

Российский физиологический журнал им. И.М. Сеченова, 2022, T. 108, № 1, стр. 36-58
Этосуксимид улучшает когнитивную гибкость при реверсивном обучении у крыс линии WAG/Rij с абсанс-эпилепсией и коморбидной депрессией
Е. А. Федосова 1, *, А. Б. Шацкова 1, К. Ю. Саркисова 1
1 Институт высшей нервной деятельности и нейрофизиологии РАН
Москва, Россия
* E-mail: ekaterina5fedosova@rambler.ru
Поступила в редакцию 08.07.2021
После доработки 07.10.2021
Принята к публикации 08.10.2021
- EDN: DCCTRK
- DOI: 10.31857/S086981392201006X
Аннотация
Крысы линии WAG/Rij являются валидной моделью абсанс-эпилепсии с коморбидной депрессией. Известно, что абсанс-эпилепсия, наряду с симптомами депрессии, сопровождается когнитивными нарушениями, такими как дефицит внимания и когнитивной пластичности, являющейся важнейшим показателем исполнительных функций. Цель настоящей работы – выяснить, имеются ли нарушения когнитивной пластичности у крыс линии WAG/Rij и связаны ли они с абсанс-эпилепсией и коморбидной депрессией. Для выявления различий в когнитивной пластичности сопоставляли обучение (прямая задача) и переучивание (обратная задача) в сложном лабиринте при пищевом подкреплении у крыс линии WAG/Rij в возрасте 2-х мес., когда симптомы абсанс-эпилепсии и коморбидной депрессии отсутствуют, и в возрасте 6-ти месяцев, когда патологический фенотип полностью выражен. Пищевую мотивацию оценивали в тесте “Novelty Suppressed Feeding” (NSF). Этосуксимид (300 мг/кг/день, 17 дней) использовали для подавления симптомов абсанс-эпилепсии и коморбидной депрессии у крыс линии WAG/Rij. Крысы Wistar служили контролем. Обнаружено, что в возрасте 6-ти мес. крысы линии WAG/Rij медленнее выполняют прямую и обратную задачи, делают больше ошибок и проявляют пониженную пищевую мотивацию в тесте NSF (больше латентный период подхода к пище в незнакомом открытом поле и меньше количество съеденной пищи в домашней клетке) по сравнению с крысами Wistar. В возрасте 2-х мес. крысы линии WAG/Rij не отличались от крыс Wistar ни по одному из показателей. Этосуксимид улучшал реверсивное обучение, уменьшал число ошибок и увеличивал пищевую мотивацию только у 6-месячных крыс линии WAG/Rij. Этосуксимид не влиял на реверсивное обучение у крыс Вистар. У 6-месячных крыс линии WAG/Rij обнаружена положительная корреляция (r = 0.66, p < 0.05) между количеством пищи, съеденной в домашней клетке (показатель пищевой мотивации), и скоростью достижения критерия обученности при выполнении обратной задачи (показатель когнитивной пластичности). Результаты свидетельствуют о том, что выявленные нарушения когнитивной пластичности у крыс линии WAG/Rij могут быть следствием пониженной пищевой мотивации как одного из симптомов депрессии, коморбидной абсанс-эпилепсии.
Крысы линии WAG/Rij являются валидной генетической моделью депрессии, ассоциированной с абсанс-эпилепсией [1–5]. Поведенческие особенности крыс линии WAG/Rij, такие как пониженное потребление и предпочтение сахарозы, повышенный уровень иммобильности в тесте вынужденного плавания [1, 2], пониженный уровень исследовательской мотивации в тестах распознавания нового объекта [6, 7] и открытого поля [2, 3], пониженное внимание в тестах распознавания нового объекта и социального распознавания [6, 7], имеют феноменологическое сходство с симптомами депрессии человека: ангедония, ощущение беспомощности, “безнадежности”, утрата интереса [1], неспособность к концентрации внимания и принятию решений [6, 7]. Депрессивно-подобные симптомы в поведении были обнаружены также у другой генетической модели абсанс-эпилепсии – у крыс линии GAERS [8].
Клинические данные свидетельствуют о том, что пациенты с детской абсанс-эпилепсией по сравнению со здоровыми детьми демонстрируют помимо симптомов депрессии когнитивные нарушения, среди которых авторы особо выделяют нарушения исполнительных функций (executive function) и направленного внимания (executive attention) [9]. В последние годы когнитивному аспекту депрессии уделяется особое внимание, так как коррекция когнитивных расстройств имеет первостепенное значение для нормализации повседневного функционирования пациентов и восстановления нарушенной трудоспособности при достижении ремиссии [10].
Данные о когнитивных функциях у экспериментальных моделей абсанс-эпилепсии немногочисленны и противоречивы. Одни исследования свидетельствуют об отсутствии дефицита обучения и памяти в тесте пассивного избегания и в водном лабиринте Морриса у 5-месячных крыс линии WAG/Rij по сравнению с крысами Wistar. Нарушения когнитивных функций в этих тестах были выявлены у крыс линии WAG/Rij только в возрасте 13-ти мес. по сравнению с крысами Wistar аналогичного возраста [11]. Не были обнаружены также нарушения рабочей памяти в радиальном 8-лучевом лабиринте у крыс линии WAG/Rij по сравнению с тремя инбредными линиями крыс (BN/Cpb, ACI и G/Cpb), у которых не наблюдается пик-волновая активность на ЭЭГ – основного симптома абсанс-эпилепсии [12]. Однако в другом тесте (платформа с отверстиями) у крыс линии WAG/Rij и G/Cpb, у которых встречаются абсансные пик-волновые разряды (ПВР) на ЭЭГ, по сравнению с крысами линии BN/Cpb и ACI, у которых ПВР практически отсутствуют, были выявлены нарушения долгосрочной памяти [12]. В других работах у 5–6-месячных крыс линии WAG/Rij были обнаружены нарушения воспроизведения следа памяти после выработки условного рефлекса пассивного избегания [1, 13], сохранность выработанного рефлекса активного избегания у них также была хуже по сравнению с крысами Wistar аналогичного возраста [13]. В нашей предыдущей работе у крыс линии WAG/Rij по сравнению с крысами Wistar были показаны нарушения эпизодической памяти в тестах распознавания нового объекта [6, 7] и социального распознавания [6], которые могут быть связаны с пониженной исследовательской мотивацией – одним из симптомов депрессии, коморбидной абсанс-эпилепсии [7].
Исполнительные функции, дефицит которых чаще всего демонстрируют пациенты с детской абсанс-эпилепсией [9], включают в себя способность планировать, принимать решения и поддерживать на необходимом уровне скорость обработки информации [14]. Вследствие этого может снижаться темп практически всех когнитивных процессов, особенно в тех ситуациях, когда требуется изменить привычную схему деятельности [14, 15]. Существенной частью исполнительных функций является когнитивная гибкость (cognitive flexibility) — способность переключаться с одного вида деятельности на другой. Высокий уровень когнитивной гибкости обеспечивает человеку и животным эффективную адаптацию к постоянно меняющимся условиям окружающей среды [16]. Для оценки когнитивной гибкости часто используют различные вариации тестов, которые предусматривают обучение – выработку животным какой-либо реакции и его последующее переучивание (реверсивное обучение, reversal learning) – переделка этой реакции на альтернативную, чаще всего противоположную исходной [16].
Как у людей [17], так и у животных, – приматов [18, 19] и крыс [20], показано, что выполнение прямой и обратной задачи в тестах на обучение с переучиванием ассоциировано с активностью орбитофронтальной и медиальной префронтальной коры головного мозга [18]. Орбитофронтальная кора получает большое число сигналов от нейронов прилежащего ядра, дорзо-медиального стриатума и миндалины [21]. Нейротоксическое поражение дорсомедиального стриатума у крыс вызывает нарушение процесса переучивания [22]. Аналогичные манипуляции с прилежащим ядром также вызывают дефицит реверсивного обучения: сходные нарушения при выполнении пространственной задачи у обезьян [23] за исключением тех видов данной методики, которые предусматривают использование визуальных стимулов [23], а также нарушение вероятностного реверсивного обучения у крыс [24]. Связи ядра базолатерального комплекса миндалины с дорсомедиальным стриатумом и с прилежащим ядром отвечают за выбор действий, позволяющих получить вознаграждение при реверсивном обучении с положительным подкреплением и избежать наказания при использовании методик с отрицательным подкреплением соответственно [25]. Взаимодействие миндалины с орбитофронтальной корой имеет решающее значение для эмоциональной оценки ожидаемых результатов или последствий [20, 24].
Первые одиночные пик-волновые разряды (ПВР) у крыс линии WAG/Rij появляются примерно в возрасте 2–3 мес. К 5–6-месячному возрасту число и длительность ПВР увеличиваются [26, 27], и по мере усиления пик-волновой активности у этих животных формируется и усиливается депрессивно-подобное поведение [1–3, 5]. В одной из предыдущих работ [28] было проведено исследование обучения с переучиванием в сложном лабиринте с пищевым подкреплением у 6-месячных крыс линии WAG/Rij по сравнению с крысами Wistar аналогичного возраста. Показано, что крысы линии WAG/Rij медленнее обучаются и переучиваются, медленнее достигают пищевого подкрепления при выполнении как прямой, так и обратной задачи и делают больше ошибок в сложном лабиринте по сравнению с крысами Wistar, у которых отсутствуют ПВР и депрессивно-подобные симптомы поведения [28].
Настоящая работа посвящена проверке предположения о том, что дефицит исполнительных когнитивных функций у крыс линии WAG/Rij в тесте реверсивного обучения в сложном лабиринте мог бы быть следствием депрессии, коморбидной абсанс-эпилепсии, а именно – пониженной пищевой мотивации как одного из проявлений депрессивно-подобного поведения. Для этого было проведено обучение с переучиванием (реверсивное обучение) в сложном лабиринте в соответствии с ранее использованной методикой [28]. Оценку пищевой мотивации проводили в тесте на тревожность и пищевую мотивацию “Novelty Suppressed Feeding” (NSF) [29]. В данной работе крыс линии WAG/Rij сравнивали с крысами Wistar в возрасте 2-х мес., когда симптомы абсанс-эпилепсии и депрессии еще отчетливо не проявляются [13, 26, 27], и в возрасте 6-ти мес., когда патологический фенотип полностью выражен [1–7]. Предполагалось, что в возрасте 2-х мес., когда у крыс линии WAG/Rij отсутствуют ПВР на ЭЭГ [26, 27] и симптомы депрессии [13], различия в реверсивном обучении в сложном лабиринте, а также в уровне пищевой мотивации в тесте NSF, у крыс линии WAG/Rij по сравнению с крысами Wistar еще не будут проявляться. С целью подавления симптомов абсанс-эпилепсии и коморбидной депрессии у 6-месячных крыс линии WAG/Rij использовали этосукcимид. Предполагалось, что подавление ПВР, вызываемое этосуксимидом, устранит симптомы депрессии, повысит пищевую мотивацию, что приведет к улучшению реверсивного обучения в сложном лабиринте у крыс линии WAG/Rij.
Животные
Работа выполнена на 26-ти крысах-самцах линии WAG/Rij и 23-х крысах-самцах Wistar. Всех животных содержали в стандартных условиях вивария со свободным доступом к пище и воде по 4–5 особей в клетке (70 × 40 × 40) в условиях естественного цикла день–ночь (день около 10 ч). Данное исследование проведено в соответствии с международными правилами содержания и обращения с животными (Директива Европейского сообщества от 22 сентября 2010 г. – Directive 2010/63/EU) и принципами, изложенными в положениях Института высшей нервной деятельности и нейрофизиологии РАН о работе с экспериментальными животными.
Схема эксперимента
Экспериментальные животные были разделены на 6 групп: крысы линии WAG/Rij в возрасте 2-х (n = 9) и 6-ти мес. (n = 17), а также крысы Wistar в возрасте 2-х (n = 8) и 6-ти мес. (n = 15). Как 6-месячных крыс линии WAG/Rij, так и крыс Wistar аналогичного возраста, делили случайным образом на две группы: экспериментальную (WAG/Rij – n = 8; Wistar – n = 7) и контрольную (WAG/Rij – n = 9; Wistar – n = 8). Животные экспериментальной группы потребляли в течение 17 дней антиабсансный препарат этосуксимид, который был добавлен в поилки с питьевой водой, животные контрольной группы потребляли только воду. Эффект этосуксимида на когнитивную пластичность и пищевую мотивацию оценивали у крыс линии WAG/Rij только в возрасте 6-ти мес., когда пик-волновая активность и симптомы депрессии полностью выражены [1–3, 5]. 2-месячные животные данной линии не потребляли этосуксимид, поскольку у крыс линии WAG/Rij в этом возрасте симптомы патологии отсутствуют [13, 26, 27], так же, как и у “нормальных” крыс Wistar. Показано, что препарат оказывает антидепрессантное влияние (уменьшение времени иммобильности в тесте вынужденного плавания) только у 5‑месячных крыс линии WAG/Rij, когда ПВР хорошо выражены. У крыс, у которых пик-волновая активность отсутствовала (крысы линии WAG/Rij 21-дневного возраста, а также крысы Wistar 21-дневного и 5-месячного возраста), этосуксимид не оказывал антидепрессантного влияния [30]. Данные экспериментальных исследований, свидетельствующих о том, что этосуксимид не обладает собственным антидепрессантным эффектом, не связанным с его антиабсансной активностью [5, 30], подтверждаются данными клиники. У пациентов с депрессивным расстройством ни после однократного перорального приема препарата, ни после его приема в течение 2-х недель не наблюдали значимого снижения показателей депрессии по сравнению с плацебо [31]. Чтобы убедиться в том, что этосуксимид не может воздействовать на исполнительные функции и пищевую мотивацию безотносительно его влияния на пик-волновую активность и поведенческие симптомы депрессии, его предъявляли крысам Wistar, у которых отсутствовала какая-либо патология, по той же схеме, что и крысам линии WAG/Rij.
Этосуксимид
(Suxilep, JENAPHARM, GmbH&Co, KG, Jena, Германия) – антиабсансный препарат первого выбора, который подавляет как пик-волновую активность, так и симптомы коморбидной депрессии [5]. Оптимальный способ для поддержания постоянной концентрации этого вещества в крови – пероральное потребление крысами этосуксимида, который добавляли в поилки с питьевой водой. Это позволяло эффективно блокировать ПВР в течение всего периода потребления данного препарата. Доза этосуксимида составляла 300 мг/кг/день, это было достаточно для того, чтобы подавить пик-волновую активность и при этом не вызвать нежелательные побочные эффекты: гипофагия, летаргия, атаксия [5, 32]. Животные потребляли этосуксимид на протяжении 17-ти дней, в течение которых производили еженедельный контроль массы тела (взвешивание) для коррекции дозы препарата в соответствии с возможными изменениями веса. Поилки с питьевой водой и этосуксимидом меняли 3 раза в неделю [5].
Поведенческие тесты
В данной работе использована схема эксперимента, которая применялась в одной из предыдущих работ [28] (рис. 1). Для оценки когнитивной гибкости и выявления различий в поведении между крысами линии WAG/Rij и крысами Wistar в возрасте 2-х и 6-ти мес., а также между контрольной и экспериментальной группами 6-месячных крыс Wistar и крыс линии WAG/Rij мы использовали тест обучения с переучиванием в сложном лабиринте и тест на тревожность и пищевую мотивацию (NSF). Процедура тестирования в сумме составляла 15 дней: 24 ч – пищевая депривация, 8 сут – обучение в тесте обучения с переучиванием в сложном лабиринте с пищевым подкреплением, 3-е сут – переучивание в тесте обучения с переучиванием в сложном лабиринте с пищевым подкреплением, 24 ч – перерыв, 24 ч – пищевая депривация, затем – тест на тревожность и пищевую мотивацию.
Рис. 1.
День достижения критерия обученности при выполнении прямой (a) и обратной (c) задачи в сложном лабиринте и латентный период взятия пищевого подкрепления при выполнении прямой (b) и обратной (d) задачи. WS – крысы Wistar, потреблявшие воду (контрольная группа), WR – крысы линии WAG/Rij, потреблявшие воду (контрольная группа), WS-ETX – 6-месячные крысы Wistar, потреблявшие ETX (экспериментальная группа), WR-ETX – 6-месячные крысы линии WAG/Rij, потреблявшие ETX (экспериментальная группа). ETX – этосуксимид (300 мг/кг/день, в течение 17 дней). **p < 0.01, ***p < 0.001 – у 6-месячных WR по сравнению с WS того же возраста. xxр < 0.01, xxxр < 0.001 – у 6-месячных крыс по сравнению с 2-месячными. #р < 0.05, ###р < 0.001 - у экспериментальной группы WR по сравнению с контрольной.

Тест обучения с переучиванием в сложном лабиринте с пищевым подкреплением. Данную методику использовали для оценки способности животных к формированию навыка (обучение; прямая задача) и его последующей переделке (переучивание; обратная задача). Экспериментальная установка “сложный лабиринт” представляет собой квадратную камеру размером 60 × 60 × 25 см, которая была разделена на 6 равных по ширине коридоров прозрачными перегородками (5 штук). Каждая из перегородок была снабжена небольшим отверстием прямоугольной формы, которое могло находиться либо в крайнем правом положении – на 10 см от правой стенки лабиринта, либо в крайнем левом – на 10 см от левой стенки. За перегородкой с одним вариантом расположения отверстия следовала перегородка с противоположным вариантом расположения. Такое устройство “сложного лабиринта” позволяло оценить способность животного к решению пространственной задачи – пройти от первого, стартового, отсека лабиринта до шестого, “целевого”, по этой, единственно возможной зигзагообразной траектории. В целевом отсеке животному предъявляли пищевое подкрепление – один стандартный сахарозный шарик (sugar dustless precision pellets), 45 мг (“BioServ”, Frenchtown, NJ, США). С целью угашения ориентировочно-исследовательской реакции и для установления первичной ассоциации между обстановкой лабиринта и получением в нем пищевого подкрепления, крыс адаптировали к условиям эксперимента. С этой целью после однодневной пищевой депривации крыс на 15 мин помещали в “сложный лабиринт”, во всех отсеках которого были равномерно разложены сахарозные шарики. Далее на протяжении всего времени тестирования в сложном лабиринте с пищевым подкреплением крысам давали корм в домашней клетке один раз в день после окончания опыта. Количество корма было не ограничено, но животные могли потреблять его только в течение 10 мин, вода присутствовала в клетке постоянно. Массу тела крыс контролировали с тем, чтобы в ходе пищевой депривации их масса снизилась не более, чем на 20% от исходной, что является необходимым, когда животных подвергают пищевой депривации.
Полное прохождение лабиринта по траектории, заданной расположениями отверстий в перегородках, в течение 3-х мин называли “правильной реакцией”. Ежедневно крыс сажали в лабиринт 5 раз подряд по 3 мин до достижения критерия обученности: воспроизведение крысой 4-х правильных реакций из 5-ти попыток при условии, что она смогла повторить этот результат на следующий день. После достижения критерия обученности перегородки лабиринта переворачивали зеркально, таким образом, что отверстие, которое было в крайнем правом положении, становилось в крайнее левое положение и наоборот. Пищевое подкрепление по-прежнему находилось в “целевом” отсеке лабиринта. Животных переучивали (reversal learning) с уже привычного способа прохождения лабиринта по зигзагообразной траектории на иной навык – по пространственно зеркальной траектории. Крысы выполняли обратную задачу до достижения того же критерия обученности.
После посадки животного в стартовый, ближайший к экспериментатору, отсек камеры регистрировали следующие показатели:
1) день достижения критерия обученности (скорость обучения) – порядковый номер того дня, когда крыса впервые выполнила 4 правильных реакций из 5-ти попыток и смогла повторить этот результат на следующий день;
2) среднее время реакции – латентный период достижения и взятия подкрепления. Оба показателя оценивали отдельно для прямой (обучение) и обратной (переучивание) задачи.
3) число ошибок – отклонения от “правильной” траектории движения, когда крыса после прохождения отверстия в перегородке в очередной отсек камеры сворачивает в “неправильную” сторону;
4) число стоек и коротких эпизодов груминга.
Тест на тревожность и пищевую мотивацию (novelty suppressed feeding test, NSF). Тест NSF направлен на оценку баланса уровня пищевой мотивации и мотивации страха (тревоги) животных [29, 33]. Крыс на 10 мин помещали в новое незнакомое “Открытое поле” (d = 80 см), в центре которого находилось пищевое подкрепление – 1 сахарозный шарик, уже знакомый животному по предыдущему тесту (обучение с переучиванием в сложном лабиринте с пищевым подкреплением). Регистрировали латентный период нахождения и поедания пищевого подкрепления в “Открытом поле”.
После изъятия крысы из незнакомого “Открытого поля” ее на 10 мин возвращали в домашнюю клетку, в которой находилось заранее взвешенное количество привычного для нее корма. Регистрировали количество корма в граммах, съеденное в домашней клетке [28, 29, 33].
Статистическая обработка данных
Статистическую обработку данных осуществляли с помощью пакета статистических программ STATISTICA 6.0. Для каждого исследуемого показателя определяли наличие выбросов по критерию Граббса (Grubb’s test). Если экстремальные значения не выходили за пределы, допустимые для нормального распределения, то применяли дисперсионный анализ (ANOVA). Результаты ANOVA контролировали с использованием непараметрического аналога ANOVA – Kruskal–Wallis H test (one-way ANOVA by ranks) и критерия Манна–Уиттни (Mann–Whitney U test). Для оценки значимости факторов “возраст” и “линия крыс” использовали двухфакторный дисперсионный анализ (two-way ANOVA). Для оценки значимости факторов “линия крыс” и “этосуксимид” у 6-месячных крыс линии WAG/Rij и крыс Wistar также использовали двухфакторный дисперсионный анализ (two-way ANOVA). Определение различий между средними значениями показателей проводили по post-hoc критерию Ньюмена–Кеулса (Newman–Keuls test). Связь между показателями оценивали по коэффициенту корреляции Пирсона.
Тест сложного лабиринта с пищевым подкреплением
Тестирование в сложном лабиринте включало в себя выполнение прямой (обучение) и обратной (переучивание) задач.
Для показателя “день достижения критерия обученности” при выполнении прямой задачи показана высокая значимость факторов “возраст” (F(1, 30) = 35.2, p < 0.001) и “линия крыс” (F(1, 30) = 23.7, p < 0.001). По данным двухфакторного дисперсионного анализа по этому показателю между крысами Wistar и крысами линии WAG/Rij отсутствовали различия в возрасте 2-х мес. (F(1, 15) = 0.001, p = 0.98). По данным однофакторного дисперсионного анализа Краскела–Уоллиса, межлинейные различия у 2-месячных животных также отсутствуют (H(1, 17) = 1.32, p = 0.25). Однако в возрасте 6-ти мес. скорость обучения у контрольных крыс линии WAG/Rij была значимо меньше по сравнению с контрольными крысами Wistar (F(1, 15) = 44.1, p < 0.001). Однофакторный дисперсионный анализ Краскела–Уоллиса подтверждает наличие значимых межлинейных различий по этому показателю в возрасте 6-ти мес. (H(1, 17) = 11.9, p < 0.001) (рис. 1a). Как 2-месячные крысы Wistar (3.9 ± 0.4 дня), так и 2-месячные крысы линии WAG/Rij (3.9 ± 0.3 дня) быстрее достигают критерия обученности при выполнении прямой задачи, чем 6-месячные крысы Wistar (4.2 ± 0.3 дня) и 6-месячные крысы линии WAG/Rij, которые не потребляли этосуксимид (7.6 ± 0.3 дня) соответственно (рис. 1a). Различия по данному показателю между 2- и 6-месячными контрольными крысами линии WAG/Rij достигали уровня статистической значимости (F(1, 16) = 106.2, p < 0.001); H(1, 18) = 14.2, p < 0.001) и оставались на уровне тенденции между 2- и 6-месячными контрольными крысами Wistar (F(1, 14) = 0.39, p = 0.54); H(1, 16) =1.11, p = 0.32) (рис. 1a).
Согласно однофакторному ANOVA, у крыс линии WAG/Rij, которые принимали этосуксимид, скорость обучения была больше по сравнению с контрольными животными той же линии (F (1, 15) = 6.9, р = 0.02), потреблявшими воду. Данные дисперсионного анализа Краскела–Уоллиса (H(2, 20) = 9.02, p = 0.01) подтверждают данные ANOVA. В отличие от межлинейных различий по этому показателю между контрольными группами животных (7.5 ± 0.4 день, WAG/Rij; 4.2 ± 0.4 день, Wistar), различия между экспериментальными крысами линии WAG/Rij (5.7 ± 0.4 день) и Wistar (5.1 ± 0.5 день), которые потребляли этосуксимид, были незначимыми (F(1, 13) = 0.59, р = 0.46); H(1, 15) = 0.69, p = 0.41) (рис. 1a). При обучении день достижения критерия обученности у экспериментальной группы крыс линии WAG/Rij не отличался от данного показателя у контрольной группы крыс Wistar (F(1, 14) = 1.09, р = 0.26); H(1, 15) = 0.69, p = 0.41). Препарат не оказывал значимого влияния на день достижения критерия обученности у крыс Wistar (F(1, 13) = 2.37, р = 0.15); (H(1, 15) = 1.92, р = 0.16) (рис. 1a).
Для среднего времени реакции значимым по данным двухфакторного дисперсионного анализа являлись оба фактора – “возраст” (F (1, 30) = 57.2, р < 0.001) и “линия крыс” (F (1, 30) = 18.1, p < 0.001). Статистически значимых отличий данного показателя не было установлено у 2-месячных крыс линии WAG/Rij по сравнению с крысами Wistar аналогичного возраста (F(1, 15) = 3.8, p = 0.07) (рис.1b). В возрасте 6-ти месяцев наблюдали статистически значимые различия между крысами линии WAG/Rij и крысами Wistar (F(1, 15) = 66.4, p < 0.001): показатель “латентный период взятия подкрепления” был больше у крыс линии WAG/Rij (79.1 ± 4.0 c), чем у крыс Wistar (32.1 ± 4.2 с) (рис. 1b). У 2-месячных крыс Wistar (29.9 ± 4.2 с) и 2‑месячных крыс линии WAG/Rij (18.3 ± 4.0 c) среднее время реакции было меньше, чем у 6-месячных крыс Wistar (32.1 ± 4.2 с) и 6-месячных крыс линии WAG/Rij (79.1 ± 4.0 c) соответственно (рис. 1b). Различия по данному показателю между 2‑месячными и 6-месячными животными достигали уровня статистической значимости только у крыс линии WAG/Rij (F(1, 16) = 185.7, p < 0.001), но не у крыс Wistar (F(1, 14) = 0.09, p = 0.77) (рис. 1b).
У крыс линии WAG/Rij с этосуксимидом (31.0 ± 4.1 с) по сравнению с контрольными животными той же линии (79.1 ± 4.0 c) показатель “латентный период взятия подкрепления” был меньше (F (1, 15) = 51.5, р < 0.001). В отличие от существенных различий по данному показателю между крысами линии WAG/Rij и крысами Wistar, потреблявшими воду, значимых различий между крысами линии WAG/Rij (31.0 ± 4.1 с) и крысами Wistar (32.7 ± 4.4 с), потреблявшими этосуксимид, не наблюдали (F (1, 13) = = 0.19, р = 0.67) (рис.1b). Препарат не оказывал значимого влияния на среднее время реакции у крыс Wistar (F (1, 13) = 0.36, р = 0.56) (рис. 1b).
При выполнении обратной задачи, то есть при переделке навыка отыскания подкрепления в сложном лабиринте, животные демонстрировали похожий на прямую задачу паттерн поведения. Для дня достижения критерия обученности показана высокая значимость обоих факторов – “возраст” (F(1, 30) = 6.5, p < 0.05) и “линия крыс” (F(1, 30) = 6.4, p < 0.05). По данным однофакторного дисперсионного анализа по данному показателю между крысами линии WAG/Rij и крысами Wistar различия в возрасте 2-х месяцев отсутствуют (F(1, 15) = 1.13, p = 0.30). Отсутствие межлинейных различий по показателю “день достижения критерия обученности” у 2-месячных животных подтверждает однофакторный дисперсионный анализ Краскела–Уоллиса (H(1, 17) =1.12, p = 0.29). Однако в возрасте 6-ти месяцев скорость переучивания у крыс линии WAG/Rij была значимо меньше по сравнению с крысами Wistar согласно данным однофакторного дисперсионного анализа (F(1, 15) = 10.3, p < 0.01). Односторонний дисперсионный анализ Краскела–Уоллиса также свидетельствует о межлинейных различиях по данному показателю в возрасте 6-ти месяцев (H(1, 17) = 5.81, p < 0.05) (рис. 1с). 2-месячные крысы линии WAG/Rij (2.0 ± 0.1 день) переучивались быстрее, чем 6-месячные крысы линии WAG/Rij (2.7 ± 0.2 день), и различия по показателю “день достижения критерия обученности” между 2-месячными и 6-месячными животными данной линии достигали уровня статистической значимости (F(1, 16) = 8.0, p = 0.01); (H(1, 18) = 6.43, p < 0.001) (рис. 1с). Между 2-месячными (2.1 ± 0.1) и 6-месячными крысами Wistar (2.0 ± 0.1) не было значимых отличий по этому показателю (F(1, 14) = 1.00, p = 0.33); (H(1, 16) =1.00, p = 0.32) (рис. 1с).
При выполнении обратной задачи крысы линии WAG/Rij, которые потребляли этосуксимид, переучивались быстрее, чем крысы линии WAG/Rij, которые потребляли воду (F (1, 15) = 23.2, р < 0.001) (рис. 1с). Данные однофакторного дисперсионного анализа Краскела–Уоллиса подтверждают различия между экспериментальной и контрольной группой крыс линии WAG/Rij (H(2, 20) = 9.01, p < 0.05). Не наблюдали значимых различий у крыс линии WAG/Rij с этосуксимидом по сравнению с аналогичной группой крыс Wistar (F (1, 13) = 0.24, р = 0.63); (H(2, 20) =9.02, p = 0.01) (рис. 1с). При переучивании день достижения критерия обученности у экспериментальной группы крыс линии WAG/Rij не отличался от данного показателя у контрольной группы крыс Wistar (F (1, 14) = 1.09, р = 0.26); H(1, 15) =0.25, p = 0.62). Прием препарата не оказал значимого влияния на день достижения критерия обученности у экспериментальной группы крыс Wistar по сравнению с контрольной (F (1, 13) = 1.16, р = = 0.30); H(1, 15) =1.14, p = 0.28) (рис. 1с).
Двухфакторный дисперсионный анализ показал, что факторы “линия крыс” (F(1, 30) = 0.162, p > 0.05) и “возраст” (F(1, 30) = 3.2, p > 0.05) не оказывали статистически значимого влияния на латентный период взятия подкрепления при переучивании. Статистически значимых различий по данному показателю не было обнаружено у крыс линии WAG/Rij (26.8 ± 7.2 с) по сравнению с крысами Wistar (27.4 ± 7.7 с) в возрасте 2 мес. (F(1, 15) = 0.001, p = 0.97) (рис. 1d). Различий по среднему времени реакции не было обнаружено также у 6-месячных животных по сравнению с 2-месячными: (F(1, 16) = 1.90, p = 0.19) – для крыс линии WAG/Rij, (F(1, 14) = 2.11, p = = 0.17) – для крыс Wistar (рис. 1d). При этом в возрасте 6-ти месяцев, по данным двухфакторного дисперсионного анализа, этот показатель у контрольной группы крыс линии WAG/Rij по сравнению с контрольной группой крыс Wistar был больше. Однако это различие проявлялось на уровне тенденции, не достигая уровня статистической значимости, и post hoc сравнение по критерию Ньюмана–Кеулса подтвердило отсутствие статистической значимости (p = 0.07). Однако однофакторный дисперсионный анализ выявил факт наличия статистической значимости, и латентный период взятия подкрепления у контрольной группы крыс линии WAG/Rij (43.9 ± 7.2 с) по сравнению с контрольной группой крыс Wistar (16.4 ± 7.7 с) был значимо больше (F(1, 15) = 50.7, p < 0.01) (рис. 1d).
При переделке навыка отыскания подкрепления в сложном лабиринте (обратная задача) у крыс линии WAG/Rij, которые потребляли этосуксимид, среднее время реакции было меньше по сравнению с контрольной группой (F (1, 15) = 71.2, р < 0.001) (рис. 1d). В отличие от значимых различий по данному показателю между крысами линии WAG/Rij (43.9 ± 7.3 с) и Wistar (16.4 ± 7.7 с), потреблявших воду, различия между крысами линии WAG/Rij (16.7 ± 2.3 с) и Wistar (16.4±7.7 с), потреблявших этосуксимид, значимые различия отсутствовали (F (1, 13) = 0.31, р = 0.59) (рис. 1d). Препарат не оказал значимого влияния на скорость выполнения обратной задачи у крыс Wistar (F (1, 13) = 0.40, р = 0.53) (рис. 1d).
Латентный период взятия пищевого подкрепления определяется общим временем, которое крысы проводят в экспериментальной установке “сложный лабиринт”. С общим временем, которое крысы проводят в лабиринте, связаны такие показатели, как число стоек и число ошибок, которые могут говорить об “отвлечении” животных от решения задачи – получение пищевого подкрепления в целевом отсеке лабиринта. Число ошибок (F(1, 15) = 0.08, p = 0.77) и число стоек (F(1, 15) = 0.40, p = 0.54) были практически одинаковыми у крыс линии WAG/Rij и крыс Wistar в возрасте 2-х мес., так же как и латентный период взятия подкрепления (прямая и обратная задачи) (рис. 2a, b). По данным однофакторного дисперсионного анализа Краскела–Уоллиса также отсутствуют межлинейные различия у 2-месячных животных по числу ошибок (H(1, 17) = 0.04, p = 0.83). Однако в возрасте 6-ти мес. как число ошибок (F(1, 15) = 41.3, p < 0.001), так и число стоек (F(1, 15) = 15.1, p < 0.01) у крыс линии WAG/Rij были больше по сравнению с крысами Wistar аналогичного возраста (рис. 2a, b). Однофакторный дисперсионный анализ Краскела–Уоллиса подтверждает межлинейные различия по числу стоек в возрасте 6-ти месяцев (H(1, 17) =11.89, p < 0.001). У крыс линии WAG/Rij число стоек в возрасте 6-ти мес. статистически значимо не отличалось от данного показателя в возрасте 2-х мес. (F(1, 16) = 0.43, p = 0.52 (рис. 2a). У крыс Wistar в возрасте 6-ти мес. число стоек было меньше, чем в 2-месячном возрасте (F(1, 14) = 13.2, p = 0.003) (рис. 2a). Число ошибок у 6-месячных крыс линии WAG/Rij было больше по сравнению с 2‑месячными животными той же линии (F(1, 16) = 13.2, p < 0.01); (H(1, 18) = 7.65, p < 0.01) (рис. 2b). У крыс Wistar число ошибок было меньше в возрасте 6-ти мес. по сравнению с 2-месячным возрастом, однако это различие не достигало уровня статистической значимости (F(1, 14) = 2.61, p = 0.13); (H(1, 16) =1.97, p = 0.16) (рис. 2 b). У крыс Wistar не наблюдали возрастных различий по числу стоек (F(1, 14) = 2.61, p = 0.13).
Рис. 2.
Число стоек (a), ошибок (b) и коротких эпизодов груминга (c) в сложном лабиринте. WS – крысы Wistar, потреблявшие воду (контрольная группа), WR – крысы линии WAG/Rij, потреблявшие воду (контрольная группа), WS-ETX – 6-месячные крысы Wistar, потреблявшие ETX (экспериментальная группа), WR-ETX – 6-месячные крысы линии WAG/Rij, потреблявшие ETX (экспериментальная группа). ETX – этосуксимид (300 мг/кг/день, в течение 17 дней). * р < 0.05, *** р < 0.001 – у 6-месячных WR по сравнению с WS того же возраста. xxр < 0.01 – у 6-месячных крыс по сравнению с 2-месячными. ###р < 0.001 – у экспериментальной группы WR по сравнению с контрольной.

У крыс линии WAG/Rij с этосуксимидом число стоек было значимо меньше по сравнению с контрольной группой (F (1, 15) = 79.2, р < 0.001); по сравнению с крысами Wistar, которые потребляли этосуксимид, у экспериментальной группы крыс линии WAG/Rij отличий не было (F (1, 13) = 0.55, р = 0.52) (рис. 2a). Между группами крыс Wistar, которые потребляли антиабсансный препарат, и крысами Wistar, которые потребляли воду, различий по числу стоек не обнаружено (F (1, 13) = 0.74, р = 0.41) (рис. 2a).
У группы крыс линии WAG/Rij, которые потребляли этосуксимид, число ошибок в сложном лабиринте было меньше по сравнению с контрольными животными той же линии (F (1, 15) = 33.9, р < 0.001). Данные однофакторного дисперсионного анализа Краскела–Уоллиса подтверждают различия между экспериментальной и контрольной группой крыс линии WAG/Rij (H(2, 20) = 13.06, p < 0.01). В отличие от различий между контрольными группами крыс линии WAG/Rij и крыс Wistar различия между экспериментальными группами крыс линии WAG/Rij и крыс Wistar, отсутствовали (F (1, 13) = 0.16, р = 0.70); (H(1, 15) =0.11, p = 0.72) (рис. 2b). Препарат не оказывал значимого влияния на число ошибок у крыс Wistar (F (1, 13) = 0.91, р = 0.40); H(1, 15) = 0.17, p = 0.68) (рис. 2b).
Число коротких эпизодов груминга у крыс линии WAG/Rij было значимо больше, чем у крыс Wistar как в возрасте 2-х месяцев (F(1, 15) = 5.62, p = 0.03), так и в возрасте 6-ти мес. (F(1, 15) = 42.4, p < 0.001) (рис. 2c). Число коротких эпизодов груминга у 6-месячных крыс линии WAG/Rij было больше, чем у 2-месячных животных этой линии (F(1, 16) = 12.2, p < 0.01) (рис. 2c). У крыс Wistar возрастных различий по данному показателю не выявлено (F(1, 14) = 0.21, p = 0.65) (рис. 2c).
У крыс линии WAG/Rij, которые принимали этосуксимид, число коротких эпизодов груминга было значимо меньше (F (1, 15) = 52.7, р < 0.001), чем у животных той же линии, потреблявших воду (рис. 2c). Между крысами линии WAG/Rij и крысами Wistar, потреблявшими этосуксимид, значимые различия по данному показателю отсутствовали (F (1, 13) = 0.83, р = 0.38) (рис. 2c). Препарат не оказывал значимого влияния на число коротких эпизодов груминга у крыс Wistar (F (1, 13) = 0.91, р = 0.40) (рис. 2c).
Cреднее время реакции (латентный период взятия подкрепления) при обучении у крыс линии WAG/Rij положительно коррелировало с числом стоек (r = 0.67, p < 0.05) (рис. 3a) и числом ошибок (r = 0.76, p < 0.05) (рис. 2b). При переучивании также были обнаружены значимые положительные корреляции между средним временем реакции и числом стоек (r = 0.58, p < 0.05) (рис. 3c) и числом ошибок (r = 0.65, p < 0.05) (рис. 3d).
Тест на тревожность и пищевую мотивацию
Рис. 3.
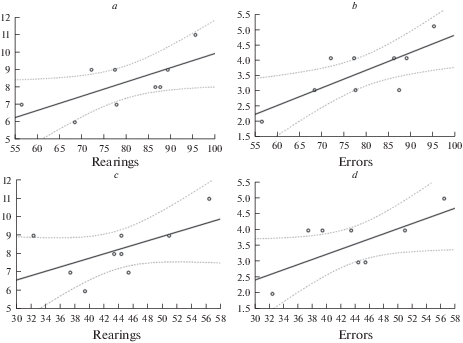
Связь между латентным периодом взятия пищевого подкрепления и числом стоек и ошибок в сложном лабиринте. По оси абсцисс – латентный период взятия подкрепления при обучении (a, b) и переучивании (c, d), по оси ординат – число стоек (a, c) и ошибок (b, d). Коэффициенты корреляции Пирсона: (a) – r = 0.67, p < 0.05. (b) – r = 0.76, p < 0.05. (c) – r = 0.58, p < 0.05. (d) – r = 0.65, p < 0.05.
Латентный период нахождения пищи в условиях новой обстановки является показателем, отражающим конфликт пищевой и оборонительной мотивации животных. Для этого показателя значимым по данным двухфакторного дисперсионного анализа являлся фактор “возраст” (F (1, 30) = 7.52, р = 0.01), незначимым – фактор “линия крыс” (F (1, 30) = 3.03, p = 0.09). В возрасте 2-х мес. у крыс линии WAG/Rij данный показатель (186.6 ± 32.3 с) статистически значимо не отличался от аналогичного показателя у крыс Wistar (172.6 ± 34.3 с) (F(1, 15) = 0.04, p = 0.84) (рис. 4a). В возрасте 6-ти мес. латентный период нахождения пищевого подкрепления у крыс линии WAG/Rij (139.2 ± 32.3 с) был больше по сравнению с крысами Wistar аналогичного возраста (36.4 ± 34.3 с) (F(1, 15) = 78.1, p < 0.001) (рис. 4a). Данный показатель у 6-месячных крыс линии Wistar был меньше, чем у 2-месячных крыс этой же линии (F(1, 14)=16.8, p < 0.01) (рис. 4a). У крыс линии WAG/Rij различий между 6-месячными и 2-месячными животными по латентному периоду нахождения пищевого подкрепления не обнаружено (F(1, 16) = 0.71, p = 0.41) (рис. 4a).
Рис. 4.
Латентный период подхода к пище в незнакомом открытом поле (a) и количество пищи, съеденное в домашней клетке (b) в тесте NSF. WS - крысы Wistar, потреблявшие воду (контрольная группа), WR – крысы линии WAG/Rij, потреблявшие воду (контрольная группа), WS-ETX – 6-месячные крысы Wistar, потреблявшие ETX (экспериментальная группа), WR-ETX – 6-месячные крысы линии WAG/Rij, потреблявшие ETX (экспериментальная группа). *** p < 0.001 – у WR по сравнению с WS. xxр < 0.01, xxxр < 0.001 – у 6-месячных крыс по сравнению с 2-месячными. ###р < 0.001 – у опытной группы WR по сравнению с контрольной.

У крыс линии WAG/Rij, которые потребляли этосуксимид (28.9 ± 7.5 с), латентный период достижения пищевого подкрепления был меньше (F (1, 15) = 97.3, p < 0.001), чем у животных той же линии, которые потребляли воду, и не отличался от такового у экспериментальной группы крыс Wistar (42.1 ± 8.0 с) (F (1, 13) = 2.11, р = 0.17) (рис. 4a). Между группами крыс Wistar, которые потребляли препарат, и крысами Wistar, потреблявшими воду, различий по латентному периоду взятия подкрепления не выявлено (F (1, 13) = 0.34, р = 057) (рис. 4a).
Количество корма, съеденное в домашней клетке, отражает собственно пищевую мотивацию животных. Для веса съеденного корма по данным двухфакторного дисперсионного анализа незначимыми являются как фактор “возраст” (F (1, 30) = = 2.59, р = 0.12), так и фактор “линия крыс” (F (1, 30) = 0.004, p = 0.95). Примечательно, что в возрасте 2-х месяцев этот показатель у крыс линии WAG/Rij (3.7 ± 0.2 г) был больше по сравнению с крысами Wistar аналогичного возраста (1.9 ± 0.2 г) (F(1, 15) = 30.5, p < 0.001) (рис. 4b). В возрасте 6 месяцев, напротив, количество корма, съеденное в домашней клетке, у крыс линии WAG/Rij (2.3 ± 0.2 г) было меньше по сравнению с крысами Wistar аналогичного возраста (4.0 ± 0.2 г) (F(1, 15) = = 41.8, p < 0.001) (рис. 4b). Вес съеденного корма у 6-месячных крыс Wistar был больше, чем у 2-месячных животных той же линии (F(1, 14) = 42.1, p < 0.001). И, напротив, данный показатель у 6-месячных крыс линии WAG/Rij был меньше, чем у 2-месячных крыс той же линии (F(1, 16) = 27.6, p < 0.001) (рис. 4b).
У крыс линии WAG/Rij, которые принимали этосуксимид (3.5 ± 0.2 г), вес съеденного пищевого подкрепления был больше (F (1, 15) = 21.9, p < 0.001), чем у контрольных животных той же линии (2.3 ± 0.2 г) (рис. 4b). Отличий от крыс Wistar, которые принимали этосуксимид (F (1, 13) = 0.43, р = 0.52), не обнаружено. Между экспериментальной (3.3 ± 0.2 г) и контрольной (4.0 ± 0.2 г) группами крыс Wistar различий по данному показателю также не выявлено (F (1, 13) = 2.11, р = 0.07) (рис. 4b).
Обнаружены значимые отрицательные корреляции между средним временем реакции (латентным периодом взятия пищевого подкрепления) при выполнении обратной задачи в сложном лабиринте и количеством пищи, потребляемой в домашней клетке в тесте NSF (r = 0.83, p < 0.05) (рис. 5a). Установлена положительная корреляция между днем достижения критерия обученности при выполнении прямой задачи в сложном лабиринте и латентным периодом нахождения пищи в тесте NSF (r = 0.46, p < 0.05) (рис. 5b).
Рис. 5.
(a) – связь между латентным периодом взятия подкрепления при выполнении обратной задачи в сложном лабиринте и количеством пищи, потребляемой в домашней клетке в тесте NSF. По оси абсцисс – латентный период взятия пищевого подкрепления при переучивании, по оси ординат – количество пищи, потребляемой в домашней клетке. (b) – связь между днем достижения критерия обученности при выполнении прямой задачи в сложном лабиринте и латентным периодом подхода к пище в условиях новой обстановки в тесте NSF. По оси абсцисс – день достижения критерия обученности при выполнении прямой задачи в сложном лабиринте, по оси ординат – латентный период подхода к пище в незнакомом открытом поле. Коэффициенты корреляции Пирсона: (a) – r = 0.83, p < 0.05. (b) – r = 0.46, p < 0.05.

ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
При выполнении прямой и обратной задачи в сложном лабиринте крысы линии WAG/Rij в возрасте 2-х мес. не отличались от крыс Wistar аналогичного возраста ни по дню достижения критерия обученности, ни по латентному периоду достижения целевого отсека камеры. Эти данные согласуется с данными, полученными в нашей предыдущей работе. Так, в тесте распознавания нового объекта у 2-месячных крыс линии WAG/Rij по сравнению с крысами Wistar аналогичного возраста нарушения эпизодической памяти отсутствовали: индекс распознавания нового объекта у крыс линии WAG/Rij значимо не отличался от аналогичного показателя у крыс Wistar [6, 7]. В возрасте 2-х месяцев у крыс линии WAG/Rij еще отсутствуют выраженные ПВР на ЭЭГ [26, 27]. Кроме того, ранее было показано, что в тесте принудительного плавания значимых различий по длительности иммобильности между 2-месячными крысами линии WAG/Rij и Wistar не выявлено, что говорит об отсутствии симптомов депрессии в поведении у крыс линии WAG/Rij в возрасте 2-х месяцев [13]. Таким образом, в том возрасте, когда у крыс линии WAG/Rij еще нет симптомов абсанс-эпилепсии [26, 27] и коморбидной депрессии [13], отсутствуют нарушения эпизодической памяти [6, 7], а также нарушения обучения и когнитивной гибкости, о чем свидетельствуют данные настоящей работы.
В настоящей работе нами получены данные, указывающие на наличие дефицита обучения и когнитивной гибкости, коморбидного депрессивно-подобного поведения, у 6-месячных крыс линии WAG/Rij. В этом возрасте при обучении навыку отыскания подкрепления в сложном лабиринте (прямая задача) крысы линии WAG/Rij в 1.8 раз медленнее достигали критерия обученности, чем крысы Wistar (рис. 1a). Время реакции (латентный период взятия подкрепления) при этом было в 2.5 раза больше у 6-месячных крыс линии WAG/Rij по сравнению с контрольными крысами Wistar (рис. 1b). С переделкой навыка (обратная задача) крысы линии WAG/Rij также справились хуже, чем крысы Wistar: крысы линии WAG/Rij в 1.4 раз медленнее достигали критерия обученности (рис. 1c) и в 2.7 раз дольше выполняли пищедобывательную реакцию по сравнению с крысами Wistar (рис. 1d). Эти данные согласуются с результатами, полученными ранее в аналогичных условиях эксперимента. Так, скорость обучения при выполнении прямой и обратной задачи в “сложном лабиринте” была значимо меньше, а время реакции, напротив, было значимо больше у крыс опытной группы (WAG/Rij) по сравнению с контрольной (Wistar) [28].
Сходные с крысами линии WAG/Rij нарушения исполнительных функций были обнаружены при инструментальном научении крыс линии GAERS распознаванию зрительных стимулов с пищевым подкреплением [34]. После того, как животные обучились прямой задаче – получать пищевое подкрепление в ответ на определенный визуальный стимул, начиналась фаза реверсивного обучения (переделки навыка), когда “правильный” и “неправильный” стимулы менялись местами. Было показано, что крысы линии GAERS медленнее обучались как прямой (только самцы), так и обратной задаче (и самцы, и самки) по сравнению с контролем (животные, у которых отсутствовали ПВР). Так, животным с абсансными ПВР требовалось больше времени на адаптацию к условиям эксперимента, они делали больше ошибок в тесте по сравнению с контролем [34]. Таким образом, можно говорить о том, что дефицит когнитивной гибкости, который демонстрируют крысы линии GAERS [34] и крысы линии WAG/Rij является характерной особенностью экспериментальных моделей абсанс-эпилепсии.
Обучение животных прямой и обратной задаче по данной методике также проводилось на каинатной модели височной эпилепсии у крыс Wistar. Однако даже по сравнению с крысами линии GAERS, животные, которые подвергались воздействию каиновой кислоты, менее успешно обучались как инструментальному навыку (прямая задача), так и его переделке (обратная задача); 60% из них вообще не достигали критерия обученности даже при обучении исходному навыку. Успех обучения (прямая задача) зависит от способности животного ассоциировать стимул с пищевым вознаграждением и помнить об этом в течение нескольких экспериментальных дней, успех переучивания (обратная задача) зависит от способности животного к торможению ранее выработанной реакции и быстрому научению новому инструментальному навыку [35].
У крыс линии WAG/Rij, которые потребляли этосуксимид, скорость достижения критерия обученности была больше (рис. 1a, c), а латентный период достижения целевого отсека камеры был меньше (рис. 1b, d) как при обучении навыку отыскания подкрепления в сложном лабиринте (прямая задача), так и при выполнении этой задачи в условиях зеркально перевернутых перегородок с отверстиями (обратная задача) по сравнению с контрольными животными той же линии. Другими словами, данный антиабсансный препарат улучшает выполнение как прямой, так и обратной задачи у крыс линии WAG/Rij. Это говорит о том, что этосуксимид положительно влияет на обучение и память, а также улучшает когнитивную гибкость – важный компонент исполнительных функций. Эти данные согласуются с результатами наших предыдущих работ, в которых было показано, что этосуксимид улучшает как кратковременную, так и долговременную эпизодическую память в тесте распознавания нового объекта [6, 7].
Такой показатель, как время реакции (латентный период взятия подкрепления), отражает скорость воспроизведения выученной реакции и определяется общим временем, которое крысы проводят в сложном лабиринте. С общим временем, проведенным животным в сложном лабиринте, связано число стоек и число ошибок. Они являются косвенным показателем внимания (избирательная направленность на достижение пищевого подкрепления, расположенного в целевом отсеке камеры) и могут говорить об “отвлечении” животных от выполнения задачи. Другими словами, чем больше латентный период взятия подкрепления – общее время, за которое крысы проходят лабиринт, тем больше они успевают выполнить стоек и тем больше совершают ошибок. Подобное предположение подтверждается корреляционным анализом: среднее время реакции при обучении у крыс линии WAG/Rij положительно коррелировало с числом стоек (r = 0.67, p < 0.05) (рис. 3a) и числом ошибок (r = 0.76, p < 0.05) (рис. 3b). При переучивании также были обнаружены значимые положительные корреляции между средним временем реакции и числом стоек (r = 0.58, p < 0.05) (рис. 3c), а также числом ошибок (r = 0.65, p < 0.05) (рис. 3d).
В данной работе было установлено отсутствие отличий числа стоек и ошибок в лабиринте между крысами линии WAG/Rij и крысами Wistar в возрасте 2-х мес. (рис. 2a, b). Это указывает на нормальное воспроизведение выученной реакции у крыс линии WAG/Rij в том возрасте, когда отсутствуют выраженная пик-волновая активность и признаки депрессивно-подобного поведения [13, 26, 27]. В возрасте 6-ти мес., когда пик-волновая активность и депрессивно-подобная симптоматика отчетливо выражена [1–5], число стоек и число ошибок было больше у крыс линии WAG/Rij по сравнению с крысами Wistar, у которых отсутствовала какая-либо патология (рис. 2a, b). Возможно, что большее по сравнению с “нормальными” крысами Wistar число “отвлечений” у крыс линии WAG/Rij может рассматриваться как показатель пониженной способности к концентрации внимания.
Прием этосуксимида значимо уменьшал как число стоек, так и число ошибок у крыс линии WAG/Rij по сравнению с животными той же линии, которые получали воду без препарата (рис. 2a, b). Это указывает на то, что антиабсансный препарат, по всей видимости, уменьшал отвлекаемость при решении задачи, то есть улучшал внимание у крыс линии WAG/Rij. Следствием уменьшения отвлекаемости при выполнении пищедобывательной реакции, вероятнее всего, стало уменьшение общего времени, проведенного животными в сложном лабиринте, и действительно, латентный период взятия подкрепления у экспериментальной группы крыс линии WAG/Rij был меньше по сравнению с контрольной группой (рис. 1c, d).
В ранее проведенном исследовании в аналогичных условиях эксперимента – “сложный лабиринт с пищевым подкреплением” – число ошибок и число стоек также были значимо больше у крыс линии WAG/Rij по сравнению с крысами Wistar [28]. С этими данными согласуются результаты ранее проведенных нами исследований распознавания нового объекта [6, 7] и социального распознавания [7] у крыс линии WAG/Rij. Число посещений пустой части арены и полный пройденный путь во время предъявления объектов для распознавания рассматривались нами как показатели “отвлечения” от исследования объектов, что было интерпретировано как дефицит внимания – направленной активности животного на исследование предъявляемых объектов. В тесте распознавания нового объекта и социального распознавания также было установлено отсутствие избирательной направленности на исследование незнакомого объекта у крыс линии WAG/Rij по сравнению с крысами Wistar, независимо от природы объекта – одушевленный (тест социального распознавания) или неодушевленный (тест распознавания нового объекта). Это также указывает на дефицит внимания у крыс линии WAG/Rij, который, возможно, является одним из поведенческих симптомов депрессии, обусловленный дисфункцией префронтальной коры [36].
Мы предполагаем, что пониженная способность к концентрации внимания (дефицит внимания) у крыс линии WAG/Rij по сравнению с крысами Wistar может быть связан с низким уровнем пищевой мотивации. Чтобы проверить эту гипотезу, мы провели тест NSF на тревожность и пищевую мотивацию. Вес съеденного корма – количество корма, съеденного в домашней клетке, характеризует именно уровень пищевой мотивации [28, 29, 33]. Установлено, что в 2 мес. вес съеденного корма у крыс линии WAG/Rij был больше по сравнению с крысами Wistar, в то время как в 6 мес. данный показатель, напротив, был меньше у крыс линии WAG/Rij по сравнению с крысами Wistar (рис. 4b). В возрасте 6-ти мес. более низкое по сравнению с “нормальными” крысами потребление пищи в домашней клетке у крыс линии WAG/Rij отражает низкий уровень пищевой мотивации. Данные корреляционного анализа подтверждают важность вклада уровня пищевой мотивации в успешное выполнение прямой и обратной задачи в сложном лабиринте. Так, обнаружены значимые отрицательные корреляции между средним временем реакции при выполнении обратной задачи в сложном лабиринте и количеством пищи, потребляемой в домашней клетке в тесте NSF (r = 0.83, p < 0.05) (рис. 5a).
Помещение животного в новое для него “Открытое поле” приводит к конфликту двух мотиваций: пищевой мотивации, которая актуализируется на фоне пищевой депривации, предшествующей проведению теста NSF, и мотивации страха (тревоги), вызываемой умеренным стрессогенным воздействием открытого незнакомого и хорошо освещенного пространства, в центре которого расположено пищевое подкрепление. Скорость, с которой животное достигнет этого пищевого подкрепления, зависит от того, какая из мотиваций является преобладающей: пищевая мотивация или страх, и она оценивается в данном тесте по показателю “латентный период нахождения пищи в условиях новой обстановки” [28, 29, 33]. В возрасте 2-х мес. по данному показателю у крыс линии WAG/Rij по сравнению с крысами Wistar не было различий. Однако в возрасте 6-ти мес. латентный период достижения пищевого подкрепления у крыс линии WAG/Rij был больше по сравнению с крысами Wistar аналогичного возраста (рис. 4a). Это свидетельствует либо о сдвиге мотивационного фона у крыс линии WAG/Rij в сторону мотивации страха (что означает повышенную тревожность), либо о пониженной пищевой мотивации, которая является одним из проявлений депрессивного состояния.
На повышенную тревожность во время выполнения задач в сложном лабиринте и тесте NSF указывает повышенное число коротких эпизодов груминга при обучении в сложном лабиринте (рис. 2с). Принято считать, что короткие (неполные) реакции груминга отражают уровень тревожности [37]. Однако различий между 6-месячными крысами линии WAG/Rij и крысами Wistar аналогичного возраста в уровне тревожности не было обнаружено ни в одном из тестов, оценивающих тревожность, – свето-темнового выбора, социального взаимодействия и приподнятого крестообразного лабиринта [1]. Повышенный по сравнению с крысами Wistar уровень тревожности в тестах свето-темнового выбора и приподнятого крестообразного лабиринта проявляли только крысы линии WAG/Rij, чувствительные к аудиогенным припадкам [1, 38]. Повышенный уровень тревожности в тестах на тревожность, таких как тест “Свето-темновой выбор” и тест “Открытое поле”, был установлен также у крыс линии WAG/Rij по сравнению с крысами Wistar только в возрасте 2-х мес. [13].
На пониженную пищевую мотивацию у крыс линии WAG/Rij указывают: 1) более низкое по сравнению с крысами Wistar количество корма, съеденного в домашней клетке (рис. 4b); 2) отрицательные корреляции между средним временем реакции при реверсивном обучении в сложном лабиринте и количеством пищи, потребляемой в домашней клетке в тесте NSF (рис. 5a). Интересно также, что нами была обнаружена положительная корреляция между латентным периодом нахождения пищи в тесте NSF и днем достижения критерия обученности при выполнении прямой задачи в сложном лабиринте (r = 0.46, p < 0.05) (рис. 5b). Таким образом, чем ниже уровень пищевой мотивации, тем хуже животные обучаются в сложном лабиринте. Существенно, что в возрасте 2-х мес. у крыс линии WAG/Rij наблюдали более высокое по сравнению с контролем потребление пищи в домашней клетке. Это, вероятно, отражает высокий уровень тревожности 2-месячных животных данной линии, что подтверждают ранее полученные данные [13].
Количество пищи, потребленной в домашней клетке, было больше (рис. 5b), в то время как латентный период ее нахождения в “Открытом поле” был меньше (рис. 5a) у экспериментальной группы крыс линии WAG/Rij, потреблявших этосуксимид, по сравнению с контрольной группой крыс той же линии. У крыс линии WAG/Rij, которые потребляли этосуксимид, число коротких эпизодов груминга было значимо меньше, чем у животных той же линии, которые потребляли воду без препарата (рис. 2c). Это указывает на то, что этосуксимид увеличивал уровень пищевой мотивации у крыс линии WAG/Rij, то есть способствовал сдвигу мотивационного фона у крыс линии WAG/Rij в сторону преобладания пищевой мотивации над мотивацией страха.
Отсутствие влияния этосуксимида на обучение, переучивание, внимание и пищевую мотивацию у “нормальных” крыс Wistar (рис. 1, 2, 4) может свидетельствовать о том, что данный препарат не обладает собственным влиянием на эти показатели, и он улучшает их у крыс линии WAG/Rij только через подавление пик-волновой активности.
Как у пациентов с эпилепсией, так и у больных депрессией, наблюдают гипофункцию дофаминергической системы мозга, которая играет ключевую роль в механизмах подкрепления, обучения и памяти [39]. Депрессивно-подобное поведение, характерное для крыс линии WAG/Rij (пониженная исследовательская активность в тесте открытого поля, повышенная иммобильность в тесте вынужденного плавания, пониженное потребление и предпочтение раствора сахарозы) [1–3, 5], имеет дофамин-зависимый характер, что подтверждено фармакологическим анализом [2].
Реверсивное обучение в сложном лабиринте, использованное в данной работе, является дофамин-зависимым процессом. С одной стороны, дофамин опосредует синаптическую пластичность в тех областях мозга, которые вовлечены в реверсивное обучение (орбитофронтальная кора, стриатум, прилежащее ядро и миндалина) [40]. Обучение и переучивание навыка выполнения пространственной задачи с положительным подкреплением применялось нами для оценки когнитивной гибкости (cognitive flexibility) – ключевого компонента исполнительных функций. Когнитивная гибкость проявляется в данном случае в том, насколько быстро животное способно затормозить навык прохождения сложного лабиринта по определенной траектории, заданной расположением отверстий в перегородках, и насколько быстро животное способно приобрести навык проходить этот лабиринт по новой траектории, заданной зеркально перевернутыми перегородками с отверстиями. Ожидание вознаграждения, пищевого подкрепления, с которым животное познакомилось во время выполнения прямой задачи, и его прогнозирование являются ключевыми факторами успешной переделки навыка прохождения сложного лабиринта, то есть выполнения обратной задачи. С помощью компьютерного анализа было обнаружено, что фазное выделение дофамина отображает несоответствие между ожидаемым и полученным результатом вознаграждения (ошибки прогнозирования вознаграждения (ОПВ), reward prediction errors (RPE)). Показано, что оптогенетическое возбуждение дофаминовых нейронов в вентральной области покрышки (ventral tegmental area, VTA) или компактной части (pars compacta) черной субстанции (Substantia nigra) (pcSn) среднего мозга улучшает эффективность пространственного реверсивного обучения [41, 42]. Результаты наблюдений за пациентами с синдромом дефицита внимания и гиперактивности, принимавшими метилфенидат (Риталин), показывают, что они лучше справляются с задачами на реверсивное обучение [43]. Напротив, фармакологическое истощение дофамина в стриатуме мармозеток приводило к грубым нарушениям выполнения обратной задачи при переделке навыка [44]. Методом быстрой циклической вольтамперометрии было зарегистрировано увеличение фазового выделения дофамина в прилежащем ядре (nucleus accumbens, NAcc) у крыс в ответ на предъявление неожиданного вознаграждения (положительная ошибка прогнозирования, positive prediction error) [45] Кроме этого, снижение уровня дофамина в NAcc у крыс вызывало нарушение реверсивного обучения [46]. Таким образом, из изложенного материала можно сделать вывод о том, что ключевую роль в контроле реверсивного обучения играют подкорковые структуры мозга, являющиеся частью мезо-кортико-лимбической дофаминергической системы. Так, фармакологическое истощение дофамина в орбитофронтальной коре не оказывает влияния на когнитивную гибкость у мармозеток [47]. Результаты, полученные в данной работе, – дефицит когнитивной гибкости и пищевой мотивации, повышенная “отвлекаемость” у 6-месячных крыс линии WAG/Rij, хорошо укладываются в существующие представления о недостаточности мезо-кортико-лимбической дофаминергической системы мозга крыс линии WAG/Rij [1, 2, 48]. В возрасте 2-х мес., когда фенотипические проявления абсанс-эпилепсии [26, 27] и депрессии [13] у крыс линии WAG/Rij отсутствуют, снижение уровня дофамина было выявлено только в префронтальной коре [13]. Вероятно, этих изменений было недостаточно, чтобы вызвать нарушения когнитивной гибкости, выявленное в настоящей работе, и эпизодической памяти, показанное в наших предыдущих работах [6, 7]. В возрасте 6-ти мес., когда пик-волновая активность и симптомы депрессии хорошо выражены, гипофункция мезо-кортико-лимбической дофаминергической системы мозга усиливается, что проявляется дефицитом дофамина в стриатуме и прилежащем ядре [48]. Вероятнее всего, что эти изменения у крыс линии WAG/Rij проявляются симптомами депрессии [1–3, 5], а также нарушениями обучения и когнитивной гибкости, установленными в данной работе, и эпизодической памяти, показанными в наших предыдущих работах [6, 7].
Таким образом, данные, полученные в настоящей работе, позволяют предположить наиболее вероятный сценарий развития дефицита когнитивной гибкости – ключевого компонента исполнительных когнитивных функций при реверсивном обучении в сложном лабиринте у крыс линии WAG/Rij. Нарушения когнитивной гибкости могут быть следствием пониженной пищевой мотивации как одного из симптомов депрессии, коморбидной абсанс-эпилепсии и обусловленной дефицитом мезо-кортико-лимбической дофаминергической системы, которая проявляется у крыс линии WAG/Rij в возрасте 6-ти мес.
Список литературы
Sarkisova K, van Luijtelaar G (2011) The WAG/Rij strain: a genetic animal model of absence epilepsy with comorbidity of depression. Prog Neuropsychopharmacol Biol Psychiatry 35(4): 854–876. https://doi.org/10.1016/j.pnpbp.2010.11.010
Sarkisova KYu, Kulikov MA, Midzianovskaia IS, Folomkina AA (2007) Dopamine-dependent character of depressive-like behavior in WAG/Rij rats with genetic absence epilepsy. Zh Vyssh Nerv Deiat Im IP Pavlova 57(1): 91–102. https://doi.org/10.1007/s11055-008-0017-z
Sarkisova KY, Midzyanovskaya IS, Kulikov MA (2003) Depressive-like behavioral alterations and c-fos expression in the dopaminergic brain regions in WAG/Rij rats with genetic absence epilepsy. Behav Brain Res 144: 211–226. https://doi.org/10.1016/s0166-4328(03)00090-1
van Luijtelaar G, Zobeiri M (2014) Progress and outlooks in a genetic absence epilepsy model (WAG/Rij). Curr Med Chem 21(6): 704–721. https://doi.org/10.2174/0929867320666131119152913
Sarkisova KYu, Kuznetsova GD, Kulikov MA, van Luijtelaar G (2010) Spike–wave discharges are necessary for the expression of behavioral depression-like symptoms. Epilepsia 51: 146–160. https://doi.org/10.1111/j.1528-1167.2009.02260.x
Leo A, Citraro R, Tallarico M, Iannone M, Fedosova E, Nesci V, De Sarro G, Sarkisova K, Russo E (2019) Cognitive impairment in the WAG/Rij rat absence model is secondary to absence seizures and depressive-like behavior. Prog Neuropsychopharmacol Biol Psychiatry 94: 109652. https://doi.org/10.1016/j.pnpbp.2019.109652
Fedosova EA, Shatskova AB, Sarkisova KY (2021) Ethosuximide increases exploratory motivation and improves episodic memory in the novel object recognition test in WAG/Rij rats with genetic absence epilepsy. Neurosci Behav Physiol 51(4): 501 – 512. https://doi.org/10.1007/s11055-021-01097-z
Jones NC, Salzberg MR, Kumar G, Couper A, Morris MJ, O’Brien TJ (2008) Elevated anxiety and depression-like behavior in a rat model of genetic generalized epilepsy, suggesting common causation. Exp Neurol 209: 254–260. https://doi.org/10.1016/j.expneurol.2007.09.026
Cheng D, Yan X, Gao Z, Xu K, Zhou X, Chen Q (2017) Common and distinctive patterns of cognitive dysfunction in children with benign epilepsy syndromes. Pediatr Neurol 72: 36–41. https://doi.org/10.1016/j.pediatrneurol.2016.12.005
Conradi HJ, Ormel J, de Jonge P (2011) Presence of individual (residual) symptoms during depressive episodes and periods of remission: a 3-year prospective study. Psychol Med 41(6): 1165–1174. https://doi.org/10.1017/S0033291710001911
Karson A, Utkan T, Balc F, Arıcıoglu F, Ates N (2012) Age-dependent decline in learning and memory performances of WAG/Rij rat model of absence epilepsy. Behav Brain Funct 8: 51. https://doi.org/10.1186/1744-9081-8-51
Inoue M, Peeters BWMM, van Luijtelaar ELJM, Vossen JMH, Coenen AML (1990) Spontaneous occurrence of spike-wave discharges in five inbred strains of rats. Physiology and Behavior 48(1): 199–201. https://doi.org/10.1016/0031-9384(90)90285-c
Fedosova EA, Sarkisova KYu, Kudrin VS, Narkevich VB, Bazyan AS (2015) Behavioral and Neurochemical Characteristics of Two Months Old WAG/Rij Rats with Genetic Absence Epilepsy. Int J Clin Exp Neurol 3(2): 32–44. https://doi.org/10.12691/ijcen-3-2-2
Hammar A, Ardal G (2009) Cognitive functioning in major depression – a summary. Front Hum Neurosci 3: 26. https://doi.org/10.3389/neuro.09.026.2009
Yang T, Zhao G, Mao R, Huang J, Xu X, Su Y, Zhu N, Zhou R, Lin X, Xia W, Wang F, Liu R, Wang X, Huang Z, Wang Y, Hu Y, Cao L, Yuan S, Wang Z, Lam RW, Chen J, Fang Y (2018) The association of duration and severity of disease with executive function: Differences between drug-naïve patients with bipolar and unipolar depression. J Affective Disorders 238: 412–417. https://doi.org/10.1016/j.jad.2018.05.051
Izquierdo A, Brigman J, Radke A, Rudebeck P, Holmes A (2016) The neural basis of reversal learning: An updated perspective. Neuroscience 345: 12–26. https://doi.org/10.1016/j.neuroscience.2016.03.021
Ghahremani DG, Monterosso J, Jentsch JD, Bilder RM, Poldrack RA (2010) Neural components underlying behavioral flexibility in human reversal learning. Cereb Cortex 20: 1843–1852. https://doi.org/10.1093/cercor/bhp247
Machado CJ, Bachevalier J (2007) The effects of selective amygdala, orbital frontal cortex or hippocampal formation lesions on reward assessment in nonhuman primates. Eur J Neurosci 25: 2885–2904. https://doi.org/10.1111/j.1460-9568.2007.05525.x
Chudasama Y, Daniels TE, Gorrin DP, Rhodes SE, Rudebeck PH, Murray EA (2013) The role of the anterior cingulate cortex in choices based on reward value and reward contingency. Cereb Cortex 23: 2884–2898. https://doi.org/10.1093/cercor/bhs266
Churchwell JC, Morris AM, Heurtelou NM, Kesner RP (2009) Interactions between the prefrontal cortex and amygdala during delay discounting and reversal. Behav Neurosci 123: 1185–1196. https://doi.org/10.1037/a0017734
Volkow ND, Fowler JS (2000) Addiction, a Disease of Compulsion and Drive: Involvement of the Orbitofrontal Cortex. Cereb Cortex 10 (3): 318–325. https://doi.org/10.1093/cercor/10.3.318
Braun S, Hauber W (2011) The dorsomedial striatum mediates flexible choice behavior in spatial tasks. Behav Brain Res 220 (2): 288–293. https://doi.org/10.1016/j.bbr.2011.02.008
Stern CE, Passingham RE (1995) The nucleus accumbens in monkeys (Macaca fascicularis). Exp Brain Res 106: 239–247. https://doi.org/10.1007/BF00241119
Dalton GL, Phillips AG, Floresco SB (2014) Preferential Involvement by Nucleus Accumbens Shell in Mediating Probabilistic Learning and Reversal Shifts. J Neurosci 34(13): 4618–4626. https://doi.org/10.1007/s00213-011-2246-z
Wassum KM, Izquierdo A (2015) The basolateral amygdala in reward learning and addiction. Neurosci Biobehav Rev 57: 271–283. https://doi.org/10.1016/j.neubiorev.2015.08.017
Coenen AM, van Luijtelaar EL (1987) The WAG/Rij rat model for absence epilepsy: age and sex factors. Epilepsy Res 1: 297. https://doi.org/10.1016/0920-1211(87)90005-2
Gabova AV, Sarkisova KYu, Fedosova EA, Shatskova AB, Morozov AA (2020) Developmental Changes in Peak-Wave Discharges in WAG/Rij Rats with Genetic Absence Epilepsy. Neurosci Behav Physiol 50: 245–252. https://doi.org/10.1007/s11055-019-00893-y
Malyshev AV, Zakharov AM, Sarkisova KYu, Dubynin VA (2012) Reverse learning in WAG/Rij rats with depression-like behavior. Zh Vyssh Nerv Deiat Im IP Pavlova 62(5): 580–590. https://doi.org/10.1007/s11055-013-9869-y
Bessa JM, Mesquita AR, Oliveira M, Pêgo JM, Cerqueira JJ, Palha JA, Almeida OF, Sousa N (2009) A trans-dimensional approach to the behavioral aspects of depression. Front Behav Neurosci 3: 1–7. https://doi.org/10.3389/neuro.08.001.2009
Саркисова КЮ, Куликов МА, Фоломкина AA (2011) Оказывает ли антиабсансный препарат этосуксимид антидепрессантный эффект? Журн высш нервн деятельн им ИП Павлова 2: 227–235. [Sarkisova KJu, Kulikov MA, Folomkina AA (2011) Does antiabsence drug ethosuximide exert antidepressant effect? Zh Vyssh Nerv Deiat Im IP Pavlova 2: 227–235. (In Russ)].
Zhang K, Jia G, Xia L, Du J, Gai G, Wang Z, Cao L, Zhang F, Tao R, Liu H, Hashimoto K, Wang G (2021) Efficacy of anticonvulsant ethosuximide for major depressive disorder: a randomized, placebo-control clinical trial. Eur Arch Psychiatry Clin Neurosci 271(3): 487–493. https://doi.org/10.1007/s00406-020-01103-4
Blumenfeld H, Klein JP, Schridde U, Vestal M, Rice T, Khera DS, Bashyal C, Giblin K, Paul-Laughinghouse C, Wang F, Phadke A, Mission J, Agarwal RK, Englot DJ, Motelow J, Nersesyan H, Waxman SG, Levin AR (2008) Early treatment suppresses the development of spike-wave epilepsy in a rat model. Epilepsia 49(3): 400–409. https://doi.org/10.1111/j.1528-1167.2007.01458.x
Mazarati A, Jones NC, Galanopoulou AS, Harte-Hargrove LC, Kalynchuk LE, Lenck-Santini PP, Medel-Matus JS, Nehlig A, de la Prida LM, Sarkisova K, Veliskova J (2018) A companion to the preclinical common data elements on neurobehavioral comorbidities of epilepsy: a report of the TASK3 behavior working group of the ILAE/AES Joint Translational Task Force. Epilepsia Open 3(1): 24–52. https://doi.org/10.1002/epi4.12236
Roebuck A, An L, Marks W, Sun N, Snatch T, Howland J (2020) Cognitive Impairments in Touchscreen-based Visual Discrimination and Reversal Learning in Genetic Absence Epilepsy Rats from Strasbourg. Neuroscience 1(430): 105–112. https://doi.org/10.1016/j.neuroscience.2020.01.028
Carron S, Dezsi G, Ozturk E, Nithianantharajah J, Jones NC (2019) Cognitive deficits in a rat model of temporal lobe epilepsy using touchscreen-based translational tools. Epilepsia 60(8): 1650–1660. https://doi.org/10.1111/epi.16291
Clark L, Cools R, Robbins TW (2004) The neuropsychology of ventral prefrontal cortex: decision-making and reversal learning. Brain Cogn 55(1): 41–53. https://doi.org/10.1016/S0278-2626(03)00284-7
Kalueff AV, Tuohimaa P (2005) The grooming analyses algorithm discriminates between different levels of anxiety in rats: potential utility for neurobehavioural stress research. J Neurosci Methods 143(2): 69–77. https://doi.org/10.1016/j.jneumeth.2004.10.001
Sarkisova KYu, Kulikov MA (2006) Behavioral characteristics of WAG/Rij rats susceptible and non-susceptible to audiogenic seizures. Behav Brain Res 166: 9–18. https://doi.org/10.1016/j.bbr.2005.07.024
Shohamy D, Adcock R (2010) Dopamine and adaptive memory. Trends Cogn Sci 14: 464–472. https://doi.org/10.1016/j.tics.2010.08.002
Calabresi P, Picconi B, Tozzi A, Di Filippo M (2007) Dopamine mediated regulation of corticostriatal synaptic plasticity. Trends Neurosci 30: 211–219. https://doi.org/10.1016/j.tins.2007.03.001
Adamantidis AR, Tsai HC, Boutrel B, Zhang F, Stuber GD, Budygin EA (2011) Optogenetic interrogation of dopaminergic modulation of the multiple phases of reward-seeking behavior. J Neurosci 31: 10829–10835. https://doi.org/10.1523/JNEUROSCI.2246-11.2011
Rossi MA, Sukharnikova T, Hayrapetyan VY, Yang L, Yin HH (2013) Operant self-stimulation of dopamine neurons in the substantia nigra. PLoS One 8: e65799. https://doi.org/10.1371/journal.pone.0065799
Clatworthy PL, Lewis SJ, Brichard L, Hong YT, Izquierdo D, Clark L (2009) Dopamine release in dissociable striatal subregions predicts the different effects of oral methylphenidate on reversal learning and spatial working memory. J Neurosci 29: 4690–4696. https://doi.org/10.1523/JNEUROSCI.3266-08.2009
Clarke HF, Hill GJ, Robbins TW, Roberts AC (2011) Dopamine, but not serotonin, regulates reversal learning in the marmoset caudate nucleus. J Neurosci 31: 4290–4297. https://doi.org/10.1523/JNEUROSCI.5066-10.2011
Klanker M, Sandberg T, Joosten R, Willuhn I, Feenstra M, Denys D (2015) Phasic dopamine release induced by positive feedback predicts individual differences in reversal learning. Neurobiol Learn Mem 125: 135–145. https://doi.org/10.1016/j.nlm.2015.08.011
Taghzouti K, Louilot A, Herman JP, Le Moal M, Simon H (1985) Alteration behavior, spatial discrimination, and reversal disturbances following 6-hydroxy-dopamine lesions in the Nucleus accumbens of the rat. Behav Neural Biol 44: 354–363. https://doi.org/10.1016/s0163-1047(85)90640-5
Clarke HF, Walker SC, Dalley JW, Robbins TW, Roberts AC (2007) Cognitive inflexibility after prefrontal serotonin depletion is behaviorally and neurochemically specific. Cereb Cortex 17: 18–27. https://doi.org/10.1093/cercor/bhj120
Саркисова КЮ, Куликов МА, Кудрин ВС, Мидзяновская ИС, Бирюкова ЛМ (2014) Возрастные изменения в поведении, в содержании моноаминов, их метаболитов и в плотности D1 и D2 дофаминовых рецепторов в структурах мозга у крыс линии WAG/Rij с депрессивноподобной патологией. Журн высш нервн деятельн им ИП Павлова 64(6): 668– 685. [Sarkisova KJu, Kulikov MA, Kudrin VS, Midzyanovskaya IS, Birioukova LM (2014) Age-related changes in behavior, in monoamines and their metabolites content, and in density of D1 and D2 dopamine receptors in the brain structures of WAG/Rij rats with depression-like pathology. Zh Vyssh Nerv Deiat Im IP Pavlova 64(6): 668–685. (In Russ)]. https://doi.org/10.7868/S0044467714060094
Дополнительные материалы отсутствуют.
Инструменты
Российский физиологический журнал им. И.М. Сеченова