

Российский физиологический журнал им. И.М. Сеченова, 2019, T. 105, № 12, стр. 1502-1513
Роль гуанилатциклазной системы в реализации осморегулирующей функции почек у крыс с различным уровнем вазопрессина в крови
П. Д. Правикова 1, *, Л. Н. Иванова 1
1 Институт цитологии и генетики СО РАН
Новосибирск, Россия
* E-mail: PollyPravi@yandex.ru
Поступила в редакцию 14.05.2019
После доработки 02.08.2019
Принята к публикации 03.08.2019
Аннотация
Исследовано влияние гуанилатциклазной системы на осморегулирующую функцию почек в экспериментах на интактных и на гидратированных крысах линии WAG, а также на вазопрессин-дефицитных крысах линии Brattleboro. Установлено, что в условиях активации гуанилатциклазной системы направленность изменений натрийуретической функции почек зависит от уровня вазопрессина в крови. У крыс WAG с нормальным уровнем вазопрессина в крови активация гуанилатциклазного каскада как в условиях стимуляции синтеза NO экзогенным донором нитропруссидом натрия (НПН), так и при устранении деградации cGMP силденафилом сопровождалась антинатрийуретической реакцией без существенных изменений параметров гидруреза. В то же время у крыс Brattleboro и у гидратированных крыс WAG в условиях применения экзогенного донора NO был выявлен натрийурез. Введение силденафила цитрата крысам Brattleboro, лишенным вазопрессина, сопровождалось иной реакцией – натрийурезом и развитием антидиуретической реакции, в то время как у гидратированных особей WAG был выявлен антидиурез без значимых изменений показателей натрийуретической функции почек. Обсуждаются возможные механизмы, лежащие в основе взаимодействия гуанилатциклазной и аденилатциклазной сигнальных систем, реализующих эффекты вазопрессина и NO на транспорт воды и натрия.
Осмоляльность внеклеточной жидкости, величину которой определяет главным образом концентрация натрия, у млекопитающих – один из главных гомеостатических параметров, в регуляции которого ведущая роль принадлежит почке. Натрий свободно проникает через поры капиллярной мембраны в гломерулярном аппарате, однако в экскретируемой моче содержится лишь небольшая фракция профильтровавшегося катиона. Реабсорбция натрия осуществляется эпителием всех сегментов нефрона и собирательных трубок и находится под контролем многих гуморальных факторов, среди которых основными являются альдостерон, ангиотензин, а также вазопрессин, способный оказывать стимулирующий или ингибирующий эффект на транспорт воды и натрия в зависимости от взаимодействия с рецепторами V2- или V1a-типа различной аффинности [1, 2].
В последние годы в модуляции осморегулирующего эффекта вазопрессина значительное внимание уделяется роли местных, тканевых факторов, выделение которых стимулирует сам гормон, формируя отрицательную обратную связь [3]. Одним из таких факторов, эндогенным субстратом для синтеза которого служит аминокислота аргинин (l-arg), является оксид азота(II), NO. В исследованиях in vitro и in vivo было показано, что NO оказывает ингибирующее влияние на реабсорбцию натрия, усиливая натрийурез [4, 5]. В настоящее время специального внимания заслуживает изучение особенностей действия NO на процессы транспорта воды и натрия в почечных канальцах в зависимости от уровня эндогенного вазопрессина в крови. Известно, что классический сигнальный путь для вазопрессина в эпителии собирательных трубок представлен сAMP/PKA-сигнальным внутриклеточным путем [6], в то время как NO реализует свой эффект через гуанилатциклазную систему путем активации NO/cGMP/PKG-сигнального каскада, все компоненты которого также экспрессируются в эпителии собирательных трубок [7]. Предполагается, что cAMP- и cGMP-зависимые внутриклеточные каскады вазопрессина и NO взаимосвязаны, и NO может осуществлять свое влияние на почечные процессы, модулируя эффект антидиуретического гормона посредством воздействия на активность аденилатциклазной системы [8]. Целью настоящего исследования явилось выявление особенностей осморегулирующей функции почек у крыс с различным уровнем вазопрессина в крови в условиях активации гуанилатциклазной системы.
МЕТОДЫ ИССЛЕДОВАНИЯ
Эксперименты проводились на половозрелых крысах линии WAG (Wistar Albino Glaxo) и гомозиготных вазопрессин-дефицитных крысах Brattleboro с наследственным дефектом синтеза нейрогипофизарного гормона вследствие мутации гена вазопрессина [9]. Для создания модели, адекватной вазопрессин-дефицитным крысам Brattleboro, использовались крысы WAG, у которых путем гидратации (отсутствие плотного корма, свободный доступ к 5%-ному раствору сахарозы в течение 3 сут.) подавлялась секреция эндогенного вазопрессина [10, 11]. Все опытные животные были из ЦКП вивария конвенциональных животных Института цитологии и генетики СО РАН, содержавшиеся на стандартном рационе со свободным доступом к питьевой воде. Все эксперименты были осуществлены в соответствии с Правилами проведения работ с экспериментальными животными (приложение к приказу Министерства здравоохранения № 755 от 12.08.1977) и международными рекомендациями по работе с экспериментальными животными [12].
Активация NO/cGMP/PKG-сигнального каскада осуществлялась двумя способами: введением экзогенного донора NO нитропруссида натрия (НПН) и селективного ингибитора фосфодиэстеразы (PDE5) – фермента деградации cGMP.
1) Активация синтеза NO нитропруссидом натрия
Для активации синтеза NO в условиях краткосрочного эксперимента использовался экзогенный донор NO (дигидрат нитропруссида натрия, НПН, Сигма, США). НПН предварительно растворяли в 5%-ном растворе глюкозы для инфузий и вводили внутрибрюшинно дважды в течение 2.5 ч в дозе 0.075 мг/0.1 мл/100 г массы тела. Контрольным животным вводили 5%-ный раствор глюкозы для инфузий объемом 0.1 мл/100 г массы тела.
Крысы двух линий были подразделены на 6 экспериментальных групп (от 7 до 12 особей в каждой):
● контрольные особи линии WAG и крысы WAG, которым вводился НПН;
● контрольные крысы Brattleboro и особи Brattleboro с введением НПН;
● крысы WAG, у которых подавлялась секреция эндогенного вазопрессина в условиях гидратирующего рациона, и гидратированные крысы WAG с введением НПН.
С учетом действия препарата [13, 14] через 30 мин после последней инъекции животных помещали в индивидуальные клетки с проволочным дном для сбора спонтанно выделяющейся мочи.
2) Ингибирование PDE5 силденафилом
С целью проверки значимости NO/цГМФ/PKG-сигнального каскада в регуляции процесса осмотического концентрирования мочи в условиях действия NO был применен силденафил цитрат (Cildenafil citrate, viagra, Сигма, США) селективный ингибитор фосфодиэстеразы 5 (PDE5), который приводит к устранению деградации cGMP. Силденафил цитрат, который предварительно растворяли в 0.9%-ном водном изотоническом растворе хлорида натрия для инъекций, однократно вводился внутрибрюшинно в дозе 0.5 мг/0.1 мл/100 г массы тела [15]. Контрольным животным вводили раствор хлорида натрия для инфузий объемом 0.1 мл /100 г массы тела.
Все животные были разделены на 6 экспериментальных групп от 7 до 10 особей в каждой:
● контрольные особи линии WAG и крысы WAG с введением силденафила;
● контрольные крысы Brattleboro и крысы Brattleboro с введением силденафила;
● крысы WAG, находившиеся в условиях гидратирующего рациона для подавления секреции эндогенного вазопрессина, и гидратированные крысы WAG с введением силденафила.
Исходя из времени действия препарата [16], через 2 ч животных помещали в индивидуальные клетки с проволочным дном для сбора спонтанно выделяющейся мочи и измерения скорости мочеотделения (диуреза, V).
После окончания экспериментального периода животных, анестезированных тиопенталом натрия (10 мг/100 г массы тела, внутрибрюшинно), декапитировали и забирали пробы крови.
В пробах плазмы крови и мочи измеряли осмоляльность криоскопическим методом (миллиосмометр МТ-2 “Буревестник”, Россия), концентрацию креатинина определяли методом Яффе на спектрофотометре (PD 303 VV; Apel, Saitama, Япония, 490 нм), содержание катионов натрия – методом пламенной фотометрии (Flame photometer 410 Sherwood, Великобритания). По стандартным формулам рассчитывали скорость клубочковой фильтрации (СКФ), индекс осмотического концентрирования (Uosm/Posm), значение реабсорбции осмотически свободной воды (${\text{T}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}^{{\text{c}}}$), клиренс осмотически активных веществ (Cосм), а также скорость экскреции (ЕNa) и экскретируемую фракцию катиона натрия (FENa).
Проверку значимости фактора генотипа (наличие/отсутствие вазопрессина) и действия препарата (активация гуанилатциклазной системы: нитропруссид натрия/сильденафил цитрат) выполняли с помощью двухфакторного дисперсионного анализа (ANOVA), используя пакет программ Statistica 8.0. Данные представлены в виде среднего значения ± ошибка среднего (M ± SEM), достоверность различий между значениями экспериментальных групп оценивали с помощью апостериорного критерия Дункана (post-hoc Duncan’s test) для множественных сравнений. Достоверными считали различия при р < 0.05.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
В настоящем исследовании в соответствии с поставленной целью были оценены параметры, характеризующие гидруретическую и натрийуретическую функции у крыс, обладающих разной способностью к синтезу эндогенного вазопрессина, в условиях активации гуанилатциклазной системы.
Гидруретическая функция у контрольных крыс линий WAG и Brattleboro, лишенных эндогенного вазопрессина, содержавшихся на стандартном питьевом режиме, существенно отличаются (табл. 1). У крыс линии WAG зарегистрирована низкая скорость мочеотделения, отражающая нормальный уровень эндогенного вазопрессина, что подтверждается положительными значениями показателей реабсорбции осмотически свободной воды. В то же время вазопрессин-дефицитные крысы Brattleboro характеризовались высокой скоростью диуреза, отрицательными показателями реабсорбции осмотически свободной воды. У крыс WAG на фоне подавления секреции эндогенного вазопрессина путем гидратации гипотоническим раствором сахарозы показатели гидруретической функции почек были аналогичны контрольным особям Brattleboro: высокая скорость мочеотделения вследствие отрицательных значений реабсорбции осмотически свободной воды (табл. 1). В то же время у гидратированных особей WAG была выявлена антинатрийуретическая реакция (табл. 1), возможно, вследствие действия гидратирующего питьевого режима, приводящего к стимуляции гормонов углеводного обмена (инсулина), участвующих в активации реабсорбции натрия в канальцах почки [17–19].
Таблица 1.
Гидруретическая функция крыс линий WAG и Brattleboro (M ± SEM)
| Показатель | Экспериментальные группы | ||
|---|---|---|---|
| WAG | Brattleboro | ||
| контроль | гидратирующий режим | контроль | |
| V, мл/ч на 100 г | 0.15 ± 0.02 | 1.09 ± 0.11*** | 0.89 ± 0.1*** |
| ${\text{T}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}^{{\text{c}}}$ мл/ч на 100 г | 0.17 ± 0.03 | –0.38 ± 0.13*** | –0.43 ± 0.09*** |
| ENa, мкмоль/ч на 100 г | 21.14 ± 4.02 | 0.61 ± 0.22*** | 7.91 ± 0.97** |
| FENa, % | 0.90 ± 0.13 | 0.10 ± 0.01*** | 0.25 ± 0.04*** |
Примечание. V – скорость мочеотделения; ${\text{T}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}^{{\text{c}}}$ – реабсорбция осмотически свободной воды, ENa– скорость экскреции натрия; FENa– экскретируемая фракция натрия. Различия достоверны при **p < 0.01; ***p < 0.001 –сравнение с контрольной группой WAG. Применялся апостериорный критерий Дункана (post-hoc Duncan’s test).
При активации синтеза NO путем введения его экзогенного донора (НПН) у крыс WAG, содержавшихся в условиях стандартного питьевого режима, не выявлено достоверных изменений как параметров гидруретической функции (рис. 1), так и скорости клубочковой фильтрации (табл. 2). У вазопрессин-дефицитных крыс Brattleboro, а также у крыс WAG при подавлении секреции эндогенного вазопрессина в условиях гидратирующего рациона активация синтеза NO при введении НПН привела к увеличению скорости мочеотделения (рис. 1), обусловленному как подавлением реабсорбции осмотически свободной воды, так и нарастанием СКФ (табл. 2).
Рис. 1.
Влияние активации синтеза NO нитропруссидом натрия (НПН) на скорость мочеотделения и реабсорбцию осмотически свободной воды (${\text{T}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}^{{\text{c}}}$) у крыс WAG и крыс Brattleboro в различных экспериментальных условиях (M ± SEM). Достоверность различий: **P < 0.01; ***P < 0.001 – сравнение с контрольной группой WAG; ###p < 0.001 – сравнение с контрольной группой Brattleboro; P < 0.01 – сравнение с группой гидратированных WAG. Применялся апостериорный критерий Дункана (post-hoc Duncan’s test).
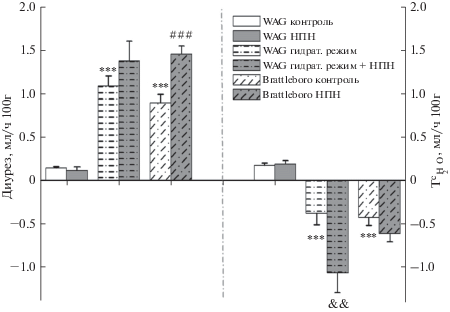
Таблица 2.
Влияние активации синтеза NO (НПН) на скорость клубочковой фильтрации (СКФ) крыс линий WAG и Brattleboro (M ± SEM)
| Экспериментальные группы | СКФ, мл/мин на 100 г | |
|---|---|---|
| контроль | НПН | |
| WAG | 0.23 ± 0.01 | 0.18 ± 0.02 |
| WAG гидратированные | 0.20 ± 0.02 | 0.33 ± 0.03&&& |
| Brattleboro | 0.34 ± 0.05 | 0.79 ± 0.01### |
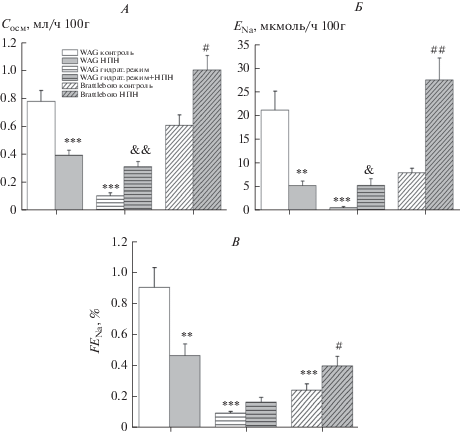
Активация синтеза NO в условиях введения НПН выявила разнонаправленную реакцию натрийуретической функции почек у крыс двух линий. У крыс WAG с высоким уровнем вазопрессина в крови при введении НПН наблюдалось падение клиренса осмотически активных веществ (рис. 2A) в связи с существенным снижением экскретируемой фракции натрия (рис. 2B) и скорости экскреции натрия (рис. 2Б). У вазопрессин-дефицитных крыс в условиях активации синтеза NO была зафиксирована натрийуретическая реакция, характеризующаяся увеличением показателя осмоляльного очищения вследствие нарастания скорости экскреции натрия (рис. 2). У гидратированных крыс WAG, характеризующихся снижением клиренса осмотически активных веществ в соответствии с низкой скоростью экскреции натрия (рис. 2), при введении НПН была выявлена тенденция к развитию натрийуретической реакции (рис. 2) аналогичной ответу, выявленному у вазопрессин-дефицитных крыс Brattleboro в условиях активации синтеза NO.
Рис. 2.
Влияние активации синтеза NO нитропруссидом натрия (НПН) на параметры натрийуретической функции почек у крыс WAG и у вазопрессин-дефицитных крыс Brattleboro в различных экспериментальных условиях (M ± SEM). А – клиренс осмотически активных веществ (Сосм); Б – скорость экскреции натрия (ENa); В – экскретируемая фракция натрия (FENa). Достоверность различий: **p < 0.01; ***p < 0.001 – сравнение с контрольной группой WAG; ##p < 0.01; ###p < 0.001 – сравнение с контрольной группой Brattleboro. Применялся апостериорный критерий Дункана (post-hoc Duncan’s test).
Исходя из схемы внутриклеточного пути действия NO, активировать N-O/сGMP/PKG-сигнальный каскад можно стимулируя как синтез NO, так и звенья гуанилатциклазного сигнального пути. В условиях введения силденафила цитрата, приводящего к повышению содержания сGMP в структурах почки [15, 20] в условиях устранения деградирующего влияния PDE5 на сGMP, у крыс WAG не было выявлено существенных изменений параметров гидруретической функции (рис. 3). В отличие от крыс WAG у крыс Brattleboro инъекция силденафила привела к падению скорости мочеотделения вследствие возрастания реабсорбции осмотически свободной воды (рис. 3). Подобный антидиуретический эффект был найден также у гидратированных крыс WAG в условиях введения сильденафила (рис. 3). Селективное ингибирование PDE5 силденафилом не сопровождалось значимыми изменениями СКФ ни у одной экспериментальной группы животных (табл. 3).
Рис. 3.
Влияние ингибитора PDE5 (силденафил цитрат) на скорость мочеотделения и реабсорбцию осмотически свободной воды (${\text{T}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}^{{\text{c}}}$) в различных экспериментальных условиях (M ± SEM). Достоверность различий: *p < 0.05; ***p < 0.001 – сравнение с контрольной группой WAG; &&&p < 0.001 – сравнение с гидратированной группой WAG; ##p < 0.01; ###p < 0.001 – сравнение с контрольной группой Brattleboro. Применялся апостериорный критерий Дункана (post-hoc Duncan’s test).
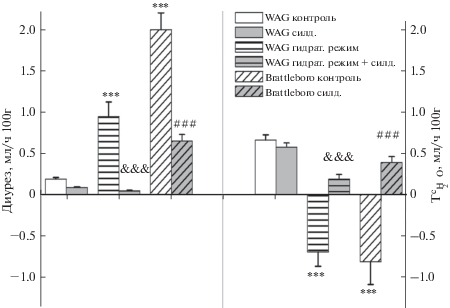
Таблица 3.
Влияние ингибитора PDE5 (силденафил цитрат) на скорость клубочковой фильтрации (СКФ) у крыс линий WAG и Brattleboro (M ± SEM)
| Экспериментальные группы | СКФ, мл/мин на 100 г | |
|---|---|---|
| контроль | силденафил | |
| WAG | 0.32 ± 0.04 | 0.28 ± 0.02 |
| WAG гидратированные | 0.27 ± 0.02 | 0.27 ± 0.04 |
| Brattleboro | 0.43 ± 0.02 | 0.46 ± 0.04 |
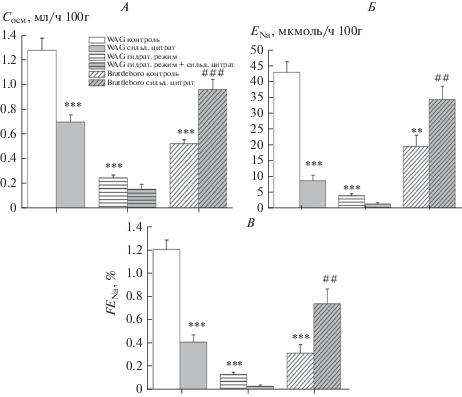
В условиях применения селективного ингибитора PDE5 сильденафила, приводящего к устранению деградации cGMP, были обнаружены разнонаправленные изменения показателей натрийуретической функции почки, которые также наблюдались в условиях применения экзогенного донора NO, НПН. У крыс WAG на фоне снижения клиренса осмотически активных веществ (рис. 4 А) был зафиксирован антинатрийурез (рис. 4 Б). У вазопрессин-дефицитных крыс Brattleboro, напротив, была выявлена натрийуретическая реакция: возрастание скорости экскреции натрия вследствие увеличенной экскретируемой фракции катиона (рис. 4 Б, В). Введение силденафила цитрата гидратированным животным WAG, у которых в условиях подавления секреции эндогенного вазопрессина базальный уровень экскреции натрия был значительно снижен, не привело к значимым изменениям натрийуретической функции почек (рис. 4 ).
Рис. 4.
Влияние ингибитора PDE5 (силденафил цитрат) на параметры натрийуретической функции почек у крыс WAG и у вазопрессин-дефицитных крыс Brattleboro в различных экспериментальных условиях (M ± SEM). А — клиренс осмотически активных веществ (Сосм); Б — скорость экскреции натрия (ENa); В — экскретируемая фракция натрия (FENa). Достоверность различий: **p < 0.01; ***p < 0.001 – сравнение с контрольной группой WAG; ##p < 0.01; ###p < 0.001 – сравнение с контрольной группой Brattleboro. Применялся апостериорный критерий Дункана (post-hoc Duncan’s test).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Согласно современным данным, NO, активирующий cGMP-зависимый сигнальный путь, подавляет реабсорбцию натрия, ингибируя Na+–K+–2Cl– котранспортер в толстом восходящем колене петли Генле путем активации фермента PDE2, который, в свою очередь, устраняет активирующее влияние аденилатциклазной системы на транслокацию котранспортера [4, 21, 22]. В отношении влияния NO на реабсорбцию натрия в собирательных трубках, а также в дистальных сегментах нефрона данные противоречивы. Так, было показано, что активация синтеза NO предсердным натрийуретическим пептидом усиливает сGMP-активированный транспорт натрия через ENaC-каналы в эпителиоцитах мочевого пузыря лягушки [23]. Кроме того, было обнаружено, что аналог cGMP 8-pСPT-cGMP стимулирует экспрессию гена, кодирующего α-субъединицу ENaC-канала [24], а также устраняет самоингибирование ENaC-каналов [25]. Однако другие исследователи сообщают, что активация синтеза NO блокирует активность ENaС-каналов и приводит к развитию натрийуреза [2, 26], а блокада нейрональной синтазы NO (NOS1), напротив, снижает экскрецию натрия [27]. Согласно нашим ранее опубликованным данным, у вазопрессин-дефицитных крыс Brattleboro и у гидратированных крыс WAG в условиях устранения тонического ингибирующего влияния NO на реабсорбцию натрия было выявлено снижение скорости экскреции натрия [28]. Ингибирующий эффект NO на реабсорбцию натрия подтверждается полученными нами экспериментальными данными: активация NO/cGMP/PKG-сигнального пути в условиях введения НПН у вазопрессин-дефицитных крыс Brattleboro и гидратированных крыс WAG сопровождается развитием натрийуретической реакции, что является, согласно данным литературы, следствием прямого тонического ингибирующего действия эндогенного NO на транспорт натрия. Устранение деградации cGMP при введении силденафила, аналогично действию НПН, сопровождалось развитием натрийуреза у вазопрессин-дефицитных крыс Brattleboro. В то же время, введение силденафила гидратированным особям WAG с частично сниженным уровнем эндогенного вазопрессина не вызвало существенных изменений натрийуретической функции, что объясняется, по-видимому, сохранением стимулирующего влияния эндогенного вазопрессина на реабсорбцию натрия. У контрольных крыс WAG антинатрийурез в условиях активации NO/cGMP/PKG-сигнального пути как при введении НПН, так и при применении сильденафила цитрата, по всей видимости, обусловлен изменениями реабсорбции натрия в СТ и в дистальных частях нефрона в результате взаимодействия аденилатциклазной и гуанилатциклазных систем при действии соответственно вазопрессина и NO. Вазопрессин, связываясь с V2-рецепторами, реализует стимулирующий эффект на реабсорбцию натрия в условиях действия сAMP-зависимого сигнального пути аналогично вазопрессин-зависимой активации транслокации AQP2 в апикальную часть мембраны эпителиоцитов собирательных трубок [29]. Гуанилатциклазная система, в свою очередь, по сGMP-зависимому пути способна оказывать активирующее влияние как на вазопрессин-регулируемую транслокацию ENaC-канала [23], так и на стимуляцию экспрессии гена, кодирующего α-субъединицу ENaC-канала [24].
Опубликованные данные о влиянии NO на транспорт воды неоднозначны [30]. Однако в последние годы многие авторы приводят доказательства подавления транспорта воды под влиянием доноров NO вследствие снижения вазопрессин-зависимого содержания cAMP в главных клетках эпителия собирательных трубок [2, 31]. Ингибирующее действие NO на вазотоцин-индуцированный осмотический ток воды было выявлено также на изолированном мочевом пузыре амфибий [32]. При этом найдено, что данный эффект является следствием активации cGMP-зависимой протеинкиназы, которая взаимодействует с гидроосмотическим эффектом вазотоцина на пост-cAMP уровне. Результаты нашего исследования показали, что активация NO/cGMP/PKG-сигнального пути не приводит к значимым изменениям гидруретической функции у крыс WAG с нормальным уровнем эндогенного вазопрессина в крови, что, видимо, связано с преобладанием активирующего влияния гормона на транспорт воды через cAMP-зависимую систему. В то же время, введение экзогенного донора NO НПН приводит к развитию диуретической реакции у вазопрессин-дефицитных крыс Brattleboro и у гидратированных крыс WAG, что является результатом увеличения скорости клубочковой фильтрации в связи с сосудорасширяющим эффектом НПН [33].
Активация гуанилатциклазной системы силденафилом путем влияния на уровень cGMP, в отличие от введения НПН, привела к усилению осмотического концентрирования у вазопрессин-дефицитных крыс Brattleboro и у гидратированных крыс WAG. В условиях стимуляции cGMP/PKG-сигнального каскада при введении ингибитора PDE5 силденафила у гидратированных крыс WAG, а также у вазопрессин-дефицитных крыс Brattleboro наблюдалось проявление антидиуретической реакции вследствие увеличения реабсорбции осмотически свободной воды без значимых изменений скорости клубочковой фильтрации, что согласуется с данными, полученными ранее. Так, например, в условиях применения ингибитора PDE5 в опытах in vivo и in vitro на вазопрессин-дефицитных крысах Brattleboro было выявлено увеличение транслокации AQP2 в апикальную часть мембраны эпителиоцитов собирательных трубок [15]. В экспериментах на изолированном сосочке мыши было показано, что NO увеличивает Ca2+/PKC/NFATc-индуцированную экспрессию AQP2 путем влияния на промотор данного гена [34 ] . Кроме того, в экспериментах in vivo на крысах с литий-индуцированными симптомами несахарного диабета в условиях введения селективного ингибитора PDE5 силденафила, было выявлено снижение полиурии и увеличение осмоляльности экскретируемой мочи [20]. Таким образом, введение сильденафила цитрата крысам в условиях врожденного отсутствия эндогенного вазопрессина или при подавлении его секреции приводит к усилению факультативной реабсорбции воды, в отличие от крыс WAG с нормальным уровнем гормона. Следовательно, в отношении влияния стимуляции NO/сGMP/PKG-сигнального пути на параметры гидруретической функции более значимый эффект принадлежит вазопрессину в реализации гидроосмотического эффекта.
Активация NO/сGMP/PKG-сигнального каскада как путем введения экзогенного донора NO (НПН), так и при устранении деградации cGMP силденафилом приводит к значительным изменениям параметров натрийуретической функции, причем направленность этих изменений зависит от уровня вазопрессина в крови. Непосредственный эффект NО, как было установлено на вазопрессин-дефицитной генетической модели Brattleboro, в условиях активации гуанилатциклазной системы проявлялся в ингибировании реабсорбции натрия, что приводило к развитию натрийуреза. У крыс WAG с врожденной способностью к синтезу вазопрессина, напротив, активация NO/cGMP/PKG- сигнального каскада синергичным способом усиливала антинатрийуретический эффект гормона.
Таким образом, гуанилатциклазная система модулирует осморегулирующую функцию почек, направленность изменения параметров которой при активации NO/cGMP/PKG-сигнального каскада реализуются путем взаимодействия гуанилатциклазного и аденилатциклазного сигнальных путей, причем базальная активность аденилатциклазного пути, зависимого от уровня вазопрессина, является определяющей.
Список литературы
Perucca J., Bichet D.G., Bardou P., Bouby N., Banki L. Sodium excretion in response to vasopressin and selective vasopressin receptor antagonists. J. Am. Soc. Nephrol. 19: 1721–1731. 2008.
Pearce D., Soundararajan R., Trimpert C., Kashlan O.B., Deen P.M.T., Kohan D. E. Collecting duct principal cell transport processes and their regulation. Clin. J. Am. Soc. Nephrol. 10: 135–146. 2015.
Moneer Z., Dyer J.L., Taylor C.W. Nitric oxide co-ordinates the activities of the capacitative and non-capacitative Ca2+ entry pathways regulated by vasopressin. Biochem. J. 370: 439–448. 2003.
Ares G.R., Caceres P.S., Ortiz P.A. Molecular regulation of NKCC2 in the thick ascending limb. Am. J. Physiol. Renal Physiol. 301: 1143–1159. 2011.
Garvin J.L., Herrera M., Ortiz P.A. Regulation of renal NaCl transport by nitric oxide, endothelin, and ATP: Clinical implications. Annu. Rev. Physiol. 73: 359–376. 2011.
Tamma G., Robben J.H., Trimpert C., Boone M., Deen P.M. Regulation of AQP2 localization by S256 and S261 phosphorylation and ubiquitination. Am. J. Physiol. Cell Physiol. 300: 636–646. 2011.
Chang J., Ding Y., Zhou Z., Nie H., Ji H. Transepital fluid and salt re-absorbtion regulated by cGK2 signals. Int. J. Mol. Sci. 19(3): 1–8. 2018.
Bouley R., Hasler U., Lu H.A., Nunes P., Brown D. Bypassing vasopressin receptor signaling pathways in nephrogenic diabetes insipidus. Semin. Nephrol. 28: 266–278. 2008.
Valtin H., Schroeder H.A. Familial hypothalamic diabetes insipidus in rats (Brattleboro rata). 1964 [classical article]. J. Am. Soc. Nephrol. 8: 1333–1341. 1997.
Yamauchi A., Nakanishi T., Takamitsu Y., Sugita M., Imai E., Noguchi T., Fujiwara Y., Kamada T., Ueda N. In vivo osmoregulation of Na/myo-inositol cotransporter mRNA in rat kidney medulla. J. Am. Soc. Nephrol. 5: 62–67. 1994.
Fathollahi A., Daneshgari F., Hanna-Mitchell A.T. Effect of polyuria on bladder function in diabetics versus non-diabetics: an article review. Curr. Urol. 8(3): 119–125. 2015.
Hubrecht R., Kirkwood J. Hertfordshire S.T.(Eds). The UFAW Handbook on the Care and Management of Laboratory Animals. 2010.
Tanahashi M., Sekizawa T., Yoshida M., Suzuki-Kusaba M., Hisa H., Satoh S. Effects of sodium nitroprusside on renal functions and NO-cGMP production in anesthetized dogs. J. Cardiovasc. Pharmacol. 33: 401–408. 1999.
Hottinger D.G., Beebe D.S., Kozhimannil T., Prielipp R.C., Belani K.G. Sodium nitroprusside in 2014: a clinical concepts review. J. Anaesthesiol. Clin. Pharmacol. 30(4): 462–471. 2014.
Bouley R., Pastor-Soler N., Cohen O., McLaughlin M., Breton S., Brown D. Stimulation of AQP2 membrane insertion in renal epithelial cells in vitro and in vivo by the cGMP phosphodiesterase inhibitor sildenafil citrate (Viagra). Am. J. Physiol. Renal Physiol. 288: 1103–1112. 2005.
Assadi F., Sharbaf F.G. Sildenafil for the treatment of congenital nephrogenic diabetes insipidus. Am. J. Nephrol. 42: 65–69. 2015.
Frindt G., Palmer L.G. Effects of insulin on Na and K transporters in the rat CCD. Am. J. Physiol. Renal Physiol. 302: 1227–1233. 2012.
Pavlov T.S., Ilatovskaya D.V., Levchenko V., Li L., Ecelbarger C.M., Staruschenko A. Regulation of ENaC in mice lacking renal insulin receptors in the collecting duct. FASEB. J. 27: 2723–2732. 2013.
Ilatovskaya D.V., Levchenko V., Brands M.W., Pavlov T.S., Staruschenko A. Cross-talk between insulin and IGF-1 receptors in the cortical collecting duct principal cells: Implication for ENaC-mediated Na+ reabsorption. Am. J. Physiol. Renal Physiol. 308: 713–719. 2015.
Sanches T.R., Volpini R.A., Massola Shimizu M.H., Bragança A.C., Oshiro-Monreal F., Seguro A.C., Andrade L. Sildenafil reduces polyuria in rats with lithium-induced NDI. Am. J. Physiol. Renal Physiol. 302: 216–225. 2012.
Mount D.B. Thick ascending limb of the loop of Henle. Clin. J. Am. Soc. Nephrol. 9: 1974–1986. 2014.
Wu P., Gao Z., Ye S., Qi Z. Nitric oxide inhibits the basolateral 10-pS Clchannels through cGMP/PKG signaling pathway in the thick ascending limb of C57BL/6 mice. Am. J. Physiol. Renal Physiol. 310: 755–762. 2016.
Yamada T., Konno N., Matsuda K., Uchiyama M. Frog atrial natriuretic peptide and cGMP activate amiloride-sensitive Na(+) channels in urinary bladder cells of Japanese tree frog, Hyla japonica. J. Comp. Physiol. B. 177: 503– 508. 2007.
Chen J., Zhang H., Zhang X., Yang G., Lu L., Lu X., Wan C., Ijiri K., Ji H., Li Q. Epithelial sodium channel enhanced osteogenesis via cGMP/PKGII/ENaC signaling in rat osteoblast. Mol. Biol. Rep. 41: 2161–2169. 2014.
Han D., Nie H., Su X., Shi X., Bhattarai D., Zhao M., Zhao R., Landers K., Tang H., Zhang L., Ji H. 8-(4-Chlorophenylthio)-Guanosine-39,59-Cyclic Monophosphate-Na Stimulates Human Alveolar Fluid Clearance by Releasing External Na1 Self-Inhibition of Epithelial Na1 Channels. Am. J. Respir. Cell Mol. Biol. 45: 1007–1014. 2011.
Hyndman K.A., Mironova E.V., Giani J.F., Dugas C., Collins J., McDonough A.A., Stockand J.D., Pollock J.S. Collecting Duct Nitric Oxide Synthase 1Я Activation Maintains Sodium Homeostasis During High Sodium Intake Through Suppression of Aldosterone and Renal Angiotensin II Pathways. J. Am. Heart Assoc. 6(10): 1–14. 2017.
Nakano D., Pollock J.S., Pollock D.M. Renal medullary ETB receptors produce diuresis and natriuresis via NOS1. Am. J. Physiol. Renal Physiol. 294: F1205–F1211. 2008.
Правикова П.Д., Иванова Л.Н. Влияние блокады синтеза оксида азота на диуретическую и натрийуретическую функции почки у крыс с различным уровнем эндогенного вазопрессина в крови. Рос. физиол. журн. им. И.М. Сеченова. 104(1): 96–102. 2018. [Pravikova P.D., Ivanova L.N. Effect of nitric oxide (II) synthesis inhibition on the diurethic and natriuretic kidney functions in rats with various blood vasopressin levels. Russ. J. Physiol. 104(1): 96–102. 2018. (In Russ.)].
Boulkroun S. Vasopressin-inducible ubiquitinspecific protease 10 increases ENaC cell surface expression by deubiquitylating and stabilizing sorting nexin 3. Am. J. Physiol. Renal Physiol. 295: 889–900. 2008.
Ortiz P.A., Garvin J.L. Role of nitric oxide in the regulation of nephron transport. Am. J. Physiol. Renal Physiol. 282: 777–784. 2002.
Hyndman K.A., Boesen E.I., Elmarakby A.A., Brands M.W., Huang P., Kohan D.E., Pollock D.M., Pollock J.S. Renal collecting duct NOS1 maintains fluid-electrolyte homeostasis and blood pressure. Hypertension. 62: 91–98. 2013.
Fock E.M., Lavrova E.A., Bachteeva V.T., Chernigovskaya E.V., Parnova R.G. Nitric oxide inhibits arginine-vasotocin-induced increase of water osmotic permeability in frog urinary bladder. Eur. J. Physiol. 448: 197–203. 2004.
Helal I., Fick-Brosnahan G., Reed-Gitomer B., Schrier R. Glomerular hyperfiltration: Definitions, mechanisms and clinical implications. Nat. Rev. Nephrol. 8: 293–300. 2012.
Borghese Albertoni M.F., Bettini L.M., Nitta C.H., de Frutos S., Majowicz M., Gonzalez Bosc L.V. Aquaporin-2 promoter is synergistically regulated by nitric oxide and nuclear factor of activated T cells. Nephron Extra. 1: 124–138. 2011.
Дополнительные материалы отсутствуют.
Инструменты
Российский физиологический журнал им. И.М. Сеченова