

Российский физиологический журнал им. И.М. Сеченова, 2022, T. 108, № 4, стр. 474-489
Нейрофизиологическое решение обратной задачи ЭЭГ исследования в покое и в условиях слухоречевой нагрузки
С. А. Гуляев *
Федеральный центр мозга и нейротехнологий Федерального медико-биологическое агентства России
Москва, Россия
* E-mail: s.gulyaev73@gmail.com
Поступила в редакцию 26.01.2022
После доработки 15.02.2022
Принята к публикации 18.02.2022
- EDN: TQHDSI
- DOI: 10.31857/S0869813922040033
Аннотация
Речь – основной механизм интеграции человека в общество и освоения им накопленных знаний, поэтому ее нарушения вызывают выраженную социальную дезадаптацию. В настоящее время требуется разработка объективного метода функционального исследования, позволяющего выявлять быстропротекающие процессы в нейронных сетях головного мозга, фиксировать их и формировать результаты исследований, связанные с базовыми анатомическими представлениями о структуре мозга. Однако такой диагностический метод должен иметь высокую степень временного разрешения, чтобы иметь возможности выявлять последовательности процессов возбуждения и торможения, протекающих в структуре нейронных сетей, формирующих речь. В нашем исследовании сделана попытка разработки технологии регистрации последовательностей активации мозговых структур при слухоречевой нагрузке на основании метода решения обратной задачи ЭЭГ. Полученные результаты демонстрируют особенности нейрофизиологической регистрации ЭЭГ-сигнала, его общую реакцию на функциональную нагрузку и связь с физиологическими состояниями мозговой ткани.
Речь – основной механизм интеграции человека в общество и освоения им накопленных знаний, поэтому ее нарушения уже более 100 лет привлекают внимание исследователей как главный субстрат социального взаимодействия. В отличие от животных, социальные аспекты человеческой жизни настолько важны, что делают существование человека вне общества неполноценным не только на психологическом, но и на физическом уровне, поскольку именно социальное взаимодействие формирует в постнатальном периоде развития новые структурные элементы, необходимые для усвоения накопленных человечеством знаний.
Клинические наблюдения за людьми, утратившими речь в результате развития заболевания, и постмортальные находки позволили исследователям сформировать четкое представление о реализации речи как активности определенных участков мозга, но бурное развитие диагностических технологий в конце прошлого века заставило пересмотреть эти “устоявшиеся” воззрения и предложить новые модели реализации речевой функции, однако их внедрение в практику потребовало изменений понимания не только паталогоанатомических принципов формирования заболевания, но и новых знаний о медицинской технологии.
УНИКАЛЬНОСТЬ РЕЧИ КАК МОДЕЛИ ФУНКЦИОНАЛЬНОЙ АКТИВНОСТИ МОЗГА
Речь – филогенетически относительно молодая мозговая функция, формирование которой строго ограничено по времени. Любое нарушение этого процесса, наблюдаемое среди феральных детей или вследствие первичного поражения слуховых центров, приводит либо к отсутствию данной функции, либо к ее замещению на иные коммуникационные системы.
Речь имеет два главных функциональных центра, отвечающих за понимание внешних фонем и за продукцию собственных. Их взаимодействие обеспечивает реализацию речевой функции, а нарушение – развитие различного рода состояний, называемых афазиями.
В 1982 г. Mishkin и Ungerleider [1], изучая слуховую кору обезьян и проводя сравнения с реализацией речевой функции у человека, формулируют новую идею реализации речевой функции, получившей название “Двухпотоковой теории”, основным постулатом которой является организация человеческой речи как двух взаимосвязанных потоков информации. Данная модель показала, что в реализации речевой функции принимает участие практически весь мозг, а не ограниченное число латерализованных корковых центров, что позволило описать практически все виды клинически проявляющихся у человека афазий. Дальнейшее развитие этой идеи позволило Sporns с соавт. [2] ввести новый термин – мозговой коннектом как общую связь между функциональными элементами в модели многоуровневой функциональной нейронной сети [3].
В результате, на сегодняшний день в научной среде возникла острая необходимость в объективно-доказательных методах, позволяющих исследовать речь как комплекс, состоящий из речевых центров и речевых связей, и способных выявлять последовательности их активаций при реализации речевой функции [4].
РАЗВИТИЕ МЕТОДОВ ИССЛЕДОВАНИЯ РЕЧЕВОЙ ФУНКЦИИ
Работы Broca и Wernicke, положившие основу научному исследованию речевой функции, реализовывали подтверждения своих клинических наблюдений последующим патоморфологическим подтверждением дефекта мозговых структур [5, 6]. Но уже к началу ХХ века стало ясно, что такой подход не позволяет произвести большие исследования на уровне мономорфных групп, поэтому внимание исследователей сосредоточилось на клинических методах исследований с помощью различных тестов и их объединений. Наиболее впечатляющих результатов в этих исследованиях добились представители школы Выгодского, а Лурия практически завершил их совершенствование, доказав, что в формировании речи наиболее важна связь активности всех речевых центров, что и формирует понятие единой речевой функции. Однако клинический метод, при всех его достоинствах, оказался чрезвычайно зависимым от субъективных факторов, а убедительных методов инструментального подтверждения клинических находок, основанных на иных, чем патоморфологическое исследование принципах, еще не было разработано [7].
В 40-х годах прошлого века Penfild, стараясь преодолеть ограничения объективных методик, построенных на постмортальном исследовании или нейрохирургическом вмешательстве, обращает внимание на развитие функциональных нарушений у больных фокальной эпилепсией и предлагает новый неинвазивный подход к исследованию функциональной активности головного мозга – “Эпилептический метод”. В 1957 г. Landau и Kleffner доказали его эффективность, открыв специфическую форму эпилептической афазии [8].
В 1980 г. Kutas и Hillyard [9] обнаружили, что восприятие слуховой парадигмы с семантическими ошибками вызывает появление негативного пика через 400 мс после предъявления стимула N400, который они связали с процессом лексико-семантической интеграции слова в контекст. Использование парадигм с синтаксическими и морфосинтаксическими аномалиями, проведенное Osterhout, Holcomb и Swinney в 1992 г., сопровождалось регистрацией позитивного потенциала через 600 мс после предъявления стимула, возникновение которого было связано с трудностями обработки синтаксической информации и процессом синтаксического реанализа [10, 11].
В настоящее время методики регистрации вызванных потенциалов активно используются для получения объективной информации о механизмах реализации речи [7, 12]. Однако такой подход не позволяет сформировать представления о реализации речевой функции как непрерывной последовательности в структуре нейронной сети и ее связи с базовыми анатомическими образованиями мозга [13].
Активно развивающаяся магнитно-резонансная томография (МРТ) внутричерепных структур, дополненная в 80-х годах прошлого века технологией определения BOLD (Вlood Оxygen Level Dependent)-сигнала, позволила визуализировать основные мозговые центры, активно участвующие в реализации речевой функции, а применение методов диффузионно взвешенных изображений сместило интерес исследователей от изучения отдельных невральных структур к исследованию их функционального взаимодействия. Тем не менее, из-за высокой инертности, функциональная МРТ (фМРТ) не позволяет отследить динамику изучаемого процесса. Решение этого вопроса представляется в виде создания гибридных технологий с попыткой получать взаимодополняющую информацию об изучаемых объектах, связях мозговых центров и последовательностях их активации, однако такие системы чрезвычайно подвержены воздействию различных артефактных сигналов, возникающих в процессе реализации исследования, что фактически нивелирует все их преимущества [14].
Таким образом, в настоящее время требуется разработка объективного метода функционального исследования, позволяющего выявлять быстропротекающие процессы в нейронных сетях головного мозга, фиксировать их и формировать результаты исследований, связанные с базовыми анатомическими представлениями о структуре мозга [15].
ТЕОРЕТИЧЕСКИЕ ОСНОВЫ ФУНКЦИОНАЛЬНОГО ИССЛЕДОВАНИЯ РЕЧЕВОГО КОННЕКТОМА НА СОВРЕМЕННОМ ЭТАПЕ
Как было сказано выше, использование физических принципов построения фМРТ изображений не отвечают временным требованиям, необходимым для исследования реализующих последовательностей работы речевого коннектома. Это ограничение можно частично обойти, выбирая методы реализации простых функциональных заданий, но тем не менее, в результате исследователь получает суммарную картину активации всей нейронной сети.
Использование стимуляционных нейрофизиологических методик, выявляющих общие ответы на унифицированный раздражитель с амплитудной суммацией реакции на повторный стимул, требует физиологической конгруэнтности нейронных сетей, формирующих речевой коннектом, что в реальных условиях не реализуемо со стороны анатомических особенностей каждого индивидуума. В результате такие исследования дают обобщенную информацию по активности главных нейронных центров, которая хорошо сочетается и гибридизируется с данными фМРТ, но по информативности не превосходит их. Также в организации данного исследования весьма трудно реализовать исследование реакции мозга именно на морфо-фонетические сочетания, которые формируют человеческую речь, избегая ложной реакции на избранные стимулы и ритмы.
Решением данного методического вопроса является анализ периодов стабилизации спектральных характеристик общего энергетического поля скальпа – ЭЭГ-микросостояний. Во время анализа спектральных ЭЭГ-топограмм Lehmann с соавт. в 1998 г. обратил внимание на феномен дискретности перехода одной энергетической конфигурации скальпового поля к другой, во время которого одно стабильное состояние фиксировалось в течение некоего периода времени, а затем быстро сменялось другим, имеющим свои временные характеристики без переходных вариантов [16]. Этот феномен был интерпретирован на представлениях, согласно которым стабильное состояние генерируется скоординированной активностью отдельных нейронных макросетей, выполняющих одну или схожие функции. Это позволило выделить несколько базовых характеристик описания каждого из микросостояний, таких как 1) процент вовлеченности микросостояния в формирование спектрального поля скальпа, характеризующий активную нейронную сеть, формирующую микросостояние; 2) продолжительность отдельного микросостояния, отражающую сохранность структуры этой макросети, и 3) частота представленности отдельного микросостояния в секунду, характеризующее передачу активности между отдельными макросетями [17].
Таким образом, в настоящее время последовательность ЭЭГ-микросостояний рассматривается исследователями как прямое отражение состояния базовых механизмов реализации высших нервных функций посредством последовательной активации нейронных макросетей головного мога [18].
Цель данной работы заключалась в попытке предложить новый методический подход к исследованию динамики мозговых процессов, лежащих в основе речевой функции человека, с помощью современных нейрофизиологических методов регистрации последовательностей динамических процессов, протекающих в отдельных нейронных макросетях головного мозга.
Важность данной проблемы обусловлена тем фактом, что на сегодняшний момент не существует методик, способных оценить полную нейрофизиологическую картину реализации речевой функции в условиях реального течения времени.
МЕТОДЫ ИССЛЕДОВАНИЯ
Испытуемые
Основная группа исследования была составлена из 33 праворуких русскоговорящих здоровых взрослых, не имевших скрытых нарушений когнитивного статуса и скрытого течения внутренних болезней.
Возрастной диапазон участников исследования колебался от 19 до 60 лет, средний возраст составил 32.4 ± 9.4 лет. По гендерному признаку добровольцы распределялись следующим образом: 11 женщин (33%) и 12 (36%) мужчин. Все обследуемые имели высшее образование – 33 (100%); на момент исследования все (100% участников исследования) работали на производстве, связанном с умственной деятельностью. Русский язык для всех участников был родным.
Этические нормы и правила
Каждому участнику был разъяснен смысл и цели исследования, а также возможные побочные эффекты. Все процедуры, выполненные в исследованиях, соответствовали этическим стандартам национального комитета по исследовательской этике и Хельсинкской декларации 1964 г. с последующими изменениями. От каждого из включенных в исследование участников было получено информированное добровольное согласие. Общий план исследования был одобрен локальным Этическим комитетом ФГБУ “ФЦМН” ФМБА России и утверждено приказом от 15.06.2021 № 148-1.
Регистрация ЭЭГ
В качестве базового метода исследования была выбрана непрерывная электроэнцефалография высокой плотности записи с последующей кластеризацией и выделением отдельных классов ЭЭГ-микросостояний. Такой режим регистрации данных лучше соответствовал идее исследования и позволял минимизировать множественные мимические артефакты, связанные с реакцией на парадигму повторяющихся раздражителей (особенно на этапе нагрузки с прослушиванием речи). Запись ЭЭГ проводилась в затемненной комнате с относительной звукоизоляцией, в состоянии расслабленного бодрствования с закрытыми глазами. Изменения исходного состояния осуществлялось либо с помощью прослушивания аудиозаписи короткого рассказа на родном языке, либо при пересказе такового. Таким образом, на выходе исследователь получал три эквивалентные по времени и физиологическому состоянию испытуемого записи, содержащие как фоновое (базовое) состояние, так и реакции мозговых структур на восприятие речи и на ее активную продукцию.
Все исследования проводили в первой половине дня, с 10 до 14 ч. Накануне пациенту разъяснялись цели и задачи, а также исключался прием снотворных или стимулирующих веществ. Ночной сон составлял не менее 8 ч, при нарушении этих условий исследование переносилось на другой день.
ЭЭГ регистрировали с помощью оригинальной 128-канальной системы Hydrocel-128 (США) с усредненным референтом. Запись, коммутация и аппаратная фильтрация биоэлектрического ЭЭ-сигнала была проведена с помощью биоусилителя “EGI-GES-300” (Magstim, США). Полученный сигнал был преобразован в цифровую форму посредством дискретизации с частотой Найквиста 500 Гц, что позволило исключать искажение сигнала на частотах от 1 до 250 Гц. Полоса пропускания сигнала составляла от 0.5 до 70 Гц с включением режекторного сетевого фильтра 50 Гц, что включало основные диапазоны интереса (1–40 Гц). Подэлектродное сопротивление электродов (импеданс) не превышало 10 кОм и контролировалось в ходе всего исследования согласно рекомендациям фирмы-производителя. Помимо этого, выключались сторонние электрические приборы, создающие паразитные электромагнитные поля, контролировалось подэлектродное сопротивление интерфейса, регулировалась температура в помещении, минимизировались паразитные движения мышц путем выбора удобной позы.
В исследовании использовался весь пул из 128 активных электродов для исключения искажения получаемого результата на этапе проводимой постобработки. Применяемые ранее попытки удаления помех методом “bad electrodes reject” из записи неизменно приводили к значимым искажениям и невозможности сравнения результатов исследования ЭЭГ, поэтому было решено отказаться от данной практики на используемой в исследовании системе записи ЭЭГ. Такой подход к организации исследования позволял уменьшить влияние физических и физиологических факторов на запись и уменьшить количество первичных артефактных сигналов.
Математическая обработка и анализ полученных результатов проводились в несколько этапов. На первом этапе полученный пул данных проходил процедуру первичной фильтрации с помощью широкополосного фильтра 1–70 Гц, т.е. стандартизацию базового монтажа в единое электродное пространство. Общая энергия поля скальпа (общая структура спектра общего скальпового поля) определялась как совокупность спектральных мощностей сигнала ЭЭГ, полученных методом быстрого преобразования Фурье для каждого электрода, и интерполяцией полученных результатов на общую площадь скальповой поверхности.
На втором этапе выполнялась процедура выделения независимых компонент сигнала (ICA) с распознаванием его вида и типа через пакет прикладных программ EEGLAB R2020a (2020 г.). Эта процедура позволяла убрать различные артефакты физической и биологической природы, которые имели частотные характеристики в пределах окна фильтра предварительной фильтрации.
На третьем этапе проводилась сегментация ЭЭГ-сигнала с выделением отдельных ЭЭГ-микросостояний методом кластеризации К-средних или методом адгезии – распыления с выделением 6 типов отдельных микросостояний (4 класса стабильных и два класса дополнительных), учитывая рекомендации Koening по вариабельности 5-го и 6-го класса и использованием плагина Microstates программного комплекса EEGLAB по методике [20] с учетом эквивалентной полярности. Выбор 6 классов ЭЭГ-микросостояний вместо 4 был сделан для возможности более полного анализа изменений биоэлектрической активности, регистрируемых во время реализации речевой функции с учетом двухпотоковой модели ее организации в головном мозге.
Заключительный этап исследования включал анализ локализации источника активности каждого из выделенных классов ЭЭГ-микросостояний по методике Pascual-Marqui (пакет прикладных программ sLORETA), методом анализа изменений спектральных характеристик энергии скальпового поля [21, 22].
В результате подобной системы обработки сигнала была получена информация о 6 отдельных классах ЭЭГ-микросостояний, включающая: время жизни микростостояния (Duration) – в мкс; частоту его регистрации в 1 с (Occurrence); объема вклада ЭЭГ-микросостояния в общую структуру спектральной плотности скальпового поля (Coverage), а также информацию об активности основного коркового поля, согласно атласа Бродмана, формирующего текущее ЭЭГ-микросостояние.
Статистическая обработка полученных результатов
Статистический анализ полученных результатов производился с помощью статистического пакета GNU-PSPP под управлением ОС Linux Mate 10.10. Общая структура статистического анализа включала следующие этапы. Проверку данных на достоверность и соответствия полученных данных модели альфа с помощью метода Кронбаха (0.5 < альфа), после которой для дальнейшего анализа использовался метод факторного анализа для выделения ведущих факторов. Для сравнения результатов применялся метод парного t-сравнения, поскольку изменения оценивались в одной группе наблюдения, соответственно поправка Бонферрони на множественность сравнений не вводилась из-за малого количества таковых. Во всех расчетах использовалась одна степень свободы, уровень доказательности был принят как альфа >0.05. Вся методика анализа была разработана согласно рекомендациям [24].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Анализ ЭЭГ-микросостояний в период расслабленного пассивного бодрствования с закрытыми глазами показал общую нестабильность последовательностей всех классов ЭЭГ-микросостояний. На рис. 1 показано процентное соотношение вклада каждого из классов ЭЭГ-микросостояний в структуру спектра общего скальпового поля во время пребывания участников эксперимента в состоянии пассивного расслабленного бодрствования.
Рис. 1.
Характеристика показателя вклада каждого из 6 классов ЭЭГ-микросостояний в формирование спектральной плотности скальпового поля в состоянии пассивного расслабленного бодрствования.
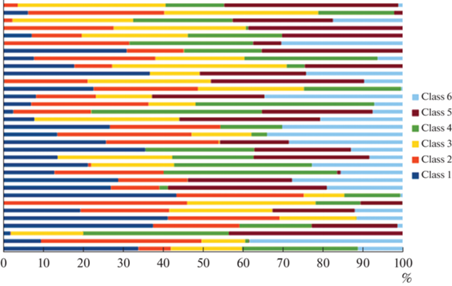
Как видно из представленного рисунка, регистрировался выраженный полиморфизм данного показателя по всем 6 исследованным классам ЭЭГ-микросостояний, формирующий хаотичное распределение на пространственной фигуре.
Однако в случае изменения условий окружающей среды, связанного с прослушиванием испытуемым текста на родном языке, картина представленности вклада всех классов ЭЭГ-микросостояний изменилась в сторону более упорядоченной последовательной структуры (рис. 2 и 3).
Рис. 2.
Нормализация и упорядочивание последовательностей 6 классов ЭЭГ-микросостояний в условиях прослушивания речи на родном языке.
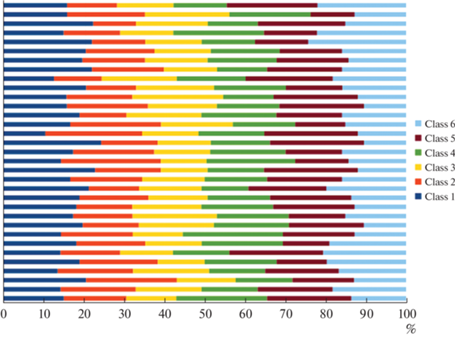
Рис. 3.
Нормализация и упорядочивание последовательностей 6 классов ЭЭГ-микросостояний в условиях активной речевой продукции на родном языке.
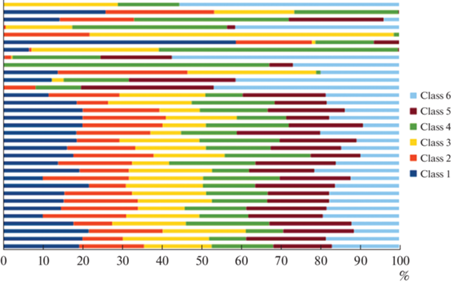
Активная речевая продукция также вызывала формирование упорядоченной структуры последовательностей ЭЭГ-микросостояний, однако у части обследуемых (10 человек) наблюдались выраженные артефакты вследствие активности артикуляционных и мимических мышц, искажающих запись и неустранимых с помощью стандартно выполняемых процедур фильтрации и независимого анализа компонент. Данные результаты представляли именно физиологические особенности, так как исследования проводилось на единой аппаратуре, в близкие моменты времени, но в разные дни, наряду с участниками, продемонстрировавшими упорядоченную структуру организации микросостояний.
Сравнительная характеристика ЭЭГ-микросостояний, регистрируемых во время реализации речевой функции
Сравнительный анализ характеристик ЭЭГ-микросостояний в состоянии расслабленного бодрствования, прослушивания и активной речевой продукции, проведенный с помощью парного t-теста показал, что все три основные показателя – продолжительность существования, частота встречаемости и процент вклада, статистически значимо (p < 0.05) отличались во время прослушивания или активной речевой продукции от состояния пассивного расслабленного бодрствования. В то же время, статистически достоверная разница между величинами основных показателей состояния прослушивания и активной речевой продукции отсутствовала (рис. 4–6).
Анализ источника активности микросостояний посредством решения обратной задачи ЭЭГ и локализации в соответствии с атласом полей Бродмана
На основании полученных данных о существовании достоверной реакции мозговых сетей на слухоречевое раздражение, было проведено исследование каждого из микросостояний с целью выделения основного источника, ответственного за формирование регистрируемого класса текущего микросостояния, с помощью процедуры анализа источников сигнала, реализованного в программном пакете eLORETA.
Рис. 4.
Парное сравнение изменений продолжительности (Duration) в секундах. 1–6-е классы ЭЭГ‑микросостояний в состоянии пассивного расслабленного бодрствования и при слухоречевой стимуляции, пара покой–прослушивание – p < 0.001; пара покой–активная речь – p = 0.002; пара прослушивание–активная речь – p = 0.9 (в представлении усредненных значений по классу ЭЭГ-микросостояний).
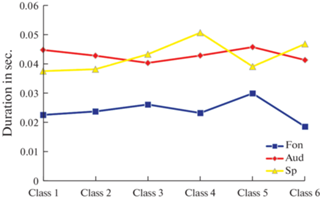
Рис. 5.
Парное сравнение изменений частот встречаемости (Occurence) 1–6 классов ЭЭГ-микросостояний в течение 1 с в состоянии пассивного расслабленного бодрствования и при слухоречевой стимуляции, пара покой–прослушивание – p < 0.001; пара покой–активная речь – p < 0.001; пара прослушивание–активная речь – p = 0.005 (в представлении усредненных значений по классу ЭЭГ-микросостояний).
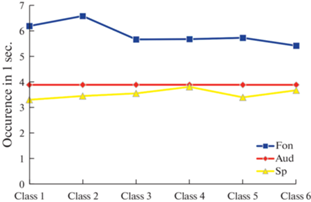
Рис. 6.
Парное сравнение изменений показателя вклада (Coverage) каждого из классов ЭЭГ-микросостояний 1–6 классов в структуру общей спектральной плотности общего поля скальпа (в %) в состоянии пассивного расслабленного бодрствования и при слухоречевой стимуляции. Пара покой–прослушивание – p < 0.005; пара покой–активная речь – p < 0.001; пара прослушивание–активная речь – p = 0.12 (в представлении усредненных значений по классу ЭЭГ-микросостояний).
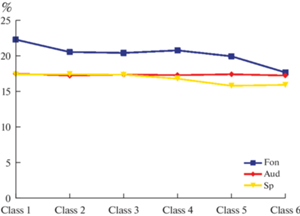
Поскольку каждое из выделенных в ходе исследования ЭЭГ-микросостояний представляло собой относительно устойчивое во времени изменение структуры спектральной характеристики поля скальпа, решение обратной задачи ЭЭГ по методике [21] позволило осуществить построение трех основных последовательностей активации полей Бродмана у каждого обследованного, которые были объединены в общие матрицы, отражающие состояние фоновой активности при пассивном расслабленном бодрствовании, слуховой нагрузке и речевой продукции для всех обследованных (рис. 7–9).
Рис. 7.
Последовательности регистрации “ритмической активности” полей Бродмана в условиях пассивного расслабленного бодрствования у обследованных, согласно классу ЭЭГ-микросостояния. (Выделены поля Бродмана, над которыми была зарегистрирована ритмическая ЭЭГ-активность).

Рис. 8.
Последовательности регистрации “ритмической активности” полей Бродмана в условиях прослушивания у обследованных, согласно классу ЭЭГ-микросостояния. (Выделены поля Бродмана, над которыми была зарегистрирована ритмическая ЭЭГ-активность).
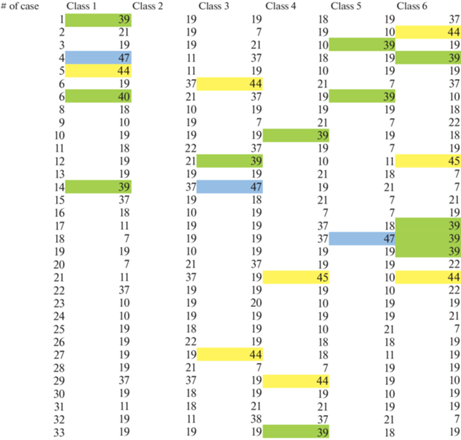
Рис. 9.
Последовательности регистрации “ритмической активности” полей Бродмана в условиях активной продукции речи на родном языке у обследованных, согласно классу ЭЭГ-микросостояния. (Выделены поля Бродмана, над которыми была зарегистрирована ритмическая ЭЭГ-активность).
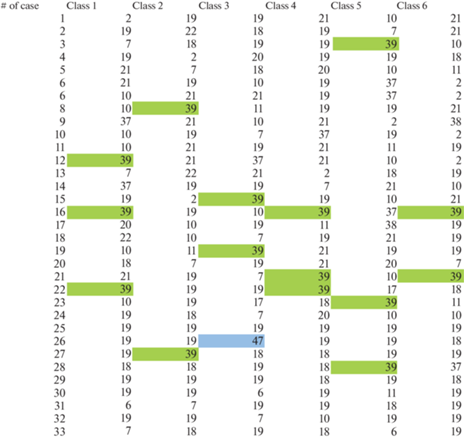
Решение обратной задачи ЭЭГ с выделением источника для каждого из исследуемых классов ЭЭГ-микросостояний (согласно представления корковых полей Бродмана) показало следующие результаты. Так, в состоянии пассивного расслабленного бодрствования преимущественно выделялось поле 47, ответственное за музыкальное восприятие, а поля речевого анализатора не демонстрировали ритмической активности. Однако во время прослушивания текста выделялись как 44-е и 45-е поля Бродмана, соответствующих корковой локализации центра Брока, так и поля 39, 40, характеризующих корковое представительство центра Вернике.
В то же время при активной речи 44-е и 45 е поля Бродмана не определись, но при этом регистрировалась ритмическая активность полей 39 и 40 (табл. 1).
Таблица 1.
Регистрация “ритмической активности” полей Бродмана в состоянии пассивного расслабленного бодрствования и во время слухоречевой нагрузки (данные 33 наблюдений)
| Абс. количество |
Процент | |
|---|---|---|
| “Активность” Центра Брока при прослушивании [поля 44, 45] | 8 | 24% |
| “Активность” Центра Вернике при прослушивании [поля 39, 40] | 8 | 24% |
| “Активность” Центра Брока при активной речи [поля 44, 45] | 0 | 0% |
| “Активность” Центра Вернике при активной речи [поля 39,40] | 16 | 48% |
| “Активность” центра музыки при прослушивании [поле 47] | 3 | 9% |
| “Активность” центра музыки при активной речи [поле 47] | 1 | 3% |
| “Активность” центра музыки в покое [поле 47] | 16 | 47% |
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Нестабильность ЭЭГ-микросостояний в период пассивного расслабленного бодрствования было описано ранее в работах [20, 25]. Однако нами был выявлен феномен формирования их спонтанной организации при предъявлении слухоречевой нагрузки. Возможно, имеет место не самоорганизация 6 классов ЭЭГ-микросостояний покоя, а организация нативного ЭЭГ-сигнала в ответ на функциональное раздражение с включением специфических нейронных сетей, требуемых для реализации выбранной функции, что подтверждалось результатами парного t-теста, продемонстрировавшими статистически значимые различия показателей ЭЭГ периода пассивного расслабленного бодрствования и ЭЭГ-периодов аудиоречевой стимуляции. Данные изменения характеризуются формированием упорядоченных последовательностей ЭЭГ-микросостояний различных классов, что, вероятно, характеризует существование единых путей передачи возбуждения между отдельными корковыми нейронными структурами, принимающими участие в реализации речевой функции. Несмотря на общую упорядоченность, эти последовательности различны для состояний прослушивания и активной речевой продукции, и, вероятно, уникальны у каждого человека в отдельности. Такая уникальность может быть как генетически детерминированной, так и развиваться вследствие уникальности развития каждого человека. Также не исключено, что эти два фактора могут оказывать сложное комбинированное влияние на процесс формирования человеческой речи.
В то же время решение обратной задачи ЭЭГ с помощью процедур локализации источника сигнала [17], основанных на данных спектральной плотности энергетического поля скальпа, не может быть гибридизировано с результатами фМРТ исследования из-за принципиальных технологических особенностей технологии исследования, демонстрирующего изменения МР характеристик гемоглобина, теряющего молекулу кислорода в функционально активных зонах.
В то же время процедура локализации источника сигнала путем решения обратной задачи ЭЭГ учитывает в качестве исходных данных характеристики спектральной плотности энергетического поля скальпа, которые выше у участков мозговой коры, продуцирующих ритмическую активность, и ниже у участков, продуцирующих низкоамплитудные хаотические сигналы [26].
Первоначальная специализация данного алгоритма, предназначенная для выявления патологических ЭЭГ-ритмов объемных внутричерепных образований или эпилептических очагов, не позволяет выделить состояние возбуждения физиологически нормальной нервной ткани, характеризующейся общей десинхронизацией и дезорганизацией ритмической активности, отражающейся на показателях спектральной плотности поля скальпа в качестве участков с минимально выраженной активностью. Данная находка объясняет большинство неудачных попыток ЭЭГ-картирования мозговых структур из-за отсутствия соответствия ЭЭГ данных, в том числе и полученных в ходе решения обратной задачи ЭЭГ, с результатами фМРТ, поскольку эти методики фиксируют совершенно различные состояния нервной ткани. фМРТ регистрирует участки повышенной активности на основании высокого потребления кислорода в фиксированном объеме нервной ткани, в то время как ЭЭГ фиксирует участок, продуцирующий специфический ритм, а не пребывающий в состоянии возбуждения.
Продукция ЭЭГ-ритма, вероятно [26], характеризует синхронную активность гигантских пирамидных клеток, которая предшествует активации полиморфного слоя мозговой коры, формируя специфическое состояние “ожидания” активности нервного центра, предшествующее функциональному возбуждению. В то же время, истинное возбуждение коркового представительства невральной функции, характеризующееся большим количеством разнонаправленных постсинаптических потенциалов полиморфного слоя коры, не будет фиксироваться из-за малой спектральной плотности характерной для хаотических и псевдохаотических процессов.
При анализе результатов, полученных при исследовании последовательностей активации полей Бродмана, важный вопрос составляла точность полученных предлагаемым методом результатов. Согласно нашим данным, точность составляла 40–50%, что в целом характерно для самого ЭЭГ-метода, используемого в качестве базовой технологии. Однако требуются дальнейшие исследования, чтобы окончательно ответить на данный вопрос с использованием более строгих критериев отбора в формировании основной группы исследования или применения близнецового метода.
ЗАКЛЮЧЕНИЕ
На основании проведенных исследований была разработана методика выделения отдельных последовательностей возбуждения полей Бродмана, соответствующих реализации речевой функции. Однако данная методика рассматривает процессы в корне отличные от процессов, регистрируемых фМРТ, фактически технология решения обратной задачи ЭЭГ позволяет определить не активные центры мозгового коннектома, участвующие в реализации функции в текущий момент времени, а те образования, которые мозг подготавливает к процессу реализации функции. В этих условиях исследование человеческой речи представляет уникальные возможности для исследователей, позволяющих более полно понять сам процесс работы головного мозга и формирования мышления.
Список литературы
Mishkin M, Ungerleider LG (1982) Contribution of striate inputs to the visuospatial functions of parieto-preoccipital cortex in monkeys. Behav Brain Res 6.1.57.77. https://doi.org/10.1016/0166.4328.82.90081
Sporns O, Tononi G, Kotter R (2005) The human connectome: A structural description of the human brain. PLoS Comput Biol 1(4): e42. https://doi.org/10.1371/journal.pcbi.0010042
Howell BR, Styner MA, Gao W (2018) The UNC/UMN Baby Connectome Project [BCP]: An overview of the study design and protocol development. Neuroimage 185.891.905. https://doi.org/10.1016/j.neuroimage.2018.03.049
Sato M (2020) The neurobiology of sex differences during language processing in healthy adults: A systematic review and a meta-analysis. Neuropsychologia 16.140.107404. https://doi.org/10.1016/j.neuropsychologia.2020.107404
Broca P (2011 [1861]) Remarks on the Seat of Spoken Language, Followed by a Case of Aphasia. Neuropsychol Rev 21.3]15: 227–229. https://doi.org/10.1007/s11065-011-9174-x
Wernicke K (1897) Grundriss der Psychiatrie in klinischen Vorlesungen. Theil II, Die paranoischen Zustände. Leipzig: Verlag von Georg Thieme. Am J Psychiatry 53[4]: 581–583. https://doi.org/10.1176/ajp.53.4.581
Карлов ВА, Шкловский ВМ, Золовкина ВС (2017) Развитие представлений об организации речевой системы. Журн неврол психиатр им СС Корсакова 117(5): 4–8. [Karlov VA, Shklovsky VM, Zolovkina VS (2017) Development of ideas about the organization of the speech system. J Neurol Psychiatry SS Korsakov 117(5): 4–8. (In Russ)]. https://doi.org/10.17116/jnevro2017117514-8
Королёва НВ, Колесников СИ, Воробьев СВ (2011) Электроэнцефалографический атлас эпилепсий и эпилептических синдромов у детей. ГЭОТАР-Медиа. [Koroleva NV, Kolesnikov SI, Vorobyov SV (2011) Electroencephalographic atlas of epilepsy and epileptic syndromes in children. GEOTAR-Media. (In Russ)]. ISBN 978-5-4235-0047-4
Kutas M, Hillyard SA (1984) Brain potentials during reading reflect word expectancy and semantic association. Nature 12-18; 307(5947): 161–163. https://doi.org/10.1038/307161a0
Osterhout L, Holcomb PJ, Swinney DA (1994) Brain potentials elicited by garden-path sentences: evidence of the application of verb information during parsing. J Exp Psychol Learn Mem Cogn 20[4]: 786–803. https://doi.org/10.1037//0278-7393.20.4.786
Alday PM, Kretzschmar F (2019) Speed-Accuracy Tradeoffs in Brain and Behavior: Testing the Independence of P300 and N400 Related Processes in Behavioral Responses to Sentence Categorization. Front Hum Neurosci 27 (13) 285. https://doi.org/10.3389/fnhum.2019.00285
Емелина ДА, Макаров ИВ, Гасанов РФ (2019) Методика вызванных потенциалов головного мозга в исследовании специфических расстройств речи у детей. Соц клин психиатрия 29(2): 104–111. [Emelina DA, Makarov IV, Gasanov RF (2019) Method of evoked potentials of the brain in the study of specific speech disorders in children. Soc Clin Psychiatry 29(2): 104–111. (In Russ)]. ISSN: 0869-4893
Kearney E, Guenther FH (2019) Articulating: The Neural Mechanisms of Speech Production. Lang Cogn Neurosci 34(9): 1214–1229. https://doi.org/10.1080/23273798.2019.1589541
Bullock M, Jackson GD, Abbott DF (2021) Artifact Reduction in Simultaneous EEG-fMRI: A Systematic Review of Methods and Contemporary Usage. Front Neurol 11;12: 622719. https://doi.org/10.3389/fneur.2021.622719
Журавлев ИВ, Нистратов АА, Поликарпов МА, Тарасов ЕФ (2019) Картирование речевых зон мозга как научная проблема. Вопр психолингвистики 3(41): 107–121. [Zhuravlev IV, Nistratov AA, Polikarpov MA, Tarasov EF (2019) Mapping of speech areas of the brain as a scientific problem. Quest Psycholinguistics 3(41): 107–121. (In Russ)]. https://doi.org/10.30982/2077.5911.2019.41.3.107.121
Lehmann D, Strik WK, Henggeler B, Koenig T, Koukkou M (1998) Brain electric microstates and momentary conscious mind states as building blocks of spontaneous thinking: I. Visual imagery and abstract thoughts. Int J Psychophysiol 29(1): 1–11. https://doi.org/10.1016/s0167-8760(97)00098-6
Lehmann D, Faber PL, Gianotti LR, Kochi K, Pascual-Marqui RD. (2005) Coherence and phase locking in the scalp EEG and between LORETA model sources, and microstates as putative mechanisms of brain temporo-spatial functional organization. J Physiol (Paris) 99(1): 29–36. https://doi.org/10.1016/j.jphysparis.2005.06.005
Van De Ville D, Britz J, Michel CM (2010) EEG microstate sequences in healthy humans at rest reveal scale-free dynamics. Proc Natl Acad Sci USA 107 (42): 18179–18184. https://doi.org/10.1073/pnas.1007841107
Abreu R, Jorge J, Leal A, Koenig T, Figueiredo P (2020) EEG Microstates Predict Concurrent fMRI Dynamic Functional Connectivity States. Brain Topogr 34(1): 41–55. https://doi.org/10.1007/s10548-020-00805-1
Mishra A, Englitz B, Cohen MX (2020) EEG microstates as a continuous phenomenon. Neuroimage 208: 116454. https://doi.org/10.1016/j.neuroimage2019.116454
Pascual-Marqui RD (2002) Standardized low-resolution brain electromagnetic tomography [sLORETA]: technical details. Methods Find Exp Clin Pharmacol 24 Suppl D: 5–12.
Lu G, Hou Y, Chen Y, Guo F (2022) Neuroimaging of EEG Rhythms at Resting State in Normal Elderly Adults: A Standard Low-Resolution Electromagnetic Tomography Study. J Clin Neurophysiol 1;39(1): 72–77. https://doi.org/10.1097/WNP.0000000000000780
Khanna A, Pascual-Leone A, Michel CM, Farzan F (2015) Microstates in resting-state EEG: current status and future directions. Neurosci Biobehav Rev 49: 105–113. https://doi.org/10.1016/j.neubiorev.2014.12.010
Sarter M, Fritschy JM (2008) Reporting statistical methods and statistical results in EJN. Eur J Neurosci 28(12): 2363–2364. PMID: https://doi.org/10.1111/j.1460-9568.2008.06581.x19087166
He B, Astolfi L, Valdes-Sosa PA, Marinazzo D, Palva S, Benar CG, Michel CM, Koenig T (2019) Electrophysiological Brain Connectivity: Theory and Implementation. IEEE Trans Biomed Eng. https://doi.org/10.1109/TBME.2019.2913928
Choi J, Lim E, Park MG, Cha W (2020) Assessing the Retest Reliability of Prefrontal EEG Markers of Brain Rhythm Slowing in the Eyes-Closed Resting State. Clin EEG Neurosci 51(5): 348–356. https://doi.org/10.1177/1550059420914832
Дополнительные материалы отсутствуют.
Инструменты
Российский физиологический журнал им. И.М. Сеченова