

Российский физиологический журнал им. И.М. Сеченова, 2022, T. 108, № 11, стр. 1442-1452
Влияние сложности задачи на величину синхронизации ЭЭГ активности бета-диапазона в сенсомоторной коре
М. В. Пронина 1, *, В. А. Пономарев 1, Ю. Д. Кропотов 1
1 Институт мозга человека им. Н.П. Бехтеревой Российской академии наук
Санкт-Петербург, Россия
* E-mail: pronina@ihb.spb.ru
Поступила в редакцию 23.08.2022
После доработки 19.09.2022
Принята к публикации 20.09.2022
- EDN: DHZVVG
- DOI: 10.31857/S0869813922110103
Аннотация
В работе исследовали влияние сложности второстепенной задачи на связанные с событиями перестройки ЭЭГ в сенсомоторной области коры при выполнении основной задачи в условиях многозадачности. 32-канальную ЭЭГ регистрировали у 24 здоровых испытуемых во время выполнения четырех тестов, сочетающих Go/NoGo и N-back задачи и различающихся по сложности. В отличие от работ, использующих классическую N-back парадигму, в которых нагрузка на рабочую память создавалась за счет необходимости удержания в памяти информации непосредственно о стимуле, для настоящего исследования мы разработали вариант N-back задачи, которая требует запоминания совершенного действия. Было показано, что повышение сложности N-back задачи приводило к уменьшению величины синхронизации бета-диапазона ЭЭГ, наблюдаемой после совершения движения в Go условии Go/NoGo теста над левой сенсомоторной областью коры (максимум в отведении C3). Можно предполагать, что увеличение нагрузки на рабочую память, независимо от типа удерживаемой в памяти информации, приводит к ослаблению процессов контроля движений, необходимых для выполнения основной задачи. Полученные результаты говорят в пользу предположения о том, что состояние многозадачности требует распределения ресурсов внимания и за счет этого ухудшает качество выполнения каждой из задач.
В современном мире состояние многозадачности становится все более распространенным и привычным. Однако одновременное выполнение нескольких задач сопряжено с риском того, что на качество выполнения приоритетной задачи могут повлиять второстепенные [1]. Ряд исследований указывает на то, что одновременное выполнение нескольких задач приводит к ухудшению когнитивных и моторных функций, таких как обработка сенсорных стимулов [2], запоминание информации [3], выполнение двигательных заданий [1], вождение автомобиля [1], поддержание баланса при ходьбе [4] и даже изменению походки [5].
Частным вариантом второстепенной задачи является необходимость удержания и/или обновления информации в рабочей памяти. Большинство литературных данных указывает на то, что увеличение нагрузки на рабочую память приводит к ухудшению выполнения приоритетной задачи [6–8]. Предполагается, что нагрузка на рабочую память снижает концентрацию внимания на основной задаче, так как ресурсы когнитивного контроля истощаются и внимание может “перетекать” на отвлекающие стимулы [6]. Согласно этой гипотезе, ухудшение выполнения основной задачи происходит только при наличии отвлекающего стимула, который похож на целевой [8]. В пользу этой гипотезы говорят результаты работы [9], в которой практически никакого влияния нагрузки на рабочую память на выполнение основной задачи выявлено не было.
Одним из наиболее часто используемых заданий для создания нагрузки на рабочую память является N-back задача [10]. В ее классическом варианте от испытуемого требуется сравнить текущий стимул с тем, которые предъявлялся в последовательности N шагов назад. В качестве стимулов могут использоваться буквы, цифры, символы и т.д., и с увеличением числа N растет нагрузка на рабочую память.
Результаты электрофизиологических исследований показывают, что увеличение нагрузки на рабочую память в N-back задаче влияет на мощность активности бета-диапазона ЭЭГ (13–30 Гц) [10], в частности снижает величину синхронизации бета-диапазона ЭЭГ, наблюдаемую после совершения произвольных и связанных с предъявлением стимула двигательных реакций над сенсомоторной областью контрлатерального полушария (реакция бета-ритма после возбуждения или rebound effect) [11, 12].
Однако в работах, использующих классическую парадигму N-back задачи, нагрузка на рабочую память создавалась за счет необходимости удержания в памяти информации непосредственно о стимуле, между тем значительный интерес представляет исследование влияния на величину бета-ритма после возбуждения других типов запоминаемой информации. Поэтому для настоящего исследования мы разработали вариант N-back задачи, которая требует запоминания совершенного действия. В качестве основной задачи мы использовали зрительный тест Go/NoGo парадигмы с предупреждающим стимулом. Этот тип теста требует как можно быстрее реагировать (например, нажатием на кнопку) на определенные стимулы и воздерживаться от нажатия на другие, тем самым задействуя процессы подготовки, совершения и торможения действия. Варианты Go/NoGo теста с предупреждающим стимулом включают дополнительный стимул, который с определенной долей вероятности указывает на то, что следующий стимул будет целевым, требуя вовлечения рабочей памяти.
Результаты предыдущих исследований указывают на то, что в случае, когда предупреждающий и целевой стимул физически идентичны, в височной коре происходит формирование модели ожидаемого стимула [13], основанной на запоминании физических характеристик стимула, если же стимулы различаются, формирование такого образа невозможно, и в рабочей памяти удерживаются другие характеристики целевого стимула, например, семантическая категория.
Так как в процессе выполнения теста испытуемому необходимо совершать движение после предъявления целевого стимула, в данных вызванной синхронизации/десинхронизации в Go условии над левой центральной корой можно наблюдать реакцию бета-ритма после возбуждения [14].
В данной работе мы исследовали влияние сложности N-back задачи на величину реакции бета-ритма после возбуждения при выполнении зрительного Go/NoGo теста. На основании литературных данных, полученных с использованием классического варианта N-back задачи, было предположено, что повышение сложности N-back задачи, требующей запоминания совершенного действия, будет снижать величину реакции бета-ритма после возбуждения в сенсомоторной коре, контралатеральной по отношению к руке, которой совершается нажатие в Go условии Go/NoGo теста. Кроме того, чтобы проверить, как необходимость удержания в памяти информации о совершенном движении будет влиять на реакцию бета-ритма после возбуждения в условиях, требующих запоминания физических характеристик целевого стимула или его семантической категории, мы использовали два варианта Go/NoGo теста с целевыми парами, состоящими из идентичных или различных изображений.
МЕТОДЫ ИССЛЕДОВАНИЯ
Испытуемые
В исследовании принимали участие 24 испытуемых в возрасте от 19 до 33 лет (4 мужчины), средний возраст 23 ± 43 года, без серьезных травм головы в анамнезе, психических, неврологических, онкологических и нейродегенеративных заболеваний, алкогольной и наркотической зависимости, не принимающие психотропные препараты. Все процедуры, выполненные в исследовании, соответствуют этическим стандартам национального комитета по исследовательской этике и Хельсинкской декларации 1964 г. и ее последующим изменениям или сопоставимым нормам этики. От каждого из включенных в исследование участников было получено информированное добровольное согласие. Процедура исследования была одобрена комиссией по этике Института мозга человека РАН 20 сентября 2018 г.
Структура тестов
Каждый тест состоял из 400 проб, каждая проба содержала два стимула, стимулами являлись изображения животных и растений. Разные сочетания стимулов составляли 4 типа проб: “животное–животное”, “животное–растение”, “растение–растение” и “растение–животное”. При этом в пробах “животное–животное” и “растение–растение” первое и второе изображения были полностью идентичны. Пробы предъявлялись на экране 19-дюймового монитора в случайном порядке с вероятностью 25%. Длительность проб – 3000 мс, интервал от начала пробы до начала предъявления первого изображения (престимульный интервал) – 300 мс, длительность каждого стимула – 100 мс, длительность интервала между двумя стимулами в паре – 1000 мс. После каждой пары изображений через 1800 мс в середине экрана на 100 мс предъявлялось изображение креста (рис. 1). Интервал между предъявлением креста и следующей пары изображений – 1900 мс. Общая продолжительность каждого теста – 35 мин.
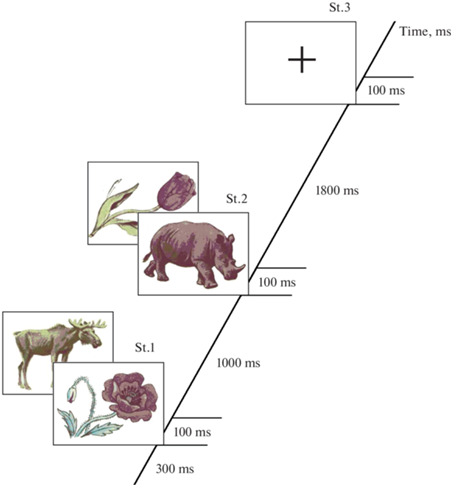
В каждом тесте испытуемые параллельно выполняли две задачи – Go/NoGo и N-back, которые, в свою очередь, имели по два варианта выполнения. Таким образом, всего использовалось 4 теста с разными сочетаниями заданий.
В тесте 1 (Ж–Ж, N–0) первым заданием было нажимать указательным пальцем правой руки на левую кнопку мыши как можно быстрее после предъявления пары картинок “животное–животное” (проба Go), не нажимая после других пар (Go/NoGo задача). Вторым заданием было нажимать после изображения креста указательным пальцем правой руки на левую кнопку мыши в случае, если текущей пробой была проба Go (N-back задача, сложность N–0), и нажимать на правую кнопку мыши средним пальцем в случае всех остальных проб. В тесте 2 (Ж–Ж, N–1) первое задание было аналогично тесту 1, вторым заданием было нажимать как можно быстрее после изображения креста на левую кнопку мыши, если проба, предшествующая последней, была пробой Go (N-back задача, сложность N–1), и нажимать на правую кнопку мыши в случае всех остальных типов проб. В тестах 3 (Ж–Р, N–0) и 4 (Ж–Р, N–1) первым заданием было нажимать как можно быстрее на левую кнопку мыши после пары картинок “животное–растение”, не нажимая после других пар. Второе задание было аналогично заданиям в тестах 1 и 2 соответственно.
ЭЭГ данные регистрировались во время выполнения 4 тестов в течение двух сеансов, два теста за один сеанс. Варианты теста предъявлялись испытуемым попарно (1, 2 и 3, 4) в случайном порядке в соответствии с рандомизационными списками.
Регистрация ЭЭГ
Регистрация электроэнцефалограммы производилась с помощью 31-канального компьютерного электроэнцефалографа “Мицар-202” (СЕ 0537) производства ООО Мицар (http://www.mitsar-eeg.ru/) и электроэнцефалографической шапочки МКС (производства ООО “Медицинские Компьютерные Системы”, Москва, Зеленоград). ЭЭГ электроды располагались на поверхности головы в соответствии с системой 10–10 в отведениях Fp1; Fpz; Fp2; F7; F3; Fz; F4; F8; FT7; FC3; FCz; FC4; FT8; T3; C3; Cz; C4; T4; TP7; CP3; CPz; CP4; TP8; T5; P3; Pz; P4; T6; O1; Oz; O2, референтные электроды – на мочках ушей, заземляющий – в лобной области. Место установки ушных электродов предварительно зачищали абразивной пастой “NuPrep”, электроды заполняли небольшим количеством геля “Унимакс”. Сопротивление электродов не превышало 5 кОм. Частота оцифровки ЭЭГ составляла 250 Гц. Параметры фильтров высокой и низкой частоты составляли соответственно 0.53 и 30 Гц, также использовался режекторный фильтр – 45–55 Гц. ЭЭГ регистрировалась в монтаже с объединенным ушным референтом.
Для контроля над правильностью выполнения тестов и измерения времени реакции регистрировался сигнал от левой и правой кнопки мыши.
Исследование проводилось в спокойной обстановке, при отсутствии отвлекающих стимулов. Во время исследования испытуемые сидели в удобном кресле на расстоянии примерно 1 м от монитора. Перед регистрацией каждого теста испытуемым подробно объяснялось задание, и проводилась серия тренировочных проб.
Обработка данных
Ошибочные пробы при выполнении Go/NoGo теста (пробы Go, в которых испытуемый не нажал на кнопку и пробы NoGo с ложными нажатиями) исключались из анализа. Их количество подсчитывалось и сравнивалось между разными тестами с использованием непараметрического критерия Манна–Уитни и многофакторного дисперсионного анализа ANOVA в рамках программного пакета Statistica 10. Также было проведено сравнение количества ошибок, допущенных при нажатии на изображение креста.
Коррекция артефактов глазных движений производилась в WinEEG автоматически методом зануления независимых компонент, соответствующих этим движениям. Кроме того, в автоматическом режиме удалялись фрагменты ЭЭГ, содержавшие активность, превышающую 100 мкВ, а также фрагменты с амплитудой сигнала 50 мкВ и выше в диапазоне 0–1 Гц, и с амплитудой 35 мкВ и выше в диапазоне 20–35 Гц. Эти пороги были ранее подобраны эмпирически [14].
ЭЭГ преобразовывалась в плотность источников тока (Current Source Density – CSD) для лучшего выявления локальных эффектов динамики спектральной плотности ЭЭГ. Применение этого метода имеет ряд преимуществ, включающих более точное определение структуры, местоположения и ориентации внутричерепных источников сигнала, а также возможности обнаружения глубоко расположенных источников [15].
В реакциях ЭЭГ различают два вида активности: вызванную и индуцированную. Вызванная активность (или связанные с событиями потенциалы) представляет собой изменения электрического потенциала, привязанные по фазе к стимулу или ответной реакции. Наиболее часто в нейрофизиологических работах исследуют именно потенциалы, связанные с событиями. Индуцированная активность, или вызванная десинхронизация/синхронизация (event-related desynchronization/synchronization – ERD/ERS), напротив, не привязана по фазе к стимулу. Такие реакции ЭЭГ описываются в виде изменений спектральной плотности электрического потенциала и их исследование дает дополнительную информацию о работе систем мозга [16].
Чтобы уменьшить влияние вызванной активности на динамику спектральной плотности ЭЭГ, усредненные потенциалы, связанные с событиями, вычитались из каждой индивидуальной пробы. В предыдущей статье авторов было показано, что вычитание ПСС из эпох ЭЭГ частично подавляет увеличение спектральной плотности сигналов тета-диапазона, хотя и не гарантирует полного удаления вызванной активности [14].
Для расчета вызванной десинхронизации/синхронизации ЭЭГ к данным применялось вейвлет преобразование исходных сигналов ЭЭГ. Для этого вычислялась свертка исходного сигнала с вейвлетом Морле. Динамика спектральной плотности вычислялась для каждого испытуемого, каждого электрода, каждого теста и каждого типа проб в отдельности. Вейвлет преобразование выполнялось для различных частот в диапазоне от 2 до 30 Гц с шагом 1 Гц. Спектральная плотность вычислялась и усреднялась по пробам одного типа. Наконец, ERD/ERS оценивалась как изменение спектральной мощности для каждой частоты отдельно в соответствие с формулой ERD/ERS(f) = (P(f, t) – PBG(f))/PBG (f), где PBG (f) – средняя мощность в интервале длительностью 300 мс перед предъявлением первого стимула в пробах.
Статистический анализ величины ERD/ERS в разных тестовых заданиях проводился с помощью основанного на кластерах теста с перестановками [17], реализованного в WinEEG на основе инструментария FieldTrip MATLAB для анализа МЭГ/ЭЭГ (свободно доступного по адресу http://fieldtrip.fcdonders.nl/). Идея этого метода основана на том, что на первом этапе выполняется сравнение сигналов в двух условиях. При этом для каждого канала и каждого временного отсчета в отдельности вычисляются значения статистик, полученных с помощью непараметрических тестов (Вилкоксона или Манна–Уитни, в зависимости от вида сравнения). Далее различия со значениями p меньше критического (p < 0.01) группируются в кластеры на основании подобия во временной и частотной областях. Наконец, для каждого из выделенных кластеров вычисляются суммы величин указанных статистик. Для оценки статистической значимости выделенных кластеров используется метод Монте-Карло, при этом исходные данные случайным образом перемешиваются. Затем снова выполняется поиск кластеров уже для перемешанных данных. Перемешивание выполнятся путем перестановок меток проб, число перемешиваний – 10 000. Далее для каждого кластера, наблюдаемого в исходных данных, вычисляется вероятность выделения кластеров в перемешанных данных с большей или равной суммой статистик. И эта вероятность принимается в качестве статистической значимости кластера. В дальнейшем рассматриваются только кластеры со статистической значимостью p < 0.05. Такой метод имеет ряд преимуществ [17] по сравнению с традиционно используемыми подходами. Он исключает необходимость субъективного выделения интервалов времени, в которых наблюдаются различия сигналов, и позволяет решить проблему множественных сравнений. И, наконец, он выполняет статистические оценки, используя непараметрические критерии.
В данной работе основанный на кластерах анализ с перестановками использовался для сравнения величины связанных с событиями перестроек ЭЭГ над левой сенсомоторной областью коры в Go условии четырех тестов попарно таким образом, чтобы целевая пара в Go/NoGo задаче была одинаковой, а сложность N-back задачи различалась, то есть тест 1 (Ж–Ж, N–0) сравнивался с тестом 2 (Ж–Ж, N–1), а тест 3 (Ж–Р, N–0) c тестом 4 (Ж–Р, N–1). Аналогично сигналы сравнивались между тестами с одинаковой сложностью N-back задачи, но разными целевыми парами в Go/NoGo задаче (тесты 1 и 3 и тесты 2 и 4). Значения статистик получали с помощью критерия Манна–Уитни.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Анализ поведенческих показателей
Средние значения количества ошибок, допущенных испытуемыми при выполнении Go/NoGo задачи в каждом из четырех типов теста (нажатие на левую кнопку мыши после пары изображений), времени реакции и ошибки времени реакции представлены в табл. 1.
Таблица 1.
Поведенческие показатели при выполнении Go/NoGo задачи в четырех тестах
| Тест | Пропуски | Ложные нажатия | Время реакции (мс) |
Ошибка времени реакции (мс) |
|---|---|---|---|---|
| Тест 1 | 4.39 | 0.78 | 393* | 12.9* |
| Тест 2 | 4.58 | 0.79 | 416 | 14.5 |
| Тест 3 | 2.60 | 1 | 481* | 17.8* |
| Тест 4 | 3.57 | 1.13 | 459 | 17.6 |
Количество допущенных ошибок между тестами не различалось. Значимые различия были получены для времени реакции (p < 0.05) и стандартного отклонения времени реакции (p < 0.01) между тестом 1 (Ж–Ж, N–0) и тестом 3 (Ж–Р, N–0). Различий по количеству ошибок и времени реакции при нажатии на изображение креста между тестами обнаружено не было.
Дисперсионный анализ показал значимое влияние на поведенческие показатели фактора “Тип целевой пробы (Ж–Ж/Ж–Р)” (p < 0.01), значимого влияния фактора “Сложность N-back задачи” и взаимодействия факторов обнаружено не было.
Анализ вызванной десинхронизации/синхронизации
Основанный на кластерах тест с перестановками при сравнении тестов между собой выявил в каждом сравнении сходные кластеры, наблюдаемые для высокочастотной активности после второго стимула в пробах Go и локализованные в левой центральной области. Величина синхронизации активности бета-диапазона была наибольшей в отведении C3 (параметры кластеров, выделенных в отведении C3, представлены в табл. 2). Кластеры с подобными параметрами также были выделены в близлежащих отведениях: при сравнении тестов 1 и 2 – в отведениях FCz (p < 0.01) и Cz (p < 0.01), при сравнении тестов 3 и 4 – в отведениях Cz (p < 0.01) и CPz (p < 0.01).
Таблица 2.
Параметры кластеров, выделенных при сравнении величины ERD/ERS над сенсомоторной корой (отведение C3) в Go пробах между тестами
| Сравнения тестов | Полярность | Латентность, мс | Частота, Гц | p < |
|---|---|---|---|---|
| 1–2 | + | 1000–1380 | 16–29 | 0.01 |
| 3–4 | + | 960–1200 | 15–30 | 0.01 |
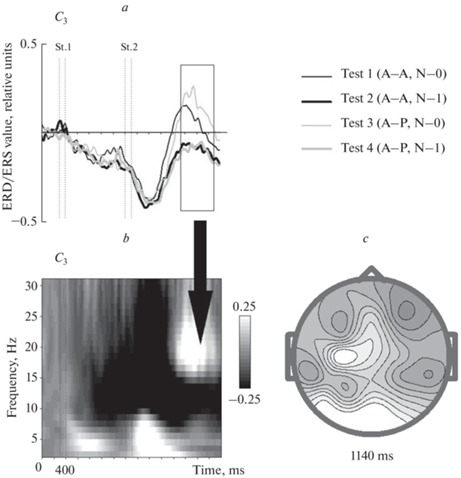
Величина вызванной синхронизации в бета-диапазоне в интервале примерно 980–1480 мс в пробе Go уменьшалась (p < 0.01) в тестах с более сложной второй задачей (тесты 2 и 4) по сравнению с тестами с более простой второй задачей (тесты 1 и 3). Статистически значимых различий по величине бета-синхронизации в левой центральной области между вариантами тестов с разными целевыми парами в Go/NoGo задаче, но одинаковой сложностью N-back задачи (тесты 1 и 3, и тесты 2 и 4), обнаружено не было.
Частотно-временное распределение и топография максимальной величины изменения спектральной плотности ЭЭГ при выполнении тестов представлены на рис. 2.
Рис. 2.
Реакция синхронизации бета-ритма после возбуждения над левой моторной корой в Go пробах. (a) – общие средние значения изменений спектральной плотности ЭЭГ для средней частоты вейвлета 20 Гц при выполнении четырех тестов. Ось X – время в мс, ось Y – величина изменения спектральной плотности относительно среднего значения в интервале перед первым стимулом (в относительных единицах). Вертикальные пунктирные линии показывают начало и окончание предъявления стимулов (St.1 и St.2). Тонкая черная линия – тест 1, толстая черная линия – тест 2, тонкая серая линия – тест 3, толстая серая линия – тест 4. Черным прямоугольником отмечен временной интервал, определенный статистическим анализом как кластер. (b) – частотно-временное представление связанного с событиями изменения спектральной плотности ЭЭГ в тесте 3. Ось X – время в мс, ось Y – частота в Гц, оттенки серого показывают величину ERD/ERS. (c) – топография распределения максимума среднего значения вызванной синхронизации на поверхности головы в тесте 3 для средней частоты вейвлета 20 Гц.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Поведенческие показатели
В результате проведенного исследования были выявлены значимые различия по времени реакции на Go стимулы и его стандартном отклонении в разных тестах. В предыдущих исследованиях было показано, что в варианте Go/NoGo теста с целевой парой, состоящей из одинаковых картинок (пара “животное–животное”, среднее время реакции примерно на 100 мс меньше, чем в варианте теста с целевой парой из разных картинок (пара “животное–растение”) [13]. Результаты сравнения времени реакции в четырех вариантах теста показывают аналогичные различия между тестами 1 (Ж–Ж, N–0) и 3 (Ж–Р, N–0). Предполагается, что различия в значении среднего времени реакции в разных целевых парах связаны с возможностью формирования модели ожидаемого стимула в височной коре в случае, когда предупреждающий стимул идентичен целевому [13]. Если же предупреждающий и целевой стимул различаются (как в паре картинок “животное–растение”) формирование такого образа невозможно, что увеличивает время, необходимое для распознавания изображения как целевого стимула.
Результаты сравнения стандартного отклонения времени реакции в разных тестах также указывают на то, что этот показатель зависит от типа целевой пары, но не от сложности второй задачи.
Мы ожидали, что увеличение нагрузки на рабочую память в тестах 2 и 4 приведет к увеличению времени реакции относительно аналогичных тестов 1 и 3, но статистически достоверной разницы выявить не удалось, возможно, из-за сравнительно небольшой выборки испытуемых или из-за того, что условие N–1 в Go/NoGo тесте не обеспечивало достаточно сильной нагрузки.
Реакция бета-ритма после возбуждения
Результаты электрофизиологических исследований показывают, что увеличение нагрузки на рабочую память в N-back задаче влияет на мощность активности бета-диапазона ЭЭГ [10], в частности на синхронизацию активности бета-диапазона в левом центральном отведении после совершения движения, которая соответствует реакции бета-ритма после возбуждения, описанной ранее [11]. Поскольку это повышение наблюдается сразу после прекращения движения, предполагается, что оно отражает состояние активного торможение моторных областей и/или обратную связь о результате действия, приходящую в сенсомоторную кору [18, 19]. Литературные данные показывают, что величина и длительность синхронизации бета-активности после совершения движения меняются в зависимости от условий выполняемой задачи, таких как скорость торможения движения [20], осуществляется воображаемое движение или реальное [21], совершается движение одним пальцем или всей рукой [11].
В данном исследовании повышение сложности второй задачи приводило к снижению реакции бета-ритма, наблюдаемой после совершения нажатия на кнопку в процессе выполнения Go/NoGo теста, так же, как и в предыдущих работах с использованием классической N-back задачи [12], и независимо от того, физические характеристики изображения или его семантическую категорию нужно было удерживать в памяти. Можно предполагать, что увеличение нагрузки на рабочую память, независимо от типа удерживаемой в памяти информации, приводит к ухудшению процесса торможения моторной системы после совершения движения [18, 19].
Ряд работ предлагает еще одну точку зрения, заключающуюся в том, что реакция бета-активности после возбуждения связана с оценкой результата действия, а именно с пересмотром существующего моторного плана на основе обновления оценки неопределенности текущей модели после анализа поступающей сенсорной информации [22–24]. В рамках этой интерпретации можно предполагать, что необходимость выполнять дополнительное сложное задание приводит к ухудшению процесса построения модели действия при выполнении основной задачи и, как следствие, негибкости моторной программы.
Полученные в работе электрофизиологические результаты согласуются с данными, представленными в работе 2004 г. [6], и говорят в пользу предположения о том, что выполнение нескольких задач одновременно требует распределения ресурсов внимания, и за счет этого может ухудшать качество выполнения каждой из них. Также можно предположить, что на выполнение более сложной задачи (в данном случае N-back задача) направляется больше ресурсов и качество выполнения более простой задачи (в данном случае Go/NoGo) снижается в большей степени.
Однако сравнение поведенческих показателей не выявило статистически достоверного влияния сложности дополнительной задачи на количество ошибок и скорость реакции при выполнении основной, несмотря на различия средних значений. Возможно, из-за высокой межиндивидуальной вариабельности данных необходимо увеличить количество испытуемых и/или увеличить сложность дополнительной задачи.
Список литературы
Scherbaum S, Gottschalk C, Dshemuchadse M, Fischer R (2015) Action dynamics in multitasking: the impact of additional task factors on the execution of the prioritized motor movement. Front Psychol 6: 934. https://doi.org/10.3389/fpsyg.2015.00934
Romeo Z, Bonato M, Zorzi M, Spironelli C (2019) Electrophysiological correlates of spatial processing during multitasking. Neuropsychologia 133: 107152. https://doi.org/10.1016/j.neuropsychologia.2019.107152
Madore KP, Khazenzon AM, Backes CW, Jiang J, Uncapher MR, Norcia AM, Wagner AD (2020) Memory failure predicted by attention lapsing and media multitasking. Nature 587(7832): 87–91. https://doi.org/10.1038/s41586-020-2870-z
Hennah C, Ellis G, Doumas M (2021) Dual task walking in healthy aging: Effects of narrow and wide walking paths. PLoS One 16(12): e0261647. https://doi.org/10.1371/journal.pone.0261647
Pizzamiglio S, Naeem U, Abdalla H, Turner DL (2017) Neural Correlates of Single- and Dual-Task Walking in the Real World. Front Hum Neurosci 11: 460. https://doi.org/10.3389/fnhum.2017.00460
Lavie N, Hirst A, de Fockert JW, Viding E (2004) Load theory of selective attention and cognitive control. J Exp Psychol Gen 133: 339–354. https://doi.org/10.1037/0096-3445.133.3.339
Capizzi M, Correa A, Sanabria D (2013) Temporal orienting of attention is interfered by concurrent working memory updating. Neuropsychologia 51(2): 326–339. https://doi.org/10.1016/j.neuropsychologia.2012.10.005
Bayramova R, Toffalini E, Bonato M, Grassi M (2021) Auditory selective attention under working memory load. Psychol Res 85(7): 2667–2681. https://doi.org/10.1007/s00426-020-01437-7
Zanto TP, Liu H, Pan P, Gazzaley A (2020) Temporal attention is not affected by working memory load. Cortex 130: 351–361. https://doi.org/10.1016/j.cortex.2020.06.008
Scharinger C, Soutschek A, Schubert T, Gerjets P (2017) Comparison of the Working Memory Load in N-Back and Working Memory Span Tasks by Means of EEG Frequency Band Power and P300 Amplitude. Front Human Neurosci 11: 6. https://doi.org/10.3389/fnhum.2017.00006
Pfurtscheller G, Lopes da Silva FH (1999) Event-related EEG/MEG synchronization and desynchronization: basic principles. Clin Neurophysiol 110: 1842–1857. https://doi.org/10.1016/s1388-2457(99)00141-8
Pesonen M, Hämäläinen H, Krause CM (2007) Brain oscillatory 4-30 Hz responses during a visual n-back memory task with varying memory load. Brain Res 1138: 171–177. https://doi.org/10.1016/j.brainres.2006.12.076
Kropotov JD, Ponomarev VA, Pronina MV, Jäncke L (2017) Functional indexes of reactive cognitive control: ERPs in cued go/no-go tasks. Psychophysiology 54(12): 1899–1915. https://doi.org/10.1111/psyp.12960
Ponomarev VA, Pronina MV, Kropotov YD (2017) Dynamics of the EEG spectral density in the θ, α, and β bands in the visual Go/NoGo task. Hum Physiol 43: 366–376. https://doi.org/10.1134/S0362119717040132
Kayser J, Tenke CE (2015) On the benefits of using surface Laplacian (Current Source Density) methodology in electrophysiology. Int J Psychophysiol 97(3): 171–173. https://doi.org/10.1016/j.ijpsycho.2015.06.001
Başar E, Düzgün A (2016) How is the brain working?: research on brain oscillations and connectivities in a new “take-off” state. Int J Psychophysiol 103: 3–11. https://doi.org/0.1016/j.ijpsycho.2015.02.007
Maris E, Oostenveld R (2007) Nonparametric statistical testing of EEG- and MEG-data. J Neurosci Methods 164(1): 177–190. https://doi.org/10.1016/j.jneumeth.2007.03.024
Wessel JR, Aron AR (2017) On the globality of motor suppression: unexpected events and their influence on behavior and cognition. Neuron 93(2): 259–280. https://doi.org/10.1016/j.neuron.2016.12.013
Barone J, Rossiter HE (2021) Understanding the Role of Sensorimotor Beta Oscillations. Front Systems Neurosci 15: 655886. https://doi.org/10.3389/fnsys.2021.655886
Heinrichs-Graham E, Kurz MJ, Gehringer JE, Wilson TW (2017) The functional role of post-movement beta oscillations in motor termination. Brain Struct Funct 222(7): 3075–3086. https://doi.org/10.1007/s00429-017-1387-1
Solis-Escalante T, Müller-Putz GR, Pfurtscheller G, Neuper C (2012) Cue-induced beta rebound during withholding of overt and covert foot movement. Clin Neurophysiol 123(6): 1182–1190. https://doi.org/10.1016/j.clinph.2012.01.013
Cao L, Hu Y-M (2016) Beta Rebound in Visuomotor Adaptation: Still the Status Quo? J Neurosci 36(24): 6365–6367. https://doi.org/10.1523/JNEUROSCI.1007-16.2016
Tan H, Wade C, Brown P (2016) Post-Movement Beta Activity in Sensorimotor Cortex Indexes Confidence in the Estimations from Internal Models. J Neurosci 36(5): 1516–1528. https://doi.org/10.1523/JNEUROSCI.3204-15.2016
Little S, Bonaiuto J, Barnes G, Bestmann S (2019) Human motor cortical beta bursts relate to movement planning and response errors. PLoS Biol 17(10): e3000479. https://doi.org/10.1371/journal.pbio.3000479
Дополнительные материалы отсутствуют.
Инструменты
Российский физиологический журнал им. И.М. Сеченова