01 ± 0.07 | 15.67 | 1.27–2.54 | 1.70 ± 0.06 | 13.45 | 1.34–2.06 | ||
| Семя Seed |
Длина, мм Length, mm |
1.55 ± 0.04 | 5.85 | 1.44–1.67 | 1.55 ± 0.04 | 10.86 | 1.37–1.92 | 1.53 ± 0.04 | 7.48 | 1.42–1.77 | |
| Ширина, мм Width, mm |
0.88 ± 0.04 | 11.27 | 0.73–1.01 | 0.94 ± 0.03 | 11.71 | 0.71–1.19 | 0.80 ± 0.03 | 13.33 | 0.66–0.98 | ||
| Длина семени/длина эрема Seed length/erem length |
0.86 ± 0.02 | 6.25 | 0.79–0.93 | 0.91 ± 0.01 | 5.80 | 0.83–1.05 | 0.81 ± 0.02 | 9.66 | 0.72–0.93 | ||
| Зародыш Embryo |
Длина, мм Length, mm |
Общая General |
1.48 ± 0.07 | 9.79 | 1.3–1.65 | 1.65 ± 0.04 | 8.58 | 1.46–2.02 | 1.7 | – | – |
| Ось зародыша Embryo axis |
0.56 ± 0.03 | 10.40 | 0.5–0.63 | 0.64 ± 0.01 | 9.24 | 0.57–0.77 | 0.67 | – | – | ||
| Семядоли Cotyledon |
1.24 ± 0.08 | 13.10 | 1.04–1.43 | 1.38 ± 0.03 | 8.75 | 1.17–1.74 | 1.38 | – | – | ||
| Ширина, мм Width, mm |
Семядоли Cotyledon |
0.85 ± 0.05 | 12.43 | 0.78–1.0 | 0.87 ± 0.02 | 7.37 | 0.77–0.99 | 0.86 | – | – | |
| Ось зародыша Embryo axis |
0.35 ± 0.02 | 10.71 | 0.32–0.39 | 0.38 ± 0.00 | 4.97 | 0.34–0.41 | 0.41 | – | – | ||
| Длина зародыша/длина эрема Embryo length/erem length |
0.80 ± 0.05 | 11.92 | 0.68–0.92 | 0.97 ± 0.01 | 5.77 | 0.91–1.14 | 0.87 | – | – | ||
| Длина оси зародыша/длина зародыша Length of embryo axis/embryo length |
0.38 ± 0.01 | 7.04 | 0.34–0.41 | 0.39 ± 0.00 | 4.96 | 0.34–0.41 | 0.40 | – | – | ||
| Длина семядолей/длина зародыша Cotyledon lengt /embryo length |
0.84 ± 0.02 | 4.94 | 0.8–0.88 | 0.84 ± 0.01 | 4.89 | 0.71–0.87 | 0.81 | – | – | ||
| Ширина семядоли/длина семядоли Cotyledon width/cotyledon length |
0.69 ± 0.03 | 8.17 | 0.61–0.75 | 0.62 ± 0.01 | 7.88 | 0.51–0.73 | 0.62 | – | – | ||
| N. bucharica | |||||||||||
| ex situ | in situ | ||||||||||
| 1 года жизни one-year-old plants |
2 года жизни two-year-old plants |
||||||||||
| Эрем Erem |
Длина, мм Length, mm |
2.38 ± 0.03 | 6.20 | 2.16–2.83 | 2.29 ± 0.03 | 8.12 | 1.98–2.63 | 2.42 ± 0.03 | 4.27 | 2.25–2.56 | |
| Ширина, мм Width, mm |
1.17 ± 0.01 | 6.74 | 1.08–1.36 | 1.07 ± 0.01 | 5.76 | 0.97–1.2 | 1.18 ± 0.01 | 4.51 | 1.11–1.29 | ||
| Ширина/длина Width / length |
0.49 ± 0.01 | 8.37 | 0.40–0.58 | 0.47 ± 0.01 | 7.73 | 0.39–0.53 | 0.49 ± 0.00 | 3.29 | 0.46–0.52 | ||
| Толщина, мм Thickness, mm |
0.75 ± 0.01 | 7.54 | 0.64–0.85 | 0.69 ± 0.01 | 7.33 | 0.54–0.82 | 0.78 ± 0.01 | 7.20 | 0.71–0.94 | ||
| Цикатрикс Cicatrix |
Длина, мм Length, mm |
0.48 ± 0.01 | 10.48 | 0.42–0.65 | 0.47 ± 0.01 | 11.14 | 0.32–0.57 | 0.50 ± 0.01 | 7.06 | 0.42–0.55 | |
| Ширина, мм Width, mm |
0.55 ± 0.01 | 12.62 | 0.4–0.69 | 0.56 ± 0.01 | 11.18 | 0.45–0.7 | 0.59 ± 0.02 | 11.9 | 0.45–0.7 | ||
| Ширина/длина Width/length |
1.15 ± 0.03 | 16.03 | 0.63–1.49 | 1.20 ± 0.03 | 13.83 | 0.9–1.59 | 1.17 ± 0.03 | 9.22 | 0.94–1.35 | ||
| Семя Seed |
Длина, мм Length, mm |
2.1 ± 0.02 | 2.67 | 2.03–2.19 | 1.99 ± 0.02 | 6.91 | 1.75–2.3 | 2.16 ± 0.04 | 6.51 | 1.92–2.35 | |
| Ширина, мм Width, mm |
1.00 ± 0.02 | 4.12 | 0.93–1.06 | 0.94 ± 0.01 | 8.89 | 0.75–1.07 | 1.02 ± 0.03 | 10.02 | 0.9–1.21 | ||
| Длина семени/длина эрема Seed length / erem length |
0.91 ± 0.01 | 3.73 | 0.87–0.98 | 0.87 ± 0.01 | 7.96 | 0.68–1.05 | 0.88 ± 0.01 | 3.99 | 0.81–0.93 | ||
| Зародыш Embryo |
Длина, мм Length, mm |
Общая General |
2.12 ± 0.03 | 4.18 | 1.96–2.19 | 2.04 ± 0.02 | 7.44 | 1.72–2.39 | 2.15 ± 0.05 | 8.22 | 1.73–2.33 |
| Ось зародыша Embryo axis |
0.8 ± 0.03 | 9.41 | 0.64–0.87 | 0.8 ± 0.01 | 9.95 | 0.62–0.94 | 0.83 ± 0.03 | 10.32 | 0.71–1.03 | ||
| Семядоли Cotyledon |
1.83 ± 0.02 | 3.60 | 1.71–1.92 | 1.71 ± 0.02 | 7.74 | 1.39–1.95 | 1.85 ± 0.06 | 9.98 | 1.38–2.00 | ||
| Ширина, мм Width, mm |
Семядоли Cotyledon |
1.05 ± 0.01 | 3.28 | 0.99–1.1 | 0.94 ± 0.02 | 9.68 | 0.78–1.1 | 1.06 ± 0.03 | 9.19 | 0.84–1.23 | |
| Ось зародыша Embryo axis |
0.42 ± 0.02 | 11.24 | 0.34–0.48 | 0.39 ± 0.01 | 10.96 | 0.29–0.48 | 0.44 ± 0.01 | 8.33 | 0.38–0.52 | ||
| Длина зародыша/длина эрема Embryo length/erem length |
0.92 ± 0.02 | 5.58 | 0.86–1.00 | 0.89 ± 0.01 | 9.86 | 0.67–1.03 | 0.88 ± 0.02 | 7.87 | 0.71–0.96 | ||
| Длина оси зародыша/длине зародыша Length of embryo axis/embryo length |
0.38 ± 0.01 | 8.51 | 0.31–0.42 | 0.39 ± 0.00 | 7.15 | 0.34–0.46 | 0.39 ± 0.01 | 7.21 | 0.36–0.44 | ||
| Длина семядолей/длина зародыша Cotyledon length/embryo length |
0.87 ± 0.01 | 1.68 | 0.84–0.88 | 0.84 ± 0.00 | 2.35 | 0.80–0.88 | 0.86 ± 0.01 | 3.61 | 0.80–0.91 | ||
| Ширина семядоли/длина семядоли Cotyledon width/cotyledon length |
0.58 ± 0.01 | 1.63 | 0.56–0.59 | 0.55 ± 0.01 | 6.90 | 0.46–0.64 | 0.57 ± 0.01 | 7.43 | 0.49–0.64 | ||
Примечание. “–” – нет данных, M – среднее арифметическое значение, m – его ошибка, V – коэффициент вариации, lim – диапазон между минимальным и максимальным значением. Note. “–” – no data, M – arithmetic mean, m – its error, V – coefficient of variation, lim – the range between the minimum and maximum values.
Масса 1000 семян изученных видов имела различные значения: N. formosa: 0.31 ± 0.01 г (in situ), 0.35 ± 0.04 г (ex situ 2018) и 0.29 ± 0.03 г (ex situ 2019); N. bucharica – 0.37 ± 0.01 г (in situ) и 0.45 ± 0.01 г (ex situ 2019). Семенная кожура семян N. formosa и N. bucharica пленчатая, светло-коричневого цвета. С брюшной стороны семени располагается халаза коричневого цвета, которая проходит от микропиле до халазальной части семени (рис. 2а, b; 4с). Эндосперм отсутствует, зародыш молочного цвета, крупный, семядоли четко отграничены от оси зародыша, основания семядолей почти смыкаются (рис. 3а, 4d). Ось зародыша состоит из радикулы, гипокотиля и апекса побега. Однако гипокотиль и апекс побега морфологически не выражены. Ось зародыша в очертании ромбовидно-каплевидной формы, наибольшая ширина находится на уровне основания семядолей (рис. 3b).
Рис. 2.
Семя Nеpeta bucharica. a – вентральная, b – дорсальная сторона. Fig. 2. Nepeta bucharica seed. a – ventral side, b – dorsal side.

Рис. 3.
Зародыш Nеpeta bucharica. a – общий вид, b – в разрезе. Fig. 3. Nеpeta bucharica embryo. a – general view, b – cross-section.
Статистический анализ размеров эремов, семени и зародыша выявил достоверные межвидовые различия у растений как в условиях in situ, так в условиях ex situ, кроме одного аллометрического признака – отношения длины семядолей к длине зародыша (табл. 2). Таким образом, пропорциональность сформированного зародыша одинакова у обоих видов независимо от мест произрастания. В условиях in situ и ex situ семядоли составляют 81–87% зародыша. Защитные покровы зародыша (эрема и семени) мягкие, тонкие, зародыш хорошо дифференцирован и имеет крупные семядоли, что обусловливает отсутствие физического покоя семян и быстрое их прорастание.
Таблица 2.
Критерий Стъюдента для межвидовой, межвозрастной и межпопуляционной достоверности различий Nepeta formosa и Nepeta bucharica Table 2. Student’s t-test of the interspecific, inter-age and inter-population significance of differences in Nepeta formosa and Nepeta bucharica
| Признак Feature |
N. formosa | N. bucharica | N. formosa–N. bucharica | ||||||
|---|---|---|---|---|---|---|---|---|---|
| ex situ: 1 год жизни –2 год жизни ex situ: one-year-old –two-year-old |
ex situ–in situ
(2 год жизни) ex situ–in situ (two-year-old) |
ex situ: 1 год жизни – 2 год жизни ex situ: one-year-old – two-year-old |
ex situ–in situ
(2 год жизни) ex situ–in situ (two-year-old) |
in situ
(2 год жизни) in situ (two-year-old) |
in situ
(1 год жизни) in situ (one-year-old) |
ex situ | |||
| Эрем Erem |
Длина Length |
1.34 | 4.44* | 2.24* | 3.32* | 2.44* | 15.94* | 10.87* | |
| Ширина Width |
2.55* | 1.62 | 5.56* | 6.78* | 3.96* | 2.90* | 3.77* | ||
| Ширина/длина Width/length |
1.56 | 1.94 | 2.4* | 2.80* | 13.49* | 9.74* | 4.12* | ||
| Толщина Thickness |
0.96 | 0.97 | 4.06* | 5.31* | 8.04* | 0.61 | 2.83* | ||
| Цикатрикс Cicatrix |
Длина Length |
1.88 | 1.8 | 1.17 | 2.96* | 19.01* | 9.45* | 12.37* | |
| Ширина Width |
1.28 | 2.17* | 0.69 | 1.42 | 15.11* | 4.24* | 1.13 | ||
| Ширина/длина Width/length |
1.43 | 3.31* | 1.10 | 0.73 | 7.41* | 9.38* | 7.89* | ||
| Семя Seed |
Длина Length |
0.08 | 0.34 | 3.61* | 3.44* | 2.32* | 13.0* | 10.94* | |
| Ширина Width |
1.17 | 3.28* | 2.83* | 2.31* | 7.43* | 2.62* | 4.73* | ||
| Длина семени/длина эрема Seed length/erem length |
2.08* | 3.42* | 2.48* | 0.31 | 6.69* | 2.26* | 2.32* | ||
| Зародыш Embry |
Длина Length |
Общая General |
2.18* | – | 1.97 | 2.03 | 2.42* | 8.07* | – |
| Ось зародыша Embryo axis |
2.22* | – | 0.04 | 1.20 | 9.91* | 5.82* | – | ||
| Семядоли Cotyledon |
1.71 | – | 3.51* | 2.33* | 3.26* | 7.05* | – | ||
| Ширина Width |
Семядоли Cotyledon |
0.43 | – | 5.32* | 3.30* | 8.01* | 3.84* | – | |
| Оось зародыша Embryo axis |
1.16 | – | 1.62 | 3.75* | 22.04* | 2.7* | – | ||
| Длина зародыша/длина эрема Embryo length/erem length |
3.29* | – | 1.35 | 0.55 | 8.59* | 2.28* | – | ||
| Длина оси зародыша/длина зародыша Length of embryo axi /embryo length |
0.22 | – | 1.23 | 0.23 | 14.63* | 0.23 | – | ||
| Длина семядолей/длина зародыша Cotyledon length/embryo length |
1.14 | – | 3.55* | 1.39 | 1.72 | 1.39 | – | ||
| Ширина семядол / длина семядоли Cotyledon width/cotyledon length |
1.82 | – | 3.13* | 3.95* | 9.85* | 3.95* | – | ||
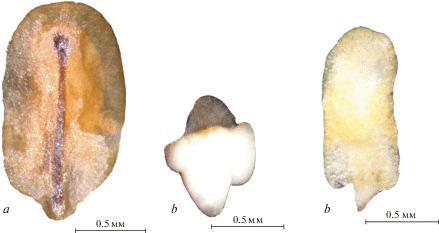
Нами отмечено, что сформированные эремы могут иметь дефектные зародыши (рис. 5). Так, из природной популяции N. formosa только 14% семян имели сформированный зародыш, а у остальной части – зародыш отсутствовал или был дефектный. У семян ex situ процент нормальных зародышей намного выше. У растений этого вида первого года жизни семена со сформированным зародышем составляют 33%, у растений второго года – 80%. У особей N. bucharica, изученных в природных популяциях, этот показатель высокий 87%. Самый низкий показатель сформированности семян N. bucharica приходится на первый год жизни – 25%, у особей второго года жизни он составляет 93%. Таким образом, условия, созданные на экспериментальных участках, благоприятны для развития семян изученных видов.
Рис. 4.
Nepeta formosa. a, b – общий вид эрема, c – семя, d – зародыш. Fig. 4. Nepeta formosa. a, b – general view of erem, c – seed, d – embryo.

В природной популяции эремы N. formosa сформировались более крупными, чем в ex situ (1.92 ± 0.04 мм и 1.71 ± 0.03 соответственно), однако размеры семян не имели достоверных различий (табл. 1, 2). Метрические значения, скорее всего, связаны с возрастом особей данного вида, что влияет на размеры зародыша, который был крупнее у растений второго года жизни. Стоит отметить, что длина зародыша составила у них 97% от длины эрема, в то время как у растений первого года жизни этот показатель достоверно ниже – 80%.
Установлено, что на размеры эремов, семян и зародышей N. bucharica влияли условия произрастания и возраст растений (табл. 1, 2). Так, эремы у растений, изученных в природных популяциях крупнее, чем в условиях ex situ (длина 2.42 ± 0.03 и 2.29 ± 0.03 мм соответственно). Одновременно с этим, проведенный анализ показал, что пропорциональность сложения зародыша одинакова (длина семядолей от длины зародыша составляла 84–86%).
В результате биометрического анализа семян растений N. formosa и N. bucharica из природных популяций и в условиях ex situ установлено, что коэффициент вариации по всем выбранным признакам не превышал 20% у обоих видов. Пропорциональность сложения зародыша, определяемая как отношение длины семядолей к длине зародыша, оказалась признаком, имеющим самую низкую изменчивость (не более 5%).
Рис. 5.
Nepeta formosa. a – семя с дефектным зародышем, b, c – дефектный зародыш. Fig. 5. Nepeta formosa. a – seed with a defective embryo, b, c – defective embryo.
Грунтовая (полевая) всхожесть семян N. bucharica (7.7%) оказалась в два раза меньше, чем лабораторная (18.7%). Первые всходы при грунтовом посеве появились на 15 день. Лабораторная всхожесть у свежесобранных семян растений N. bucharica и N. formosa, выращенных в условиях ex situ, составила около 60%, у семян природных популяций в 3.3–3.9 раза меньше. В лабораторных условиях семена прорастали на 2–4 день (рис. 6). Они прорастали одинаково быстро в течение 8–10 дней, энергия прорастания семян N. formosa варьировала от 8 до 48%, N. bucharica – 17–40%. Интенсивность энергии прорастания была высокая (более 60%), то есть большинство семян прорастали за 5 дней (табл. 3). Для сравнительного анализа всхожести свежесобранных семян и семян первого года хранения мы выбрали семена N. formosa. Оказалось, что всхожесть семян после 8 мес. хранения достигла 72%, но для этого потребовалось 140 дней. Динамика прорастания семян у изученных видов имела схожий характер и различия наблюдались в зависимости от срока хранения и их происхождения (ex situ и in situ).
Рис. 6.
Динамика прорастания семян Nepeta bucharica и Nepeta formosa. По горизонтали – дни, по вертикали – всхожесть, %. Fig. 6. Dynamics of seed germination in Nepeta bucharica and Nepeta formosa. X-axis – days; y-axis – germination, %.

Таблица 3.
Характеристика прорастания семян N. bucharica и N. formosa при различных условиях опыта Table 3. Germination of N. bucharica and N. formosa seeds under different experimental conditions
| Вид Species |
Происхождение, период
хранения (месяцы) Origin, duration of storage, (months) |
Всхожесть,% Germination, % |
Период, дни Period, days |
Энергия прорастания, % Germination energy, % |
Интенсивность энергии
прорастания, % Intensity of germination nergy, % |
||
|---|---|---|---|---|---|---|---|
| до прорастания prior to germination |
продолжительност
опыта duration of experiment |
прорастания germination |
|||||
| N. bucharica | in situ, 8 | 17–20 | 2 | 81 | 3–8 | 17–19 | 90–100 |
| ex situ, 1 | 62 | 4 | 12 | 10 | 40 | 65 | |
| N. formosa | in situ, 8 | 9–24 | 4 | 81 | 30 | 8–16 | 75–89 |
| ex situ, 8 | 22–72 | 4 | 140 | 120–140 | 12–48 | 41–67 | |
| ex situ, 1 | 58 | 4 | 12 | 10 | 35 | 60 | |
ЗАКЛЮЧЕНИЕ
В результате изучения эремов и семян центральноазиатских видов Nepeta formosa и N. bucharica из природных популяций (in situ) на территории Республики Кыргызстан (Ферганский хребет, Таласский Алатау) и полученных в условиях ex situ (Центральный сибирский ботанический сад СО РАН) выявлены особенности их морфологии и дана оценка жизнеспособности семян на основе показателей энергии прорастания, грунтовой и лабораторной всхожести. Эремы изученных видов сплюснутые в дорсивентральном направлении: N. formosa – овальной формы, темно-коричневого цвета, N. bucharica – обратнояйцевидной формы, коричневого цвета. Защитные покровы зародыша мягкие и тонкие, зародыш хорошо дифференцирован, имеет крупные семядоли. Эндосперм отсутствует. Зародыш крупный, семядоли четко отграничены от оси зародыша, которая в очертании ромбовидно-каплевидной формы. Пропорциональность сформированного зародыша одинакова у обоих видов независимо от мест произрастания (в условиях in situ и ex situ семядоли составляют 81–87% зародыша). Семена без периода покоя. Условия ex situ являются благоприятными для формирования семян изученных видов, что подтверждается лабораторной всхожестью и наличием полноценных семян. Выявлено, что возраст растений и условия произрастания больше влияют на размеры эремов и семян N. bucharica, чем N. formosa. Лабораторная всхожесть семян, полученных ex situ (около 60%) в 3–4 раза выше, чем семян из природных популяций. По совокупности полученных данных (морфологии семян и биологии их прорастания) установлено, что репродуктивная способность N. bucharica более адаптивна в ответных реакциях на изменение экологических и погодных условий по сравнению с N. formosa.
Список литературы
Растительные ресурсы СССР: Цветковые растения, их химический состав, использование семейства Hippuridaceae – Lobeliaceae. 1991. СПб. 200 с.
Растительные ресурсы России: дикорастущие цветковые растения, их компонентный состав и биологическая активность. 2011. Т. 4. СПб.; М. 630 с.
Pooter H.Z., Nicolai B., Buyck L.F., Goetghebeur P., Schamp N.M. 1987. The essential oil of Nepeta nuda. Identification of a new nepetalactone triastereoisomer. – Phytochemistry. 26(8): 2311–2314. https://doi.org/10.1016/S0031-9422(00)84709-3
Тропникова И.В., Буданцев А.Л., Зенкевич И.Г. 1998. Содержание и состав эфирных масел видов рода Nepeta L. – Растительные ресурсы. 34(4): 84–103.
Formisano C., Rigano D., Senatore F. 2011. Chemical constituents and biological activities of Nepeta species. – Chem. Biodivers. 8(10): 1783–1818. https://doi.org/10.1002/cbdv.201000191
Работягов В.Д., Аксенов Ю.В. 2014. Компонентный состав эфирного масла из видов рода Nepeta L. – Фармация и фармакология. 2(6(7)): 25–28. https://doi.org/10.19163/2307-9266-2014-2-6(7)-25-28
Astashenkov A.Yu., Cheryomushkina V.A., Myrzagalieva A.B., Medeubaeva B. 2019. Ontogenesis, estimation of coenopopulation state and component composition in Nepeta densiflora individuals of East Kazakhstan. – International Journal of Environmental Studies. 76(4): 634–647. https://doi.org/10.1080/00207233.2018.1509496
Sarikurkcu C., Eskici M., Karanfil A., Tepe B. 2019. Phenolic profile, enzyme inhibitory and antioxidant activities of two endemic Nepeta species: Nepeta nuda subsp. glandulifera and N. cadmea. – South African J. Botany. 120: 298–301. https://doi.org/10.1016/j.sajb.2018.09.008
Letchamo W., Korolyk E.A., Tkachev A.V. 2005. Chemical screening of essential oil bearing flora of Siberia IV. Composition of the essential oil of Nepeta sibirica L. Tops from Altai region. – J. Essential Oil Research. 17(5): 487–489. https://doi.org/10.1080/10412905.2005.9698973
Мяделец М.А., Домрачев Д.В., Водолазова С.В. 2012. Анализ химического состава эфирных масел Nepeta sibirica L., Thymus petraeus L. и Schizonepeta multifida L., произрастающих на территории Республики Хакасия. – Химия растительного сырья. 4: 119–124. https://elibrary.ru/item.asp?id=18834497
Sharma A., Shahzad B., Rehman A., Bhardwaj R., Landi M., Zheng B. 2019. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. – Molecules. 24(13): 2452. https://doi.org/10.3390/molecules24132452
Tepe B., Daferera D., Tepe A.-S., Polissiou M., Sokmen A. 2007. Antioxidant activity of the essential oil and various extracts of Nepeta flavida Hub.-Mor. from Turkey. – Food Chemistry. 103(4): 1358–1364, https://doi.org/10.1016/j.foodchem.2006.10.049
Крылов Г.В. 1992. Травы жизни и их искатели. Томск. 320 с.
Miceli N., Taviano M.F., Giuffrida D., Trovato A., Tzakou O., Galati E. 2005. Anti-inflammatory activity of extract and fractions from Nepeta sibthorpii Bentham. – J. Ethnopharmacology. 97(2): 261–266. https://doi.org/10.1016/j.jep.2004.11.024
Мяделец М.А., Водолазова С.В. 2006. Антимикробная активность сухих экстрактов и эфирных масел из надземной части видов сем. Lamiaceae L. – Вопросы общей ботаники: традиции и перспективы. Ч. 2. Казань. С. 74–76.
Işcan G., Köse Y., Demirci B., Başer K.H.C. 2011. Anticandidal activity of the essential oil of Nepeta transcaucasica Grossh. – Chemistry and biodiversity. 8(11): 2144–2148. https://doi.org/10.1002/cbdv.201100091
Köksal E., Tohma H., Kılıç Ö., Alan Y., Aras A., Gülçin İ., Bursal E. 2017. Assessment of antimicrobial and antioxidant activities of Nepeta trachonitica: analysis of its phenolic compounds using HPLC-MS/MS. – Sci. Pharm. 85(2): 24. https://doi.org/10.3390/scipharm85020024
Schultz G., Simbro E., Belden J., Zhu J., Coats J. 2004. Catnip, Nepeta cataria (Lamiales: Lamiaceae) – a closer look: seasonal occurrence of nepetalactone isomers and comparative repellency of three terpenoids to insects. – Environmental Entomology. 33(6): 1562–1569. https://doi.org/10.1603/0046-225X-33.6.1562
Zahirnia A., Boroomand M., Nasirian H., Salehzadeh A., Soleimani-Asl S. 2019. Comparing cytotoxicity of propoxur and Nepeta crispa (Lamiales: Lamiaceae) essential oil against invertebrate (Sf9) and vertebrate (L929) cell lines. – Vet. World. 12(11): 1698–1706. https://doi.org/10.14202/vetworld.2019.1698-1706
Budantsev A.L., Lobova T.A. 1997 Fruit morphology, anatomy and taxonomy of tribe Nepeteae (Labiatae) – Edinburgh J. Bot. 54(2): 183–216. https://doi.org/10.1017/S0960428600004029
Cheryomushkina V.A., Astashenkov A.Yu., Saidov D. 2016. Individual development of Nepeta bucharica Lipsk. (Lamiaceae) under different habitat conditions. – J. Biology and Nature. 6(1): 1–7. https://www.ikprress.org/index.php/JOBAN/article/view/1152
Асташенков А.Ю., Черемушкина В.А., Курочкина Н.Ю. 2019. Особенности онтогенеза и побегообразования эндемика Центральной Азии Nepeta formosa Kudr. (Lamiaceae). – Известия высших учебных заведений. Поволжский регион. Естественные науки. 3: 24–33. https://izvuz_est.pnzgu.ru/en3319
Пояркова А.И. 1954. Семейство Lamiaceae. – В кн. Флора СССР. М.; Л. Т. 20. С. 286–360.
Цукерваник Т.И. 1987. Род Nepeta. – В кн. Определитель растений Средней Азии. Т.9. Ташкент. С. 44–52.
Артюшенко З.М. Федоров А.А. 1986. Атлас по описательной морфологии высших растений. Плод. Л. 392 с.
Артюшенко З.М. Федоров А.А. 1989. Атлас по описательной морфологии высших растений. Семя. Л. 360 с.
Терехин Э.С. 1996. Семя и семенное размножение. СПб. 377 с.
Флоря В.Н. 1987. Интродукция и акклиматизация растений в Молдавии (лекарственные, витаминоносные, медоносные). Кишинев. 296 с.
Елисафенко Т.В. 2012. Изучение особенностей латентного периода растений на примере видов секции Mirabiles рода Viola (Violaceae). I. Семенная продуктивность и биология прорастания семян. – Раст. мир Азиатской России. 2(10): 66–72. https://elibrary.ru/item.asp?id=18263310
Лакин Г.Ф. 1973. Биометрия. М. 342 с.
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы