

Российский физиологический журнал им. И.М. Сеченова, 2019, T. 105, № 10, стр. 1215-1224
Паттерны двигательной активности вибрисс в состояниях сна и бодрствования у новорожденных крыс
А. Р. Насретдинов 1, Г. Р. Валеева 1, Р. Н. Хазипов 1, 2, *
1 Казанский федеральный университет
Казань, Россия
2 Институт нейробиологии Средиземноморья
Марсель, Франция
* E-mail: roustem.khazipov@inserm.fr
Поступила в редакцию 29.06.2019
После доработки 25.07.2019
Принята к публикации 26.07.2019
Аннотация
Цель настоящей работы заключалась в описании примитивных движений вибрисс новорожденных крыс (4–7 день после рождения) линии Вистар в состояниях сна и бодрствования, а также в исследовании корреляции между движениями вибрисс и движениями тела животного. Движения вибрисс регистрировались с помощью высокоскоростной видеорегистрации, а поведенческие состояния животного определялись с помощью регистрации электромиографической активности шейных мышц и одновременной регистрации движений конечностей животного с помощью пьезоэлементов. Было обнаружено, что во время активного сна преимущественно наблюдаются кратковременные движения вибрисс, и значительная часть этих движений возникает одновременно с кратковременными вспышками активности в шейных мышцах и кратковременными миоклоническими подергиваниями тела животного. Во время периодов бодрствования наблюдались преимущественно комплексные длительные движения вибрисс, которые сопровождались длительным повышенным тонусом шейных мышц и комплексными движениями тела и конечностей. Оба типа движений вибрисс происходили преимущественно в ростральном и каудальном направлениях, а амплитуда движений во время бодрствования лишь ненамного превосходила амплитуду движений вибрисс во время сна. Полученные данные свидетельствуют о том, что временнáя организация двигательной активности вибрисс новорожденных крыс значительно отличается в состояниях сна и бодрствования, и что движения вибрисс в обоих поведенческих состояниях с высокой степенью коррелируют с движениями тела животного. Предполагается, что генерализованные примитивные движения вибрисс и тела у новорожденных крыс обусловлены активностью единого центрального паттерн-генератора как в состоянии активного сна, так и в состоянии бодрствования.
Вибриссы – длинные жесткие волосы, располагающиеся на мордочке животного – являются у грызунов (например, у крыс и мышей), а также у ряда других млекопитающих специализированным органом чувств. В отличие от обычных волосков шерсти и подшерстка, которые в основном выполняют теплоизолирующую функцию, вибриссы грызунов предназначены для активного тактильного исследования внешних объектов. Вибриссы своим основанием погружены в подкожный фолликул, к которому подходит около 200 нервных окончаний тригеминального нерва, а положение самого фолликула, угол его наклона и, соответственно, положение самой вибриссы в пространстве контролируются мышечными группами протракторов и ретракторов [1–3]. При исследовании внешних объектов вибриссы осуществляют петлеобразные движения с частотой около 10 Гц преимущественно в ростро-каудальном направлении (так называемый “вискинг”), а тактильная информация, получаемая в результате этих движений, проводится по восходящим путям в соматосенсорную кору головного мозга для анализа расположения и текстуры исследуемого объекта [4, 5]. Представительством каждой из вибрисс в специализированном участке соматосенсорной коры, известной как бочонковая кора, является кортикальная бочонковая колонка, представляющая собой высокоупорядоченную нейрональную сеть, состоящую у крыс примерно из 10 тысяч нейронов [6]. О важности системы вибрисс свидетельствует непропорционально большой размер представительства вибрисс на соматотопической карте соматосенсорной коры крыс – так называемом “ратункулусе” – сравнимый с корковым представительством рук на аналогичном “гомункулусе” у человека.
Формирование системы вибрисс и соответствующих карт в нервной системе крыс происходит в эмбриональном и раннем постнатальном периоде [7–9]. При этом развитие этой упорядоченной системы обусловлено как молекулярными, генетически предопределенными механизмами, так и активностью. Благодаря своей относительной простоте организации, а также удобству манипуляций на уровне сенсорной периферии, система вибрисс стала одной из основных моделей для онтогенетических исследований. Так, было показано, что сенсорная информация, поступающая от вибрисс, играет важную роль в формировании соматосенсорных карт коры головного мозга во время так называемого “критического периода” развития – первой недели после рождения животного [10–13]. Предыдущие исследования также показали, что активация сенсорного входа от вибрисс у новорожденных крыс происходит не только в результате пассивной стимуляции вибрисс (например, со стороны матери и других крысят в гнезде), но и благодаря обратной сенсорной связи, которая активируется во время движений вибрисс самого крысенка [14]. Также было обнаружено, что, несмотря на то, что активный вискинг с характерными высокочастотными упорядоченными движениями вибрисс наблюдается лишь начиная с конца второй недели после рождения [15–17], для новорожденного животного характерны другие, более примитивные формы двигательной активности вибрисс. Эти ранние формы движений вибрисс включают в себя кратковременные движения миоклонического типа (небольшая часть из которых может быть классифицирована как “твитчи” – изолированные подергивания одной вибриссы, а другая часть как коллективные подергивания вибрисс), а также более сложные и более длительные комплексные движения, что в совокупности схоже с организацией примитивных движений тела и конечностей новорожденного [14, 18, 19]. Было также показано, что кратковременные движения вибрисс возникают в основном во время фазы активного сна, и что эти движения ассоциированы с кратковременным повышением активности шейных мышц [18]. Однако до сих пор остается малоизученной комплексная организация примитивных движений вибрисс в контексте поведенческих состояний новорожденных крыс, а также их связь с двигательной активностью тела и конечностей животного. В связи с этим целью настоящей работы являлось комплексное описание примитивных форм движений вибрисс новорожденных крыс в состояниях сна и бодрствования, а также исследование связи между движениями вибрисс и примитивными движениями тела животного.
МЕТОДЫ ИССЛЕДОВАНИЯ
Исследование проводилось в соответствии с принципами Базельской декларации и рекомендациями биоэтического комитета Казанского государственного медицинского университета (протокол N9-2013). Эксперименты проводились на новорожденных крысах линии Вистар возраста P4–7 дней после рождения (P0 соответствует дню рождения). Общая схема эксперимента представлена на рис. 1А. Подготовка к экспериментам в условиях фиксации головы животного проводилась аналогично описанным ранее методам [14, 20]. Вибриссы подстригались до длины 1 мм и окрашивались с помощью черной гуаши, на поверхность кожи рядом с вибриссами наносились метки для последующего разделения движений вибрисс и смещений кожи (рис. 1Б). Регистрация движений вибрисс проводилась с частотой 100 кадров в секунду с помощью высокоскоростной видеокамеры (Promon 501, AOS Technologies, США) и макрообъектива TEC-M55 (Computar, США). Движения конечностей регистрировались с помощью пьезоэлектрических датчиков KPEG165 (Kingstate, Тайвань). Электромиографическая (ЭМГ) активность шейных мышц регистрировалась с помощью стальных игольчатых электродов c толщиной кончика не более 50 мкм. Регистрация двигательной активности проводилась не менее 15 мин для каждого животного. Сигналы, полученные от пьезоэлектрических датчиков и ЭМГ, усиливались и оцифровывались с частотой дискретизации 32 кГц с использованием системы сбора данных Digital Lynx SX (Neuralynx, США). Трекинг движений вибрисс проводился с помощью программного обеспечения ProAnalyst (Xcitex, США). По результатам трекинга движения вибрисс представлялись в виде векторизованной механограммы, для которой проводилась z-oценка, и по превышению порога z = 0 осуществлялась детекция движений. Аналогичным образом определялись периоды повышенной активности на ЭМГ и механограмме конечности. Периоды одновременной повышенной активности на ЭМГ и механограмме конечности общей длительностью более 1 с считались состоянием бодрствования, остальные периоды были отнесены к состоянию сна. Обработка экспериментальных данных и статистический анализ (ранговый тест Уилкоксона) проводились в программной среде Matlab (MathWorks, США). Данные в тексте представлены в виде среднего значения со стандартной ошибкой.
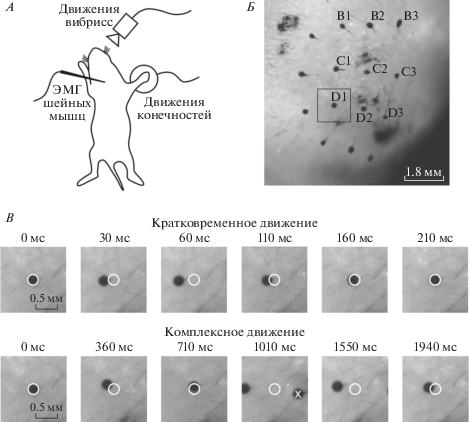
Рис. 1.
Типы движений вибрисс новорожденных крыс. А – Схема эксперимента по одновременной регистрации движений вибрисс, электромиографической (ЭМГ) активности шейных мышц и движений конечности. Б – Фотография правой стороны мордочки крысы с окрашенными кончиками вибрисс. Квадратом выделена область вокруг вибриссы D1, примеры движений которой представлены на В. В – Примеры кратковременного и комплексного движений вибриссы D1. Для каждого типа движений представлены 6 кадров с указанием времени относительно первого кадра, который иллюстрирует положение покоя непосредственно перед началом движения. Исходное положение покоя вибриссы обозначено белой окружностью. Белым значком Х на 4-м кадре внизу обозначена соседняя вибрисса D2.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИX ОБСУЖДЕНИЕ
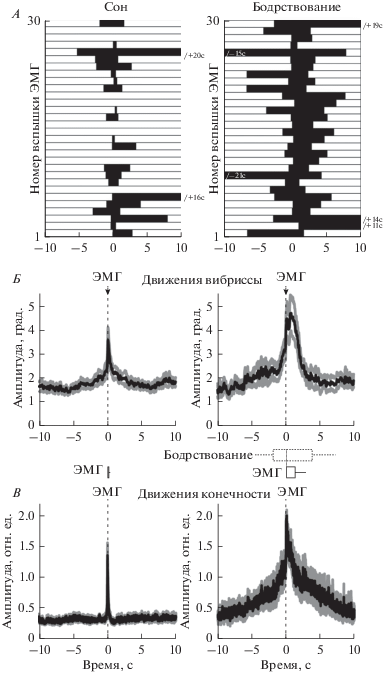
Видеорегистрация движений вибрисс у новорожденных крыс выявила довольно широкий репертуар двигательных паттернов, которые различались по своей длительности, а также по направлению и амплитуде движений вибрисс. На рис. 1В приведены примеры двух наиболее часто встречающихся типов движений: (1) кратковременных миоклонических движений, во время которых вибрисса смещалась на небольшое время в одном направлении, а затем быстро возвращалась в положение покоя, и (2) более длительных комплексных движений, во время которых вибриссы совершали длительные движения по сложной и иногда повторяющейся траектории. Анализ двигательной активности вибрисс, который включал в себя определение времен начала и длительности смещения вибрисс, а также амплитуды и направления этих смещений для каждого из движений вибрисс, проводился отдельно для состояний сна и бодрствования. Состояние животного определялось на основании комплексного анализа электромиографической (ЭМГ) активности шейных мышц и движения конечностей (рис. 2). За бодрствование принимались эпизоды длительностью более 1 с, сопровождающиеся повышением ЭМГ активности шейных мышц и двигательной активности конечностей, определенные на основе методов, предложенных в работе [21]. За периоды сна принимались эпизоды атонии шейных мышц, на фоне которых возникали кратковременные (менее 1 с) вспышки ЭМГ-активности шейных мышц и кратковременные миоклонические подергивания конечностей. На основании группового анализа было выявлено, что бóльшую часть времени новорожденные крысы проводили в состоянии активного сна, каждый из эпизодов которого в среднем длился 33.3 ± ± 3.0 с. Эпизоды сна перемежались периодами бодрствования, длящимися в среднем 3.9 ± 0.3 с, что в целом согласуется с преобладанием активного сна и сравнительно краткой длительностью эпизодов как сна, так и бодрствования у новорожденных животных [21]. Пример одновременной регистрации движения вибриссы D1, ЭМГ шейных мышц и движений задней конечности приведен на рис. 2. Как следует из приведенного примера, эпизоды бодрствования сопровождались как длительными комплексными движениями вибриссы, так и повышенным тонусом шейных мышц и движениями конечностей. В свою очередь, движения вибрисс во время сна имели преимущественно небольшую длительность и часто возникали в ассоциации с кратковременными вспышками ЭМГ-активности шейных мышц и миоклоническими подергиваниями конечностей. Дальнейший количественный анализ движений вибрисс в состояниях сна и бодрствования проводился относительно начала вспышек ЭМГ-активности шейных мышц, которые регистрировались во время этих состояний. На графиках, представленных на рис. 3А, приведены примеры временных соотношений движений вибрисс относительно начала вспышек ЭМГ-активности шейных мышц во время сна и бодрствования. Движения вибрисс наблюдались на фоне большей части (81 ± 3%) кратковременных ЭМГ-вспышек шейных мышц в состоянии сна (n = 5 крыс). Следует, однако, отметить, что во время сна 51 ± 2% движений вибрисс возникали спонтанно, на фоне отсутствия надпороговой ЭМГ-активности шейных мышц (n = 5 крыс). В состоянии бодрствования движения вибрисс возникали практически во всех (99 ± 1%) эпизодах бодрствования, все из которых, по определению, сопровождались длительными вспышками ЭМГ-активности шейных мышц (n = 5 крыс). В результате, корреляция движений вибрисс с ЭМГ-активностью шейных мышц наблюдалась как во время сна, так и во время бодрствования (рис. 3Б). Движения конечностей также коррелировали со вспышками активности шейных мышц в обоих поведенческих состояниях (рис. 3В). Анализ временных соотношений между началами вспышек ЭМГ-активности шейных мышц и движениями вибрисс и конечностей во время сна выявил, что задержки между началом движений конечности и вибриссы после начала вспышки ЭМГ составляли соответственно 4 ± 3 мс и 10 ± 5 мс (n = 5 крыс).
Рис. 2.
Движения вибрисс новорожденной крысы во время сна и бодрствования. А – Пример механограммы вибриссы D1 крысы возраста P6 во время эпизодов сна и бодрствования, определенных на основе ЭМГ-активности шейных мышц и механограммы задней конечности. Прямоугольниками над механограммами вибриссы и конечности, а также над ЭМГ обозначены соответствующие задетектированные события. На механограмме вибриссы пунктирной линией показан порог детекции движений. Б – Фрагменты записи (выделены прямоугольниками на А) во время состояний бодрствования и сна на развернутой временной шкале.

Рис. 3.
Движения вибрисс и конечностей относительно вспышек ЭМГ-активности шейных мышц у новорожденных крыс во время сна и бодрствования. А – Диаграмма движений вибриссы (черные прямоугольники) относительно случайно выбранных 30 ЭМГ-вспышек шейных мышц во время сна и бодрствования у новорожденной крысы возраста Р6. Время 0 с соответствует началу ЭМГ-вспышек. Б, В – Механограммы вибрисс (Б) и конечностей (В), усредненные относительно начала ЭМГ-вспышек во время сна и бодрствования (черным цветом обозначено среднее значение, серым – стандартная ошибка; n = 5 крыс возраста Р4–7). На вставках в центре между Б и В в виде прямоугольников указаны средние значения длительности и соответствующие стандартные отклонения ЭМГ-вспышек, начала которых использовались для усреднения механограмм вибрисс и конечностей во время сна и бодрствования; выше справа также указаны средние времена начала и окончания эпизодов бодрствования (±стандартные отклонения) относительно начала ЭМГ-вспышек.
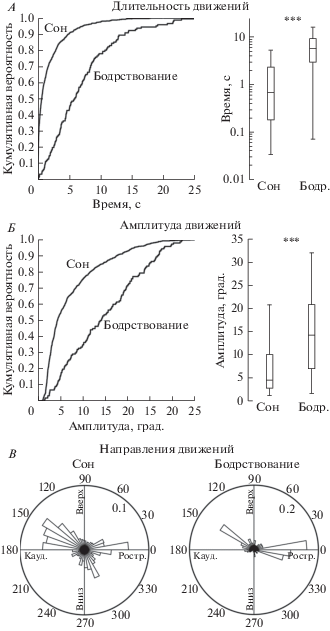
Далее был проведен анализ амплитудно-временных свойств, а также направлений движений вибрисс во время сна и бодрствования (рис. 4). Как следует из кумулятивных гистограмм, длительность движений вибрисс во время бодрствования была существенно больше, чем во время сна (рис. 4А). Статистический анализ выявил, что движения вибрисс во время периодов бодрствования имеют длительность 6.9 ± 0.5 с (n = 140 движений), что значительно превышает длительность движений вибрисс во время сна (1.8 ± 0.1 с; n = 862 движения; n = 5 крыс; p < 0.001; рис. 4А). Также, как следует из кумулятивных гистограмм, амплитуда движений вибрисс во время бодрствования была существенно больше, чем во время сна (рис. 4Б), и статистический анализ выявил, что амплитуда движения вибрисс во время периодов бодрствования имела амплитуду 14.6 ± 0.7° (максимальный угол отклонения вибриссы от положения покоя, n = 140 движений; n = 5 крыс), что превышало значения амплитуд движений вибрисс во время сна (7.3 ± 0.2°; n = 862 движения; n = 5 крыс; р < 0.001; рис. 4Б). Аппроксимация круговых гистограмм направлений отклонений вибрисс с помощью распределений Гаусса позволила выявить следующие наиболее вероятные направления отклонений: 5 ± 4°, 155 ± 7°, 285 ± 6° в состоянии бодрствования и 5 ± 5°, 145 ± 3° в состоянии сна, что соответствует преимущественно ростральному и каудальному направлениям (рис. 4В).
Рис. 4.
Амплитудно-временные характеристики движений вибрисс во время сна и бодрствования. А, Б – Кумулятивные графики распределения длительности (А) и амплитуды (Б) движений вибрисс во время сна и бодрствования (слева) и соответствующие статистические диаграммы (справа) с указанием медианы, 25 и 75 квартилей и максимальных/минимальных значений. *** р < 0.001. В – Нормализованные круговые гистограммы направлений отклонений вибрисс от положения покоя при движениях во время сна и бодрствования (n = 5 животных); углы 0° и 180° соответствуют ростральному и каудальному направлениям отклонения вибрисс, углы 90° и 270° соответствуют направлениям вверх и вниз, значения 0.1 и 0.2 отражают вероятностные характеристики направлений 10% и 20% соответственно (сумма всех значений на гистограмме = 1).
Таким образом, результаты настоящего исследования свидетельствуют о том, что двигательная активность вибрисс новорожденных крыс в значительной степени зависит от поведенческого состояния животного. Если в состоянии сна движения вибрисс имеют короткую длительность и по своим свойствам схожи с миоклоническими подергиваниями тела животного [20, 22, 23], то в состоянии бодрствования вибриссы совершают длительные комплексные движения, более схожие с комплексными генерализованными движениями тела животного [22, 24]. Также обнаруженная нами высокая степень корреляции между ЭМГ-активностью шейных мышц, движениями конечностей и вибрисс свидетельствуют об общем контроле движений как вибрисс, так и тела животного со стороны центрального паттерн-генератора примитивных движений новорожденных крыс. В дальнейших исследованиях представляет интерес идентифицировать нейрональные сети, которые участвуют в контроле ранних форм двигательной активности, а также определить возрастные изменения сенсорно-моторных сетей, которые обуславливают транcформацию ранних паттернов двигательной активности в зрелые паттерны по мере взросления животного, включая исчезновение спонтанных миоклонических подергиваний и кратковременных движений вибрисс во время активного сна, а также возникновение активного вискинга. Также представляет интерес то, какие паттерны активности возникают в бочонковой коре во время различных типов движений вибрисс в состояниях сна и бодрствования, и какова их роль в пластичности синапсов и в формировании соматотопических карт коры головного мозга.
Список литературы
Dorfl J. The musculature of the mystacial vibrissae of the white mouse. J. Anat. 135: 147–154. 1982.
Szwed M., Bagdasarian K., Ahissar E. Encoding of vibrissal active touch. Neuron. 40: 621–630. 2003.
Petersen C.C. The functional organization of the barrel cortex. Neuron. 56: 339–355.2007.
McElvain L.E., Friedman B., Karten H.J., Svoboda K., Wang F., Deschenes M., Kleinfeld D. Circuits in the rodent brainstem that control whisking in concert with other orofacial motor actions. Neuroscience. 368: 152–170. 2018.
Moore J.D., Mercer Lindsay N., Deschenes M., Kleinfeld D. Vibrissa self-motion and touch are reliably encoded along the same somatosensory pathway from brainstem through thalamus. PLoS. Biol. 13: e1002253. 2015.
Feldmeyer D., Brecht M., Helmchen F., Petersen C.C., Poulet J.F., Staiger J.F., Luhmann H.J., Schwarz C. Barrel cortex function. Prog. Neurobiol. 103: 3–27. 2013.
Erzurumlu R.S., Gaspar P. Development and critical period plasticity of the barrel cortex. Eur. J. Neurosci. 35: 1540–1553. 2012.
Luhmann H.J., Khazipov R. Neuronal activity patterns in the developing barrel cortex. Neuroscience. 368: 256–267. 2017.
Lopez-Bendito G. Development of the thalamocortical interactions: Past, present and future. Neuroscience. 385: 67–74. 2018.
Van der Loos H., Woolsey T.A. Somatosensory cortex: Structural alterations following early injury to sense organs. Science. 179: 395–398. 1973.
Simons D.J., Land P.W. Early experience of tactile stimulation influences organization of somatic sensory cortex. Nature. 326: 694–697. 1987.
Fox K. A critical period for experience-dependent synaptic plasticity in rat barrel cortex. J. Neurosci. 12: 1826–1838. 1992.
Feldman D.E., Brecht M. Map plasticity in somatosensory cortex. Science. 310: 810–815. 2005.
Akhmetshina D., Nasretdinov A., Zakharov A., Valeeva G., Khazipov R. The nature of the sensory input to the neonatal rat barrel cortex. J. Neurosci. 36: 9922–9932. 2016.
Welker W.I., Johnson J.I.Jr., Pubols B.H.Jr. Some morphological and physiological characteristics of the somatic sensory system in raccoons. Am. Zool. 4: 75–94. 1964.
Landers M., Philip Zeigler H. Development of rodent whisking: Trigeminal input and central pattern generation. Somatosens. Mot. Res. 23: 1–10. 2006.
Grant R.A., Mitchinson B., Prescott T.J. The development of whisker control in rats in relation to locomotion. Dev. Psychobiol. 54: 151–168. 2012.
Tiriac A., Uitermark B.D., Fanning A.S., Sokoloff G., Blumberg M.S. Rapid whisker movements in sleeping newborn rats. Curr. Biol. 22: 2075–2080. 2012.
Mizuno H., Ikezoe K., Nakazawa S., Sato T., Kitamura K., Iwasato T. Patchwork-type spontaneous activity in neonatal barrel cortex layer 4 transmitted via thalamocortical projections. Cell Rep. 22: 123–135. 2018.
Khazipov R., Sirota A., Leinekugel X., Holmes G.L., Ben Ari Y., Buzsaki G. Early motor activity drives spindle bursts in the developing somatosensory cortex. Nature. 432: 758–761. 2004.
Blumberg M.S., Seelke A.M., Lowen S.B., Karlsson K.A. Dynamics of sleep-wake cyclicity in developing rats. Proc. Natl. Acad. Sci. U.S.A. 102: 14860–14864. 2005.
Gramsbergen A., Schwartze P., Prechtl H.F. The postnatal development of behavioral states in the rat. Dev. Psychobiol. 3: 267–280. 1970.
Tiriac A., Sokoloff G., Blumberg M.S. Myoclonic twitching and sleep-dependent plasticity in the developing sensorimotor system. Curr. Sleep Med. Rep. 1: 74–79. 2015.
Mohns E.J., Blumberg M.S. Neocortical activation of the hippocampus during sleep in infant rats. J. Neurosci. 30: 3438–3449. 2010.
Дополнительные материалы отсутствуют.
Инструменты
Российский физиологический журнал им. И.М. Сеченова