

Растительные ресурсы, 2022, T. 58, № 1, стр. 87-99
Элементный состав доминирующих видов растений в среднетаежных ельниках (на примере Республики Коми)
Е. А. Робакидзе 1, *, К. С. Бобкова 1, С. И. Наймушина 1
1 Институт биологии Коми научного центра Уральского отделения Российской академии наук
г. Сыктывкар, Россия
* E-mail: robakidze@ib.komisc.ru
Поступила в редакцию 16.08.2021
После доработки 09.11.2021
Принята к публикации 02.12.2021
- EDN: AUFAXM
- DOI: 10.31857/S0033994622010113
Аннотация
Исследовано содержание химических элементов (N, P, K, Ca, Mg, Na, Al, Fe, Mn) в разновозрастной хвое и ветвях Picea obovata Ledeb., листьях Vaccinium vitis-idaea L. и V. myrtillus L., зеленых (Pleurozium schreberi, Dicranum polysetum) и сфагновых (Sphagnum sp.) мхах в еловых лесах средней тайги (Республика Коми). Сравнительный анализ минерального состава доминантных видов проведен в ельниках разнотравно-черничном на подзолистой почве и чернично-сфагновом на подзолисто-глееватой почве. Установлена возрастная динамика содержания исследуемых элементов в хвое P. obovata. Подтверждено, что с увеличением возраста хвои происходит уменьшение содержания биофильных элементов (N, P, K) и возрастает содержание Ca, Al, Mn. Суммарное содержание химических элементов в хвое P. obovata не зависит от ее возраста. Не выявлено значимой зависимости массовой доли азота и серы в хвое P. obovata от условий произрастания. Установлено, что в разных типах еловых лесов отсутствуют достоверные различия в суммарном содержании зольных элементов и валового азота в листьях Vaccinium vitis-idaea и V. myrtillus. В зеленых мхах суммарное содержание зольных элементов в разных типах еловых лесов практически одинаково. Сфагновые мхи в ельнике разнотравно-черничном содержат меньшее количество зольных элементов, чем в ельнике чернично-сфагновом. Достоверные различия в содержании валового азота в зеленых и сфагновых мхах в разных условиях произрастания отсутствуют.
На территории европейского Северо-Востока России леса являются важнейшим компонентом природных ландшафтов. Развитие экономики здесь неразрывно связано с повышением уровня рационального использования лесных ресурсов, организацией новых форм производства, использующих как древесину, так и другие виды растительного сырья. Еловые насаждения в этом регионе, сформированые в основном елью сибирской (Picea obovata Ledeb.), занимают более половины лесопокрытой площади [1, 2]. Ельники, являющиеся зональным типом растительности, выступают важным центром биоразнообразия. Одновременно они выполняют существенную средообразующую функцию [3]. Одним из условий устойчивого управления лесами является изученность состояния лесных экосистем. Любая система контроля природной среды складывается из экологического мониторинга и анализа полученных данных, на основе которого принимаются решения о перспективах функционирования и практического использования экосистем [4].
На территории Республики Коми в подзоне средней тайги еловые леса произрастают на площади 6.74 млн га, из них 74% относятся к спелым и перестойным насаждениям. Старовозрастные ельники относятся, главным образом, к зеленомошной, долгомошной и сфагновой группам типов [3]. В еловых сообществах важными в ценотическом и ресурсном отношении видами являются Vaccinium vitis-idaea L. и V. myrtillus L. [5]. При оценке функционирования еловых фитоценозов и качества лесосырьевых ресурсов представляет интерес изучение химического состава компонентов сообществ [6–9]. Показано, что концентрация химических элементов в органах растений характеризуется большой изменчивостью в зависимости от видовой принадлежности и возраста, региональных климатических особенностей, сезонного развития и лесорастительных условий [10–15].
В надземной части древесных растений выделяется две основные группы органов и ряд фракций, существенно различающихся по биологической роли и специфике участия в процессах обмена веществ: а) фотосинтетические части 1-летних побегов, 2-летних побегов, побегов старше двух лет (отдельно хвоя и ветви); б) ствол и скелетные ветви, имеющие опорные и проводящие функции (древесина, кора, ветви живые неохвоенные, корни). Фотосинтезирующие фракции растений играют основную роль в потреблении зольных элементов и азота. Известно также, что химический состав физиологически активных ассимилирующих органов хвойных растений определяется поглощением элементов из почвы и атмосферы, ретранслокацией их из более старых тканей и выщелачиванием атмосферными осадками. Растения на 50–98% состоят из воды. Сухое вещество, остающееся после высушивания, наполовину состоит из углерода. Минеральная часть (зола) составляет от 0.2 до 20% сухого веса. В листьях растений содержание золы относительно велико. Состав золы зависит от содержания минеральных веществ в почве и любой химический элемент, имеющийся в данном местообитании, может быть обнаружен в растении [16, 17].
Целью данной работы является сравнительный анализ элементного состава доминирующих видов растений фитоценозов среднетаежных еловых лесов (Picea obovata Ledeb.) в разных условиях произрастания.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Работа выполнена в ельниках подзоны средней тайги Республики Коми на территории Ляльского лесоэкологического стационара Института биологии Коми НЦ УрО РАН (62°17′ с.ш, 50°40′ в.д.). Исследования проводились на постоянных пробных площадях (ППП), заложенных в естественно развивающихся спелых ельниках разнотравно-черничном (ППП 38) и чернично-сфагновом (ППП 8). Тип леса определен по [18]. Пробные площади заложены согласно ОСТ 56-69-83 (1983) [19]. Таксационная обработка материала выполнена в соответствии с методическими требованиями [20] (табл. 1).
Таблица 1.
Лесоводственно-таксационная характеристика древостоев ельников Table 1. Mensurational characteristics of spruce tree stands
| Тип леса (№ ППП) Type of forest (site number) |
Состав древостоя1 Composition of tree stand1 |
Возраст, лет Age, year |
Число деревьев, экз./га Number of trees, pcs./hectare |
Запас древесины, м3/га Total stock, m3/hectare |
Средняя высота, м Average height, m |
Средний диаметр, см Average diameter, сm |
||
|---|---|---|---|---|---|---|---|---|
| растущие growing |
сухие dry |
растущие growing |
сухие dry |
|||||
| Ельник разнотравно-черничный Mixed herbs-bilberry spruce forest (PSP 38) | 7Е3Б+С ед. П 7S3B+P single F |
90–170 | 958 | 224 | 333.0 | 14.35 | 18.0 | 22.0 |
| Ельник чернично-сфагновый Bilberry-sphagnum spruce forest (PSP 8) | 9Е1Б+С ед. П 9S1B+P single F |
106–200 | 625 | 44 | 194.0 | 3.4 | 16.0 | 20.0 |
Ельник разнотравно-черничный (Piceеtum herboso-myrtillosum) располагается на очень пологом юго-восточном склоне. Подобные элементы мезорельефа представляют собой зоны выноса и частичного транзита воды, минеральных элементов, химических соединений и характеризуются относительной дренированностью почвенного профиля. Древесный ярус состоит из Picea obovata, Abies sibirica Ledeb., Betula pubescens Ehrh., B. pendula Roth, Pinus sylvestris L., имеет состав 7Е3Б+С ед. П (табл. 1). Древостой накапливает 333 м3/га стволовой древесины. В подлеске встречаются единично кусты Juniperus cоmmunis L., Rosa acicularis Lindl., Sorbus aucuparia L., Salix sp. Травяно-кустарничковый ярус имеет простое синузиальное строение, в его составе представлены типичные бореальные виды: черника (Vaccinium myrtillus L.), брусника (Vaccinium vitis-idaea L.), майник двулистный (Maianthemum bifolium (L.) F.W. Schmidt.), линнея северная (Linnaea borealis L.), седмичник европейский (Trientalis europaea L.), голокучник обыкновенный (Gymnocarpium dryopteris (L.) Newm.) и др. Моховой покров, почти сплошной, образуют Hylocomium splendens (Hedw.) Schimp., Pleurozium schreberi (Brid.) Mitt. с незначительной примесью Dicranum polysetum Sw., Polytrichum commune Hedw., встречаются небольшие пятна сфагновых мхов (Sphagnum sp.)11.
Микрорельеф биогенный сформирован отпавшими и заросшими мхами деревьями, старыми пнями. Почва диагностируется как подзолистая с микропрофилем подзола остаточно-карбонатная [22]. Она характеризуется отчетливой дифференциацией на генетические горизонты. Под подстилочно-торфянистым горизонтом О, мощностью до 6 см (в нижней части присутствуют послепожарные угольки), формируется элювиальный горизонт ELg (6–15 см) – серовато-белесой окраски, супесчаный, с заметными признаками оглеения в виде сизых пятен. На глубине 15–25 см сформирован горизонт EL[e-hf], имеющий неоднородную окраску: серовато-светло-бурый с ржавыми пятнами средний суглинок с резким переходом в субэлювиальный горизонт BEL (25–40 см). С глубины 40 см располагается текстурный горизонт ВТ – темно-бурый тяжелый суглинок, который переходит в горизонты ВС–Сса – бурый тяжелый суглинок с включениями окатанных карбонатных пород.
Ельник чернично-сфагновый (Piceеtum myrtilloso-sphagnosum) располагается на довольно ровной территории. Древесный ярус фитоценоза состоит из ели Picea obovata и сопутствующих пород: березы, с единичным участием сосны и пихты, имеет состав 9Е1Б+С ед. П (табл. 1). Древостой аккумулирует 194 м3/га стволовой древесины. В подлеске встречаются кусты можжевельника, шиповника, рябины, жимолости. Травяно-кустарничковый ярус имеет проективное покрытие 60–70%, его образуют бореальные виды: черника, брусника, майник двулистный, линнея северная, седмичник европейский, хвощ лесной (Equisetum sylvaticum L.), осока шаровидная (Carex globularis L.), костяника (Rubus saxatilis L.), княженика (Rúbus árcticus L.), щитовник мужской (Dryopteris filix-mas (L.) Schott.), грушанка (Pirola rotundifolia L.), золотая розга (Solidago virgaurea L.), герань лесная (Geranium silvaticum L.), ожика волосистая (Luzula pilosa (L.), дудник (Angelica sylvestris L.). Моховой ярус с покрытием 80–90% образован в основном сфагновыми мхами, на прикомлевых возвышениях встречаются пятна Pleurozium schreberi, Hylocomium splendens. Состав растений нижних ярусов свидетельствует об олиготрофных условиях.
В почвенном профиле мощность горизонта О составляет 11–13 см, горизонт отличается четкой дифференциацией на подгоризонты по степени разложенности. Ниже формируются элювиальные и субэлювиальные горизонты (ELg/BELg) с отчетливо выраженными признаками оглеения в виде сизых и сизовато-бурых пятен. Тяжелый гранулометрический состав нижележащих горизонтов (BT1g–BT2g) затрудняет внутрипочвенный отток атмосферных осадков и определяет застойно-промывной тип водного режима. Средняя и нижняя часть почвенного профиля также имеют признаки оглеения, что позволяет отнести данную почву к подзолистым глееватым [22, 23].
Для изучения химического состава растений были отобраны образцы следующих компонентов и фракций: хвоя ели 1–5 года жизни, которая была разделена по годам; тонкие ветви ели последних пяти лет без разделения по возрасту; листья Vaccinium vitis-idaea и Vaccinium myrtillus, зеленые (Pleurozium schreberi, Dicranum polysetum) и сфагновые (Sphagnum sp.) мхи. Сбор материала производился в четвертой декаде июля–начале августа в 2019 г. Образцы растений напочвенного покрова отбирали методом укосов [24]. Образцы хвои и ветвей отбирали у 10–12 деревьев P. obovata на каждой ППП с южной стороны из средней части кроны [25]. Образцы высушивали до воздушно-сухого состояния и размалывали в электрической мельнице до порошкообразной массы [26]. Проводили химический анализ только одной средней пробы образцов растений с каждой ППП. Определено содержание следующих химических элементов: N, P, K, Ca, Mg, Na, Al, Fe, Mn, S.
Химический анализ выполнен в аккредитованной экоаналитической лаборатории Института биологии Коми НЦ УрО РАН (аттестат аккредитации № РОСС RU.0001.511257 от 26 февраля 2014 г.) по аттестованным методикам количественного химического анализа. Валовое содержание элементов минерального питания (калий, кальций, магний, фосфор, марганец, железо, натрий, алюминий, сера) в растительных образцах определяли методом атомно-эмиссионной спектрометрии с индуктивно-связанной плазмой (Спектрометр атомно-эмиссионный с индуктивно-связанной плазмой Spectro Cirosccd, Германия). Валовое содержание азота определяли методом газовой хроматографии на элементном анализаторе (ЕА 1110 (CHNS-O, Италия).
Статистическую обработку данных проводили, используя пакет программ Microsoft Excel 2010 (лицензия Института биологии Коми НЦ УрО РАН). В таблицах указаны средние арифметические значения и их стандартные ошибки.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Элементный состав хвои P. obovata
Хвойные древесные растения имеют существенные отличия от покрытосеменных растений по скорости и направленности обменных процессов в связи с особенностями структуры и многолетним характером развития ассимиляционных органов. Они способны сохранять ассимиляционный аппарат в течение многих лет и функционировать в экстремальных условиях благодаря глубокой перестройке ферментативных систем и выработке биохимических защитных приспособлений, выражающихся в накоплении липидов, смол, углеводов [27, 28]. Продолжительность жизни хвои ели в условиях cредней тайги достигает 14 лет. В процессе ее развития происходят значительные изменения в содержании и соотношении отдельных элементов минерального питания, которые в большей степени выражены в хвое 1–5-го года жизни. Однолетняя хвоя ели является наиболее зольной ее частью [29, 30].
Сравнительный анализ минерального состава хвои ели в исследованных ельниках разнотравно-черничном и чернично-сфагновом показывает, что возрастная динамика содержания исследуемых элементов имеет разные тенденции. Динамика концентрации большинства элементов (P, K, Ca, Na, Al, Mn) в хвое P. obovata с высокой степенью достоверности описывается логарифмической кривой (рис. 1). С увеличением возраста хвои в ней достоверно уменьшается содержание K и P: наиболее высокие концентрации этих элементов наблюдаются в 1-летней хвое. Содержание Ca, Mn и Al в хвое старших возрастов достоверно увеличивается, что свидетельствует о слабой подвижности этих элементов. Содержание элементов, способных к ретранслокации внутри растений (N, P, K) снижается, а концентрация элементов средней подвижности (Mg) не снижается в хвое старших возрастов. Такие закономерности содержания химических элементов в хвое ели разного возраста отмечались и ранее [10, 15, 31].
Рис. 1.
Массовая доля элементов в воздушно-сухом веществе хвои Picea obovata в среднетаежных еловых лесах: 1 – логарифмический тренд данных ППП 8; 2 – логарифмический тренд данных ППП 38; R2 – коэффициент детерминации. По горизонтали – возраст хвои, лет, по вертикали – массовая доля, %. Fig. 1. The mass fraction of elements in spruce (Picea obovata) needles air-dry matter in the middle taiga spruce forest: 1 – logarithmic data trend for PSP 8; 2 – logarithmic data trend for PSP 38; R2 – determination coefficient. X-axis – age of needles, y-axis – mass fraction, %.

Из зольных элементов в однолетней хвое P. obovata накапливается наибольшее количество K, который в растениях способствует синтетической деятельности клеток в качестве активатора многих ферментов. Содержание K в хвое ели первого года было на одном уровне в обоих сообществах (1.20 ± 0.5%). Данные по концентрации этого элемента в 1-летней хвое ели в исследуемых нами среднетаежных ельниках несколько превышают показатели, приведенные для Picea abies L. этой же зоны в условиях Карелии Н.И. Казимировым и Р.М. Морозовой [15]. Относительно высокое накопление K 1-летней хвоей, видимо, является приспособительной реакцией, адаптацией ели к суровым условиям Севера и способствует более энергичному и полному усвоению углекислоты за короткий вегетационный период [32]. Достаточный уровень содержания K, характеризующий обеспеченность ели этим элементом, составляет 0.40% [33]. Можно считать, что рассматриваемые еловые насаждения достаточно обеспечены этим элементом (рис. 1). Для сравнения отметим, что в среднетаежных ельниках в условиях Карелии и северотаежных ельниках Кольского п-ва соответствующие величины составляют 0.46–0.48 и 0.80–1.40% от массы сухой хвои [10, 15].
Из биофильных элементов, содержащихся в однолетней хвое P. obovata, второе место по количеству занимает Ca. Большая потребность ели, как и многих других растений, в этом элементе обусловлена тем, что он входит в состав клеточных стенок, участвует в регулировании азотного и калийного обмена. В исследованных нами еловых лесах содержание Ca в 1-летней хвое ели было на одном уровне в обоих сообществах (0.46 ± 0.1%). Концентрация Ca достоверно увеличивается с возрастом до 1.2 ± 0.3% в хвое 5-го года (рис. 1). В исследованиях Н.А. Артемкиной и др. [34] отмечено накопление Са в хвое P. abies ssp. obovata в условиях северной тайги Кольского п-ва с возрастом: от 0.39% в хвое текущего года до 1.4% в хвое 5–7-го года. Выявлено значительное снижение концентрации Са в хвое ели в дефолиирующих сообществах и техногенных редколесьях [35]. Как было показано ранее [29] такое снижение связано с обеднением почв этим элементом в результате реакций замещения в почвенном поглощающем комплексе основных катионов протонами и катионами тяжелых металлов, входящих в состав выбросов.
Концентрация P – важнейшего элемента, влияющего на содержание хлорофилла и ассимиляцию углекислоты [36], в 1-летней хвое ели в разнотравно-черничном и чернично-сфагновом ельниках составляет 0.23–0.25 ± 0.08% от возд.-сухой массы. В хвое P. abies в ельниках зеленомошных на подзолистых почвах в Карелии содержание Р равно 0.13%, а на Кольском полуострове – 0.19% [10, 15], что значительно меньше, чем в хвое P. obovata в исследованных сообществах. С увеличением возраста массовая доля P уменьшается и составляет в среднем 0.17% в хвое 4-го и 5-го года. В целом содержание Р в хвое ели в ельнике чернично-сфагновом достоверно меньше, чем в ельнике разнотравно-черничном. Оптимальная обеспеченность ели этим элементом наблюдается при его концентрации в хвое, равной 0.14% от массы сухого вещества [33]. Следовательно, недостатка фосфора исследуемые ельники не испытывают.
В хвое P. obovata в значительном количестве содержится Mg, который, как составная часть хлорофилла, оказывает прямое влияние на фотосинтез [36]. Согласно нашим данным, в обоих ценозах содержание этого элемента не зависит от возраста хвои (рис. 1). Выявлена более высокая концентрация Mg в разновозрастной хвое ели в чернично-сфагновом ельнике. В однолетней хвое изученных ельников этот элемент находится в пределах 0.09 ± 0.02–0.11 ± 0.01% от массы сухого вещества. Согласно имеющимся данным [33], минимальное количество Mg в однолетней хвое необходимое для нормальной обеспеченности им ели, составляет 0.07–0.10%. Исходя из этого, можно считать, что ель в исследованных сообществах не испытывает недостатка в этом элементе питания.
Согласно полученным данным, в обоих сообществах содержание Mn и Al в хвое ели достоверно возрастает с увеличением ее возраста и является значимо более низким в чернично-сфагновом ельнике по сравнению с разнотравно-черничным (рис. 1).
В содержании Na в хвое ели наблюдаются достоверные возрастные изменения: в ельнике чернично-сфагновом концентрация Na возрастает от 0.0009 ± 0.0004% от возд.-сухой массы в 1-летней хвое до 0.0015 ± 0.0006% в 4-летней, в ельнике разнотравно-черничном — от 0.0011 ± 0.0004% в 1-летней хвое до 0.0015 ± 0.0005% в 5-летней. Cодержание Fe не зависит от возраста хвои. Не выявлено достоверных различий в содержании этих элементов в хвое ели в разных типах леса.
Суммарное содержание зольных элементов в хвое P. obovata не зависит от ее возраста в обоих исследуемых сообществах (рис. 2). В ельнике чернично-сфагновом оно находится в пределах от 1.95 ± 0.27% от возд.-сухой массы в 1-летней хвое до 2.09 ± 0.29% в 4-летней хвое, а в ельнике разнотравно-черничном – от 2.06 ± 0.28% в 1-летней хвое до 1.98 ± 0.26% в 5-летней хвое. Как видно из данных рис. 2, в исследуемых типах еловых сообществ различия по сумме элементов в хвое ели находятся в пределах ошибки.
Рис. 2.
Суммарное содержание химических элементов в пробах хвои Picea obovata в ельниках чернично-сфагновом (ППП 8) и разнотравно-черничном (ППП 38): 1 – однолетняя, 2 – двухлетняя, 3- трехлетняя, 4 – четырехлетняя; 5 – пятилетняя хвоя. По горизонтали – номер участка, по вертикали – массовая доля, %. Fig. 2. The total elemental content in the samples of spruce (P. obovata) needles in bilberry-sphagnum spruce forest (PSP 8) and mixed herbs-bilberry spruce forest (PSP 38): 1 – one-year-old needles; 2 – two-year needles; 3 – three-year needles; 4 – four-year needles; 5 – five year old needles. X-axis – number of PSP, y–axis – mass fraction, %.
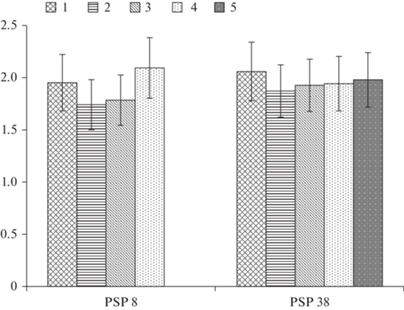
Полученные нами величины концентрации зольных элементов в хвое P. obovata ниже величин, установленных для хвои P. abies в ельниках зеленомошной группы на подзолистых почвах в условиях Карелии и Кольского полуострова [10, 15]. В условиях Карелии содержание зольных элементов в хвое ели достигает 3.27%, а на Кольском п-ве – 2.42%. В 35-летних ельниках Московской обл. зольность 1-летней хвои Picea abies cоставляет 3.72, двухлетней – 4.83, многолетней 6.24% [37], что также значительно выше показателей, полученных нами в ельниках средней тайги.
Анализ динамики содержания валового N в хвое P. obovata показал его значимое снижение с увеличением возраста хвои в обоих исследуемых сообществах (рис. 1). В ельнике чернично-сфагновом содержание N с возрастом уменьшается от 1.23 ± 0.14 до 1.01 ± 0.11% возд. сух. в-ва, а в ельнике разнотравно-черничном – от 1.30 ± 0.14 до 0.93 ± 0.10%.
Содержание общей S (рис. 1) в хвое P. obovata в ельнике чернично-сфагновом составляет от 0.09 ± 0.0 в 1-летней хвое до 0.11 ± 0.05% возд. сух. в-ва в 4-тилетней, в ельнике разнотравно-черничном – от 0.11 ± 0.08 в 1-летней хвое до 0.10 ± 0.05% в 5-летней. Достоверных различий массовой доли S в хвое в зависимости от ее возраста и типа еловых сообществ не выявлено.
Согласно приведенным шкалам обеспеченности фитоценозов минеральными компонентами питания на основе их содержания в однолетней хвое ели [33, 38], исследуемые еловые фитоценозы следует считать достаточно обеспеченными N, K, P, Ca, Mn, Mg.
Содержание химических элементов в ветвях ели
В чернично-сфагновом и разнотравно-черничном ельниках в тонких ветвях P. obovata концентрации всех исследуемых элементов (K, Ca, P, Mg, Mn, Al, Na, Fe, N) и суммарное содержание зольных элементов достоверно не различаются (табл. 2).
Таблица 2.
Содержание химических элементов в компонентах фитомассы среднетаежных еловых лесов, массовая доля элементов в воздушно-сухом веществе, (%) Table 2. Elemental composition of the phytomass components of the middle taiga spruce forests, mass fraction on air dry basis, (%)
| Компонент Component |
K | Ca | P | Mg | Mn | Al | Na | Fe | Nобщ. Ntot |
Сумма без Nобщ Amount without Ntot |
|---|---|---|---|---|---|---|---|---|---|---|
| Ельник разнотравно-черничный (ППП 38) Mixed herbs-bilberry spruce forest (PSP 38) | ||||||||||
| Ветви ели Branches of spruce |
0.5 ± 0.02 | 0.54 ± 0.16 | 0.09 ± 0.02 | 0.06 ± 0.02 | 0.06 ± 0.02 | 0.009 ± 0.002 | 0.003 ± 0.001 | 0.006 ± 0.002 | 0.74± 0.08 | 1.27 ± 0.16 |
| Листья брусники Leaves of cowberry |
0.54 ± 0.20 | 0.47 ± 0.14 | 0.11 ± 0.03 | 0.15± 0.04 | 0.14 ± 0.04 | 0.01 ± 0.002 | 0.001 ± 0.00 | 0.003 ± 0.001 | 1.20 ± 0.13 | 1.42 ± 0.28 |
| Листья черники Leaves of bilberry |
0.95 ± 0.40 | 0.83 ± 0.25 | 0.18 ± 0.05 | 0.19 ± 0.06 | 0.36 ± 0.11 | 0.019 ± 0.005 | 0.001 ± 0.000 | 0.006± 0.001 | 2.25 ± 0.25 | 2.54 ± 0.49 |
| Зеленые мхи Green mosses |
0.59 ± 0.12 | 0.35 ± 0.10 | 0.16 ± 0.05 | 0.10 ± 0.03 | 0.06 ± 0.01 | 0.012 ± 0.003 | 0.005 ± 0.002 | 0.012 ± 0.003 | 0.94 ± 0.10 | 1.29 ± 0.17 |
| Сфагновые мхи Sphagnum mosses |
0.80 ± 0.06 | 0.27 ± 0.08 | 0.15 ± 0.04 | 0.11 ± 0.03 | 0.05 ± 0.01 | 0.008 ± 0.002 | 0.006 ± 0.003 | 0.006 ± 0.002 | 1.00 ± 0.11 | 1.40 ± 0.11 |
| Ельник чернично-сфагновый (ППП 8) Bilberry-sphagnum spruce forest (PSP 8) | ||||||||||
| Ветви ели Branches of spruce |
0.45 ± 0.08 | 0.70 ± 0.17 | 0.09 ± 0.02 | 0.08 ± 0.02 | 0.04 ± 0.01 | 0.009 ± 0.002 | 0.003 ± 0.001 | 0.006 ± 0.002 | 0.75 ± 0.08 | 1.38 ± 0.19 |
| Листья брусники Leaves of cowberry |
0.58 ± 0.09 | 0.60 ± 0.18 | 0.11 ± 0.03 | 0.12 ± 0.04 | 0.13 ± 0.04 | 0.008 ± 0.002 | 0.0007 ± 0.0002 | 0.003 ± 0.0001 | 1.23 ± 0.14 | 1.55 ± 0.21 |
| Листья черники Leaves of bilberry |
1.1 ± 0.04 | 0.90 ± 0.30 | 0.18 ± 0.05 | 0.20 ± 0.06 | 0.28 ± 0.05 | 0.023 ± 0.001 | 0.001 ± 0.0004 | 0.007 ± 0.002 | 2.15 ± 0.24 | 2.69 ± 0.32 |
| Зеленые мхи Green mosses |
0.67 ± 0.09 | 0.34 ± 0.07 | 0.16 ± 0.05 | 0.10 ± 0.03 | 0.05 ± 0.01 | 0.013 ± 0.003 | 0.007 ± 0.002 | 0.013 ± 0.004 | 0.91 ± 0.10 | 1.35 ± 0.13 |
| Сфагновые мхи Sphagnum mosses |
0.90 ± 0.10 | 0.29 ± 0.09 | 0.18 ± 0.05 | 0.14 ± 0.04 | 0.04 ± 0.01 | 0.006 ± 0.001 | 0.008 ± 0.003 | 0.006 ± 0.001 | 1.10 ± 0.12 | 1.57 ± 0.15 |
Элементный состав листьев кустарничков Vaccinium myrtillus и Vaccinium vitis-idaea. Кустарнички характеризуются азотно-кальциево-калиевым типом накопления минеральных элементов. Суммарное содержание зольных элементов в листьях V. vitis-idaea в исследуемых типах ельников (1.42–1.55%) находится на одном уровне (табл. 2). Различий в содержание элементов в листьях V. vitis-idaea в разных типах сообществ еловых лесов не выявлено. Более ранние исследования [39] показали, что содержание зольных элементов в листьях брусники в ельниках, расположенных в фоновом районе, составляет 1.19% от массы сухого вещества. В еловых насаждениях, загрязненных выбросами целлюлозно-бумажного производства, зольность листьев брусники превышает фоновые значения в 1.3 раза и изменяется от 1.44 до 1.79%.
Суммарное содержание зольных элементов в листьях V. myrtillus в ельнике разнотравно-черничном и ельнике чернично-сфагновом (2.54–2.69%) не имеет значимых различий (табл. 2). Надо отметить достаточно высокое содержание Mn в листьях черники в обоих еловых фитоценозах, что характерно для видов р. Vaccinium [10]. Содержание валового азота в листьях V. myrtillus в ельниках обоих типов одинаково. Наши исследования по влиянию выбросов целлюлозно-бумажного производства на элементный состав листьев кустарничков [39] показали, что в еловых лесах отмечается увеличение содержания зольных элементов в листьях черники при приближении к источнику эмиссии с превышением контрольных значений в среднем в 3 раза.
Среди зольных элементов в листьях V. vitis-idaea и V. myrtillus в исследуемых еловых фитоценозах доминируют Ca и K. Высокое содержание кальция является характерной особенностью видов рода Vaccinium. Эти растения относятся к типичным кальцифагам [7]. На долю Ca в сумме зольных элементов приходится 33–39% в листьях брусники и 33% в листьях черники. Содержание K в листьях брусники составляет 37–38%, в листьях черники – 37–41% (рис. 3). Значительное участие в структуре зольного состава у этих кустарничков принимают Mg, P и Mn. В целом, V. vitis-idaea отличается несколько меньшей зольностью и содержанием азота по сравнению с V. myrtillus.
Рис. 3.
Структура содержания зольных элементов в листьях Vaccinium vitis-idaea L. и V. myrtillus L. в еловых фитоценозах. По горизонтали – доля, %. Fig. 3. The structure of mineral content in cowberry and bilberry leaves in spruce phytocenoses. X-axis – proportion, %.

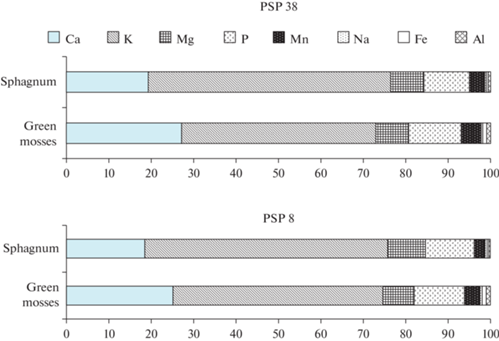
Содержание химических элементов в зеленых (Pleurozium schreberi, Dicranum polysetum) и сфагновых (Sphagnum sp.) мхах
Концентрации всех исследуемых минеральных элементов, суммарное содержание зольных элементов, а также содержание общего азота в зеленых мхах в ельниках разнотравно-черничном и чернично-сфагновом находится на одном уровне (1.29–1.35%) (табл. 2). То же касается содержания зольных элементов и общего азота в сфагновых мхах, произрастающих в этих сообществах (табл. 2). Сравнительный анализ содержания минеральных элементов и азота в зеленых и сфагновых мхах не выявил принципиальных различий.
Среди зольных элементов в побегах Pleurozium schreberi, Dicranum polysetum и Sphagnum sp. так же, как в листьях кустарничков, доминируют Ca и K (рис. 4). Однако, надо отметить, что в отличие от V. vitis-idaea и V. myrtillus, мхи характеризуются более высоким содержанием K по сравнению с Ca. На долю K в сумме зольных элементов у Pleurozium schreber и Dicranum polysetum приходится 46–50%, у Sphagnum sp. – 57%. Участие Ca в составе зольных элементов в побегах зеленых мхов составляет 25–27%, сфагновых мхов – 18–19%. Кроме того, значительное участие в структуре элементного состава мхов принимают Mg, P и Mn. Доля Mn в побегах Pleurozium schreberi, Dicranum polysetum и Sphagnum sp. варьирует от 3 до 5%, что значительно меньше, чем в листьях V. vitis-idaea и V. myrtillus (8–14%). Вместе с тем, побеги и зеленых, и сфагновых мхов отличаются более высокой по сравнению с листьями кустарничков долей Р (11–12 и 7–8% соответственно).
Рис. 4.
Структура содержания зольных элементов в зеленых (Pleurozium schreberi, Dicranum polysetum) и сфагновых (Sphagnum sp.) мхах в еловых фитоценозах. По горизонтали – доля, %. Fig. 4. The structure of mineral content in green (Pleurozium schreberi, Dicranum polysetum) and sphagnum (Sphagnum sp.) mosses in spruce phytocenoses. X-axis – proportion, %.
Отсутствие различий или незначительные различия в содержании исследуемых элементов в разных видах растений в ельнике разнотравно-черничном и ельнике чернично-сфагновом, возможно, обусловлено тем, что эти сообщества произрастают на близких по механическому составу и химическим свойствам почвах. Почвы супесчаные, подстилаемые суглинками, обладают высокой кислотностью. Поглощенных оснований в почвах мало. В минеральных горизонтах резко падает содержание гумуса и азота. Подзолистые почвы ельника разнотравно-черничного характеризуются относительно благоприятными условиями влажности. Влагозапас теплого периода держится в доступной форме. Почвы умеренно дренированы, отмечается периодическое переувлажнение в весеннее-летний период. Фитоценоз ельника чернично-сфагнового входит в динамический ряд заболачивания ельников черничных; подзолисто-глееватая почва этого сообщества в весеннее-летнее время находится почти постоянно в переувлажненном состоянии [3].
ЗАКЛЮЧЕНИЕ
Проведен сравнительный анализ содержания азота и зольных элементов в ассимилирующих органах Picea obovata и доминирующих растений нижних ярусов в естественно развивающихся среднетаежных ельниках разнотравно-черничном и чернично-сфагновом на территории Республики Коми. Согласно полученным результатам, содержание основных элементов минерального питания в хвое ели, листьях кустарничков, зеленых и сфагновых мхах в разных типах ельников, развивающихся на почвах близких по механическому составу и химическим свойствам в большинстве случаев близки между собой. Влажность почв не оказывает существенного влияния на элементный состав растений. Данные по элементному составу растений исследуемых еловых лесов могут быть использованы при проведении комплексного экологического мониторинга лесных сообществ средней тайги в качестве фоновых.
Список литературы
Чертовской В.Г. 1978. Еловые леса европейской части СССР. М.: Лесная пром-ть. 176 с.
Бобкова К. С. Перчаткин П.А., Свойкин В.Ф. 2000. Древесные ресурсы. В кн.: Лесное хозяйство и лесные ресурсы Республики Коми. М. С. 331–369.
Бобкова К.С. Еловые леса средней подзоны тайги. 2006. – В кн.: Коренные еловые леса Севера: биоразнообразие, структура, функции. СПб. P. 99-158.
Булгаков Н.Г. 2002. Индикация состояния природных экосистем и нормирование факторов окружающей среды: обзор существующих подходов. – Успехи современной биологии. 122(2): 115–135.
Лесное хозяйство и лесные ресурсы Республики Коми. 2000. М. 512 с.
Митрофанов Д.П. 1977. Химический состав лесных растений Сибири. Новосибирск. 120 с.
Ingestad T. 1973. Mineral nutrient requirements of Vaccinium vitis-idaea and V. myrtillus. – Physiol. Plant. 29: 239–246. https://doi.org/10.1111/j.1399-3054.1973.tb03099.x
Tamminen P., Starr M., Kubin E. 2004. Element concentrations in boreal, coniferous forest humus layers in relation to moss chemistry fnd soil factors. – Plant and Soil. 259(1–2): 51–58. https://doi.org/10.1023/B:PLSO.0000020940.74369.22
Бобкова К.С., Тужилкина В.В., Робакидзе Е.А. 2005. Ресурсная характеристика древесной зелени Piceae obovata (Pinaceae) в лесах Северо-Востока Европейской России. — Раст. ресурсы. 41(3): 53—64.
Лукина Н.В., Никонов В.В. 1996. Биогеохимические циклы в лесах Севера в условиях аэротехногенного загрязнения. Апатиты. Ч. 1. 213 с. Ч. 2. 192 с.
Родин Л.Е., Базилевич Н.И. 1952. Динамика органического вещества и биологический круговорот зольных элементов и азота в основных типах растительности земного шара. М.; Л. 456 с.
Barcan V.Sh., Kovnatsky E.F., Smetannikova M.S. 1998. Absorption of Heavy Metals in Wild Berries and Edible Mushrooms in an Area Affected by Smelter Emission. – Water, Air and Soil Pollution. 103(1–4): 173–195. https://doi.org/10.1023/A:1004972632578
Дроздова И.В., Алексеева-Попова Н.В. 2008. Оценка макро- и микроэлементного состава некоторых полезных растений Полярного Урала. – Раст. ресурсы. 44 (4): 116–122.
Барановская Н.В., Черненькая У.В. 2015. Особенности накопления химических элементов в чернике обыкновенной (Vaccinium myrtillus) на территории Западной Сибири. – Фундаментальные исследования. 2 (часть 2): 299–306. https://s.fundamental-research.ru/pdf/2015/2-2/36806.pdf
Казимиров Н.И., Морозова Р.М. 1975. Биологический круговорот веществ в ельниках Карелии. Л. 175 с.
Либберт Э. 1976. Физиология растений. Мир. 576 с.
Веретенников А.В. 1987. Физиология растений с основами биохимии. М. 256 с.
Сукачев В.Н., Зонн С.В. 1961. Методические указания к изучению типов леса. М. 144 с.
ОСТ 56-69-83. Пробные площади лесоустроительные. Метод закладки. 1983. М. 60 с.
Лесотаксационный справочник для Северо-Востока европейской части СССР. 1986. Архангельск. 358 с.
Черепанов С.К. 1995. Сосудистые растения России и сопредельных государств. СПб. 990 с.
Полевой определитель почв России. 2008. М. 88 с. http://www.esoil.ru/images/stories/pdf/Field_guide.pdf
Zhangurov E.V., Lebedeva M.P., Zaboeva I.V. 2011. Microstructure of Genetic Horizons in Automorphic Soils of the Timan Ridge. – Eurasian Soil Science. 44: 261–271. https://doi.org/10.1134/S1064229311030203
Методы изучения лесных сообществ. 2002. СПб. 240 с.
Helmisari H.S. 1992. Spatial and age-related variation in nutrient concentration of Pinus sylvestris needles. – Silva Fennica. 26(3): 145–153. https://doi.org/10.14214/sf.a15643
Родин Л.Е., Ремезов Н.П., Базилевич Н.И. 1968. Методические указания к изучению динамики и биологического круговорота в фитоценозах. Л. 143 с.
Козина Л.В. 1995. Метаболизм фотоассимилятов и передвижение веществ у хвойных. Владивосток. 126 с.
Судачкова Н.Е. 1977. Метаболизм хвойных и формирование древесины. Новосибирск. 229 с.
Лукина Н.В., Никонов В.В. 1998. Питательный режим лесов северной тайги: природные и техногенные аспекты. Апатиты. 316 с.
Бобкова К.С. 1993. Биологический круговорот азота и зольных элементов в сосновых биогеоценозах. – В кн.: Эколого-физиологические основы продуктивности сосновых лесов европейского Севера. Сыктывкар. С.127–148.
Робакидзе Е.А., Торлопова Н.В. 2015. Сравнительный анализ элементного состава разновозрастной хвои Piceae obovata (Pinaceae) в условиях аэротехногенного загрязнения целлюлозно-бумажного производства. – Раст. ресурсы. 51(2): 167–180. https://www.elibrary.ru/item.asp?id=23285890
Манаков К.Н., Никонов В.В. 1981. Биологический круговорот минеральных элементов и почвообразование в ельниках Крайнего Севера. Л. 196 с.
Huettl R.F. 1993. Mg deficiency – a new phenomenon in declining forests – symptoms and effects, causes, recuperation. Huettl and Mueller-Dombois (Eds) Forest Decline in the Atlantic and Pacific Region. Springer. Verlag Berlin Heidelberg. P. 97–114. https://doi.org/10.1134/S1995425519070023
Артемкина Н.А., Данилова М.А., Лукина Н.В. 2019. Пространственное варьирование содержания фенольных соединений и элементов питания в хвое ели северотаежных лесов. – Лесоведение. 4: 243–256. https://doi.org/10.1134/S1995425519070023
Сухарева Т.А., Лукина Н.В. 2014. Минеральный состав ассимилирующих органов хвойных деревьев после снижения уровня атмосферного загрязнения на Кольском полуострове. – Экология. 2: 97—104. https://www.elibrary.ru/item.asp?id=21249090
Веретенников А.В. 2006. Физиология растений. М.: Академический проспект. 480 с.
Второва В.Н., Холопова Л.Б. 2010. Распределение биофильных элементов в кроне ели европейской. – Лесоведение. 3: 71–80. https://www.elibrary.ru/item.asp?id=13917414
Wehrmann I. 1963. Möglichkeiten und Grenzen der Blattanalyse in der Fore-swirtschaft. – Landwirtsch. Forsch. 16(2): 130–145.
Робакидзе Е.А., Торлопова Н.В. 2013. Изменение видового состава напочвенного покрова ельников и минерального состава листьев Vaccinium vitis-idaea и V. myrtillus (Ericaceae) в условиях аэротехногенного загрязнения в Республике Коми. – Растительные ресурсы. 49(1): 65–77. https://www.elibrary.ru/item.asp?id=18762302
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы