

Растительные ресурсы, 2021, T. 57, № 4, стр. 340-358
Воздействие загрязнения почв тяжелыми металлами на пространственное распределение биомассы напочвенного покрова и запаса лесной подстилки в сосновых лесах Кольского полуострова
И. В. Лянгузова 1, *, П. А. Примак 2, Ф. С. Салихова 1, Е. Н. Волкова 1, А. И. Беляева 1
1 Ботанический институт им. В.Л. Комарова РАН
г. Санкт-Петербург, Россия
2 Санкт-Петербургский государственный университет
г. Санкт-Петербург, Россия
* E-mail: ILyanguzova@binran.ru
Поступила в редакцию 13.03.2021
После доработки 14.07.2021
Принята к публикации 12.09.2021
Аннотация
Приведены результаты исследования внутриценотической неоднородности почвенно-растительного покрова в средневозрастных сосновых лесах лишайниково-зеленомошной группы типов леса на фоновой территории Кольского полуострова и в пределах буферной и импактной зон комбината “Североникель” (Мурманская обл.). Выявлены особенности в накоплении биомассы различных компонентов напочвенного покрова (лишайников, мхов, кустарничков), а также запасов растительного опада и лесной подстилки в зависимости от положения в тессере, под которой понимается совокупность приствольного, подкронового и межкронового пространств деревьев Pinus sylvestris L. или Betula pubescens Ehrh. Показано, что закономерности распределения биомассы компонентов напочвенного покрова и мортмассы в тессерах принципиально различаются в фоновых условиях и при аэротехногенном загрязнении. В условиях загрязнения распределение биомассы лишайников по микросайтам несколько выравнивается по сравнению с фоновыми сообществами, а контрастность распределения биомассы мхов, надземных органов кустарничков и массы опада существенно возрастает. Наблюдаемые явления обусловлены увеличением запаса тяжелых металлов в органогенном горизонте почв, неравномерностью его распределения по площади, а также снижением средообразующей роли деревьев.
Гетерогенность фитоценоза является одним из признаков его строения и необходимой формой осуществления оптимального оборота потоков вещества и энергии [1, 2]. В лесных сообществах деревья – эдификаторы фитоценоза, могут рассматриваться в качестве “экосистемных инженеров” [3], формирующих пространственную структуру и существенно преобразующих физико-химическую среду. В редкостойных северотаежных лесах гетерогенность и контрастность ценотических условий проявляется особенно ярко. Хорошо известно, что кроны деревьев перераспределяют атмосферные осадки и трансформируют их химический состав, влияют на световой, тепловой и ветровой режимы местообитаний, мохово-лишайниковый и травяно-кустарничковый ярусы, возобновление древесных растений [2, 4–10]. Микромозаика растительности в биогеоценозах определяет пространственную гетерогенность лесных почв, что наиболее четко отражается на морфологических, физико-химических и биологических свойствах их органогенных горизонтов, формирующихся из растительного опада [11–16]. В наших исследованиях, вслед за Л.О. Карпачевским [11, 13], мы придерживаемся термина “тессера”, под которым понимается совокупность приствольного, подкронового и межкронового пространств, различающихся локальными экологическими условиями; эта совокупность представлена разными типами микросайтов.
Исследования средообразующей роли деревьев в большинстве случаев осуществлялись на фоновых территориях, не испытывающих воздействия локальных источников загрязнения. Изучение микромозаики напочвенного покрова и верхних горизонтов почв в пределах лесных биогеоценозов преимущественно проводилось в северотаежных еловых лесах или в темнохвойных лесах средней тайги, где внутриценотическая мозаичность выражена наиболее контрастно [2, 4, 7, 17–21]. От приствольных пространств к межкроновым в напочвенном покрове увеличивается разнообразие экологических свит, общее число видов, долевое участие неморальных, светолюбивых и требовательных к общему содержанию элементов питания видов, уменьшаются толщина и запас лесных подстилок [22]. Значительно меньше работ посвящено изучению пространственной гетерогенности напочвенного покрова и верхних горизонтов почв в светлохвойных (сосновых, лиственничных) лесах, что, по-видимому, связано с тем, что они отличаются более равномерной освещенностью [5–7, 23–27].
Аэротехногенное загрязнение окружающей среды оказывает существенное воздействие на структуру и продуктивность фитоценозов, а также вносит дополнительный вклад в формирование пространственной неоднородности накопления и разложения органического вещества, способствует изменению фракционного состава, запасов и скорости разложения растительного опада и приводит к трансформации свойств лесных подстилок [14–16, 26, 28–32].
Цель настоящей работы – оценка пространственного распределения биомассы живого напочвенного покрова, запаса растительного опада, лесной подстилки и тяжелых металлов в средневозрастных северотаежных сосновых лесах, находящихся на фоновой территории и под воздействием аэротехногенного загрязнения выбросами комбината по производству цветных металлов.
Комбинат “Североникель” вступил в строй в 1938 и до 1968 гг. использовал руду местного происхождения, а затем перешел на руду Норильского месторождения, что привело к резкому увеличению объемов атмосферных выбросов диоксида серы. Согласно опубликованным данным [33], в период 1981–1990 гг. ежегодный объем атмосферных выбросов SO2 ОАО “Североникель” превышал в среднем 220 тыс. т, твердых веществ – 16 тыс. т, затем происходило постепенное снижение объемов выбросов, и к концу XX в. они составляли соответственно 45.8 и 6.0 тыс. т в год. В настоящее время ежегодный объем выбросов SO2 и твердых веществ на комбинате “Североникель” составляет соответственно 35–37 и 2.9–3.4 тыс. т в год.
МАТЕРИАЛ И МЕТОДЫ
Исследования проводили в средневозрастных сосновых лесах, расположенных в фоновом районе Кольского полуострова в среднем течении р. Ливы, и на территории буферной и импактной зон комбината “Североникель” (г. Мончегорск, Мурманская обл.). Давность последнего пожара в исследованных сообществах составляет в среднем 90 лет.
Пробные площади (ПП) размером 20 × 20 м были заложены в лишайниково-зеленомошных сосновых лесах в фоновом районе (ПП1) на расстоянии 80 км от комбината “Североникель” и на территории буферной (ПП2 и ПП3) и импактной (ПП4 и ПП5) зон. ПП2 и ПП4 удалены от комбината соответственно на расстояние 27 и 10 км в северо-восточном, а ПП3 и ПП5 – на 35 и 15 км в юго-западном направлении.
Краткая таксационная характеристика древостоев представлена в табл. 1. Древесный ярус исследованных сообществ сформирован Pinus sylvestris L. с участием Betula pubescens Ehrh. В травяно-кустарничковом ярусе фоновых сосновых лесов доминирующими видами являются кустарнички Vaccinium vitis-idaea L., V. myrtillus L., Empetrum hermaphroditum Hagerup, в мохово-лишайниковом – зеленые мхи Pleurozium schreberi (Brid.) Mitt., Dicranum sp. и лишайники Cladonia rangiferina (L.) Weber ex F.H.Wigg., Cl. stellaris (Opiz.) Pouzar & Vězda, Cl. arbuscula (Wallr.) Flot. На загрязненной территории в травяно-кустарничковом ярусе доминируют те же виды кустарничков, а в мохово-лишайниковом ярусе преобладают лишайники р. Cladonia с шиловидными и сцифовидными подециями. Согласно современной классификации почв России [34, 35], исследуемые почвы относятся к Al-Fe-гумусовым подзолам или к Albic Rustic Podzols, согласно классификации WRB [36].
Таблица 1.
Краткая таксационная характеристика древостоев на пробных площадях и их географическое расположение Table 1. Brief characteristics of the forest stands on sample plots and their location (SP)
| Номер ПП Number of sample plot |
Координаты, расстояние
и направление от комбината Location, distance and direction from the plant |
Вид, Species | Возраст, лет Age, years |
Диаметр на высоте
1.3 м, см Diameter at breast height, cm |
Высота, м Нeight, m |
Плотность, экз./га Density, ind./ha |
Сумма площадей сечений, м2/га Basal area, m2/ha |
|---|---|---|---|---|---|---|---|
| Фоновый район Background |
|||||||
| 1 | N 67°49′ E 31°20′ 80 км, ЮЗ 80 km, SW |
С/P | 70 | 11.9 | 11.5 | 1160 | 13 |
| Б/B | – | 7.9 | 7.7 | 480 | 2.3 | ||
| Буферная зона Buffer zone |
|||||||
| 2 | N 68°06′82″ E 33°19′45″ 35 км СВ 35 km, NE |
С/P | 80 | 9.1 | 9.7 | 2960 | 19 |
| Б/B | – | 2.8 | – | 290 | 0.2 | ||
| 3 | N 67°38′17″ E 32°42′23″ 35 км, ЮЮЗ 35 km, SSW |
С/P | 80 | 7.9 | 8.1 | 2880 | 14 |
| Б/B | – | 3.6 | – | 460 | 0.5 | ||
| Импактная зона Impact zone |
|||||||
| 4 | N 68°00′38″ E 32°55′54″ 10 км СВ 10 km, NE |
С/P | 80 | 6.6 | 9.2 | 4300 | 15 |
| Б/B | – | 3.9 | – | 700 | 0.9 | ||
| 5 | N 67°44′22″ E 32°46′45″ 15 км ЮЮЗ 15 km SSW |
С/P | 80 | 5.1 | 7.2 | 6300 | 13 |
| Б/B | – | 6 | – | 110 | 0.3 | ||
На каждой ПП были заложены трансекты, на которых с интервалом 1 м размещали учетные площадки размером 10 × 10 см. В зависимости от положения в тессере были выделены следующие типы микросайтов: А – в приствольной зоне в 20 см от стволов деревьев сосны и березы с 4-х сторон света; Б – в подкроновых пространствах деревьев сосны и березы; В – на межкроновых участках в “окнах” полога древостоя. В связи с практически полным отсутствием деревьев березы, на ПП2 исследования проводили только в сосновой тессере. Всего заложено 350 учетных площадок, обследовано 32 дерева сосны и 21 дерево березы.
На каждой площадке был вырезан монолит размером 10×10 см до верхнего минерального горизонта. С 4-х сторон монолита измерена толщина лесной подстилки. Монолит был разобран на живую биомассу и мортмассу. В живую биомассу входили следующие компоненты: надземные и подземные части сосудистых растений, зеленые части мхов и живые части лишайников. Мортмасса – слой отмерших растительных остатков, в котором, согласно современным представлениям [11–13, 37] выделяют 3 подгоризонта. Подгоризонт АОL, образованный свежим растительным опадом, все фракции которого (мелкие ветви, кора, хвоя, листья, шишки) еще сохраняют свою морфологическую структуру (этот слой мы будем в дальнейшем называть растительным опадом), и подгоризонты AOF и AOH, в совокупности образующие органогенный горизонт (О) почвы или лесную подстилку. Все образцы живой биомассы и мортмассы были высушены до воздушно-сухого состояния и взвешены. Запас живой биомассы и мортмассы представлен в г/м2.
Содержание кислоторастворимых форм Ni, Cu, Co было определено в вытяжке 1.0 н HCl из образцов лесной подстилки (соотношение 1 : 25) методом атомно-абсорбционной спектрофотометрии в 3-кратной повторности [38]. Как и другие сильные кислоты, 1.0 н HCl позволяет анализировать не только доступные растениям формы тяжелых металлов, но и потенциально мобилизуемые, что лучше отражает общий уровень их поступления в среду по сравнению с другими экстрагентами, такими, например, как ацетатно-аммонийный буфер или комплексоны [16, 39].
Для оценки уровня загрязнения почв тяжелыми металлами использовали два критерия: индекс техногенной нагрузки и запас Ni и Cu в лесной подстилке. Выбор органогенного горизонта подзолов обусловлен, с одной стороны, высокой сорбционной емкостью лесной подстилки, которая является основным депо тяжелых металлов в лесных экосистемах; с другой стороны, тем, что именно в этом горизонте располагаются подземные побеги и корни кустарничков. Индекс техногенной нагрузки (ИТН) представляет собой превышение суммарного содержания кислоторастворимых форм преобладающих металлов (Ni, Cu и Co) в подстилке над их фоновым содержанием [38]. Запас (г/м2) Ni и Cu в лесной подстилке был рассчитан как произведение концентрации металла (мг/кг) на запас подстилки (г/м2) на каждой учетной площадке.
Проверка выборок исследуемых параметров на соответствие закону нормального распределения показала, что распределения большинства параметров значимо отличаются от нормального распределения. В связи с этим при оценке значимости различий использовали непараметрические критерии Краскела–Уоллиса (H) и Манна–Уитни (z), при уровне значимости р < 0.05 различия считали достоверными.
РЕЗУЛЬТАТЫ
Биомасса компонентов напочвенного покрова. В фоновом сосняке (ПП1) величина надземной биомассы лишайников достоверно увеличивается от приствольных к межкроновым пространствам, как в сосновой, так и в березовой тессерах (рис. 1). В отличие от лишайников, биомасса мхов и надземных частей кустарничков в пределах березовой тессеры распределены равномерно (рис. 2, 3). В подкроновых микросайтах сосновой тессеры биомасса мхов является достоверно более низкой, а биомасса кустарничков, наоборот, более высокой по сравнению с другими типами микросайтов. В обеих тессерах не выявлено достоверных различий в величине надземной биомассы мхов и кустарничков в приствольных и межкроновых микросайтах.
Рис. 1.
Надземная биомасса лишайников в сосновой (а) и березовой (б) тессерах в исследуемых сосновых сообществах. Типы микросайтов: А – приствольные; B – подкроновые; C – межкроновые. По горизонтали – номер пробной площади (ПП); по вертикали – биомасса, г/м2. Fig. 1. Above-ground lichen biomass in pine (a) and birch (b) tessera of the studied Scots pine communities. Microsite types: A – at tree base, B – under crown, C – on canopy gaps. X-axis – number of sample plot; y-axis – above-ground lichen biomass stock, g/m2.
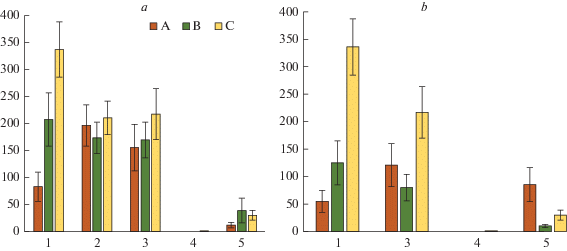
Рис. 2.
Биомасса мхов в сосновой (а) и березовой (б) тессерах в исследуемых сосновых сообществах. Типы микросайтов: А – приствольные; Б – подкроновые; C – межкроновые. По горизонтали – номер пробной площади (ПП); по вертикали – биомасса, г/м2. Fig. 2. Above-ground moss biomass in pine (a) and birch (b) tessera of the studied Scots pine communities. Microsite types: A – at tree base, B – under crown, C – on canopy gaps. X-axis – number of sample plot; y-axis – above-ground moss biomass stock, g/m2.
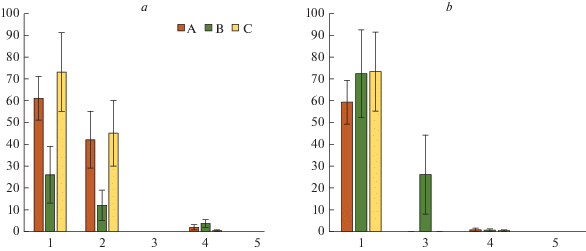
Рис. 3.
Надземная биомасса кустарничков в сосновой (а) и березовой (б) тессерах в исследуемых сосновых сообществах. Типы микросайтов: А – приствольные; Б – подкроновые; В – межкроновые. По горизонтали – номер пробной площади (ПП); по вертикали – биомасса, г/м2. Fig. 3. The above-ground biomass of dwarf shrubs in pine (a) and birch (b) tessera in the studied Scots pine communities. Microsite types: A – at tree base, B – under crown, C – on canopy gaps. X-axis – number of sample plot; y-axis – above-ground dwarf shrubs biomass stock, g/m2.
На территории буферной зоны воздействие аэротехногенного загрязнения на накопление органического вещества компонентами напочвенного покрова в сосновой и березовой тессерах проявляется по-разному (рис. 1–3). В обеих тессерах надземная биомасса лишайников вблизи стволов деревьев возрастает в 2–2.4 раза по отношению к фоновой величине, напротив, в межкроновых пространствах она достоверно снижена в 1.2–1.6 раз, а в подкроновых достоверно не отличается от фоновых значений (рис. 1). Мхи сохранились только на ПП2, благодаря почти вдвое меньшему уровню загрязнения почв, но их биомасса снижена в 1.5–2.2 раза по сравнению с фоновыми значениями; на ПП3 небольшая биомасса мхов отмечена лишь в подкроновом пространстве березы (рис. 2).
Величина надземной биомассы кустарничков по-разному связана с положением в тессере: на ПП2 в сосновой, а на ПП3 в березовой тессерах она достоверно меньше в межкроновых микросайтах по сравнению с приствольными и подкроновыми пространствами, в то время как в сосновой тессере на ПП3 этот показатель в среднем одинаков в разных микросайтах (рис. 3). Сравнительный анализ показал, что надземная биомасса кустарничков в отдельных типах микросайтов может достоверно не отличаться от соответствующих фоновых значений, быть меньше в 1.3–2.4 раза или даже в 1.4–2 раза больше.
В пределах импактной зоны для большинства компонентов напочвенного покрова зарегистрированы наименьшие величины надземной биомассы (рис. 1–3). На ПП4 практически полностью отсутствуют лишайники (рис. 1), зато встречаются ранне-сукцессионные виды мхов (Pohlia nutans, Polytrichum spp.), формирующие незначительную биомассу преимущественно в сосновой тессере (рис. 2). На ПП5, наоборот, полностью отсутствуют мхи, но присутствуют лишайники, в основном, ранне-сукцессионные виды р. Cladonia и Trapeliopsis granulosa. В сосновой тессере основная часть биомассы лишайников сосредоточена в подкроновом и межкроновом пространстве, а в березовой тессере, наоборот, преимущественно вблизи ствола (рис. 1, 2). Величина надземной биомассы кустарничков существенно варьирует в зависимости от положения в тессере (рис. 3). В сосновой тессере на обеих ПП импактной зоны биомасса многократно возрастает от приствольных к межкроновым микросайтам, а в березовой тессере максимальные ее значения отмечаются в подкроновых зонах. Вблизи стволов сосны и березы регистрируется резкое снижение надземной биомассы кустарничков по отношению к фоновым величинам, в межкроновых микросайтах ее величина достоверно не отличаются от фоновой. Под кронами сосен в импактной зоне надземная биомасса кустарничков снижается в 3.3–4.3 раза, а под кронами березы, напротив, возрастает в 1.6–5.7 раз по сравнению с фоновыми величинами.
Сравнительный анализ соотношения биомассы надземных и подземных частей кустарничков показал неоднозначность изменения этих показателей, как по градиенту аэротехногенного загрязнения, так и в зависимости от положения в тессере (рис. 4). В пределах буферной зоны соотношение массы надземных и подземных частей кустарничков в разных микросайтах обеих тессер существенно не отличается от такового в фоновом сосняке, при этом не выявляется закономерного изменения этого соотношения от приствольных к межкроновым микросайтам. В импактной зоне доля надземной биомассы кустарничков последовательно возрастает от 2.0–5.8% вблизи стволов деревьев сосны и березы до 30–36% в межкроновых пространствах, за исключением березовой тессеры на ПП5, где наиболее высокая доля надземной биомассы (56%) наблюдается в подкроновом пространстве (рис. 4). Следует отметить, что практически во всех случаях биомасса подземных частей кустарничков существенно (в 1.5–51 крат) превышает биомассу надземных частей. Кроме того, необходимо подчеркнуть, что интервал варьирования последнего показателя очень широк, от 2 до 56%.
Рис. 4.
Соотношение биомассы подземных (1) и надземных (2) частей кустарничков в сосновой (а) и березовой (б) тессерах в исследуемых сосновых сообществах. Типы микросайтов: А – приствольные; B – подкроновые; C – межкроновые. По горизонтали – тип микросайта; по вертикали – доля биомассы, %. Fig. 4. Proportion of the below- (1) and aboveground (2) dwarf shrubs biomass in pine (a) and birch (b) tessera of the studied Scots pine communities. Microsite types: A – at tree base, B – under crown, C – on canopy gaps. X-axis – microsite type; y-axis – biomass relative stock, %.

Для северотаежных лесов характерно преобладание накопления органического вещества в мохово-лишайниковом ярусе по сравнению с травяно-кустарничковым, что и наблюдается в исследуемом фоновом сосняке, где в обеих тессерах доля травяно-кустарничкового яруса плавно снижается от 35–37% вблизи стволов деревьев до 12% в межкроновых пространствах (рис. 5). На территории буферной зоны, как в сосновой, так и в березовой тессере сохраняется преобладание накопления органического вещества в мохово-лишайниковом ярусе по отношению к травяно-кустарничковому ярусу, при этом возрастание доли мохово-лишайникового яруса в общей биомассе от приствольных к межкроновым микросайтам выражено менее ярко и не столь последовательно по сравнению с фоновым сосняком, особенно в сосновой тессере (рис. 5). Вклад травяно-кустарничкового яруса в общую биомассу напочвенного покрова варьирует в пределах 16–50%.
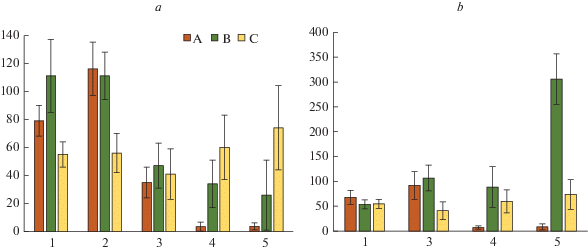
Рис. 5.
Соотношение надземной биомассы мохово-лишайникового (1) и травяно-кустарничкового (2) ярусов в разных типах микросайтов в сосновой (а) и березовой (б) тессерах в исследуемых сосновых сообществах. Типы микросайтов: А – приствольные; Б – подкроновые; В – межкроновые. По горизонтали – тип микросайта; по вертикали – доля биомассы, %. Fig. 5. Proportion of the lichen-moss (1) and dwarf shrub-herbaceous (2) layers biomass stock in pine (a) and birch (b) tessera of the studied Scots pine communities. Microsite types: A – at tree base, B – under crown, C – on canopy gaps. X-axis – microsite type; y-axis – biomass relative stock, %.

В пределах импактной зоны соотношение биомассы мохово-лишайникового и травяно-кустарничкового ярусов принципиально изменяется (рис. 5). Вклад мохово-лишайникового яруса в общую биомассу резко уменьшается вплоть до полного отсутствия, а вклад травяно-кустарничкового яруса, наоборот, существенно увеличивается, в некоторых микросайтах практически до 100%. При этом регистрируется уменьшение доли участия мохово-лишайникового яруса в общей биомассе от приствольных к межкроновым зонам в обеих тессерах. Иными словами, при сильном аэротехногенном загрязнении более высокая доля биомассы лишайников характерна для приствольных зон в отличие от противоположной закономерности в фоновых условиях.
Запас растительного опада в тессерах. В фоновом сосняке (ПП1) запас растительного опада в сосновой тессере снижается в 2 раза от приствольных к межкроновым участкам, в березовой тессере эта закономерность менее выражена, запас опада здесь снижается лишь в 1.5 раза (табл. 2).
Таблица 2.
Масса (г/м2) опада и подстилки в тессерах исследованных сосновых лесов Table 2. Stock (g/m2) of plant waste and forest litter in different microhabitats of pine forests
| Компонент Component |
Микросайты Microsites |
Критерий
Краскела–Уоллиса (H) Kruskal–Wallis H test |
Уровень
значимости (p) Significance level (p) |
||
|---|---|---|---|---|---|
| приствольные at tree base |
подкроновые under crowns |
межкроновые on canopy gaps |
|||
| ППП1, SP1 | |||||
| Масса опада Litterfall mass |
$\frac{{1700 \pm 102*}}{{1250 \pm 111}}$ | $\frac{{1215 \pm 103}}{{870 \pm 90}}$ | 835 ± 50 | $\frac{{26.0}}{{16.8}}$ | $\frac{{0.000}}{{{\mathbf{0}}.{\mathbf{002}}}}$ |
| Масса подстилки Litter layer mass |
$\frac{{3940 \pm 2873}}{{770 \pm 390}}$ | $\frac{{3240 \pm 4703}}{{350 \pm 437}}$ | 2880 ± 270 | $\frac{{12.3}}{{10.2}}$ | $\frac{{0.004}}{{{\mathbf{0}}.{\mathbf{006}}}}$ |
| Толщина подстилки, см Litter layer thickness, cm |
$\frac{{5.9 \pm 0.4}}{{4.7 \pm 0.4}}$ | $\frac{{3.8 \pm 0.4}}{{5.3 \pm 0.7}}$ | 2.7 ± 0.3 | $\frac{{16.8}}{{13.6}}$ | $\frac{{0.001}}{{{\mathbf{0}}.{\mathbf{002}}}}$ |
| ПП2, SP2 | |||||
| Масса опада Litterfall mass |
1655 ± 164 | 1480 ± 76 | 1465 ± 159 | 1.89 | 0.39 |
| Масса подстилки Litter layer mass |
5050 ± 654 | 4650 ± 413 | 4230 ± 441 | 0.69 | 0.71 |
| Толщина подстилки, см Litter layer thickness, cm |
5.4 ± 0.4 | 3.9 ± 0.3 | 3.1 ± 0.3 | 17.7 | 0.000 |
| ПП3, SP3 | |||||
| Масса опада Litterfall mass |
$\frac{{2440 \pm 2931}}{{515 \pm 364}}$ | $\frac{{1980 \pm 2662}}{{735 \pm 596}}$ | 702 ± 177 | $\frac{{14.7}}{{10.4}}$ | $\frac{{{\mathbf{0}}{\mathbf{.001}}}}{{{\mathbf{0}}.{\mathbf{005}}}}$ |
| Масса подстилки Litter layer mass |
$\frac{{5510 \pm 9164}}{{100 \pm 820}}$ | $\frac{{8950 \pm 2200}}{{6630 \pm 1245}}$ | 6420 ± 1560 | $\frac{{2.62}}{{2.64}}$ | $\frac{{0.270}}{{0.267}}$ |
| Толщина подстилки, см Litter layer thickness, cm |
$\frac{{3.7 \pm 0.4}}{{3.3 \pm 0.8}}$ | $\frac{{4.5 \pm 0.8}}{{5.1 \pm 0.8}}$ | 3.1 ± 0.5 | $\frac{{2.75}}{{3.56}}$ | $\frac{{0.25}}{{0.17}}$ |
| ПП4, SP4 | |||||
| Масса опада Litterfall mass |
$\frac{{2660 \pm 1661}}{{110 \pm 98}}$ | $\frac{{1900 \pm 130}}{{1260 \pm 170}}$ | 1040 ± 107 | $\frac{{29.0}}{{0.91}}$ | $\frac{{{\mathbf{0}}{\mathbf{.000}}}}{{0.63}}$ |
| Масса подстилки Litter layer mass |
4690 ± 437 4185 ± 657 |
3780 ± 349 5310 ± 777 |
5190 ± 993 | $\frac{{1.87}}{{1.28}}$ | $\frac{{0.39}}{{0.53}}$ |
| Толщина подстилки, см Litter layer thickness, cm |
$\frac{{3.8 \pm 0.5}}{{3.5 \pm 0.5}}$ | $\frac{{3.1 \pm 0.4}}{{4.7 \pm 0.5}}$ | 2.3 ± 0.4 | $\frac{{6.0}}{{10.0}}$ | $\frac{{{\mathbf{0}}{\mathbf{.049}}}}{{{\mathbf{0}}{\mathbf{.007}}}}$ |
| ПП5, SP5 | |||||
| Масса опада Litterfall mass |
$\frac{{1800 \pm 133}}{{1210 \pm 97}}$ | $\frac{{1250 \pm 131}}{{945 \pm 60}}$ | 920 ± 99 | $\frac{{19.5}}{{4.8}}$ | $\frac{{{\mathbf{0}}{\mathbf{.0001}}}}{{0.09}}$ |
| Масса подстилки Litter layer mass |
$\frac{{6875 \pm 475}}{{6885 \pm 362}}$ | $\frac{{6980 \pm 692}}{{7690 \pm 557}}$ | 6750 ± 478 | $\frac{{0.2}}{{1.9}}$ | $\frac{{0.90}}{{0.39}}$ |
| Толщина подстилки, см Litter layer thickness, cm |
$\frac{{5.8 \pm 0.4}}{{6.1 \pm 0.3}}$ | $\frac{{4.6 \pm 0.3}}{{6.4 \pm 0.5}}$ | 3.3 ± 0.4 | $\frac{{21.6}}{{23.4}}$ | $\frac{{0.000}}{{{\mathbf{0}}.{\mathbf{000}}}}$ |
На территории буферной зоны характер изменения запасов растительного опада различается, как на ПП2 и ПП3, так и в тессерах (табл. 2). На ПП2 запас растительного опада не связан с положением в тессере. На ПП3 запас растительного опада в 2.2–3.5 раза снижается от приствольных к межкроновым микросайтам в обеих тессерах, при этом вблизи стволов сосны накапливается в 1.6 раза больше опада по сравнению с приствольными микросайтами в березовой тессере, а в подкроновых пространствах сравниваемых тессер достоверные различия отсутствуют.
На обеих ПП импактной зоны в сосновой тессере запас растительного опада в 2–2.6 раза снижается от приствольных к межкроновым зонам (табл. 2). В березовых тессерах на обеих ПП отсутствует закономерное изменение запаса растительного опада в зависимости от положения в тессере.
Толщина и запас лесной подстилки в тессерах. В фоновом районе (ПП1) толщина и запас лесной подстилки, как в сосновой, так и в березовой тессере в приствольных и подкроновых зонах достоверно ниже, чем в межкроновом пространстве (табл. 2).
В буферной зоне на ПП2 толщина лесной подстилки в 1.7 раза снижается от приствольных к межкроновым микросайтам, в то время как на ПП3 различия в толщине подстилки недостоверны в обеих тессерах (табл. 2). Запас подстилки не связан ни с положением в тессере, ни с видовой принадлежностью дерева, однако в большинстве случаев значения этого показателя достоверно (в 1.1–2.8 раза) больше его фоновых величин.
В импактной зоне на ПП4 и ПП5 характер изменения толщины подстилки как в сосновой, так и в березовой тессерах сходен: ее величина достоверно снижается от приствольных и подкроновых к межкроновым микросайтам (табл. 2). Запас подстилки на обеих ПП не связан ни с положением в тессере, ни с видовой принадлежностью дерева-эдификатора, но он в среднем в 1.7 раз больше по сравнению с его величиной в фоновом районе (лишь в отдельных случаях на ПП 4 различия с фоновыми показателями в сравниваемых типах микросайтов могут отсутствовать).
Вариабельность индекса техногенной нагрузки и запасов тяжелых металлов в тессерах. В фоновом сосновом лесу (ПП1) среднее значение суммарной концентрации кислоторастворимых форм Ni, Cu и Со в органогенном горизонте (лесной подстилке) Al-Fe-гумусовых подзолов составляет 20.0 ± 0.5 мг/кг, при этом не выявлено достоверных различий в их содержании в разных типах микросайтов, поэтому при расчете индекса техногенной нагрузки суммарное содержание этих металлов было принято за 1.
В пределах буферной зоны, как в сосновой, так и в березовой тессере индекс техногенной нагрузки достоверно снижается от приствольных к межкроновым пространствам (Н = 6.4–26.9, р = = 0.000–0.04). Однако величины индексов техногенной нагрузки на ПП2 и ПП3 существенно различаются. На ПП2 его среднее значение равно 8.7 ± 0.3 отн. ед., составляя в приствольных, подкроновых и межкроновых микросайтах соответственно 9.6 ± 0.3, 9.3 ± 0.4, 7.2 ± 0.3 отн. ед. На ПП3 среднее значение (15.8 ± 1.1 отн. ед.) является более высоким; в соответствующих типах микросайтов в сосновой тессере величины индекса составляют 17.0 ± 1.3, 17.7 ± 1.0, 13.1 ± 1.3 отн. ед., в березовой тессере –18.5 ± 1.3, 12.2 ± 1.1, 13.4 ± ± 3.3 отн. ед., при этом различия между сосновой и березовой тессерами недостоверны. Столь существенные различия в значениях индекса техногенной нагрузки на двух исследованных пробных площадях в буферной зоне обусловлены преобладанием ветров, имеющих юго-юго-западное направление от г. Мончегорска, и, соответственно, большим количеством полиметаллической пыли, переносимой именно в этом направлении.
На территории импактной зоны средние значения индекса техногенной нагрузки одинаковы на ПП4 и ПП5 – 116 ± 5 и 119 ± 7 отн. ед., и в 7.5–13.6 раз больше соответствующих величин в буферной зоне. Диапазоны варьирования этого показателя достаточно широки – 11–182 и 66–218 отн. ед. соответственно. На обеих ПП в разных типах микросайтов величины индекса техногенной нагрузки достоверно не различаются, но на ПП4 индекс техногенной нагрузки в сосновой тессере достоверно больше по отношению к его значениям в березовой тессере (z = 3.61, p = 0.000).
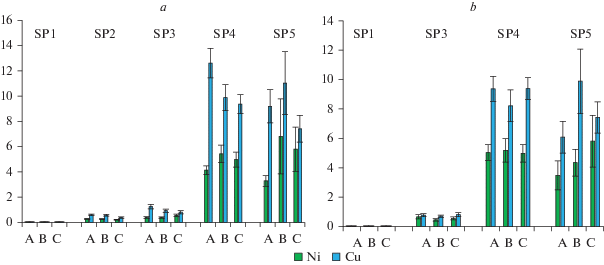
Распределение запасов тяжелых металлов в лесной подстилке по площади фитоценоза имеет свои особенности, обусловленные как химической природой металла, так и биогеоценотической обстановкой, и отличается от распределения концентраций тяжелых металлов. Прежде всего следует отметить, что наименьшие значения запаса регистрируются для кобальта, они варьируют от 0.004 (в подстилке фонового сосняка) до 0.24 г/м2 (в импактной зоне), поэтому в дальнейшем мы их не будем учитывать. В зонах аэротехногенного загрязнения запасы тяжелых металлов в подстилке различаются значительно сильнее, чем концентрации: так например, в сосновых тессерах превышение фоновых величин в запасах Ni и Cu достигают в буферной зоне 9–33, в импактной – 170–390 крат; в березовых тессерах 18–28 и 140–325 крат соответственно.
Характер изменения запасов тяжелых металлов в тессерах неодинаков, как в зависимости от тессеры, так и от металла (рис. 6). В условиях аэротехногенного загрязнения во всех случаях более высокие величины запаса фиксируются для меди, в частности, в импактной зоне они превышают соответствующие значения запасов никеля в среднем в 2 раза (рис. 6). Независимо от уровня аэротехногенной нагрузки запасы тяжелых металлов как в сосновой, так и в березовой тессерах значимо не различаются в разных типах микросайтов (H = 0.19–4.87, p > 0.05). В березовой тессере на ПП3 и ПП4 запас меди является достоверно меньшим (z = 2.11–2.29, p = 0.02–0.03) по сравнению с сосновой. Столь неоднозначный характер изменения запасов тяжелых металлов, скорее всего, связан с отсутствием достоверных различий в запасах лесной подстилки в разных типах микросайтов независимо от уровня аэротехногенного загрязнения (табл. 2), а концентрации кислоторастворимых форм меди в подстилке всегда больше соответствующего содержания никеля.
Рис. 6.
Запасы Ni и Cu в лесной подстилке в сосновой (а) и березовой (б) тессерах в исследуемых сосновых сообществах. Типы микросайтов: А – приствольные; Б – подкроновые; В – межкроновые. По горизонтали – тип микросайта; по вертикали – запас металла, г/м2. Fig. 6. Stock of Ni and Co in litter layer of pine (a) and birch (b) tessera of the studied Scots pine communities. Microsite types: A – at tree base, B – under crown, C – on canopy gaps. X-axis – microsite type; y-axis – stock of metal, g/m2.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Результаты многолетнего мониторинга уровня загрязнения верхнего органогенного горизонта Al-Fe-гумусовых подзолов показывают, что несмотря на 5–8-кратное снижение объемов атмосферных выбросов комбинатом “Североникель” на территории буферной зоны уровень загрязнения лесной подстилки продолжает увеличиваться, а на территории импактной зоны практически не снижается [40, 41, 44–46]. В пределах буферной и импактной зон концентрации кислоторастворимых форм тяжелых металлов в подстилке в среднем в 5–25 и 80–190 раз соответственно превышают региональные фоновые значения. Еще более впечатляющие различия наблюдаются в запасах тяжелых металлов в органогенном горизонте почв. В буферной зоне превышение фоновых значений достигает для Ni – 9–18, для Cu – 18–33 крат, а в импактной зоне соответственно 140–175 и 290–390 крат, что обусловлено меньшей скоростью разложения растительных остатков и соответственно повышенным накоплением массы подстилки [14–16, 26, 42, 43]. Сохранение высокого уровня загрязнения почв тяжелыми металлами препятствует восстановлению биоты, что хорошо задокументировано в районах воздействия атмосферных выбросов предприятий цветной металлургии [44–49].
В фоновых условиях наиболее четкие закономерности в накоплении органического вещества в зависимости от положения в тессере наблюдаются для надземной биомассы лишайников, для мхов и кустарничков более характерно незакономерное варьирование биомассы по площади фитоценоза (рис. 1–3), что, возможно, связано с микромасштабным пространственным варьированием условий экотопа. В разных типах еловых лесов выявлена высокая степень варьирования запаса надземной биомассы напочвенного покрова в пределах тессер [22, 50]. Авторы установили, что внутрибиогеоценозное варьирование надземной биомассы напочвенного покрова превышает межбиогеоценозное варьирование, при этом фактором, определяющим увеличение биомассы напочвенного покрова от приствольных к межкроновым пространствам, является освещенность. Исследования ценопопуляций Vaccinium myrtillus в южно-таежных ельниках и среднетаежных сосняках показали, что внутриценозный оптимум для большинства показателей черники отмечается на наноповышениях и в основных микрогруппировках, приуроченных, как правило, к оптимальным по освещенности участкам, при этом худшие абсолютные значения большинства показателей регистрируются в сосняках на приствольных повышениях [51]. В редкостойных северотаежных лесах фактор освещенности не играет столь существенной роли, как в еловых лесах южной тайги, однако, как в еловых, так и в сосновых фоновых лесах Кольского полуострова влияние деревьев на формирование биомассы напочвенного покрова остается по-прежнему значительным [25]. Авторы указывают, что в сосняках кустарничково-лишайниковых наименьший запас надземной биомассы напочвенного покрова регистрируется в приствольных зонах (758 г/м2), а максимальный – в подкроновых пространствах (1414–1482 г/м2) деревьев сосны, при этом запас биомассы в межкроновых (730 г/м2) и приствольных зонах достоверно не различается.
Как показали наши предшествующие исследования [27], максимальные величины запаса надземной биомассы напочвенного покрова отмечаются в наиболее сухом лишайниковом сосновом редколесье, а минимальные – в более влажном сосняке зеленомошном, при этом наиболее контрастно запасы биомассы нижних ярусов распределены по площади фитоценоза в сосновом редколесье, а наиболее выровнены – в сосняке зеленомошном. Рассматриваемые нами сообщества сосновых лесов находятся лишь на промежуточной стадии постпирогенного восстановления (давность последнего пожара составляет 90 лет), когда общее проективное покрытие и высота травяно-кустарничкового и мохово-лишайникового ярусов еще не достигли стабилизации, регистрируемой при давности пожара свыше 100 лет [28]. Именно это обстоятельство объясняет несколько сниженные величины запасов компонентов напочвенного покрова в исследуемых сосновых лесах по сравнению с данными, приводимыми В.В. Никоновым с соавт. [25].
Воздействие аэротехногенного загрязнения диоксидом серы совместно с полиметаллической пылью привело к изменению видового состава и структуры мохово-лишайникового яруса в буферной зоне (вплоть до полного выпадения доминанта мохового покрова Pleurozium schreberi на ПП3) и смене кустистых лишайников, доминирующих в фоновом сосняке, лишайниками с шиловидными и сцифовидными подециями. В результате нарушения видовой структуры и проективного покрытия мохово-лишайникового яруса под воздействием аэротехногенного загрязнения на территории буферной зоны создаются более контрастные гидротермические условия, что было отмечено нами ранее [52]. Сочетанное воздействие этих факторов привело к 1.5–2-кратным различиям в величине биомассы этого яруса на ПП2 и ПП3 по сравнению с фоновым сообществом и способствовало замедлению скорости разложения растительных остатков и накоплению мортмассы опада, которая в 1.2–1.5 раза превышает соответствующую величину в фоновом сосняке.
На территории импактной зоны ранне-сукцессионные виды мхов (Pohlia nutans, Polytrichum spp.) формируют незначительную биомассу преимущественно в сосновой тессере, а лишайники р. Cladonia с шиловидными и сцифовидными подециями накапливают небольшой по величине запас биомассы в обеих типах тессер. В условиях аэротехногенного загрязнения наиболее устойчивым является травяно-кустарничковый ярус [28, 53], что и обусловливает более значительное накопление биомассы кустарничков по отношению к биомассе компонентов мохово-лишайникового яруса (рис. 1–3). Как следствие различной устойчивости нижних ярусов северотаежных лесов, изменяется структура биомассы напочвенного покрова (рис. 5). Если в фоновом районе и буферной зоне основной вклад в общую биомассу вносят мхи и лишайники, то в импактной зоне значительно возрастает доля участия кустарничков.
Логично предположить, что распределение по площади фитоценоза толщины и запаса лесной подстилки должно быть связано с запасом растительного опада, однако, как оказалось, согласованное изменение указанных параметров регистрируется не всегда (табл. 2). Если толщина подстилки и масса опада в большинстве случаев закономерно снижаются от приствольных к межкроновым зонам, то масса подстилки в условиях аэротехногенного загрязнения не связана с положением в тессере. Возможно, это обусловлено высокой степенью вариабельности параметров лесных подстилок, что отмечается в ряде работ [22, 50, 54].
Сравнительный анализ различий в накоплении органического вещества компонентами сообществ лишайниково-зеленомошных сосновых лесов в разных типах микросайтов позволяет сделать ряд заключений, касающихся изменения средообразующей роли древесного яруса в условиях аэротехногенного загрязнения. Если принять за 1 величины исследованных параметров на межкроновых участках, то оказывается, что хорошо выраженное в обеих тессерах фонового сосняка последовательное возрастание биомассы лишайников от приствольных к межкроновым зонам существенно сглаживается в сосновой тессере буферной зоны за счет исчезновения различий между приствольными и межкроновыми участками, и в меньшей мере в березовой тессере (табл. 3).
Таблица 3.
Соотношение биомассы компонентов напочвенного покрова, мортмассы опада и подстилки и толщины подстилки в разных типах микросайтов исследованных сосновых лесов Table 3. The ratio of the biomass of ground layer components, mortmass of plant waste and forest litter and the thickness of forest litter in different microsites of Scots pine forests
| Компонент Component |
ПП1 SP1 |
ПП2 SP2 |
ПП3 SP3 |
ПП4 SP4 |
ПП5 SP5 |
|---|---|---|---|---|---|
| Лишайники Lichens |
$\frac{{0.25:0.6:{{1}^{1}}}}{{0.16:0.4:1}}$ | 0.9 : 0.8 : 1 | $\frac{{0.7:0.8:1}}{{0.6:0.4:1}}$ | $\frac{0}{0}$ | $\frac{{0.4:1.3:1}}{{2.8:0.3:1}}$ |
| Мхи Mosses |
$\frac{{0.8:0.4:1}}{{0.8:1:1}}$ | 0.9 : 0.3 : 1 | – | $\frac{{5:9:1}}{{2:1.5:1}}$ | |
| Кустарнички: Dwarf-shrubs |
|||||
| надземные части aboveground parts |
1.4 : 2 : 1 1.2 : 1 : 1 |
2.1 : 2 : 1 | $\frac{{0.9:1.1:1}}{{2.2:{\text{ }}2.6:1}}$ | $\frac{{0.1:0.6:1}}{{0.1:1.5:1{\text{ }}}}$ | $\frac{{0.1:0.4:1}}{{0.1:4.1:1}}$ |
| подземные части belowground parts |
$\frac{{1.2:1.3:1}}{{1.7:2.6:1}}$ | 1.3 : 1.3 : 1 | $\frac{{0.3:0.4:1}}{{4.5:5.4:1}}$ | $\frac{{1:1.2:1}}{{1.1:2.9:1}}$ | $\frac{{1.1:1.1:1}}{{0.5:1.4:1}}$ |
| Масса опада Litterfall mass |
$\frac{{2.0:1.5:1}}{{1.5:1:1}}$ | 1.1 : 1 : 1 | $\frac{{3.5:2.8:1}}{{2.2:3.9:1}}$ | $\frac{{2.6:1.8:1}}{{1.4:1.2:1}}$ | $\frac{{2.0:1.4:1}}{{1.3:1.0:1}}$ |
| Масса подстилки Litter layer mass |
$\frac{{1.4:1.1:1}}{{1.3:1.2:1}}$ | 1.2 : 1.1 : 1 | $\frac{{0.9:1.4:1}}{{0.6:1.0:1}}$ | $\frac{{0.9:0.7:1}}{{0.8:1.0:1}}$ | $\frac{{1.0:1.0:1}}{{1.0:1.1:1}}$ |
| Толщина подстилки, см Litter layer thickness, cm |
$\frac{{2.2:1.4:1}}{{1.7:2.0:1}}$ | 1.7 : 1.3 : 1 | $\frac{{1.2:1.5:1}}{{1.0:1.6:1}}$ | $\frac{{1.7:1.3:1}}{{1.1:2.0:1}}$ | $\frac{{1.8:1.4:1}}{{1.8:1.9:1}}$ |
В импактной зоне соотношение величин биомассы лишайников в разных микросайтах различается в зависимости от видовой принадлежности дерева. В сосновой тессере направленность изменения этого параметра от приствольных участков к межкроновым аналогична наблюдаемой в фоновом сообществе, а в березовой тессере изменяется на обратную. Как в сосновой, так и в березовой тессере контрастность различий микросайтов несколько снижается по сравнению с фоновой зоной.
Соотношение величин биомассы мхов в приствольном, подкроновом и межкроновом пространстве сосновой тессеры, наблюдаемое в фоновой и буферной зонах, в импактной зоне изменяется на обратное (табл. 3). Кроме того, в импактной зоне существенно возрастает контрастность распределения биомассы мхов в разных микросайтах по сравнению с фоновыми условиями. Наиболее ярко это выражено в сосновой тессере, где мхи в преимущественно приурочены к приствольным и подкроновым участкам.
Различия в величинах надземной биомассы кустарничков в разных частях сосновой и березовой тессер в фоновой и буферной зонах в большинстве случаев не превышают 2-х крат. В отдельных случаях, в частности в березовой тессере фонового сосняка и в сосновой тессере на ПП3 в буферной зоне, надземная биомасса кустарничков практически равномерно распределена по разным микросайтам (табл. 3). Наиболее контрастные различия регистрируются в импактной зоне, где величина надземной биомассы кустарничков возрастает в 10 раз от участков вблизи стволов сосны или березы к межкроновым пространствам.
Биомасса подземных частей растений в сосновых тессерах, как в фоновом сообществе, так и в условиях аэротехногенного загрязнения распределена практически равномерно, за исключением ПП3, где ее величина существенно выше на межкроновых участках по сравнению с приствольными и подкроновыми (табл. 3). В березовых тессерах подземные части растений, в основном, сконцентрированы в подкроновых зонах независимо от уровня аэротехногенного загрязнения.
Контрастность в распределении запаса растительного опада по типам микросайтов наиболее ярко выражена в сосновых тессерах, где в приствольных зонах его величина во всех исследованных сообществах, за исключением ПП2, в 2–3.5 раза больше, чем в межкроновых пространствах (табл. 3). В березовых тессерах контрастность в распределении массы опада выражена очень слабо, за исключением ПП5 в импактной зоне. В отличие от опада, масса лесной подстилки относительно равномерно распределена в пределах сосновой и березовой тессер, как в фоновых условиях, так и в условиях загрязнения (табл. 3). Контрастность пространственного распределения толщины подстилки выражена более четко. Это относится как к сосновым, так и к березовым тессерам: вблизи стволов и в подкроновом пространстве деревьев толщина подстилки в большинстве случаев 1.5–2 раза больше, чем в межкроновых микросайтах (табл. 3).
Определенные трудности в оценке влияния дерева-эдификатора на формирование микромозаики напочвенного покрова в северотаежных сосновых лесах обусловлены рядом причин. Во-первых, особенностями структуры крон сосны обыкновенной, их более низкой плотностью, ажурностью по сравнению с кронами ели. Во-вторых, более быстрой скоростью разложения лиственного опада березы пушистой по сравнению с опадом хвои сосны обыкновенной, что приводит к большему накоплению мортмассы в приствольных зонах сосны по отношению к межкроновым пространствам [55]. В-третьих, высокой степенью неоднородности уровня загрязнения лесной подстилки тяжелыми металлами, связанной в том числе с деятельностью микромицетов [14, 16, 52, 56]. В результате формируются микролокусы или “микросайты выживания” с меньшим уровнем токсичности почвы, где могут сохраняться и выживать растения, что приводит к тому, что состояние экосистем в зоне экстремального загрязнения варьирует в широком диапазоне и не всегда согласуется с уровнем концентраций основных металлов-загрязнителей в почвах [45, 46]. В-четвертых, различиями в видовом составе микроорганизмов, обладающих разными жизненными стратегиями [57]. В-пятых, сглаживанием различий в химическом составе атмосферных осадков, выпадающих в межкроновых и подкроновых пространствах в условиях сильного аэротехногенного загрязнения, где охвоенность крон существенно снижена [8, 10].
ЗАКЛЮЧЕНИЕ
Исследование, проведенное в средневозрастных сосновых лесах Кольского полуострова, позволило оценить степень внутриценотической неоднородности распределения биомассы компонентов напочвенного покрова, мортмассы опада и лесной подстилки в фоновых условиях и по градиенту аэротехногенного загрязнения в зонах воздействия выбросов медно-никелевого комбината.
Характер распределения биомассы компонентов напочвенного покрова и мортмассы в тессерах принципиально различается в фоновых условиях и при аэротехногенном загрязнении. В сообществах фоновых сосновых лесов только биомасса лишайников возрастает от приствольных к межкроновым пространствам, как в сосновой, так и в березовой тессерах. Величина надземной биомассы мхов и кустарничков не связана с положением в тессере, в то время как запас мортмассы убывает от приствольных к межкроновым микросайтам.
В условиях аэротехногенного загрязнения распределение биомассы компонентов напочвенного покрова и мортмассы по микросайтам изменяется по-разному. Распределение биомассы лишайников несколько выравнивается, напротив, контрастность распределения биомассы мхов и надземных органов кустарничков существенно возрастает (до 5–10 крат, против 1.5–2 крат в фоновых условиях). Заметно (с 1.5–2 крат до 3–4 крат) возрастает контрастность распределения массы опада, тогда как распределение массы подстилки существенно не меняется по сравнению с фоновыми условиями. Логично предположить, что причины этих явлений состоят в существенном изменении уровня загрязнения верхнего органогенного горизонта почв по мере приближения к источнику загрязнения и неравномерности распределения тяжелых металлов по площади сообществ, а также в изменении состояния главного эдификатора сообществ – соснового древостоя.
В условиях загрязнения кроны деревьев сосны приобретают еще бóльшую ажурность, снижается охвоенность ветвей, перераспределение количества и трансформация химического состава осадков кронами сосны существенно ослабляется, что приводит к снижению роли хвойных деревьев в распределении органического вещества по площади фитоценоза. При совместном произрастании деревьев Pinus sylvestris L. и Betula pubescens Ehrh. более четко различия запасов био- и мортмассы компонентов почвенно-растительного покрова в разных микросайтах выражены в сосновых тессерах по сравнению с березовыми.
В зонах аэротехногенного загрязнения наблюдается не только увеличение общего запаса тяжелых металлов в органогенном горизонте почв, но и усиление контрастности его распределения по площади. Степень контрастности слабо повышается (~ на 20%) по сравнению с фоновыми условиями в буферной зоне и очень резко проявляется в импактной (в 3–16 раз), и если в первом случае причиной является перераспределение загрязнителей кронами деревьев, то во втором влияние древесного яруса уже не диагностируется. При современных объемах атмосферных выбросов комбината “Североникель”, сниженных в 5–8 раз по отношению к их максимальным величинам, запасы тяжелых металлов в органогенном горизонте Al-Fe-гумусовых подзолов на территории импактной зоны остаются очень высокими, что препятствует даже началу процессов восстановления напочвенного покрова сообществ сосновых лесов.
Список литературы
Ипатов В.С. 2013. Вновь о понятии “фитоценоз” – Бот. журн. 98(4): 481–486. https://doi.org/10.1134/S1234567813040046
Лебедева В.Х., Ипатов В.С., Тиходеева М.Ю. 2015. Неоднородность пространственной структуры живого напочвенного покрова в лесных сообществах. – Вестник СПбГУ. Сер. 3. Биология. 2: 32–46. https://biocomm.spbu.ru/article/view/859/750
Jones C.G., Lawton J.H., Shachak M. 1994. Organisms as ecosystem engineers. – Oikos. 69(3): 373–386. https://doi.org/10.2307/3545850
Лебедева В.Х., Тиходеева М.Ю., Ипатов В.С. 2005. Влияние древесного полога на виды напочвенного покрова в ельнике чернично-зеленомошном. – Бот. журн. 90(3): 400–410.
Лебедева В.Х., Тиходеева М.Ю., Ипатов В.С. 2006. Оценка влияния деревьев на виды травяно-кустарничкового и мохового ярусов в сосняке чернично-зеленомошном. – Бот. журн. 91(2): 176–192.
Лебедева В.Х., Тиходеева М.Ю., Ипатов В.С. 2016. О неоднородности растительного покрова лугов и лесов. – Бот. журн. 101(4): 358–376. https://doi.org/10.1134/S0006813616040025
Лукина Н.В., Ершов В.В., Горбачева Т.В., Орлова М.А., Исаева Л.Г., Тебенькова Д.Н. 2018. Оценка состава почвенных вод северотаежных хвойных лесов фоновых территорий индустриально развитого региона. – Почвоведение. 3: 284–296. https://doi.org/10.7868/S0032180X18030036
Ershov V.V., Isaeva L.G., Sukhareva T.A., Lukina N.V., Danilova M.A., Smirnov V.E. 2020. Assessment of the composition of rain deposition in coniferous forests at the Northern tree line subject to air pollution. – Russian J. Ecology. 51(4): 319–328. https://doi.org/10.1134/S1067413620040050
Клименко Д.Е., Остахова А.Л. 2020. Моделирование потерь дождевых осадков в кронах хвойных древостоев. – Лесоведение. 4: 335–345. https://doi.org/10.31857/S0024114820040063
Ershov V.V., Lukina N.V., Orlova M.A., Zukert N.V. 2016. Dynamics of snowmelt water composition in conifer forests exposed to airborne industrial pollution. – Rus. J. Ecology. 47(1): 46–52. https://doi.org/10.1134/S1067413616010045
Карпачевский Л.О. 1981. Лес и лесные почвы. М. 264 с.
Карпачевский Л.О. 1983. Подстилка – особый биогоризонт лесного биогеоценоза. – Роль подстилки в лесных биогеоценозах. М. 88–89.
Карпачевский Л.О., Зубкова Т.А., Ташнинова Л.Н., Руденко Р.Н. 2007. Почвенный покров и парцеллярная структура лесного биогеоценоза. – Лесоведение. 6: 107–113.
Воробейчик Е.Л., Пищулин П.Г. 2009. Влияние отдельных деревьев на рН и содержание тяжелых металлов в лесной подстилке в условиях промышленного загрязнения. – Почвоведение. 8: 927–937.
Воробейчик Е.Л., Пищулин П.Г. 2011. Влияние деревьев на скорость деструкции целлюлозы в почвах в условиях промышленного загрязнения. – Почвоведение. 5: 597–610.
Воробейчик Е.Л., Пищулин П.Г. 2016. Промышленное загрязнение снижает роль деревьев в формировании структуры полей концентраций тяжелых металлов в лесной подстилке. – Экология. 5: 323–334. https://doi.org/10.7868/S0367059716050164
Орлова М.А., Лукина Н. В., Камаев И. О., Смирнов В. Э., Кравченко Т.В. 2011. Мозаичность лесных биогеоценозов и продуктивность почв. – Лесоведение. 6: 39–48.
Орлова М.А., Лукина Н.В., Смирнов В.Э., Артемкина Н.А. 2016. Влияние ели на кислотность и содержании элементов питания в почвах северотаежных ельников кустарничково-зеленомошных. – Почвоведение. 11: 1355–1367. https://doi.org/10.7868/S0032180X16110071
Смирнова О.В., Алейников А.А., Семиколенных А.А., Бовкунов А.Д., Запрудина М.В., Смирнов Н.С. 2011. Пространственная неоднородность почвенно-растительного покрова темнохвойных лесов в Печоро-Илычском заповеднике. – Лесоведение. 6:67–78.
Луговая Д.Л., Смирнова О.В., Запрудина М.В., Алейников А.А., Смирнов В.Э. 2013. Микромозаичная организация и фитомасса напочвенного покрова в основных типах темнохвойных лесов Печоро-Илычского заповедника. – Экология. 1: 3–10. https://doi.org/10.7868/S0367059713010083
Артемкина Н.А., Орлова М.А., Лукина Н.В. 2018. Микромозаика растительности и вариабельность химического состава L-горизонтов подстилки северотаежных ельников кустарничково-зеленомошных. – Лесоведение. 2: 97–106. https://doi.org/10.7868/S002411481802002X
Семенюк О.В., Телеснина В.М., Богатырев Л.Г., Бенедиктова А.И., Кузнецова Я.Д. 2020. Оценка внутрибиогеоценозной изменчивости лесных подстилок и травяно-кустарничковой растительности в еловых насаждениях. – Почвоведение. 1: 31–43. https://doi.org/10.31857/S0032180X2001013X
Демьянов В. А. 1982. Влияние Larix gmelinii (Pinaceae) на строение растительного покрова на верхней границе древесной растительности. – Бот. журн. 62(4): 500–507.
Демьянов В.А. 1992. Ценотическая роль Pinus sylvestris L. в лесных сообществах Кольского севера в условиях техногенного загрязнения. – Изв. РАН. Сер. биол. 1: 52–57.
Никонов В.В., Лукина Н.В., Смирнова Е.В., Исаева Л.Г. 2002. Влияние Picea obovata и Pinus sylvestris на первичную продуктивность нижних ярусов хвойных лесов Кольского полуострова. – Ботан. журн. 87(8):107–119.
Иванова Е.А., Лукина Н.В., Данилова М.А., Артемкина Н.А., Смирнов В.Э., Ершов В.В., Исаева Л.Г. 2019. Влияние аэротехногенного загрязнения на скорость разложения растительных остатков в сосновых лесах на северном пределе распространения. – Лесоведение. 6: 533–546. https://doi.org/10.1134/S0024114819060044
Лянгузова И.В., Примак П.А. 2019. Пространственное распределение запасов напочвенного покрова и лесной подстилки в средневозрастных сосновых лесах Кольского полуострова. – Растительные ресурсы. 4: 473–489. https://doi.org/10.1134/S003399461904006X
Динамика лесных сообществ Северо-Запада России. 2009. СПб. 276 с.
Иванова Е.А., Лукина Н.В. 2017. Варьирование массы и фракционного состава древесного опада в сосняках кустарничково-лишайниковых при аэротехногенном загрязнении. – Лесоведение. 5: 47–58. https://doi.org/10.7868/S0024114817050059
Одинцов П.Е., Караванова Е.И., Степанова А.А. 2018. Трансформация водорастворимых органических веществ подстилок подзолов фоновых и техногенных территорий Кольского полуострова. – Почвоведение. 8: 1022–1032. https://doi.org/10.1134/S0032180X18080099
Zvereva E.L., Kozlov M.V. 2004. Facilitative effects of top-canopy plants on four dwarf shrub species in habitats severely disturbed by pollution. – J. Ecol. 92(2): 288–296. https://doi.org/10.1111/j.0022-0477.2004.00854.x
Zvereva E.L., Kozlov M.V. 2007. Facilitation of bilberry by mountain birch in habitat severely disturbed by pollution: Importance of sheltering. – Environ. Exp. Bot. 60(2): 170–176. https://doi.org/10.1016/j.envexpbot.2006.10.005
Кольская ГМК. Норникель. http://www/kolagmk.ru
Классификация и диагностика почв России. 2004. Смоленск. 342 с.
Переверзев В.Н. 2011. Почвообразование в лесной зоне Кольского полуострова. – Вестн. Кольского научного центра РАН. 2: 74–82.
Мировая реферативная база почвенных ресурсов 2014, исправленная и дополненная версия 2015. Международная система почвенной классификации для диагностики почв и создания легенд почвенных карт. Доклады о мировых почвенных ресурсах № 106. ФАО, Рим. 203 с. http://www.fao.org/3/i3794ru/I3794RU.pdf World Reference Base for Soil Resources. International soil classification system for naming soils and creating legends for soil maps. Rome: FAO, 2014. 191 p.
Базилевич Н.И., Титлянова А.А. 2008. Биотический круговорот на пяти континентах: азот и зольные элементы в природных наземных экосистемах. Новосибирск. 381 с.
Методы изучения лесных экосистем. 2002. СПб. 240 с.
Ладонин Д.В. 2002. Соединения тяжелых металлов в почвах – проблемы и методы изучения. – Почвоведение. 6: 682–692.
Лянгузова И.В. 2016. Тяжелые металлы в северотаежных экосистемах России. Saarbrucken (Германия). 260 с.
Лянгузова И.В. 2017. Динамические тренды содержания тяжелых металлов в растениях и почвах при разном режиме аэротехногенной нагрузки. – Экология. 4: 250–260. https://doi.org/10.7868/S0367059717040114
Воробейчик Е.Л. 1995. Изменение мощности лесной подстилки в условиях химического загрязнения. – Экология. 4: 278–284.
Воробейчик Е.Л. 2002. Изменение пространственной структуры деструкционного процесса в условиях атмосферного загрязнения лесных экосистем. – Изв. АН. Сер. Биол. 3: 368–379.
Лянгузова И.В., Ярмишко В.Т., Евдокимов А.С., Беляева А.И. 2018. Состояние сосновых лесов Кольского полуострова на фоне снижения объемов атмосферных выбросов предприятием цветной металлургии. – Растительные ресурсы. 4: 516–531. https://doi.org/10.1134/S0033994618040039
Кашулина Г.М. 2017. Экстремальное загрязнение почв выбросами медно-никелевого предприятия на Кольском полуострове. – Почвоведение. 7: 860–873. https://doi.org/10.7868/S0032180X17070036
Кашулина Г.М. 2018. Мониторинг загрязнения почв тяжелыми металлами в окрестностях медно-никелевого предприятия на Кольском полуострове. – Почвоведение. 4: 493–505. https://doi.org/10.7868/S0032180X1804010X
Воробейчик Е.Л., Кайгородова С.Ю. 2017. Многолетняя динамика содержания тяжелых металлов в верхних горизонтах почв в районе воздействия медеплавильного завода в период сокращения объемов его выбросов. – Почвоведение. 8: 1009–1024. https://doi.org/10.7868/S0032180X17080135
Воробейчик Е.Л., Трубина М.Р., Хантемирова Е.В., Бергман И.Е. 2014. Многолетняя динамика лесной растительности в период сокращения выбросов медеплавильного завода. – Экология. 6: 448–458. https://doi.org/10.7868/S0367059714060158
Трубина М.Р., Воробейчик Е.Л., Хантемирова Е.В., Бергман И.Е., Кайгородова С.Ю. 2014. Динамика лесной растительности после снижения промышленных выбросов: быстрое восстановление или продолжение деградации? – Доклады Академии наук. 458(6): 721–725. https://doi.org/10.7868/S0869565214300252
Телеснина В.М., Семенюк О.В., Богатырев Л.Г. 2017. Свойства лесных подстилок во взаимосвязи с напочвенным покровом в лесных экосистемах Подмосковья (на примере УОПЭЦ “Чашниково”). – Вест. МГУ. Сер. 17. Почвоведение. 4: 11–20.
Тюлин С.Я., Мазная Е.А. 1991. Некоторые характеристики структуры ценопопуляций черники. – Изучение лекарственных растений Карелии. Петрозаводск. 79–102.
Лянгузова И.В., Примак П.А., Волкова Е.Н., Салихова Ф.С. 2020. Пространственное распределение запасов напочвенного покрова и лесной подстилки в фоновых и дефолиирующих сосновых лесах Кольского полуострова. – Раст. ресурсы. 56(4): 335–350. https://doi.org/10.31857/S0033994620040068
Баккал И.Ю., Горшков В.В. 2003. Характеристика травяно-кустарничкового яруса сосновых лесов в условиях атмосферного загрязнения (Кольский полуостров). – Раст. ресурсы. 39(4): 20–35.
Демаков Ю.П., Исаев А.В., Шарафутдинов Р.Н. 2017. Роль лесной подстилки в борах Марийского Заволжья и вариабельность ее параметров. – В сб.: Научные Труды Государственного природного заповедника “Большая Кокшага”. Вып. 8. С. 15–43.
Swift M.J., Heal O.W., Anderson J.M. 1979. Decomposition in terrestrial ecosystem. Oxford: Blackwell Scientific Publication. 372 p.
Воробейчик Е.Л., Позолотина В.Н. 2003. Микромасштабное пространственное варьирование фитотоксичности лесной подстилки. – Экология. 6: 420–427.
Благодатская Е.В., Пампура Т.В., Богомолова И.Н. и др. 2008. Влияние выбросов мeдно-никелевого комбината на микробные сообщества почв лесных биогеоценозов Кольского полуострова. – Изв. РАН. Сер. биологическая. 2: 232–242.
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы