

Российский физиологический журнал им. И.М. Сеченова, 2019, T. 105, № 4, стр. 447-455
Кинематика локомоторных движений крыс в течение 7-дневного вывешивания
А. А. Попов 1, В. А. Ляховецкий 1, 3, Н. С. Меркульева 1, 2, 3, О. В. Горский 1, 2, 3, Е. Ю. Баженова 1, П. Е. Мусиенко 1, 2, 3, 4, *
1 Институт физиологии им. И.П. Павлова РАН
Санкт-Петербург, Россия
2 Институт трансляционной биомедицины СПбГУ
Санкт-Петербург, Россия
3 Российский научный центр радиологии и хирургических технологий
им. академика А.М. Гранова
Санкт-Петербург, Россия
4 Клиника детской хирургии и ортопедии МЗ РФ
Санкт-Петербург, Россия
* E-mail: pol-spb@mail.ru
Поступила в редакцию 06.02.2019
После доработки 22.02.2019
Принята к публикации 23.02.2019
Аннотация
Целью работы было изучение динамики изменений сенсомоторного контроля ходьбы в условиях вывешивания задних конечностей. Эксперименты проводились на крысах, у которых кинематика локомоторных движений оценивалась до, во время и после окончания 7-дневного пребывания задних конечностей в антиортостатическом положении по Morey–Holton. При локомоции на тредбане по окончании вывешивания обнаружено увеличение экстензии в голеностопном и коленном суставах. Анализ динамики изменений кинематических параметров локомоторных движений в безопорном положении показал прогрессирующее увеличение экстензии коленного и голеностопного суставов, но снижение экстензии в тазобедренном в течение 7 дней вывешивания. Это соответствовало вынужденному положению задних конечностей при пассивном висении, которое характеризуется гиперэкстензией коленных и голеностопных суставов, но гиперфлексией тазобедренных. На основании полученных данных сделан вывод о том, что устойчивая динамика изменений кинематики ходьбы, которая характеризует работу спинальных нейронных сетей центрального локомоторного генератора, ответственных за формирование двигательного паттерна, может быть вызвана прогрессирующей реорганизацией системы проприоцептивных связей в условиях измененной конфигурации позы тела и суставов конечностей во время вывешивания.
На сегодняшний день модель вывешивания задних конечностей крысы по Новикову–Ильину [1] в модификации Morey–Holton [2], создающая разгрузку задних конечностей, является общепринятой для изучения эффектов гипогравитации [3–6]. В данной модели, как и в случае гипогравитации, у животных наблюдается определенный комплекс изменений состояния опорно-двигательного аппарата в виде атрофии волокон скелетных мышц [7], снижения процента миозина 1 типа в тонической разгибательной камбаловидной мышце [4, 8–10], изменения активности мышц голени [5]. Некоторые из этих изменений прогрессируют со временем [3], иные кратковременны, например, выраженное снижение активности камбаловидной мышцы в первые сутки вывешивания сменяется постепенным возвращением к исходным значениям на 14 день вывешивания [11].
Модель вывешивания задних конечностей интересна и для изучения центральных механизмов контроля позы и локомоции. После вывешивания отмечены изменения не только структуры и активности единичной мышцы, но и координированной работы разных мышц и суставов при локомоции в целом [5, 12, 13]. В частности, после 7-дневного вывешивания выявлено увеличение задержки между окончанием активности передней большеберцовой и началом активности камбаловидной мышц [5], а также длительности фазы опоры локомоторного цикла [14]. Изучение кинематики ходьбы по тредбану после длительного 14-дневного вывешивания у крыс показало гиперэкстензию голеностопного сустава при завершении фазы опоры [15]. Также после вывешивания выявляется экстензорная установка коленных суставов [13]. Кроме того, гиперэкстензия коленных и голеностопных суставов, обусловленная 9-недельным вывешиванием, не восстанавливалась до исходного уровня даже через 8 нед. [12]. Динамические изменения кинематических характеристик в ходе вывешивания, которые могут помочь выявить механизмы происходящей адаптивной перестройки контроля позы и локомоции, мало изучены.
Согласно современным представлениям специализированная нейронная сеть, называемая центральным генератором паттерна (central pattern generator, CPG), является ключевым элементом системы локомоторного контроля [16–18]. Уместно предположить, что адаптивные реакции локомоторной системы как во время, так и после вывешивания, в значительной степени связаны с изменением активности CPG. На сегодняшний день выполнен ряд исследований ходьбы по тредбану до и после вывешивания [5, 12, 13], однако, детального изучения кинематических характеристик локомоторной активности в безопорном положении не проводилось. Также не были описаны адаптивные двигательные изменения в тазобедренном суставе, которые, могут повлиять как на активность мышц голени [19, 20], так и CPG [16].
В соответствие с вышесказанным, в настоящей работе была исследована динамика кинематических характеристик локомоторной активности до, в течение 7 дней вывешивания и после его окончания с целью выяснения сенсомоторных механизмов развивающихся патологических изменений.
МЕТОДЫ ИССЛЕДОВАНИЯ
Исследование проведено на самцах крыс линии Вистар (n = 4) в возрасте 8 нед. Все процедуры проводились согласно Директиве Совета ЕС (86/609/EЭC от 24 ноября 1986 г.), “Правилам проведения работ c использованием экcпеpиментальныx животныx” (пpиказ МЗ CCCP № 775 от 12.08.1977), закону “О защите животныx от жеcтокого обpащения” (гл. IV, cт. 10, 4679/11 ГК от 01.12.1999).
Вывешивание осуществлялось по Новикову–Ильину в модификации Morey–Holton [2, 21]. К хвосту животного с помощью ленты крепился поворотный механизм, подвешенный к потолку клетки. Данный механизм позволял животному контактировать передними лапами с основанием клетки и передвигаться во всех направлениях. Задние конечности были свободны от внешних воздействий и не имели возможности контакта с поверхностью. Угол наклона туловища равнялся приблизительно 30°. Корм и вода постоянно были доступны в неограниченных количествах.
Движения по тредбану. Локомоция животных длилась 1 ч. Скорость движения ленты тредбана в течение первых 5-ти мин составляла 5 м/мин, затем – 8 м/мин. Из всего объема движений подбирались отрезки с непрерывной однонаправленной ходьбой животного с опережающей скоростью, содержащие в среднем 2–3 повторяющихся цикла ходьбы.
Видео-регистрация осуществлялась видеокамерами, расположенными сбоку от тредбана. После бритья латеральной поверхности задней конечности на стороне, обращенной к видеокамере, маркером на коже ставились метки размером приблизительно 2 мм. Метки располагались в проекции передней верхней подвздошной ости, большом вертеле бедренной кости, коленном суставе, латеральной лодыжке, пятом плюсно-фаланговом суставе.
Анализ данных. Измерение углов суставов на видеокадре происходило на основе координат маркерных меток на коже животного. Локомоторный цикл делился на фазу переноса и опоры. Во время переноса последовательно происходит сгибание (F) в трех основных суставах конечности и затем разгибание в коленном и голеностопном суставах (E1). Во время фазы опоры тазобедренный сустав монотонно разгибается, а в двух дистальных суставах происходит прогибание (подфаза E2), а потом разгибание (подфаза E3) [22]. Определение значений углов голеностопного, коленного, тазобедренного суставов проводилось в подфазу E3 (максимальное разгибание суставов) и F (максимальное сгибание суставов) локомоторного цикла (рис. 1А, Б). У вывешенных животных регистрировались эпизоды локомоторной активности в воздухе. Измерения в данном случае проводились в момент максимального разгибания и сгибания всех суставов задней конечности, соответствующих подфазам Е3 и F у животных, локомотирующих на тредбане.
Рис. 1.
Кинематика локомоторной активности крысы по тредбану: A – максимальная экстензия суставов задней конечности в фазу E3 опоры, Б – максимальная флексия суставов задней конечности в фазу F переноса. В – Модель антиортостатического вывешивания задних конечностей крысы по Morey–Holton. Г – Изменение суставных углов задней конечности в пассивном положении во время вывешивания. В-0 – сразу после вывешивания, В-1 – через 1 сутки, В-3 – через 3 суток, В-7 – через 7 суток вывешивания.

Анализ кинематических характеристик локомоторной активности осуществляли на следующих временных этапах эксперимента: до вывешивания; вскоре после начала вывешивания (В-0), через сут (В-1), 3 сут (В-3), 7 сут (В-7), и после его окончания.
Значимость отличий между группами значений параметров определялась на уровне p < 0.05 с помощью метода иерархических линейных моделей [23]. Статистическая обработка данных проводилась в среде технических вычислений Matlab R2016b (Matworks Inc., version 9.1.0.441655).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Изменение конфигурации позы тела и суставов конечностей во время вывешивания
Сразу после вывешивания установилась вынужденная конфигурация позы тела с положением таза и пояса задних конечностей выше передних и, как следствие, пассивным флексорным положением тазобедренного сустава и экстензорным положением коленного и голеностопного суставов (рис. 1В). К 7-м суткам вывешивания выявлено прогрессирование экстензии коленных и голеностопных суставов, однако положение бедра не изменилось (рис. 1Г).
Динамика значений углов суставов нижних конечностей при локомоции по тредбану и во время вывешивания
Голеностопный сустав
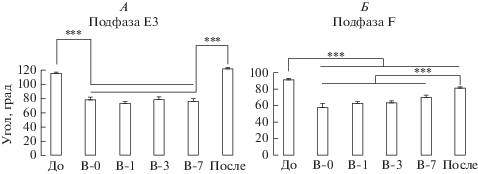
Во время вывешивания, несмотря на безопорное положение задних конечностей, регистрировались эпизоды локомоторной активности в воздухе. Немедленно после вывески животных была выявлена увеличенная экстензия угла голеностопного сустава в аналоге подфазы Е3. Выявлена положительная динамика данного явления: достоверное приращение к 3-му дню вывешивания и продолжающееся недостоверное приращение к 7-му дню; в любом случае, в моменты времени “В-1…7” и “После” величина угла голеностопного сустава оставалась значимо выше, чем в момент времени “До” (рис. 2А). Схожая динамика, но более значимая, наблюдалась и в подфазу F (рис. 2Б), где сразу после вывешивания наблюдалось значимое увеличение угла экстензии голеностопного сустава, который продолжал достоверно увеличиваться к 7 дню. Примечательно, что при локомоции на тредбане значение угла в подфазе F не отличалось от контрольных значений до вывешивания.
Рис. 2.
Динамика значений углов голеностопного сустава крыс в момент максимальной экстензии всех суставов нижней конечности (А, подфаза Е3) и в момент максимальной флексии всех суставов нижней конечности (Б, подфаза F). “До” – контроль до вывешивания, В-0 – сразу после вывешивания, В-1 – через 1 сутки, В-3 – через 3 суток, В-7 – через 7 суток, “После” – после окончания вывешивания.

Коленный сустав
Сразу после вывески животных отмечена значимо увеличенная экстензия угла коленного сустава в Е3. Положительной динамики изменения угла коленного сустава, в отличие от голеностопного сустава, не установлено, однако величина угла осталась значимо выше, чем до вывешивания (рис. 3А). В подфазу F так же, как и в голеностопном суставе, обнаружено значимое увеличение угла экстензии и положительная динамика этого явления в течение 7 суток. После постановки на тредбан угол экстензии коленного сустава снизился, но остался значимо выше, чем до вывешивания (рис. 3Б).
Рис. 3.
Динамика значений углов коленного сустава крыс в момент максимальной экстензии всех суставов нижней конечности (А, подфаза Е3) и в момент максимальной флексии всех суставов нижней конечности (Б, подфаза F). Обозначения экспериментальных групп, как в рис. 2.

Тазобедренный сустав
В противоположность голеностопному и коленному суставам, тазобедренный показал значимое снижение значения угла, как в подфазе Е3 (рис. 4А), так и в подфазе F (рис. 4Б), сразу после вывешивания, которое не показало значимой динамики изменений в течение 7 сут. При постановке на тредбан значение угла для подфазы Е3 вернулось к исходному контрольному значению до вывешивания, но для подфазы F осталось сниженным.
Рис. 4.
Динамика значений углов тазобедренного сустава крыс в момент максимальной экстензии всех суставов нижней конечности (А, подфаза Е3) и в момент максимальной флексии всех суставов нижней конечности (Б, подфаза F). Обозначения экспериментальных групп, как в рис. 2.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Проведенный в настоящей работе анализ кинематики локомоторной активности показал, что сразу после вывешивания наблюдается значительное увеличение экстензии угла коленного и голеностопного суставов как в момент максимальной экстензии (Е3), так и максимальной флексии (F) в сравнении с показателями при локомоции по тредбану. В противоположность этому в тазобедренном суставе выявлено значительно более флексорное положение в Е3 и F. Это может говорить о рефлекторном ответе со стороны нервной системы как на отсутствие опорной загрузки, так и на вынужденное положение суставов нижней конечности при вывешивании.
Способность к альтернирующим локомоторным движениям сохранялась в течение 7 дней вывешивания. Однако наблюдались специфические изменения локомоторного паттерна, которые сохранялись и после прекращения вывешивания. Исследование динамики изменений углов суставов задних конечностей показало, что к 7-м суткам вывешивания экстензия голеностопного и коленного суставов значимо увеличивается по сравнению со значениями на первые сутки. Схожие данные о снижении рабочего диапазона движений голеностопного сустава в сторону экстензии к 7-м суткам вывешивания сообщались ранее [8]. Однако значения угла тазобедренного сустава как в пассивном положении, так и при альтернирующих движениях, не показали динамики за 7 суток вывешивания. Кроме того, значение угла тазобедренного сустава в момент максимальной флексии конечности в воздухе (F) было близко к значению угла в пассивном положении. Следовательно, рабочий диапазон движения бедра в течение 7 суток вывешивания соответствовал в основном экстензии от его исходного пассивного положения.
Как было сказано выше, в основе ритмических локомоторных движений лежит работа центрального генератора паттерна [16–18]. Если принять во внимание условное разделение CPG на уровни генерации ритма (генераторный) и формирования двигательного паттерна (паттерновый) [18], а также выявленную в нашем эксперименте сохранность способности задних конечностей к ритмичным альтернирующим движениям на тредбане и в воздухе, то наиболее очевидной причиной обнаруженных изменений можно полагать развивающуюся реорганизацию паттернового уровня CPG.
Сенсорные сигналы как от опорных рецепторов стопы, так и от проприорецепторов мышц, модулируют работу CPG [24]. Было показано, что вероятным триггерным фактором инициации фазы переноса конечности является специфический проприоцептивный вход от мышц, действующих на тазобедренный сустав [16], и что именно положение бедра, а не стопы или голени, имеет решающее значение для инициации переноса конечности [17]. Согласно результатам, полученным нами при анализе динамики конфигурации позы тела и суставов задней конечности при вывешивании, тазобедренный сустав немедленно оказывается в более флексорном положении и остается таким в течение всего периода вывешивания. В противоположность этому коленный и голеностопный суставы за 7 сут вывешивания изменяют свою конфигурацию постепенно как при пассивном висении, так и при локомоторных движениях в воздухе. Эти данные могут подтверждать гипотезу об особом значении положения бедра для модуляции работы СPG. Такая динамика адаптации суставных углов к условиям вывешивания также хорошо согласуется с данными [25] о проксимально-дистальной иерархии систем координат: сначала для выполнения моторной задачи в измененных условиях калибруются системы координат, связанные с проксимальными суставами.
Обнаруженные изменения кинематики ходьбы по тредбану после 7 сут двигательной разгрузки в виде экстензорной установки коленных и голеностопных суставов сходны с данными, полученными при 14-дневном вывешивании [13, 15]. Было выдвинуто предположение [14], что гиперэкстензия может возникать из-за разбалансировки длины флексоров и экстензоров и развивающейся гипервозбудимости экстензорных мотонейронов в ответ на снижение активности первичных афферентов [26] от соответствующих экстензорных мышц, находящихся в укороченном положении. Этому способствует вынужденное состояние стопы при вывешивании, при котором экстензорные мышцы голеностопного сустава находятся в укороченном состоянии, а флексорные – в растянутом [6, 27]. О значимой роли проприорецепторов мышц экстензоров стопы можно судить и по тому, что при экспериментальной активации проприорецепторов путем вибрации ахиллового сухожилия [28], либо создания постоянного растянутого положения мышц-экстензоров [6], эффекты, возникающие после состояния вывешивания, в значительной степени нивелируются. Мы считаем, что схожим механизмом можно объяснить экстензорную установку коленных суставов, развивающуюся через 7 дней вывешивания, при котором экстензоры колена также находятся в укороченном состоянии. Таким образом, наша работа показывает, что уже к 7-м суткам вывешивания формируются изменения локомоторного контроля, схожие с более длительными сроками вывешивания и связанные с нарушением ключевых модулирующих соматосенсорных влияний на CPG. Предполагается, что причиной изменения кинематических характеристик локомоции может быть не только отсутствие кожного входа от опорных рецепторов стоп, но и прогрессирующая реорганизация системы проприоцептивных связей в условиях измененной конфигурации позы тела и суставов конечностей во время вывешивания. Представляется интересным дальнейшее изучение влияния вывешивания на механизмы контроля позы и локомоции, в частности, сопоставление происходящих изменений активности отдельных мышц задней конечности и работы центрального локомоторного генератора.
Список литературы
Novikov V.E., Ilyin E.A. Age-related reactions of rat bones to their unloading. Aviat. Space En-viron. Med. 52(9): 551–553. 1981.
Morey-Holton E.R., Globus R.K. Hindlimb unloading rodent model: technical aspects. J. Appl. Physiol. (1985). 92(4): 1367–1377. 2002.
Alford E.K., Roy R.R., Hodgson J.A., Edgerton V.R. Electromyography of rat soleus, medial gastrocnemius, and tibialis anterior during hindlimb suspension. Exp. Neurol. 96(3): 635–649. 1987.
Ishihara A., Kawano F., Ishioka N., Oishi H., Higashibata A., Shimazu T., Ohira Y. Effects of running exercise during recovery from hindlimb unloading on soleus muscle fibers and their spinal motoneurons in rats. Neurosci. Res. 48(2): 119–127. 2004.
Canu M.H., Falempin M. Effect of hindlimb unloading on two hindlimb muscles during treadmill locomotion in rats. Eur. J. Appl. Physiol. Occup. Physiol. 75(4): 283–288. 1997.
Fujita N., Fujimoto T., Tasaki H., Arakawa T., Matsubara T., Miki A. Influence of muscle length on muscle atrophy in the mouse tibialis anterior and soleus muscles. Biomed. Res. Feb; 30(1): 39–45. 2009.
Winiarski A.M. Roy R.R., Alford E.K., Chiang P.C., Edgerton V.R. Mechanical properties of rat skeletal muscle after hindlimb suspension. Exp. Neurol. 96(3): 650–660. 1987.
Riley D.A., Slocum G.R., Bain J.L., Sedlak F.R., Sowa T.E., Mellender J.W. Rat hindlimb unloading: soleus histochemistry, ultrastructure, and electromyography. J. Appl. Physiol. 69(1): 58–66. 1990.
Григорьев А.И., Козловская И.Б., Шенкман Б.С. Роль опорной афферентации в организации тонической мышечной системы. Рос. физиол. журн. им. И.М. Сеченова. 90(5): 508–521. 2004. [Grigoryev A.I., Kozlovskaya I.B., Shenkman B.S. The role of support afferentation in the organization of the tonic musculaar system. Russ. J. Physiol. 90(5): 508–521. 2004. (In Russ.)].
Shenkman B.S. From Slow to Fast: Hypogravity-Induced Remodeling of Muscle Fiber Myosin Phenotype. Acta Naturae. 4(31): 47–59. 2016.
Kawano F., Ishihara A., Stevens J.L., Wang X.D., Ohshima S., Horisaka M., Maeda Y., Nonaka I., Ohira Y. Tension- and afferent input-associated responses of neuromuscular system of rats to hindlimb unloading and/or tenotomy. Am. J. Physiol. Regul. Integr. Comp. Physiol. 287(1): R76–R86. 2004.
Ohira Y., Nomura T., Kawano F., Sato Y., Ishihara A., Nonaka I. Effects of nine weeks of unloading on neuromuscular activities in adult rats. J. Gravi.t Physiol. 9(2): 49–59. 2002.
Tajino J., Ito A., Nagai M., Zhang X., Yamaguchi S., Iijima H., Aoyama T., Kuroki H. Discordance in recovery between altered locomotion and muscle atrophy induced by simulated microgravity in rats. J. Mot. Behav. 47(5): 397–406. 2015.
Canu M.H., Falempin M. Effect of hindlimb unloading on locomotor strategy during treadmill locomotion in the rat. Eur. J. Appl. Physiol. Occup. Physiol. 74(4): 297–304. 1996.
Canu M.H., Garnier C., Lepoutre F.X., Falempin M. A 3D analysis of hindlimb motion during treadmill locomotion in rats after a 14-day episode of simulated microgravity. Behav. Brain Res. 2005. 157(2): 309–321.
Duysens J., Pearson K.G. From cat to man: basic aspects of locomotion relevant to motor rehabilitation of SCI. NeuroRehabilitation. 10(2): 107–118.1998.
Grillner S., Rossignol S. On the initiation of the swing phase of locomotion in chronic spinal cats. Brain Res. 146(2): 269–277. 1978.
McCrea D.A., Rybak I.A. Organization of mammalian locomotor rhythm and pattern generation. Brain Res. Rev. 57. 134–146. 2008.
Knikoua M., Rymera W. Hip angle induced modulation of H reflex amplitude, latency and duration in spinal cord injured humans. Clin. Neurophysiol. 113. 1698–1708. 2002.
Kamiya A., Tanabe S., Muraoka Y., Masakado Y. Modulation of the soleus H-reflex during static and dynamic imposed hip angle changes. Int. J. Neurosci. 116(9): 1045–1053. 2006.
Musacchia X.J., Fagette S. Weightlessness simulations for cardiovascular and muscle systems: validity of rat models. J. Gravit. Physiol. 4(3): 49–59. 1997.
Шик М.Л. Управление наземной локомоцией млекопитающих животных. В кн.: Физиология движения. Л. Наука. 234–275. 1976. [Management of terrestrial locomotion of mammals. In the book: Physiology of motion. Leningrad. Nauka. 234–275. 1976. (In Russ.)].
Aarts E., Verhage M., Veenvliet J.V., Dolan C.V., van der Sluis S. A solution to dependency: using multilevel analysis to accommodate nested data. Nat. Neurosci. 17: 491–496. 2014.
Rossignol S. Neuronal control of stereotypic limb movements. In: Handbook of physiology (eds. Rowell L.B. and Sheperd J.T.). 173–216. Oxford Univers. Press. New York. 1996.
Berthoz A. Reference frames for the perception and control of movement. In: Brain and Space. Ed. J. Paillard. Oxford. Oxford Univers. Press. 81–111. 1991.
Webb C.B., Cope TC. Modulation of la EPSP amplitude: the effects of chronic synaptic inactivity. J. Neurosci. 12(1): 338–344. 1992.
Huckstorf B.L., Slocum G.R., Bain J.L., Reiser P.M., Sedlak F.R., Wong-Riley M.T., Riley D.A. Effects of hindlimb unloading on neuromuscular development of neonatal rats. Brain Res. Dev. Brain Res. 7; 119(2): 169–178. 2000.
Falempin M., Albon S. Influence of brief daily tendon vibration on rat soleus muscle in non-weight-bearing situation. J. Appl. Physiol. 87(1): 3–9. 1999.
Дополнительные материалы отсутствуют.
Инструменты
Российский физиологический журнал им. И.М. Сеченова