

Российский физиологический журнал им. И.М. Сеченова, 2022, T. 108, № 10, стр. 1305-1315
Выполнение произвольных движений в условиях активации системы непроизвольного внимания
В. М. Князева 1, *, А. М. Плакхин 1, А. А. Александров 1
1 Санкт-Петербургский государственный университет
Санкт-Петербург, Россия
* E-mail: v.m.knyazeva@spbu.ru
Поступила в редакцию 14.07.2022
После доработки 12.08.2022
Принята к публикации 23.08.2022
- EDN: GWXONL
- DOI: 10.31857/S0869813922100090
Аннотация
В работе исследовалось влияние процессов, сопряженных с активацией системы автоматического обнаружения изменений в слуховой среде и переключения внимания, на вызванные потенциалы, связанные с подготовкой и реализацией произвольного движения. Во время эксперимента испытуемые производили самоинициированные нажатия на клавишу указательным пальцем правой руки одновременно с прослушиванием двух типов стимульных последовательностей: парадигма одд-болл и контрольная парадигма. Испытуемых просили сосредоточить внимание на моторном задании и игнорировать звуковые стимулы. Парадигма одд-болл представляла собой последовательность повторяющихся (стандартных) и отклоняющихся (девиантных) стимулов, предъявление которых приводит к активации системы непроизвольного внимания. Анализ распределения числа нажатий относительно времени подачи звукового стимула показал, что в парадигме одд-болл достоверно увеличивается число нажатий, совершенных на интервале 800–1200 мс после предъявления стимула по сравнению с интервалом, который соответствует моменту предъявления стимула (0–400 мс). При анализе потенциала готовности было обнаружено значительное увеличение амплитуды билатерально симметрично распределенного компонента на интервале от –300 до –600 мс до начала движения, совершаемого одновременно с прослушиванием стимулов в парадигме одд-болл по сравнению с контрольной парадигмой. Полученные результаты позволяют говорить об активирующем эффекте процессов, связанных с вовлечением системы непроизвольного внимания, на ранние компоненты подготовки движения.
Регистрация электроэнцефалограммы в момент подготовки и совершения произвольного движения позволяет установить, что указанные процессы сопровождаются формированием медленного отрицательного потенциала, известного как потенциал готовности (ПГ). В последнее время интенсивно разрабатываются интерфейсы “мозг–компьютер”, основанные на классификации паттернов ПГ, что определяет необходимость исследования нейрофизиологических механизмов этого процесса, а также поиска новых подходов к улучшению распознавания потенциалов в реальном времени [1].
ПГ имеет преимущественно центральное распределение и может быть зарегистрирован за несколько секунд до совершения движения. В среднем за 300–500 мс до начала движения наблюдается резкое увеличение негативности сигнала, которое выделяют в отдельный компонент – поздний ПГ [2, 3]. Поздний ПГ имеет асимметричное распределение с максимумом над контралатеральными областями моторной коры (для движений руки) [4].
Наиболее вероятным источником позднего ПГ является контралатеральная моторная кора (поля 3 и 4 по Бродману), однако корковые генераторы раннего ПГ остаются предметом для дискуссий [5]. Ряд авторов полагает, что в случае движений руки основными генераторами раннего ПГ являются билатеральная дополнительная и пре-дополнительная моторная области (поле 6) и цингулярная моторная кора [6]. С другой стороны, результаты построения источников активности показывают, что наилучший результат обнаруживается при моделировании генератора раннего ПГ в билатеральной моторной коре с минимальным вкладом дополнительной моторной области в генерацию сигнала [7]. Исследования с применением функциональной магнитно-резонансной томографии показали, что большинство испытуемых демонстрируют активацию в области дополнительной моторной коры, в то время как у части испытуемых также наблюдается активация контралатеральной моторной коры за 1 с до начала движения [8]. Таким образом, все гипотезы относительно корковых генераторов раннего ПГ можно разделить на три группы: источником ПГ является только билатеральная дополнительная моторная область, только моторная кора или же имеет место совместная активация этих двух областей [5].
Время возникновения, амплитуда и топографическое распределение ранней и поздней составляющих ПГ зависят от вида движения, его темпа, скорости, силы, точности и сложности моторного ответа, а также от тренировки, обучения, готовности совершить движение и некоторых других параметров [9]. На ранний ПГ влияют такие когнитивные факторы, как степень нисходящего контроля со стороны ЦНС, необходимость планирования и выбора движения, тогда как на поздний ПГ больше влияют факторы, связанные с самим движением, такие как точность, дискретность и сложность. Это согласуется с вероятной локализацией генераторов двух компонент ПГ: билатеральная премоторная кора и латеральная премоторная кора для раннего ПГ и контралатеральная прецентральная кора для позднего ПГ. Однако точная связь каждого компонента с намерением совершить движение остается до конца не исследованной [10].
В известном эксперименте, проведенном исследовательской группой под руководством Libet [11], была произведена оценка времени возникновения сознательного желания совершить движение относительно времени начала ПГ. После совершения простого движения, такого как поднятие пальца, участников эксперимента просили сообщить, где находилась вращающаяся точка на циферблате в тот момент, когда они впервые осознали желание двигаться. Было обнаружено, что ПГ начинается примерно за 1.5–2.0 с до фактического возникновения намерения действовать, таким образом, показывая, что подготовка движения начинается раньше, чем это осознается человеком.
Представленная интерпретация эксперимента Libet не является единственной. Согласно так называемой стохастической модели принятия решения [12, 13] при отсутствии побуждающих команд точный момент принятия решения о совершении движения определяется случайными колебаниями активности нейронов (которым соответствует ранний ПГ). Эти спонтанные колебания приводят к превышению порогового значения группой нейронов, ответственных за определенное действие (поздний ПГ), что сопровождается субъективным принятием решения о совершении движения.
Теория сэмплирования медленных потенциалов коры основывается на аналогичном предположении [14]. В течение негативной фазы постоянных спонтанных колебаний корковой активности, возникающих с частотой менее 1 Гц, происходит повышение возбудимости нейронных сетей, связанных с инициацией движения. Во время этих флуктуаций переживается неспецифическое желание совершить движение, что приводит к повышению вероятности спонтанного произвольного движения во время негативной фазы/негативного пика этих флуктуаций.
Кроме этого, существует также теория, что ПГ, формирующийся при совершении произвольных движений, идентичен условной негативной волне и является коррелятом процесса ожидания [15]. С другой стороны, существуют данные о том, что условная негативная волна является комбинацией ПГ и отдельного, связанного с ожиданием, компонента [16]. Также было высказано предположение, что условная негативная волна и негативность, связанная с ожиданием стимула, могут быть связаны с вниманием и иметь общие нервные генераторы в теменной области, но различаться в лобных областях [17]
Таким образом, вопрос о связи ПГ и других медленноволновых потенциалов остается открытым. Также до сих пор точно неизвестно, какие компоненты ПГ, кроме латерализованного ПГ, действительно связаны с подготовкой движения [18] и является ли ранний ПГ нейронным коррелятом планирования или подготовки моторного акта, сознательного намерения совершить движение, их комбинации или ни одного из вышеперечисленного [10, 13]. В связи с этим представляет интерес изучить зависимость между ПГ и процессами внимания, оказывающими активирующее действие на процессы подготовки движения.
МЕТОДЫ ИССЛЕДОВАНИЯ
В эксперименте приняли участие 14 человек (из них 5 мужчин; средний возраст 24 ± 4 лет). Все испытуемые были правшами и сообщили об отсутствии неврологических заболеваний, нормальном слухе и нормальном или скорректированном зрении. Все процедуры, выполненные в исследованиях с участием людей, соответствуют этическим стандартам национального комитета по исследовательской этике и Хельсинкской декларации 1964 г. и ее последующим изменениям или сопоставимым нормам этики. От каждого из включенных в исследование участников было получено информированное добровольное согласие. Экспериментальные процедуры были одобрены Этическим комитетом Санкт-Петербургского государственного университета.
Во время эксперимента испытуемые производили произвольные самоинициированные нажатия на клавишу указательным пальцем правой руки. Испытуемых просили избегать ритмических движений, варьировать время между нажатиями и совершать нажатие достаточное количество раз для набора статистики (точная цифра не оговаривалась). Во время эксперимента через головные телефоны предъявлялись последовательности звуковых стимулов, которые испытуемых просили игнорировать. Каждому участнику в случайном порядке без перерыва предъявлялись два типа стимульных последовательностей: парадигма одд-болл и контрольная парадигма. Парадигма одд-болл состояла из двух типов стимулов, проигрываемых в псевдослучайном порядке: звуковой тон частотой 1000 Гц и вероятностью предъявления 85% (стандартный стимул) и звуковой тон частотой 1200 Гц и вероятностью предъявления 15% (девиантный стимул). Возникновение отклоняющихся (девиантных) стимулов в последовательности повторяющихся стандартных приводит к активации системы непроизвольного внимания, что отражается в генерации негативности рассогласования в вызванных потенциалах. Контрольная парадигма состояла только из стимулов частотой 1000 Гц. В каждой парадигме предъявлялась 1000 стимулов. Длительность стимула составляла 50 мс (время нарастания и спада 5 мс), интенсивность – 60 дБ, межстимульный интервал варьировал от 800 до 900 мс. Для создания и предъявления стимулов использовалась программа “Presentation” (Neurobehavioral Systems, США).
У каждого испытуемого в обеих парадигмах рассчитывалось общее количество нажатий, а также проводился кросскорреляционный анализ моментов нажатий и времени предъявления девиантных и контрольных стимулов. Для того, чтобы число взятых в анализ контрольных стимулов было равно числу девиантных стимулов, из всех стимулов, предъявляемых в контрольной парадигме были взяты в анализ 150 стимулов, совпадающих с девиантными стимулами по времени предъявления в последовательности. Число нажатий, совершенных относительно времени подачи звукового стимула на интервале от –400 до 1600 мс с шагом 400 мс было рассчитано с использованием алгоритма, входящего в пакет MLIB [19] для среды MATLAB (MathWorks, США).
Регистрация электроэнцефалограммы проводилась при помощи хлорсеребряных электродов, размещенных в отведениях: F3, Fz, F4, C3, Cz, C4, Pz согласно международной системе 10–20. Референтный электрод находился на кончике носа, заземляющий – на лбу. Электроокулограмма регистрировалась при помощи электрода, расположенного около височного угла левого глаза. Сопротивление электродов не превышало 5 кОм. Для регистрации ЭЭГ использовался цифровой электроэнцефалограф Мицар-ЭЭГ-05/70-202 совместно с пакетом программного обеспечения для регистрации и обработки электроэнцефалограммы WinEEG (ООО Мицар, Россия).
Для анализа ПГ были взяты данные 10 испытуемых (из них 2 мужчин; средний возраст 23 ± 3 лет), совершивших более 30 нажатий (среднее число нажатий 87). ЭЭГ сигнал был отфильтрован в полосе 0.032–40 Гц. Продолжительность анализируемой эпохи составляла 3000 мс. Для коррекции изолинии использовался интервал от 2800 до 2000 мс до начала нажатия. Пробы, амплитуда сигнала в которых превышала по амплитуде ±100 мкВ на любом из каналов были удалены из анализа.
Все данные имели нормальное распределение согласно критерию Колмогорова–Смиронова. Статистический анализ среднего количества нажатий и количества нажатий, совершенных относительно времени подачи звукового стимула, проводился с использованием непараметрического критерия Уилкоксона. Анализ амплитуды потенциала готовности проводился с использованием дисперсионного анализа с повторными измерениями (IBM SPSS Statistic v. 21, IBM Corporation, Нью-Йорк, США) на интервалах от –1200 до 0 мс с шагом 300 мс с внутригрупповыми факторами “Парадигма” (парадигма одд-болл/контрольная парадигма), “Фронтальность” (фронтальные/центральные отведения) и “Латерализация” (левое полушарие/медианные отведения/правое полушарие). В случае значимого отклонения от предположения о сферичности, оцениваемого с помощью критерия Моучли, применялась поправка статистики F-критерия по методу Гринхауса–Гейссера.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Поведенческие данные
Среднее количество нажатий, совершенное испытуемыми в парадигме одд-болл было на 27% больше (М = 85, SD = 64), чем в контрольной парадигме (М = 73, SD = 53), однако статистический анализ с помощью критерия Уилкоксона показал только маргинально достоверные отличия между полученными значениями (p = 0.06). Следует отметить, что полученные данные имеют значительную разницу между испытуемыми: максимальное число нажатий оставило 316 (165 – в парадигме одд-болл и 151 – в парадигме контроль), а минимальное – 22 (11 – в парадигме одд-болл и 11 – в парадигме контроль).
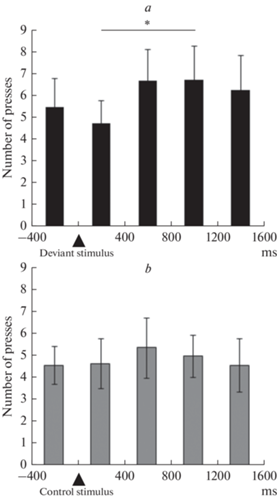
На рис. 1 представлено распределение числа нажатий относительно времени подачи звукового стимула на интервале от –400 до 1600 мс с шагом 400 мс. Было проведено сравнение суммарного количества нажатий, совершенных на указанном интервале с помощью непараметрического критерия Уилкоксона. Обнаружено достоверное увеличение числа нажатий на интервале 800–1200 мс после предъявления девиантного стимула по сравнению с интервалом 0–400 мс (р = 0.015) в парадигме одд-болл. В контрольной парадигме достоверных отличий обнаружено не было.
Рис. 1.
Распределение среднего числа нажатий (± ошибка среднего) относительно времени подачи девиантного и контрольного стимула на интервале от –400 до 1600 мс с шагом 400 мс в парадигме одд-болл (а) и контрольной парадигме (b). Момент предъявления стимула обозначен стрелкой. * – р < 0.05 по результатам статистического анализа с помощью критерия Уилкоксона.
Потенциал готовности
Для анализа ПГ были взяты данные 10 испытуемых, совершивших более 30 нажатий (среднее число нажатий 87). Визуальный анализ данных показал наличие медленно нарастающей негативности, начинающейся примерно за 1000 мс до начала движения в центральных отведениях, которую мы определили как ПГ. На рис. 2 представлен ПГ, зарегистрированный в центральных отведениях в парадигме одд-болл и контрольной парадигме. В табл. 1 представлены средние значения амплитуды ПГ на интервалах от –1200 до 0 мс с шагом 300 мс.
Рис. 2.
Потенциал готовности, зарегистрированный в центральных отведениях в парадигме одд-болл (черная линия) и контрольной парадигме (серая линия). Момент совершения нажатия показан вертикальной пунктирной линией. Серым прямоугольником выделен интервал –300 до –600 мс. * – р < 0.05 по результатам дисперсионного анализа с повторными измерениями.
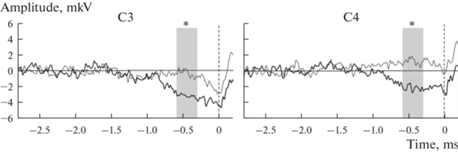
Таблица 1.
Средние значения (M, мкВ) и значения стандартной ошибки среднего (SEM) амплитуды потенциала готовности
| Парадигма | Отведение | ||||||
|---|---|---|---|---|---|---|---|
| F3 | Fz | F4 | C3 | Cz | C4 | ||
| от –300 до 0 мс | |||||||
| Контрольная парадигма | M | –0.80 | 0.22 | 2.76 | –1.62 | –1.05 | 0.84 |
| SEM | 1.90 | 1.97 | 2.43 | 1.56 | 1.89 | 1.69 | |
| Парадигма одд-болл | M | –3.66 | –3.90 | –2.74 | –3.81 | –3.17 | –2.16 |
| SEM | 1.24 | 1.04 | 1.01 | 0.79 | 0.67 | 0.70 | |
| от –600 до –300 мс | |||||||
| Контрольная парадигма | M | –0.24 | 0.81 | 3.11 | –0.36 | –0.32 | 1.13 |
| SEM | 1.50 | 1.69 | 2.48 | 1.26 | 1.68 | 1.51 | |
| Парадигма одд-болл | M | –3.83 | –4.02 | –2.82 | –3.19 | –2.79 | –2.17 |
| SEM | 0.91 | 0.75 | 0.61 | 0.57 | 0.53 | 0.57 | |
| от –900 до –600 мс | |||||||
| Контрольная парадигма | M | –0.58 | 0.38 | 2.62 | –0.41 | –0.38 | 0.89 |
| SEM | 1.48 | 1.52 | 2.31 | 1.28 | 1.57 | 1.37 | |
| Парадигма одд-болл | M | –2.26 | –2.44 | –1.75 | –1.63 | –1.25 | –1.01 |
| SEM | 0.66 | 0.51 | 0.45 | 0.30 | 0.38 | 0.37 | |
| от –1200 до–900 мс | |||||||
| Контрольная парадигма | M | –1.31 | –0.11 | 2.07 | –0.87 | –0.55 | 0.56 |
| SEM | 1.07 | 1.06 | 1.81 | 0.88 | 1.08 | 0.88 | |
| Парадигма одд-болл | M | –0.88 | –1.05 | –0.06 | –0.94 | –0.37 | –0.19 |
| SEM | 0.74 | 0.62 | 0.96 | 0.44 | 0.43 | 0.44 | |
Дисперсионный анализ показал достоверное влияние факторов “Фронтальность” (F(9,1) = 5.380, p = 0.046, η2 = 0.374) и “Латерализация” (F(9,1) = 7.163, p = 0.019, η2 = 0.443) на интервале от –300 до 0 мс до начала движения. На указанном интервале наблюдается увеличение негативности сигнала в центральных отведениях и в левом полушарии.
На интервале от –600 до –300 мс было обнаружено достоверное влияние факторов “Парадигма” (F(9.1) = 8.586, p = 0.017, η2 = 0.488), а также пограничное к значению р < 0.05 взаимодействие факторов “Парадигма” × “Фронтальность” (F(9.1) = 3.726, p = 0.086, η2 = 0.293). Апостериорный анализ парных сравнений показал достоверное увеличение негативности ответа в парадигме одд-болл по сравнению с контрольной парадигмой как в центральных (р = 0.044), так и во фронтальных отведениях (р = 0.016). В парадигме одд-болл наблюдалась достоверно более негативная амплитуда вызванных потенциалов (ВП) во фронтальных отведениях чем в центральных (р = 0.033), в то время как в контрольной парадигме эти различия не достигают уровня статистической значимости (р = 0.171).
На интервалах от –900 до –600 мс и от –1200 до –900 мс до начала движения достоверных влияний факторов или взаимодействия факторов обнаружено не было.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В представленной работе изучалось влияние активации процессов непроизвольного внимания, связанных с генерацией негативности рассогласования, на ВП, сопровождающие подготовку и совершение произвольного движения.
Мы обнаружили, что при совершении произвольных движений в парадигме одд-болл наблюдается некоторое увеличение числа нажатий по сравнению с контрольной парадигмой, однако не достигающее уровня статистической значимости. Анализ распределения числа нажатий относительно времени подачи звукового стимула показал, что в парадигме одд-болл число нажатий, совершенных на интервале 800–1200 мс после предъявления стимула достоверно больше по сравнению с интервалом, соответствующим моменту предъявления стимула (0–400 мс). Полученные данные позволяют говорить о наличии неравномерности в распределении числа нажатий в парадигме одд-балл по сравнению с контрольной парадигмой.
В парадигме одд-болл при различии физических характеристик в последовательно предъявляемых стимулах генерируется так называемая негативность рассогласования – компонент ВП отражающий процессы непроизвольного переключения внимания [20, 21]. Отмечают, что активация системы автоматического обнаружения изменений в слуховой среде и переключения внимания может оказывать стимулирующее действие и приводить к снижению времени реакции и количества ошибок при ответе на стимул [22, 23], а также уменьшать скорость развития мышечного утомления [24, 25]. Мы предполагаем, что процессы, связанные с генерацией негативности рассогласования, также могут оказывать активирующее влияние на процесс подготовки произвольного самоинициированного движения.
Согласно стохастической модели принятия решения и теории сэмплирования медленных потенциалов коры в отсутствие сильного волевого сигнала выбор точного момента начала самоинициированного движения определяется моментом превышения некоторого порогового уровня активности группой нейронов интеграторов [12–14]. Рост числа нажатий после предъявления девиантного стимула может быть следствием неспецифического активирующего влияния со стороны систем внимания, усиливающих свою активность после генерации негативности рассогласования.
Данные регистрации ПГ также показывают значительное увеличение амплитуды ВП в парадигме одд-болл на интервале от –600 до –300 мс до начала движения. Следует отметить, что на этом интервале ПГ является билатерально симметричным, что выражается в отсутствие достоверного влиянии фактора “Латерализация” на указанном интервале. На более позднем интервале от –300 до 0 мс наблюдается достоверная разница между амплитудой ответа для правого и левого полушария, в то время как увеличение амплитуды ПГ в парадигме одд-болл не достигает уровня статистической значимости.
Согласно литературным данным, при выполнении простых самопроизвольных движений в медленном темпе ранний ПГ начинает развиваться билатерально примерно за 2 с до начала движения в пре-дополнительной моторной области (pre-SMA) и в собственно дополнительной моторной области (SMA), а вскоре после этого в латеральной премоторной коре. Примерно за 400 мс до начала движения в контралатеральной моторной коре и латеральной премоторной коре возникает еще один более крутой отрицательный наклон (поздний ПГ) [10, 26]. Таким образом, обнаруженное в парадигме одд-болл достоверное увеличение амплитуды ПГ на интервале от –600 до –300 мс соответствует билатерально симметричному ПГ, генерируемому предположительно в дополнительной моторной области и латеральной премоторной коре.
Установлено, что субъективное отношение испытуемого к стимулу, например, сосредоточение или отвлечение внимания, меняет распределение и параметры ВП. Предыдущие исследования показали, что выполнение сложных комплексных действий, требующих соответственно большего объема внимания, приводит к увеличению амплитуды позднего ПГ [27–29]. В работе Keller и Heckhausen [30] было проведено сравнение ПГ при совершении произвольных движений в классической парадигме Libet и непроизвольных движений. Было установлено, что и произвольные, и непроизвольные движения вызывают ПГ, однако при совершении произвольных движений ПГ достоверно более выражен. Более того, авторы установили различия в распределении потенциала: при совершении непроизвольных движений активация наблюдалась преимущественно в контрлатеральных областях моторной коры, в то время как при совершении произвольных движений были задействованы области дополнительной моторной коры. Аналогичный результат был получен в ряде других работ [31, 32], где было показано, что внимание (и, предположительно, осознание намерения действовать) приводит к увеличению амплитуды ПГ, зарегистрированного над медиальными лобно-центральными областями в течение 500 мс до начала движения.
Можно предположить, что полученное в нашем исследовании увеличение амплитуды ПГ является результатом повышения концентрации произвольного внимания на текущей моторной задаче, которое необходимо для преодоления постоянного рефлекторного отвлечения внимания девиантными стимулами. Кроме того, следует отметить, что одновременно с увеличением ПГ в одд-болл парадигме наблюдается смещение максимума амплитуды ВП к фронтальным отведениям, что может говорить о большем вкладе дополнительной моторной области в генерацию потенциала. Таким образом, повышение внимания к задаче ожидаемо вызывает повышение активации центральной нервной системы, что приводит к усилению активности областей коры, отвечающих за подготовку движения [33].
Полученные данные также согласуются с результатами ряда исследований условной негативной волны вызванных потенциалов мозга человека [34, 35], что может говорить о сходных нейронных процессах, лежащих в основе генерации этих медленных потенциалов. Недавние исследования [36] также подтверждают эту гипотезу, демонстрируя влияние системы внимания на условно-негативной волне, приводящее к усилению нейронной активности в области дополнительной моторной коры.
Следует отметить, что мониторинг и классификация паттернов ПГ в режиме реального времени является одним из перспективных направлений для разработки интерфейсов мозг–компьютер [1, 37]. Полученные в нашем исследовании результаты по усилению выраженности ПГ в условиях активации системы внимания в парадигме одд-болл могут найти применение в такого рода исследованиях как одного из возможных механизмов улучшения качества различения одиночных ВП.
Список литературы
Кирой В, Лазуренко Д, Шепелев И, Асланян Е, Миняева Н, Бахтин О (2017) Нейротехнологии: нейро-БОС и интерфейс “мозг – компьютер”. Изд-во Южн федерал универ. Ростов-на-Дону. [Kiroy V, Lazurenko D, Shepelev I, Aslanyan E, Minyaeva N, Bakhtin O (2017) Neurotechnologies: neuro-BFB and brain-computer interface. Southern Federal Univ Press. Rostov-on-Don. (In Russ)].
Deecke L, Grozinger B, Kornhuber HH (1976) Voluntary finger movement in man: Cerebral potentials and theory. Biol Cybern 23: 99–119. https://doi.org/10.1007/BF00336013
Shibasaki H, Barrett G, Halliday E, Halliday A (1980) Components of the movement-related cortical potential and their scalp topography. Electroencephalogr Clin Neurophysiol 49: 213–226. https://doi.org/10.1016/0013-4694(80)90216-3
Shibasaki H, Barrett G, Halliday E, Halliday AM (1981) Cortical potentials associated with voluntary foot movement in man. Electroencephalogr Clin Neurophysiol 52: 507–516. https://doi.org/10.1016/0013-4694(81)91426-7
MacKinnon CD (2003) Recordings of Movement-Related Potentials Combined with PET, fMRI of MEG. In: Jahanshahi M, Hallett M (eds) The Bereitschaftspotential. Springer New York 95–111.
Deecke L, Kornhuber HH (2003) Human Freedom, Reasoned Will, and the Brain: The Bereitschaftspotential Story. In: Jahanshahi M, Hallett M (eds) The Bereitschaftspotential. Springer New York 283–320.
Botzel K, Paulus W, Scherg M (1993) Bereitschaftspotential: is there a contribution of the supplementary motor area? Electroencephalogr Clin Neurophysiol Evoked Potentials 89: 187–196. https://doi.org/10.1016/0168-5597(93)90132-9
Ball T, Schreiber A, Feige B, Wagner M, Lecking CH, Kristeva-Feige R (1999) The role of higher-order motor areas in voluntary movement as revealed by high-resolution EEG and fMRI. Neuroimage 10: 682–694. https://doi.org/10.1006/nimg.1999.0507
Lang W (2003) Surface Recordings of the Bereitschaftspotential in Normals. In: Jahanshahi M, Hallett M (eds) The Bereitschaftspotential. Springer New York 19–34.
Shibasaki H, Hallett M (2006) What is the Bereitschaftspotential? Clin Neurophysiol 117: 2341–2356.https://doi.org/10.1016/j.clinph.2006.04.025
Libet B, Gleason CA, Wright EW, Pearl DK (1983) Time of conscious intention to act in relation to onset of cerebral activity (readiness-potential): The unconscious initiation of a freely voluntary act. Brain 106: 623–642. https://doi.org/10.1093/brain/106.3.623
Schurger A, Mylopoulos M, Rosenthal D (2016) Neural Antecedents of Spontaneous Voluntary Movement: A New Perspective. Trends Cogn Sci 20: 77–79. https://doi.org/10.1016/j.tics.2015.11.003
Schurger A, Sitt JD, Dehaene S (2012) An accumulator model for spontaneous neural activity prior to self-initiated movement. Proc Natl Acad Sci U S A 109: E2904–E2913.https://doi.org/10.1073/pnas.1210467109
Schmidt S, Jo HG, Wittmann M, Hinterberger T (2016) “Catching the waves” – slow cortical potentials as moderator of voluntary action. Neurosci Biobehav Rev 68: 639–650. https://doi.org/10.1016/j.neubiorev.2016.06.023
Schlegel A, Alexander P, Sinnott-Armstrong W, Roskies A, Tse PU, Wheatley T (2013) Barking up the wrong free: Readiness potentials reflect processes independent of conscious will. Exp Brain Res 229: 329–335. https://doi.org/10.1007/s00221-013-3479-3
Damen EJP, Brunia CHM (1987) Changes in Heart Rate and Slow Brain Potentials Related to Motor Preparation and Stimulus Anticipation in a Time Estimation Task. Psychophysiology 24: 700–713. https://doi.org/10.1111/j.1469-8986.1987.tb00353.x
Kotani Y, Ohgami Y, Arai J, Kiryu S, Inoue Y (2011) Motor and nonmotor components of event-brain potential in preparation of motor response. J Behav Brain Sci 1: 234–241.https://doi.org/10.4236/jbbs.2011.14030
Schurger A, Hu P Ben, Pak J, Roskies AL (2021) What Is the Readiness Potential? Trends Cogn Sci 25: 558–570. https://doi.org/10.1016/j.tics.2021.04.001
Stüttgen M (2022) MLIB – toolbox for analyzing spike data. https://www.mathworks.com/matlabcentral/fileexchange/37339-mlib-toolbox-for-analyzing-spike-data
Näätänen R, Gaillard AWK, Mäntysalo S (1978) Early selective-attention effect on evoked potential reinterpreted. Acta Psychol (Amst) 42: 313–329. https://doi.org/10.1016/0001-6918(78)90006-9
Näätänen R, Astikainen P, Ruusuvirta T, Huotilainen M (2010) Automatic auditory intelligence: An expression of the sensory-cognitive core of cognitive processes. Brain Res Rev 64: 123–136.
Aleksandrov AA, Babanin ME, Stankevich LN (2003) Mechanisms of Generation of Mismatch Negativity and Their Role in the Recognition of Brief Auditory Stimuli. Neurosci Behav Physiol 33: 867–872. https://doi.org/10.1023/A:1025992620828
Aleksandrov AA, Starostina LV, Stankevich LN (2005) Characteristics of evoked potentials of the human brain during recognition of short acoustic stimuli of different frequencies. Neurosci Behav Physiol 35: 209–213. https://doi.org/10.1007/s11055-005-0015-3
Aleksandrov AA, Knyazeva VM (2017) Effects of Cognitive Loading on the Development of Muscle Fatigue. Neurosci Behav Physiol 47: 960–966. https://doi.org/10.1007/s11055-017-0497-9
Aleksandrov AA, Knyazeva VM, Stankevich LN, Dmitrieva ES, Shestakova AN (2016) Mismatch negativity affects muscle fatigue during repeated contraction trials of different durations. Front Physiol 7: 13. https://doi.org/10.3389/fphys.2016.00013
Di Russo F, Berchicci M, Bozzacchi C, Perri RL, Pitzalis S, Spinelli D (2017) Beyond the “Bereitschaftspotential”: Action preparation behind cognitive functions. Neurosci Biobehav Rev 78: 57–81. https://doi.org/10.1016/j.neubiorev.2017.04.019
Simonetta M, Clanet M, Rascol O (1991) Bereitschaftspotential in a simple movement or in a motor sequence starting with the same simple movement. Electroencephalogr Clin Neurophysiol Evoked Potentials 81: 129–134. https://doi.org/10.1016/0168-5597(91)90006-J
Kitamura JI, Shibasaki H, Takagi A, Nabeshima H, Yamaguchi A (1993) Enhanced negative slope of cortical potentials before sequential as compared with simultaneous extensions of two fingers. Electroencephalogr Clin Neurophysiol 86: 176–182. https://doi.org/10.1016/0013-4694(93)90005-G
Masaki H, Takasawa N, Yamazaki K (1998) Enhanced negative slope of the readiness potential preceding a target force production task. Electroencephalogr Clin Neurophysiol – Evoked Potentials 108: 390–397. https://doi.org/10.1016/S0168-5597(98)00019-7
Keller I, Heckhausen H (1990) Readiness potentials preceding spontaneous motor acts: voluntary vs. involuntary control. Electroencephalogr Clin Neurophysiol 76: 351–361. https://doi.org/10.1016/0013-4694(90)90036-J
Takashima S, Cravo AM, Sameshima K, Ramos RT (2018) The effect of conscious intention to act on the Bereitschaftspotential. Exp Brain Res 236: 2287–2297. https://doi.org/10.1007/s00221-018-5302-7
Takashima S, Najman FA, Ramos RT (2019) Disruption of volitional control in obsessive-compulsive disorder: Evidence from the Bereitschaftspotential. Psychiatry Res – Neuroimaging 290: 30–37. https://doi.org/10.1016/j.pscychresns.2019.06.007
Freude G, Ullsperger P, Kruger H, Pietschmann M (1988) Bereitschaftspotential in preparation to mental activities international. J Psychophysiol 6: 291–297. https://doi.org/10.1016/0167-8760(88)90016-5
Birbaumer N, Elbert T, Lutzenberger W, Rockstroh B, Schward J (1981) EEG and slow cortical potentials in anticipation of mental tasks with different hemispheric involvement. Biol Psychol 13: 251–260. https://doi.org/10.1016/0301-0511(81)90040-5
Simons RF, Öhman A, Lang PJ (1979) Anticipation and Response Set: Cortical, Cardiac, and Electrodermal Correlates. Psychophysiology 16: 222–233. https://doi.org/10.1111/j.1469-8986.1979.tb02982.x
Liu Y, Zhang D, Ma J, Li D, Yin H, Luo Y (2013) The attention modulation on timing: an event-related potential study. PLoS One 8: e66190. https://doi.org/10.1371/journal.pone.0066190
Волкова КВ, Дагаев НИ, Киселев АС, Касумов ВР, Александров МВ, Осадчий АЕ (2017) Интерфейс мозг-компьютер: опыт построения, использования и возможные пути повышения рабочих характеристик, Журн высш нервн деятельн им ИП Павлова 67: 504–520. [Volkova KV, Dagaev NI, Kiselev AS, Kasumov VR, Aleksandrov MV, Osadchiy AE (2017) Brain-computer interface: experience in building, using and possible ways to improve performance, J Higher Nerv Activity named after IP Pavlova 67: 504–520. (In Russ)]. https://doi.org/10.7868/s0044467717040128
Дополнительные материалы отсутствуют.
Инструменты
Российский физиологический журнал им. И.М. Сеченова