

Российский физиологический журнал им. И.М. Сеченова, 2022, T. 108, № 1, стр. 24-35
Методологические и вычислительные аспекты извлечения обширных мышечных синергий при локомоциях умеренной интенсивности
С. А. Моисеев 1, *, А. М. Пухов 1, Е. А. Михайлова 1, Р. М. Городничев 1
1 Великолукская государственная академия физической культуры и спорта
г. Великие Луки, Россия
* E-mail: sergey_moiseev@vlgafc.ru
Поступила в редакцию 10.09.2021
После доработки 08.10.2021
Принята к публикации 08.10.2021
- EDN: JPINGF
- DOI: 10.31857/S0869813922010101
Аннотация
Современные подходы к изучению двигательных синергий, как правило, включают рассмотрение синергетических эффектов на одном, реже двух уровнях организации нервной системы, и их взаимодействие в контексте решения разнообразных двигательных задач, в том числе локомоций. Однако даже при рассмотрении схожих по биомеханической структуре движений наблюдаются расхождения в некоторых аспектах изучения мышечных синергий, что объясняют применением разных методологических и вычислительных подходов, не учитывающих специфику рассматриваемых двигательных актов. Цель исследования заключалась в поиске оптимальных параметров предварительной обработки электромиографических сигналов и оценке их влияния на результаты извлечения обширных мышечных синергий в процессе локомоций умеренной интенсивности. Исследования выполнены на 10-ти спринтерах мужского пола, которые выполняли бег с умеренной интенсивностью на тредбане. Регистрировали электромиограммы 16‑ти поверхностных скелетных мышц туловища, верхних и нижних конечностей. Мышечные синергии извлекали с помощью методов матричной факторизации с применением различных процедур предобработки исходных сигналов. Установлено, что лучшие результаты по извлечению синергий дает применение полосового фильтра в диапазоне 20–450 Гц с использованием после выпрямления дополнительной фильтрации 20 Гц. Количество извлекаемых синергий из разных периодов цикла бегового шага оказалось больше при использовании метода главных компонент и меньше при факторном анализе, а по показателю объясняемой дисперсии оба метода давали лучшие результаты, чем при рассмотрении целого цикла бегового шага. Количество синергий пропорционально снижалось по мере уменьшения количества исходных данных, а качество извлечения синергий снижалось при уменьшении количества анализируемых мышц. Результаты реконструкции исходных электромиографических сигналов методами факторизации данных оказываются более эффективными при рассмотрении различных периодов в сравнении с целым циклом бегового шага. Показатели извлечения мышечных синергий чувствительны к исходному набору электромиограмм, включенных в анализ.
Современные подходы к изучению двигательных синергий, как правило, включают рассмотрение синергетических эффектов на одном, реже двух уровнях организации нервной системы и их взаимодействие в контексте решения разнообразных двигательных задач, причем, вопрос интеграции разных уровней центральной нервной системы в формировании и координации синергий зачастую упускается. Рассматриваемые произвольные движения человека при изучении синергий чаще всего включают локомоторные модели, задачи постурального контроля и некоторые естественные произвольные движения, выполняемые верхними конечностями. Как правило, для выявления синергетических паттернов взаимодействия скелетных мышц применяют алгоритмы факторизации данных, однако получаемые при их использовании результаты существенно расходятся в некоторых аспектах даже при исследовании схожих по биомеханической структуре двигательных актов [1–4]. Это связывают с тем, что применяются разные методологические и вычислительные подходы, не учитывающие специфику рассматриваемых движений и подчас не имеющие строгой стандартизации [5, 6]. В связи с этим методологическая основа и некоторые вычислительные аспекты изучения синергетических эффектов нуждаются в уточнении и дополнении. Кроме того, такие сведения будут способствовать более глубокому пониманию нейронных механизмов, лежащих в основе управления произвольными движениями человека, и могут быть полезны при решении прикладных вопросов совершенствования специализированных двигательных навыков в спорте. Исходя из этого, цель работы заключалась в поиске оптимальных параметров предварительной обработки электромиографических сигналов и оценке их влияния на результаты извлечения обширных мышечных синергий в процессе локомоций умеренной интенсивности.
МЕТОДЫ ИССЛЕДОВАНИЯ
В исследовании приняли участие 10 испытуемых мужского пола, специализирующихся в спринтерском беге. Эксперименты выполнены на базе Научно-исследовательского института проблем спорта и оздоровительной физической культуры ФГБОУ ВО “ВЛГАФК” в лаборатории физиологии нервной и мышечной систем. Все исследования проведены с соблюдением требований и принципов биомедицинской этики, сформулированными в Хельсинской декларации 1964 г., и одобрены биоэтическим комитетом Великолукской государственной академии физической культуры и спорта. Каждый участник представил добровольное письменное информированное согласие на участие в исследованиях.
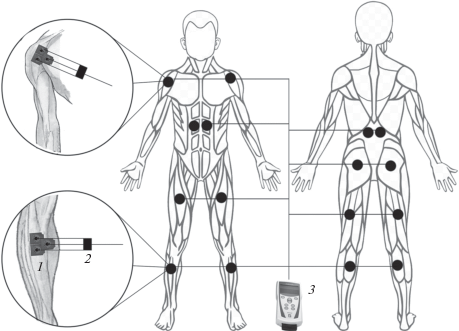
Испытуемые выполняли бег на беговой дорожке HP Cosmos (Австрия) с интенсивностью, определяемой диапазоном сердечных сокращений от 140 до 150 уд./мин. Посредством 16-канальной телеметрической системы МЕ6000 и программного обеспечения MegaWin (MegaElectronics LTD, Финляндия) регистрировали электромиограммы (ЭМГ) 16-ти поверхностных билатеральных мышц туловища, верхних и нижних конечностей: передней большеберцовой, икроножной (медиальная головка), прямой бедра, двуглавой бедра, передней части дельтовидной, выпрямляющей позвоночник (уровень L2), прямой живота (верхняя часть), большой ягодичной. Электромиограф располагался на поясе испытуемого. Регистрацию ЭМГ осуществляли накожными одноразовыми самоклеющимися электродами с токопроводящим гелем с активной площадью контакта 2.5 см2, 36 × 45 мм (Swaromed, Австрия). Электроды накладывались биполярно, при этом активный располагался в области проекции двигательной точки исследуемой мышцы, а референтный прикреплялся по ходу ее волокон с межэлектродным расстоянием 2 см [7]. Для подавления шумов и повышения чистоты записи ЭМГ предусматривался дополнительный электрод заземления на каждой мышце. Расположение отводящих электродов при регистрации ЭМГ представлено на рис. 1. ЭМГ-сигналы усиливали биоусилителем (МЕ 6000) с полосой пропускания частот от 10 до 10 000 Гц и оцифровывали с частотой 2000 Гц. Зарегистрированные интерференционные ЭМГ обрабатывались полосовым фильтром с полосой пропускания 25–250 Гц (25–300, 25–450 Гц), силой подавления 60 дБ, затем выпрямлялись с интервалом усреднения 0.002 с и повторно применяли фильтр низких частот (ФНЧ) 80 или 20 Гц, используя программное обеспечение MegaWin. Обработанные ЭМГ экспортировали в систему Statistica (StatSoft, Inc., version 10) для дальнейшего анализа.
Рис. 1.
Схематическое расположение отводящих электродов при регистрации ЭМГ в условиях бега по тредбану с умеренной интенсивностью. 1 – отводящие электроды, 2 – предусилитель, 3 – электромиограф.
Для определения граничных моментов периодов бегового шага использовали систему 3D-видеозахвата “Qualisys” (Qualisys, Швеция), синхронизированную с биомонитором. Она включала 8 высокоскоростных камер Oqus, располагаемых по периметру вокруг испытуемого. Частота кадров видеозахвата составляла 500 Гц. Для регистрации перемещения сегментов тела использовали светоотражающие маркеры, закрепленные на антропометрических точках тела, совпадающих с осями вращения в суставах. Периоды опоры и переноса определялись моментами постановки конечной антропометрической точки правой нижней конечности на ленту тредбана и ее отрыва. Первичную обработку видеоряда осуществляли в программной среде Qualisys Track Manager. Оцифрованные данные, содержащие координаты антропометрических точек сегментов тела в трехмерном пространстве, и маркеры, определяющие граничные моменты периодов бегового шага, экспортировались в систему Statistica.
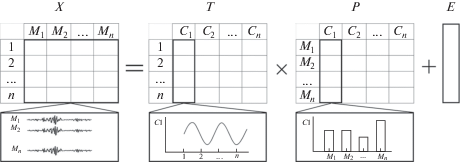
В системе Statistica формировали матрицу исходных данных (X), размерностью (I × J), где I – нормированное число точек целого цикла бегового шага или его отдельных периодов, а J – число независимых переменных (вариационных рядов ЭМГ и координат антропометрических точек сегментов тела). Координаты точек не использовались в процессе извлечения синергий, а применялись только для соотнесения данных ЭМГ с периодами бегового шага. В общей сложности в анализ были включены данные 150 полных циклов. Методы снижения размерности данных используют новые, формальные переменные, являющиеся линейной комбинацией исходных переменных, с помощью которых исходная матрица X разлагается на произведение двух матриц: X = T × P + E, где T – матрица счетов (“scores”), P – матрица нагрузок (“loadings”), E – матрица остатков (рис. 2). Матрица нагрузок несет информацию о взаимосвязи или независимости переменных относительно новых, формальных переменных, полученных в процессе разложения матриц – “векторы синергии”. Отношение определенной мышцы к выявленной синергии определяли по коэффициенту взаимосвязи, превышающему 0.7.
Рис. 2.
Схема разложения исходной матрицы методами определения главных компонент. X – исходная матрица, T – матрица счетов, P – матрица нагрузок, E – матрица остатков. М – ЭМГ одной из мышц, С – главный компонент, 1, 2, … n – значение в момент времени.
Матрица счетов (коэффициенты мышечной активации) определяет временную организацию выявленных мышечных синергий. Количество новых компонент (факторов) вычисляется алгоритмами факторного анализа (FA) и метода главных компонент (PCA), а доля объясняемой дисперсии каждым компонентом указывает на качество реконструкции исходных данных. Рассматривали факторы (компоненты), имеющие собственные значения больше единицы и описывающие не менее 10% общей дисперсии. Факторы (компоненты), отвечающие таким требованиям, определяли количество выявленных мышечных синергий в целом цикле бегового шага и его отдельных периодах. Сравнение коэффициентов мышечной активации и векторов синергии при использовании разных методов извлечения синергий осуществляли с помощью анализа значений кросскорреляционных функций с учетом смещения максимума, где 1 – полное соответствие, 0 – отсутствие взаимосвязи.
Математико-статистическая обработка данных выполнена в Statistica 10.0 и включала расчет среднего арифметического (M), ошибки среднего арифметического (m), коэффициента вариации (CV). Для оценки достоверности различий при сравнении параметров в разных условиях использовали однофакторный дисперсионный анализ (ANOVA) с Post-Hoc анализом по критерию Newman–Keuls. Различия считали значимыми при p < 0.05.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Исходя из цели работы, были проанализированы параметры компонент при использовании различных методов их извлечения. Установлено, что фильтрация исходных ЭМГ в диапазоне 25–250 (300, 450) Гц существенно не влияла на количество компонент и долю объясняемой дисперсии, однако их величины существенно отличались при использовании разных методов извлечения (табл. 1). Так, методом главных компонент извлекались не более 2.9 компонент (синергий), а доля дисперсии находилась в диапазоне 43.6–44.7%. Факторный анализ показывал 5 синергий с долей дисперсии около 60%. Повторная обработка после усреднения фильтром низких частот 80 Гц достоверно повышала оба показателя при использовании PCA, а при FA существенно не изменилась (p > 0.05). Аналогично, с применением ФНЧ 20 Гц возрастало количество извлекаемых компонент PCA и практически не менялось при FA. Доля объясняемой дисперсии в обоих случаях достоверно возрастала и достигала 80% (табл. 1). Таким образом, применение полосового фильтра 25–450 Гц с дополнительной фильтрацией ФНЧ 20 Гц после выпрямления показывало наилучшие результаты по количеству извлекаемых компонент и доли объясняемой дисперсии при использовании как PCA, так и FA.
Таблица 1.
Статистические параметры извлекаемых компонент (синергий) при различных режимах фильтрации исходных ЭМГ-сигналов (n = 150)
| Параметры фильтрации | Стат. параметры | PCA | FA | ||
|---|---|---|---|---|---|
| кол-во извлекаемых компонент (ед.) | доля дисперсии (%) | кол-во извлекаемых компонент (ед.) | доля дисперсии (%) | ||
| 25–250 Гц | M ± m | 2.90 ± 0.10 | 44.70 ± 1.49 | 5.10 ± 0.10 | 61.40 ± 0.65 |
| CV | 10.90 | 10.55 | 6.20 | 3.36 | |
| 25–300 Гц | M ± m | 2.80 ± 0.13 | 43.60 ± 1.84 | 5.00 ± 0.00 | 60.20 ± 0.76 |
| CV | 15.06 | 13.34 | 0 | 3.98 | |
| 25–450 Гц | M ± m | 2.80 ± 0.13 | 43.80 ± 1.87 | 5.00 ± 0.00 | 60.70 ± 0.86 |
| CV | 15.06 | 13.52 | 0 | 4.46 | |
| 25–450 Гц +ФНЧ 80 Гц | M ± m | 3.50 ± 0.17 | 58.10 ± 1.93* | 4.90 ± 0.10 | 68.50 ± 1.21 |
| CV | 15.06 | 10.53 | 6.45 | 5.60 | |
| 25–450 Гц +ФНЧ 20 Гц | M ± m | 4.90 ± 0.41 | 80.60 ± 2.36* | 5.00 ± 0.00 | 82.90 ± 0.59* |
| CV | 26.26 | 9.25 | 0 | 2.24 | |
На рис. 3 представлены образцы ЭМГ поверхностных скелетных мышц в процессе локомоций. В результате применения к исходным данным фильтра 25–450 Гц с повторной фильтрацией ФНЧ 20 Гц электромиограммы фактически представляли собой концентрированные пики активности, однако такая обработка существенно не влияла на среднюю амплитуду ЭМГ. Например, величина ЭМГ-активности передней большеберцовой мышцы правой нижней конечности до фильтрации составляла 71.66 ± 0.82 мкВ, а икроножной медиальной – 85.53 ± 1.58 мкВ. После обработки изменения средней амплитуды ЭМГ названных мышц не превышали 6% (p > 0.05). Достоверных различий средней амплитуды ЭМГ в полном цикле бегового шага, а также в периодах опоры и переноса отдельно при использовании различных параметров фильтрации исходных ЭМГ-сигналов зарегистрировано не было.
Рис. 3.
Образцы ЭМГ скелетных мышц туловища, верхних и нижних конечностей при выполнении локомоций умеренной интенсивности в различных периодах бегового шага. (a) – интерференционная, (b) – выпрямленная, (c) – фильтрованная полосовым фильтром 25–450 Гц, (d) – фильтрованная полосовым фильтром 25–450 Гц + повторная фильтрация ФНЧ 20 Гц.

В различных периодах бегового шага извлеченные мышечные синергии имели некоторые отличия в сравнении их со структурой целого цикла бегового шага. Так, метод главных компонент показывал 6.5 в периоде опоры и 6.2 компонент в периоде переноса (табл. 2). Доля дисперсии в этом случае превышала 90%. Методом факторного анализа извлекалось не более четырех компонент с долей объясняемой дисперсии менее 88%.
Таблица 2.
Статистические параметры извлекаемых компонент (синергий) в различных периодах бегового шага (n = 150)
| Периоды бегового шага | Стат. параметры | PCA | FA | ||
|---|---|---|---|---|---|
| кол-во извлекаемых компонент (ед.) | доля дисперсии (%) | кол-во извлекаемых компонент (ед.) | доля дисперсии (%) | ||
| Опора | M ± m | 6.50 ± 0.34 | 96.00 ± 1.00 | 3.70 ± 0.21 | 87.8 ± 0.80 |
| CV | 16.62 | 3.29 | 18.24 | 2.88 | |
| Перенос | M ± m | 6.20 ± 0.33 | 91.60 ± 1.40 | 4.20 ± 0.20 | 84.00 ± 0.87 |
| CV | 16.66 | 4.83 | 15.06 | 3.27 | |
Таким образом, количество извлекаемых синергий из разных периодов цикла бегового шага оказывалось больше при использовании PCA и меньше при FA, однако по показателю объясняемой дисперсии оба метода давали лучшие результаты, чем при рассмотрении целого цикла бегового шага. Следует также отметить очень низкие показатели вариативности дисперсии, что указывает на высокую плотность данных при их межиндивидуальном рассмотрении. Количество извлекаемых синергий оказывалось более вариативным, CV здесь достигали 16.62%, однако они также оценивались как низкие.
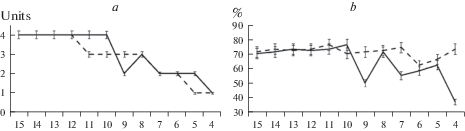
Исходя из цели работы, были проанализированы результаты извлечения мышечных синергий из разных наборов исходных данных ЭМГ. Показано, что качество извлечения синергий при достижении порога в 11 мышц снижается. Так, количество синергий пропорционально снижалось по мере уменьшения количества исходных данных при использовании обоих методов анализа (рис. 4а). По показателю доли объясняемой дисперсии результаты оказались неоднозначными. Факторный анализ демонстрировал стабильные показатели независимо от количества исходных ЭМГ-сигналов, а качество извлечения мышечных синергий снижалось при уменьшении количества анализируемых мышц (рис. 2b).
Рис. 4.
Количество извлекаемых мышечных синергий (a) и доля дисперсии (b) с различным набором исходных ЭМГ сигналов и при использовании разных методов извлечения мышечных синергий. По оси абсцисс – количество мышц. Сплошная линия – результаты при использовании факторного анализа, пунктирная линия – результаты при использовании метода главных компонент.
Дальнейший анализ был сконцентрирован на рассмотрении различий пространственно-временной структуры мышечных синергий, выявляемых при использовании разных методов их извлечения. Следует отметить, что первая компонента при использовании обоих методов объясняла большую часть общей дисперсии, поэтому в работе будет представлены данные о структуре первой синергии. Установлено, что активность первой мышечной синергии снижалась в первой четверти периода опоры, затем возрастала и снова снижалась к завершению периода (рис. 5a). При использовании разных методов извлечения мышечных синергий была установлена высокая взаимосвязь их коэффициентов мышечной активации (rmax = 0.84 ± 0.05). Пространственная структура первой извлеченной мышечной синергии включала ЭМГ-активность дельтовидных мышц и большой ягодичной мышцы правой стороны, причем, она сохранялась при использовании и факторного анализа и метода главных компонент (рис. 3b). В последнем случае была выявлена дополнительная мышца, входящая в структуру этой же синергии – прямая бедра правой нижней конечности. Также было установлено высокое соответствие пространственной структуры данной синергии в периоде опоры при использовании FA и PCA.
Рис. 5.
Пространственно-временная структура мышечной синергии при использовании разных методов их извлечения в процессе локомоций умеренной интенсивности. По оси абсцисс на панели a – коэффициенты активации синергии (у. е.), на панели b – векторы синергии (у. е.). 1 – передняя большеберцовая правая, 2 – икроножная правая, 3 – прямая бедра правая, 4 – двуглавая бедра правая, 5 – передняя большеберцовая левая, 6 – икроножная левая, 7 – прямая бедра левая, 8 – двуглавая бедра левая, 9 – дельтовидная правая, 10 – дельтовидная левая, 11 – выпрямляющая позвоночник правая, 12 – выпрямляющая позвоночник левая, 13 – прямая живота правая, 14 – большая ягодичная правая, 15 – прямая живота левая, 16 – большая ягодичная левая.

ЭМГ-активность дельтовидной мышцы правой верхней конечности и большой ягодичной мышцы левой нижней конечности определяла характерные паттерны временной структуры синергии в периоде переноса (рис. 5). Так, было выявлено три пика активности синергии в первых двух и последней четвертях периода. Также была выявлена дополнительная мышца методом главных компонент, входящая в структуру синергии – прямая бедра левой нижней конечности. В целом коэффициенты мышечной активации и векторы синергии демонстрировали высокую степень соответствия при использовании различных методов факторизации данных.
При рассмотрении структуры целого цикла бегового шага было выявлено пять пиков активности синергии, обусловленные, преимущественно, активностью дельтовидных мышц обеих верхних конечностей (рис. 5). Следует отметить, что использование PCA в данном случае давало несколько лучшие результаты, чем FA в плане определения пространственной структуры извлеченных мышечных синергий. Тем не менее, коэффициенты кросскорреляционных функций оценивались как высокие при анализе соответствия коэффициентов мышечной активации и векторов синергии – 0.92 ± 0.02 и 0.83 ± 0.26 соответственно.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Применяемые в нашей работе методы факторизации данных являются широко используемыми для извлечения мышечных синергий. При этом приводятся доказательства в пользу того, что получаемые результаты отражают реальные аспекты организации паттернов активации скелетных мышц, лежащие в основе двигательного поведения человека, а не являются вычислительным артефактом [1, 8, 9]. Однако применяются различные параметры предварительной обработки данных, включающие фильтрацию в диапазоне от 25 до 850 Гц, с последующей фильтрацией или без нее. Нам удалось приблизиться к результатам, описанным в работах ряда авторов по изучению локомоторной активности разной интенсивности, а именно, количество извлекаемых мышечных синергий достигало в наших исследованиях пяти при уровне объясняемой дисперсии около 80%, в литературных данных, как правило, приводят порог 90%, однако есть работы, где ее порог устанавливается и на более низком уровне [9–11]. Ряд авторов применяют метод неотрицательной матричной факторизации как наиболее эффективный, на их взгляд, метод при работе с положительными значениями вариационных рядов данных ЭМГ [5]. Наряду с этими исследованиями приводятся и другие данные, свидетельствующие о не меньшей эффективности использования факторного анализа и других методов снижения размерности данных. Тем не менее, использование разных методов при изучении схожих по биомеханической структуре движений может давать несколько различающиеся результаты. Есть данные, что даже несущественные различия в позиционировании отводящих электродов при регистрации ЭМГ могут влиять на сходство синергетических паттернов мышечных активаций.
Количество извлекаемых мышечных синергий может быть критерием, указывающим на нейромышечную стратегию центральной нервной системы, предусматривающую достижение оптимальных параметров моторного выхода для обеспечения стабильности конечного результата движения [12, 13]. Большее количество синергий может свидетельствовать о более сложной стратегии управления, вследствие включения в управляющую систему значительного количества управляемых элементов – синергий [11, 12, 14]. Логично предположить, что целый беговой шаг по своей координационной структуре может быть значительно сложнее, чем движения, составляющие структуру его отдельных периодов, однако в таком случае количество синергий, задействованных в их реализации, должно быть несколько меньше. Результаты наших исследований показывают обратное, оказывается, что управление движением на уровне подзадач, т.е. в отдельных периодах движения, характеризуется не меньшей сложностью, на что указывает количество извлекаемых мышечных синергий и величина объясняемой дисперсии, достигающая описанной в литературе пороговой величины – 90%.
Известно, что центральная нервная система не генерирует каждый раз новые мышечные синергии, а гибко комбинирует существующий набор синергий в соответствии с изменяющимися условиями выполнения двигательной задачи, постулируется, что множество мышечных синергий могут быть активированы одновременно, и каждая мышца может быть представлена в паттернах нескольких синергий [15, 16]. Исходя из этого, мышечные синергии, извлекаемые из одного набора исходных ЭМГ-данных, в структуре цикла бегового шага не являются результатом сложения синергетических эффектов, наблюдаемых в отдельных периодах цикла, а вероятно, являются частью двигательной программы, направленной на поддержание оптимальных параметров локомоций при заданной интенсивности работы.
Возможные причины расхождений в результатах при рассмотрении схожих по биомеханической структуре движений заключаются в применении различных процедур предварительной обработки ЭМГ данных (фильтрации, нормализации и др.), использовании разных методов извлечения синергетических эффектов, вариативности исходного набора ЭМГ, включенных в анализ. Выбор скелетных мышц для анализа является непростой задачей, поскольку синергетические эффекты не могут быть выявлены непосредственно в ходе эксперимента, а становятся доступны после применения процедур математической обработки зарегистрированных сигналов. На наш взгляд, вопрос количества ЭМГ, включаемых в анализ при извлечении мышечных синергий, является скорее поисковой задачей, но чаще их состав определяется логическим путем или на основе анализа турн-амплитудных характеристик электромиограмм. Наши данные свидетельствуют, что включение в анализ менее одиннадцати ЭМГ снижает количество мышечных синергий и эффективность их извлечения.
Анализ данных литературы показывает, что при изучении локомоторной активности нижних конечностей регистрируют электрическую активность поверхностных мышц одной, реже двух ног, крайне редко в анализ включают мышцы спины [13, 16–18]. Тот факт, что при таком существенном варьировании скелетных мышц в разных исследованиях выявляются от 4 до 6 синергий позволяет предположить, что синергетические паттерны взаимодействия скелетных мышц могут быть обнаружены в любом наборе исходных ЭМГ данных. Более важным является вопрос установления оптимальных границ, в рамках которых результатам можно дать физиологическую интерпретацию. Так, выявляются синергии, состоящие из двух или одной мышцы, что нередко встречается при использовании подхода к изучению мышечных синергий методами сокращения размерности данных, что противоречит самой концепции синергизма, как механизма снижения вычислительной нагрузки на центральную нервную систему посредством объединения управляемых элементов (мышц) в модули с меньшей размерностью. В одной из работ было показано, что характерные синергетические паттерны могут быть обнаружены в активности отдельных двигательных единиц m. flexor digitorum superficialis при помощи PCA [19]. Таким образом, выбор скелетных мышц и вопрос их количества в процессе изучения синергетических эффектов скорее определяется целями и задачами конкретного исследования.
Сравнение коэффициентов мышечной активации и векторов синергий в нашем исследовании показывает высокое их сходство при использовании разных методов извлечения мышечных синергий, но PCA выявляет большее количество мышц, определяющих пространственную их организацию. Таким образом, оба метода могут быть использованы для выявления синергетических эффектов при локомоциях умеренной интенсивности, однако метод главных компонент дает несколько лучшие результаты.
ЗАКЛЮЧЕНИЕ
Таким образом, применяемый в работе методологический подход, включающий рассмотрение синергетических эффектов мышц туловища, верхних и нижних конечностей, позволяет эффективно выявлять мышечные синергии в структуре цикла бегового шага. Повышение качества реконструкции исходных ЭМГ-сигналов может быть достигнуто применением полосового фильтра с полосой пропускания 20–450 Гц с использованием после выпрямления дополнительной фильтрации 20 Гц. Такая предобработка существенно не влияет на величину средней амплитуды ЭМГ, но достоверно повышает качество реконструкции исходных данных.
Извлечение мышечных синергий методами факторизации данных оказывается более эффективным при рассмотрении различных периодов в сравнении с целым циклом бегового шага. Количество извлекаемых мышечных синергий пропорционально снижается по мере уменьшения числа исходных ЭМГ-сигналов при использовании FA и PCA, что необходимо учитывать при определении минимального количества электромиограмм, включаемых в анализ.
Выявлено высокое соответствие коэффициентов мышечной активации и векторов мышечной синергии при использовании FA и PCA, однако векторы синергии, определяемые методом главных компонент, представлены бόльшим количеством скелетных мышц при рассмотрении целого цикла бегового шага и его отдельных периодов. Это указывает на целесообразность применения PCA для анализа пространственной структуры мышечных синергий при локомоциях умеренной интенсивности.
Список литературы
Tresch M, Cheung V, d’Avella A (2006) Matrix factorization algorithms for the identification of muscle synergies: evaluation on simulated and experimental data sets. J Neurophysiol l95 (4): 2199–2212. https://doi.org/10.1152/jn.00222.2005
Escalona M, Bourbonnais D, Goyette M, Le Flem D, Duclos C, Gagnon D (2021) Effects of varying overground walking speeds on lower-extremity muscle synergies in healthy individuals. Motor Control 25(2): 234–251. https://doi.org/10.1123/mc.2020-0008
Kerkman J, Bekius A, Boonstra T (2020) Muscle Synergies and Coherence Networks Reflect Different Modes of Coordination During Walking. Front Physiol 11: 751. https://doi.org/10.3389/fphys.2020.00751
Mileti I, Serra A, Wolf N, Munoz-Martel V, Ekizos A, Palermo E, Arampatzis A, Santuz A (2020) Muscle activation patterns are more constrained and regular in treadmill than in overground human locomotion. Front Bioeng Biotechnol 8: 581619. https://doi.org/10.3389/fbioe.2020.581619
Rabbi M, Pizzolato C, Lloyd D, Carty C, Devaprakash D, Diamond L (2020) Non-negative matrix factorization is the most appropriate method for extraction of muscle synergies in walking and running. Sci Rep 10(1): 8266. https://doi.org/10.1038/s41598-020-65257-w
Turpin N, Uriac S, Dalleau G (2021) How to improve the muscle synergy analysis methodology? Eur J Appl Physiol 121(4): 1009–1025. https://doi.org/10.1007/s00421-021-04604-9
Altenburger K, Bumke O, Foerster O (1937) Allgemeine Neurologie. Handbuch der Neurologie. Berlin.
Cheung V, Cheung B, Zhang J, Chan Z, Ha S, Chen C, Cheung R (2020) Plasticity of muscle synergies through fractionation and merging during development and training of human runners. Nat Commun 11(1): 4356. https://doi.org/10.1038/s41467-020-18210-4
Lambert-Shirzad N, Van der Loos N (2017) On identifying kinematic and muscle synergies: a comparison of matrix factorization methods using experimental data from the healthy population. J Neurophysiol 117(1): 290–302. https://doi.org/10.1152/jn.00435.2016
Gui K, Zhang D (2016) Influence of locomotion speed on biomechanical subtask and muscle synergy. J Electromyogr Kinesiol 30: 209–215. https://doi.org/10.1016/j.jelekin.2016.07.010
Hajiloo B, Anbarian M, Esmaeili H, Mirzapour M (2020) The effects of fatigue on synergy of selected lower limb muscles during running. J Biomech 16: 103–109. https://doi.org/10.1016/j.jbiomech.2020.109692
Santuz A, Brüll L, Ekizos A (2020) Neuromotor Dynamics of Human Locomotion in Challenging Settings. Science 23(1): 100796. https://doi.org/10.1016/j.isci.2019.100796
Chia Bejarano N, Pedrocchi A, Nardone A, Schieppati M, Baccinelli W, Monticone M, Ferrigno G, Ferrante S (2017) Tuning of muscle synergies during walking along rectilinear and curvilinear trajectories in humans. Ann Biomed Eng 45: 1204–1218. https://doi.org/10.1007/s10439-017-1802-z
Santuz A, Ekizos A, Kunimasa Y (2020) Lower complexity of motor primitives ensures robust control of high-speed human locomotion. Heliyon 6(10): e05377. https://doi.org/10.1016/j.heliyon.2020.e05377
Amundsen HS, Van Acker G, Luchies C, Cheney P (2018) Muscle synergies obtained from comprehensive mapping of the cortical forelimb representation using stimulus triggered averaging of EMG activity. J Neurosci 38(41): 8759–8771. https://doi.org/10.1523/JNEUROSCI.2519-17.2018
Nishida K, Hagio S, Kibushi B, Moritani T, Kouzaki M (2017) Comparison of muscle synergies for running between different foot strike patterns. PLoS One 12(2): e0171535. https://doi.org/10.1371/journal.pone.0171535
Saito A, Tomita A, Ando R (2018) Muscle synergies are consistent across level and uphill treadmill running. Sci Rep 8: 5979. https://doi.org/10.1038/s41598-018-24332-z
Barroso F, Torricelli D, Moreno J, Taylor J, Gomez-Soriano J, Bravo-Esteban E, Piazza S, Santos C, Pons J (2014) Shared muscle synergies in human walking and cycling. J Neurophysiol 112(8): 1984–1998. https://doi.org/10.1152/jn.00220.2014
Madarshahian S, Letizi J, Latash ML (2021) Synergic control of a single muscle: The example of flexor digitorum superficialis. J Physiol 599(4): 1261–1279. https://doi.org/10.1113/JP280555
Дополнительные материалы отсутствуют.
Инструменты
Российский физиологический журнал им. И.М. Сеченова