

Российский физиологический журнал им. И.М. Сеченова, 2023, T. 109, № 2, стр. 214-228
Применение метода цветового кодирования для анализа эффектов галоперидола на поведение животных в тесте “Открытое поле”
М. С. Макаров 1, Ю. И. Сысоев 2, 3, 4, 5, *, М. К. Гузенко 1, В. А. Приходько 2, 5, Э. Коркотян 1, 6, **, С. В. Оковитый 2, 5
1 Пермский государственный национальный исследовательский университет
Пермь, Россия
2 Санкт-Петербургский государственный химико-фармацевтический университет
Санкт-Петербург, Россия
3 Институт физиологии им. И.П. Павлова РАН
Санкт-Петербург, Россия
4 Институт трансляционной биомедицины, Санкт-Петербургский государственный университет
Санкт-Петербург, Россия
5 Институт мозга человека им. Н.П. Бехтеревой РАН
Санкт-Петербург, Россия
6 Институт им. Вейцмана
Реховот, Израиль
* E-mail: susoyev92@mail.ru
** E-mail: eduard.korkotian@weizmann.ac.il
Поступила в редакцию 17.09.2022
После доработки 18.12.2022
Принята к публикации 09.01.2023
- EDN: NDUCZC
- DOI: 10.31857/S0869813923020085
Аннотация
Разработка новых подходов к анализу данных поведенческих тестов имеет важное значение как для фундаментальных исследований, так и для оценки эффектов фармакологических агентов в экспериментальной нейрофармакологии. Одной из фармакологических групп, представляющей большой интерес для изучения, являются антипсихотические средства, применяемые для лечения различных психических заболеваний (например, расстройств шизофренического спектра). Несмотря на то, что данные препараты оказывают выраженное влияние на поведение животных в простых поведенческих тестах, их эффект часто трудно отличить от действия других препаратов, угнетающих центральную нервную систему, например, седативных средств. В настоящей работе предложен метод анализа структуры поведенческих паттернов у крыс с использованием приема цветового кодирования видеоряда на примере эффектов типичного нейролептика галоперидола в тесте “Открытое поле”. Эксперименты были выполнены на белых беспородных крысах-самцах массой 250–300 г. С помощью языка программирования Python в среде Google Colab 3-минутные видеозаписи поведения крыс в тесте “Открытое поле” обрабатывали с применением библиотеки OpenCV. Цветовое кодирование позволило маркировать разными цветами и совмещать в одном кадре настоящее (t = 0), ближайшее будущее (t = 0.33 с) и несколько более отдаленное будущее положение животного в пространстве (t = 1.66 с). При использовании предлагаемого метода было обнаружено специфическое влияние галоперидола на паттерн поведения крыс, не выявляемое с использованием традиционных подходов к анализу поведения животных в данном тесте. Примененный метод цветового кодирования с использованием трех временных точек показал эффективность для анализа и обработки данных, оказавшись сопоставимым с ранее описанными в литературе подходами с применением как традиционных, так и компьютеризированных методов визуализации.
ВВЕДЕНИЕ
Одним из распространенных и не до конца изученных психических расстройств, которым страдает около 1% населения, является шизофрения [1]. Несмотря на то, что механизмы ее развития не вполне понятны, предложен ряд фармакологических мишеней, действуя на которые, антипсихотические средства могут купировать симптомы этого заболевания [2]. Тем не менее, используемые в настоящее время препараты не оправдывают в полной мере ожидания врачей и пациентов, поскольку недостаточно действуют на так называемые “минус”-симптомы и когнитивные нарушения. Более того, при приеме типичных антипсихотических средств у больных неизбежно развиваются такие нежелательные лекарственные реакции, как паркинсонизм и дистония [3]. Все это определяет важность поиска новых препаратов, направленных на лечение шизофрении.
Для выполнения данной задачи необходимы эффективные методы скрининга новых потенциально активных соединений. Большое значение имеют поведенческие тесты с использованием мелких лабораторных животных, поскольку такие эксперименты моделируют отдельные аспекты реальной клинической ситуации. Кроме того, поведенческие тесты не требуют больших материальных затрат, позволяя при этом ответить на вопрос, является ли изучаемое соединение психоактивным [4].
Выявление именно антипсихотической активности у потенциальной молекулы не является рутинной задачей. Так, если действие анксиолитических или, например, седативных препаратов можно выявить в тестах “Открытое поле” (ОП) или “Приподнятый крестообразный лабиринт” без какой-либо подготовки испытуемых животных, то в случае потенциального нейролептика необходимо предварительное моделирование шизофрении. На сегодняшний день предложено множество моделей этого заболевания, которые можно разделить на фармакологические, хирургические и генетические [5]. Каждая из моделей достаточно неплохо отражает те или иные симптомы шизофрении у людей, однако все они требуют затрат времени экспериментатора и, самое главное, расхода лабораторных животных.
Возникает справедливый вопрос: возможно ли выявление специфического антипсихотического действия у препарата с использованием простых поведенческих установок без предварительного моделирования заболевания? Для этого необходим поиск новых подходов к анализу данных, получаемых в ходе видеорегистрации поведения животных. Важно, что одной из основных трудностей в интерпретации результатов поведенческих тестов остается субъективность оценки тех или иных критериев разными исследователями, а также излишнее упрощение и однонаправленность выбранных методов анализа [6].
В настоящем исследовании использован метод выявления динамических паттернов поведения с помощью цветового кодирования видеоряда для анализа влияния галоперидола на поведение у лабораторных крыс в тесте ОП. Галоперидол был выбран в качестве эталонного типичного антипсихотика, обладающего сильным антипсихотическим действием без выраженного седативного эффекта [7, 8]. В качестве установки для оценки поведения испытуемых животных был выбран тест ОП как наиболее простой и часто используемый при изучении поведения мелких лабораторных животных [9].
МЕТОДЫ ИССЛЕДОВАНИЯ
Эксперименты проведены на 20 белых беспородных крысах-самцах массой 250– 300 г. Крыс содержали в стандартных условиях при 12-часовом цикле свет/темнота и со свободным доступом к воде и корму.
Животные были разделены на 2 группы (опытная и контрольная) по 10 особей. Опытным животным вводили раствор галоперидола (ООО “Велфарм”, Россия) внутрибрюшинно в дозе 0.3 мг/кг в объеме 0.5 мл. В качестве контроля вводили физиологический раствор в том же объеме. Время экспозиции препарата составляло 20 мин [10], после чего крысу помещали в установку ОП (ООО “НПК Открытая наука”, Россия), представлявшую собой круглую арену черного цвета диаметром 97 см, расчерченную на 19 одинаковых по площади сегментов и оборудованную отверстиями в полу. Видеорегистрацию проводили в течение 3 мин с помощью камеры Canon 5D (Canon Inc., Япония) при частоте 30 кадров в секунду и уровне освещенности 250 люкс (с равномерным освещением всей площади арены без градиента освещенности). Участок помещения с тестовой установкой ограничивали от остального рабочего пространства при помощи однотонных медицинских ширм. Управление работой камеры происходило удаленно при помощи программного обеспечения EOS Utillity (Canon Inc., Япония); изображение выводилось на монитор экспериментатора.
При анализе видеоматериалов фиксировали следующие поведенческие паттерны: повороты головы/туловища вправо, влево, движение вперед, разворот назад, стойка на задних лапах с опорой на стенку и без опоры, обследование отверстий, обнюхивание пространства, груминг и замирание. Валидация общей активности на основе подсчета паттернов производилась стандартным методом ручного подсчета пересечений секторов. Часть видеозаписей просматривали в исходном виде, при этом выделяли и фиксировали основные паттерны. Далее с помощью языка программирования Python в среде Google Colab записи обрабатывали с применением библиотеки OpenCV. При помощи цветового кодирования отображали различными цветами и совмещали в одном кадре настоящее (t = 0), ближайшее будущее (t = 0.33 с) и несколько более отдаленное будущее (t = 1.66 с) положения животного.
Статистическую обработку данных проводили с использованием t-критерия Стьюдента или U-критерия Манна–Уитни при помощи программ PAST 4.03 и Excel 2016 (Microsoft, США). Нормальность распределения данных в выборках определяли с помощью критерия Шапиро–Уилка. При изучении зависимости двигательной активности от фактора времени использовали двухфакторный дисперсионный анализ ANOVA для повторных изменений с последующим попарным сравнением post hoс на основе критерия Тьюки. Различия между группами признавались статистически значимыми при р < 0.05. Данные на столбчатых диаграммах приведены как среднее арифметическое ± стандартная ошибка среднего.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
Тест ОП позволяет оценить двигательную активность, ориентировочно-исследовательское поведение и уровень тревожности у крыс и мышей [11]. Анализ двигательных поведенческих паттернов является основой для теста ОП, поэтому в первую очередь из всего комплекса выявленных паттернов исследовали именно двигательные.
Среди локомоторных паттернов для оценки общей двигательной активности в центре и на периферии арены были учтены: повороты на 180°, вправо, влево, движение вперед, а также переориентация головы вправо, влево и вперед. Статистически значимые отличия в двигательной активности контрольных и получавших галоперидол животных были получены только на периферии арены. В группе галоперидола двигательная активность оказалась достоверно ниже: суммарное число паттернов за все время записи составило 19.3 ± 1.2 по сравнению с 42.2 ± 4.2 в контрольной группе (р < 0.001) (рис. 1a).
Рис. 1.
Общая двигательная активность в группах контроля и галоперидола. (a) – Среднее суммарное число двигательных паттернов в периферийных и центральных сегментах “Открытого поля” в контрольной группе и группе галоперидола за все время записи (3 мин). (b) – Изменение двигательной активности животных в контрольной группе и группе галоперидола в тесте “Открытое поле” в течение последовательных 30-секундных отрезков времени. *** – p < 0.001.
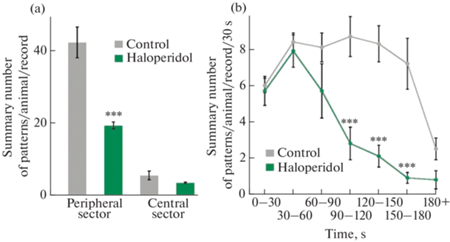
Стандартный метод определения двигательной активности с помощью подсчета пересечений секторов выявил ту же закономерность: 92.3 ± 10.5 и 32.9 ± 4.9 пересечений секторов в группе контроля и галоперидола соответственно (p < 0.001). Разница в количестве пересечений секторов для одной и той же группы объясняется на уровне подходов: при подсчете выбранных нами паттернов события фиксируются только при инициализации действий, после которых животное может продолжать движение, пересекая несколько секторов. В перспективе можно использовать оба метода для выявления нарушений в структурах, ответственных за инициализацию и планирование движений. В центральных сегментах в это же время четко прослеживалась гораздо более низкая локомоторная активность; статистически значимых отличий в центре поля между группами обнаружено не было (рис. 1a).
Беспокойное поведение (тревожность) во многих исследованиях определяется по относительной длительности нахождения животных на открытом пространстве или в центре арены по сравнению с локализацией у стенок или в укрытии [12], хотя в целом факт предпочтения периферии объясняется стремлением грызунов избегать освещенных и открытых пространств [9].
На рис. 1b представлены средние суммы паттернов за каждые последующие 30 с эксперимента. Таким образом, общее время записи было сведено к 7 временным зонам. Прослеживалась динамика подъемов и спадов двигательной активности, однако в течение первой минуты эксперимента активность у животных обеих групп была статистически неотличима. Начиная со второй минуты, у животных, получивших галоперидол, наблюдалось заметное снижение активности. В контрольной группе аналогичное снижение также имело место, однако наблюдалось оно лишь на последней минуте записи, где средние числа паттернов у крыс обеих групп вновь сближались.
Данная тенденция к снижению активности могла быть следствием того, что на фоне галоперидола развивался дофаминергический дефицит, косвенно влияющий на локомоцию и вызвавший гипокинезию. Нейротрансмиттер дофамин играет важную роль в модуляции моторных функций [13], и дефицит дофамина или блокада дофаминовых рецепторов приводят к развитию ряда патологических форм поведения, в том числе и касающихся локомоторной активности [14].
Путем дисперсионного анализа для повторных измерений обнаружена достоверная разница между группами, усиленная фактором времени (p < 0.001), а с помощью попарного post hoc анализа на основе критерия Тьюки было установлено, что динамика активности исследованных групп статистически значимо отличается в периоды: 90–120, 120–150 и 150–180 с (p < 0.001). Иными словами, животные, получавшие галоперидол, гораздо раньше переставали передвигаться по ОП, чем контрольные особи.
Ранее было выявлено, что галоперидол, как и многие другие типичные антипсихотики, обладает значительным побочным влиянием на двигательную активность и вызывает выраженные экстрапирамидные расстройства, что клинически проявляется тремором, брадикинезией, ригидностью мышц и др. [15]. Данное обстоятельство не могло не сказаться на характере активности крыс. Частота и длительность случаев замирания в группах контроля и галоперидола статистически значимо различались (рис. 2a и 2b). В целом, движения на фоне действия галоперидола были несколько замедлены и сопровождались, как было сказано выше, поворотами головы (рис. 2c). Эпизоды периодического ускорения и замедления нередко наблюдались также и в группе контроля, поскольку являются элементом нормальной локомоции у крыс [16–18]. Однако важно отметить, что чередования ускорения и замедления отмечалось в группе, получавшей галоперидол, значительно реже, чем в контроле, что указывает на появление выраженной стереотипности и монотонности локомоции под влиянием препарата (рис. 2d). Однообразие поведения, и, вероятно, связанное с ним снижение поисковой деятельности, проявлялись более редким совершением стоек на задних лапах с опорой (рис. 2e) (4.5 ± 1.0 и 1.3 ± 0.4 для групп контроля и галоперидола соответственно, p < 0.01), а также снижением числа обнюхиваний отверстий (лунок) в арене (6.9 ± 1.9 в группе контроля и 2.0 ± 0.8 в группе галоперидола, p < 0.05).
Рис. 2.
Частота некоторых поведенческих паттернов, наблюдаемых у крыс в тесте “Открытое поле” в группах контроля и галоперидола. (a) – Среднее число эпизодов замирания на месте. (b) – Суммарная длительность эпизодов замирания. (c) – Число эпизодов замедления в движении с одновременными поворотами головы или “кивками” ею. (d) – Число случаев чередования ускорения и замедления. (e) – Среднее число стоек на задних лапах, совершенных без опоры на стенку арены. (f) – Случаи шаткой, неуверенной или “хромающей” походки. ***p < 0.001, **p < 0.01, *p < 0.05.
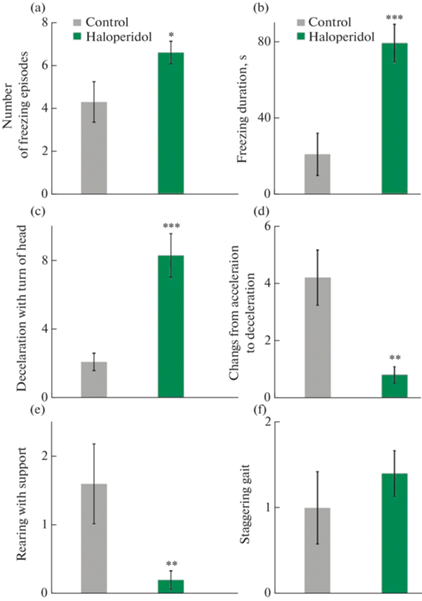
В то же время для остальных общепринятых поведенческих паттернов, таких как стойка на задних лапах без опоры, принюхивание к воздуху как одном из проявлений ориентировочного рефлекса, а также груминг, различия не были выявлены.
Применение метода временного кодирования, позволяющего совместить в одном кадре три последовательных состояния: в настоящем, близком (через 0.33 с) и более отдаленном (через 1.66 с) будущем, позволило наиболее объективно (насколько это возможно в принципе) зафиксировать то или иное поведенческое событие. Примеры поведенческих паттернов, выявленных с помощью цвето-временного кодирования, показаны на рис. 3.
Рис. 3.
Типичные примеры некоторых поведенческих паттернов, отображенных при помощи цвето-временного кодирования в настоящем, близком (через 0.33 с) (серым и желтым) и более отдаленном (через 1.66 с) (красным) будущем. (a) – Принюхивание к воздуху над ареной. (b) – Обнюхивание стенок арены. (с) – Груминг. (d) – Исследование отверстия в поверхности арены. (е) – Стойка без опоры на стенку арены. (f) – Стойка с опорой на стенку арены. Стрелками отмечены области, принципиальные для идентификации того или иного паттерна.
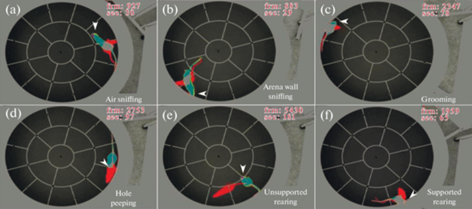
Так, на рис. 3a показан типичный пример кратковременного принюхивания к воздуху (желтым, стрелка), которое быстро сменяется иным поведением (красным). Аналогичным образом выглядит обнюхивание стенок арены (3b, желтым, стрелка) с той разницей, что оно продолжает происходить и в отдаленном будущем, со смещением (красным). Типичный пример груминга отражен на панели (c), а исследование отверстия — на панели (d). Панели (e) и (f) демонстрируют кратковременные (только серым, стрелки) стойки без опоры и с опорой на стенку, в обоих случаях сменившиеся в будущем иными типами поведение (красным).
Вышеперечисленные показатели поведенческой активности, как правило, говорят о степени ориентировочно-исследовательской активности, тревожности животных и об эмоциональной составляющей как таковой. Так, исследование, проведенное Звездочкиной с соавт. [14], выявило выраженное снижение ориентировочно-исследовательского поведения у крыс в тесте ОП после введения галоперидола, а также снижение частоты паттерна груминга, что свидетельствовало о повышении тревожности животных и развитии тормозных процессов в ЦНС.
Однако такой показатель, как груминг, нельзя определить в качестве однозначного маркера тревоги, поскольку он также может означать комфортное и спокойное состояние, в котором пребывает животное. Исходя из этого, изменчивость такого параметра не стоит рассматривать в качестве маркерного показателя эффекта галоперидола или какого-либо другого психоактивного соединения. Нельзя также отрицать, насколько большую роль играет эмоциональная составляющая в двигательных функциях, поскольку было выяснено, что именно мезокортиколимбический путь модулирует реакции, которые связаны со страхом и тревожностью [19].
К сожалению, учет таких параметров поведения затруднен и сопряжен с риском чрезмерной субъективности оценок. Гораздо более эффективным и обоснованным представляется анализ простых двигательных паттернов, интерпретация которых однозначна. К ним относятся: движение вперед, разворот на 180°, повороты влево и вправо, а также повороты головы. На рис. 4a–f отражено попарное сравнение этих двигательных паттернов у двух групп крыс. Статистический анализ показал, что различия значимы для всех паттернов, кроме поворотов головы.
Рис. 4.
Среднее число простых локомоторных событий, зарегистрированных в группах контроля и галоперидола. (a) – Движение вперед. (b) – Разворот на 180°. (с) – Поворот корпуса влево. (d) – Поворот корпуса вправо. (e) – Поворот головы влево. (f) – Поворот головы вправо. ***p < 0.001, **p < 0.01, *p < 0.05.
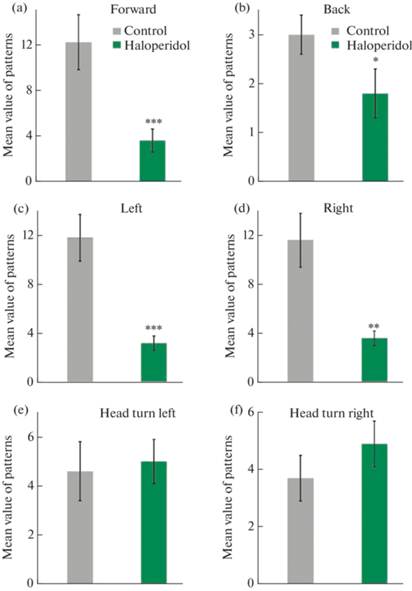
Среднее количество движений вперед в контрольной группе составило 12.2 ± 2.4 против 3.6 ± 1.0 в группе галоперидола (p < 0.01) (рис. 3a), разворотов на 180° – 3.3 ± 0.4 против 1.8 ± 0.5 в группе галоперидола (p < 0.05) (рис. 3b), поворотов вправо или влево – 11.8 ± 1.9 и 11.6 ± 2.2 против 3.2 ± 0.6 и 3.6 ± 0.6 соответственно в группе галоперидола (p < 0.001 и р < 0.01 соответственно) (рис. 3c, d). В совокупности эти данные явно указывают на сниженную базовую локомоторную активность, включающую в себя движение вперед и повороты, в то время как другие моторные акты (повороты головы), ориентировочные рефлексы и общий уровень тревожности остаются примерно одинаковыми.
Однако остается открытым вопрос о том, в какой мере текущее положение животного или его легкое смещение в течение 0.3 с, может служить предиктором дальнейшего движения. Иными словами, есть ли связь между вектором кратковременного смещения и вектором более отдаленного движения? И в том случае, если такая зависимость существует среди интактных животных, то в какой мере она сохраняется или нарушается под влиянием галоперидола?
Так, благодаря цветовому кодированию, соответствующему трем вышеуказанным временным точкам, нами была сделана попытка предсказать будущее положение животных по определенным закономерностям. Для этого нами были выбраны 4 паттерна в отсроченном будущем: движение вперед, разворот на 180°; повороты вправо или влево. Согласно этому набору, собранные данные были разделены на четыре соответствующие группы.
Далее нами было выдвинуто предположение, что поворот головы, хвоста или корпуса животного в ту или иную сторону служит признаком, по которому можно определять вероятность возникновения тех или иных паттернов в будущем, отстоящем на 1.66 с. Также обратило на себя внимание то, что животные зачастую поворачивают хвост и/или голову в направлении будущего движения. Поэтому для анализа было выбрано 3 этих фактора в настоящем и ближайшем будущем. Каждый из факторов соотносился с будущим поведением следующим образом: обращены ли голова, хвост или корпус животного в ту же сторону, куда оно направится через 1.66 с, а именно: вперед, назад, вправо или влево.
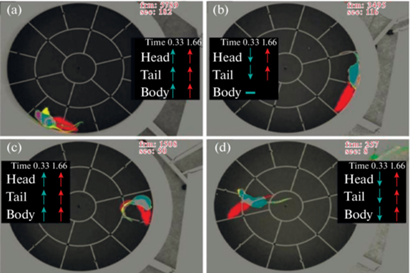
На рис. 5 показаны примеры того, каким образом производилась сортировка в соответствии с указанными факторами. Панель (a) рисунка демонстрирует сдвиг (желтым) головы, хвоста и всего корпуса, примерно сонаправленный будущему положению (красным). На панели (b) показано очень небольшое противонаправленное смещение головы и хвоста (желтым) относительно будущего (красным). Панель (c) демонстрирует выраженную сонаправленность, тогда как панель (d) – слабую противонаправленность.
Рис. 5.
Примеры сонаправленного и противонаправленного смещения головы, хвоста и корпуса крыс в настоящем и через 0.33 с (серым и желтым) относительно положения через 1.66 с (красным). (a) – Пример разворота на 180°. (b) – Пример движения вперед. (с, d) – Примеры поворотов вправо и влево.
Голова, хвост и корпус животных чаще всего принимали независимое положение, поэтому для дальнейшего анализа рассматривались отдельно. При анализе полученных данных относительно положения головы, хвоста и туловища в качестве предсказательных критериев будущего движения были выявлены разнообразные тенденции, которые существенно варьировали от одного животного к другому, а также между основными видами локомоции: движения вперед, назад, поворотов влево и вправо. Однако зависели ли эти тенденции от действия галоперидола? Для ответа на этот вопрос было необходимо вычислить соотношение сонаправленности и противонаправленности у каждого животного в каждом из двигательных паттернов. Так как отсутствовали какие-либо убедительные основания для рассмотрения левых и правых поворотов по отдельности, они были объединены в общий паттерн. При анализе прямолинейного движения примерно в равном числе случаев голова, хвост и корпус занимали сонаправленное и противонаправленное положения. Однако в иных случаях эти части тела чаще занимали положение, противонаправленное движению, чем сонаправленное ему. В связи с этим была применена пропорция противонаправленности к сонаправленности, а не наоборот, хотя принципиального значения это не имело. Таким образом, вычислялся своеобразный “индекс направленности”, который представлял собой пропорцию суммарного числа противонаправленных положений головы, хвоста и корпуса к сонаправленным для каждого из трех двигательных паттернов: движение вперед, назад, поворот. Все пропорции вычислялись индивидуально для каждого животного, после чего были сведены в таблицу.
Алгоритм расчета “индекса направленности”:
1. Отбирались паттерны движения вперед, разворота, поворотов вправо или влево.
2. Для каждого животного в каждом паттерне учитывались положение головы, туловища и хвоста как в настоящем и ближайшем будущем (0.33 с) времени, так и в дальнем будущем (т.е. через 1.66 с). Если положение какой-то из частей тела в настоящем и ближайшем будущем времени совпадало с положением той же части тела в дальнем будущем, то это движение называлось сонаправленным, в противном случае – противонаправленным. Например, на рис. 5b видно, как голова, туловище и хвост развернуты вправо, значит, в паттерн поворота для данного животного мы добавляем по одному сонаправленному движению для каждой части тела.
3. Далее для каждого животного в каждом из паттернов для каждой части тела суммировалось количество сонаправленных и противонаправленных движений, затем рассчитывалось отношение противонаправленных движений к сонаправленным.
В таком виде данные приняли удобный вид для проведения двухфакторного сравнения. Первый фактор отражал индексы направленности для головы, хвоста и туловища, распределенные между строками с 1-й по 10-ю (для головы), с 11-й по 20-ю (для хвоста) и с 21-й по 30-ю (для корпуса). Второй фактор соответствовал группам (контроль или галоперидол), расположенным в двух колонках. Результаты двухфакторного дисперсионного анализа, проведенного в среде MATLAB R2020a, представлены в табл. 1–3.
Таблица 1.
Двухфакторный дисперсионный анализ движения вперед (контроль/галоперидол)
| Движение вперед | Сумма квадратов (SS) | Степени свободы (df) | Дисперсия (MS) | F-критерий | Вероятность Prob > F |
|---|---|---|---|---|---|
| Столбцы (группа животных) | 0.0001 | 1 | 0.00009 | 0 | 0.99 |
| Строки (часть тела) | 27.85 | 9 | 3.1 | 2.97 | 0.01 |
| Взаимодействие | 7.8 | 9 | 0.9 | 0.8 | 0.6 |
| Ошибка | 41.7 | 40 | 1.04 | ||
| Итого | 77.4 | 59 |
Таблица 2.
Двухфакторный дисперсионный анализ движения назад (контроль/галоперидол)
| Движение назад | Сумма квадратов (SS) | Степени свободы (df) | Дисперсия (MS) | F-критерий | Вероятность Prob > F |
|---|---|---|---|---|---|
| Столбцы (группа животных) | 10.5 | 1 | 10.5 | 10.7 | 0.002 |
| Строки (часть тела) | 16.1 | 9 | 1.8 | 1.9 | 0.1 |
| Взаимодействие | 15.4 | 9 | 1.7 | 1.7 | 0.1 |
| Ошибка | 39.5 | 40 | 0.99 | ||
| Итого | 81.6 | 59 |
Таблица 3.
Двухфакторный дисперсионный анализ поворотов влево или вправо (контроль/галоперидол)
| Повороты | Сумма квадратов (SS) | Степени свободы (df) | Дисперсия (MS) | F-критерий | Вероятность Prob > F |
|---|---|---|---|---|---|
| Столбцы (группа животных) | 47.85 | 1 | 47.85 | 30.05 | 0.0000005 |
| Строки (часть тела) | 126.7 | 19 | 6.7 | 4.19 | 0.000003 |
| Взаимодействие | 100.3 | 19 | 5.3 | 3.32 | 0.0001 |
| Ошибка | 127.4 | 80 | 1.6 | ||
| Итого | 402.2 | 119 |
Из табл. 1 следует, что при движении вперед индекс направленности достоверно зависел от частей тела животных, которые вели себя по-разному. В то же время введение галоперидола никак не влияло на пропорцию противонаправленности к сонаправленности (рис. 6) – соответственно, не было и взаимодействия между факторами.
Рис. 6.
Индекс направленности головы, хвоста и корпуса крыс в тесте “Открытое поле” перед одним из простых локомоторных паттернов: движения вперед, назад (с разворотом на 180°) и поворотами влево или вправо. Индекс направленности рассчитывался индивидуально для каждого животного и для каждого из локомоторных паттернов путем деления числа противонаправленных локаций головы, хвоста или корпуса на число сонаправленных положений. Различия между группами контроля и галоперидола статистически достоверны для разворотов назад и поворотов влево или вправо, но не для движения вперед (см. табл. 1– 3), **p < 0.01, ***p < 0.001.
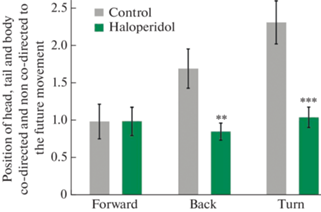
Из табл. 2 следует, что фактор группы животных (контроль или галоперидол) статистически значимо (p < 0.01) влияет на индекс направленности. В то же время фактора влияния положения головы, хвоста или корпуса, а также их взаимодействия с группой животных не отмечалось. Следовательно, гипотеза о влиянии галоперидола на планирование паттерна будущей локомоции подтверждается – после введения галоперидола противонаправленность относительно будущего движения перестает доминировать над сонаправленностью, как это наблюдается в контроле (рис. 6).
Наконец, табл. 3, в которой объединены сведения о поворотах влево и вправо, демонстрирует очень высокий уровень статистической значимости того, что индекс направленности зависит от принадлежности животных к одной из групп. Тем не менее, эффект связан и с положением конкретной части тела животного: головы, хвоста или корпуса. Более того, между указанными факторами наблюдается статистически значимое взаимодействие и, таким образом, эффект галоперидола в наибольшей степени отражается на положении головы и корпуса и в меньшей степени – на положении хвоста. Но в любом случае, как и при движении назад, введение галоперидола резко снижает частоту противонаправленного положения частей тела перед поворотами (рис. 6, справа).
Таким образом, можно утверждать, что галоперидол в гораздо большей степени, по сравнению с контролем, ассоциируется с сонаправленной ориентацией тела при поворотах в стороны и разворотах назад, тогда как при движении прямо он не оказывает никакого воздействия (рис. 6). Иными словами, в определенных случаях животное становится более предсказуемым, что может быть связано с брадикинезией, вызванной галоперидолом.
Установлено, что антагонист дофаминовых D2-рецепторов галоперидол при внутрибрюшинном введении крысам-самцам вызывает двигательный дефицит в тесте “Открытое поле”, особенно во второй половине периода тестирования (90–180 с). Обнаружено снижение двигательной активности вплоть до ее полного отсутствия как в центре, так и на периферии арены. Выявлены случаи неустойчивости позы и замедленности движений под воздействием галоперидола, а также периодические эпизоды довольно длительного замирания. Все вышеперечисленные наблюдения предварительно указывают на вероятность проявления двигательных нарушений, сходных с таковыми при болезни Паркинсона, что косвенно указывает на блокаду дофаминовых рецепторов в нигростриарном пути под влиянием галоперидола [20].
Примененный нами метод цветового кодирования с использованием трех временных точек показал эффективность в анализе и обработке данных. Полученные результаты соответствуют литературным данным, в которых применяли как традиционные, так и компьютеризированные методы визуализации. Неоспоримым преимуществом метода является то, что метод открывает новые возможности в работе с биологическими изображениями при использовании относительно несложной методики, расширяя аналитический арсенал исследователя и снижая степень человеческого фактора за счет объективизации процесса. Он позволяет регистрировать достаточно мелкие движения и визуализировать динамику объектов, совмещенных в одном кадре. Мы считаем, что данный метод в комплексе с другими поможет точнее и быстрее производить оценку эффектов психоактивных соединений при проведении их фармакологического скрининга.
Список литературы
Harris L, Wayland P, Umrania Y, Krishnamurthy D, Rahmoune H, Bahn S (2013) Schizophrenia: metabolic aspects of aetiology, diagnosis and future treatment strategies. Psychoneuroendocrinology 38(6): 752–766. https://doi.org/10.1016/j.psyneuen.2012.09.009
Калитин КЮ, Спасов АА, Муха ОЮ, Придворов ГВ, Липатов ВА (2021) Фармакологические мишени и механизм действия антипсихотических средств в рамках нейрохимической теории патогенеза шизофрении. Рос физиол журн им ИМ Сеченова 107(8): 927–954. [Kalitin KY, Spasov AA, Mukha OU, Pridvorov GV, Lipatov VA (2021) Pharmacological Targets and the Mechanism of Action of Antipsychotic Agents in the Framework of the Neurochemical Theory of the Pathogenesis of Schizophrenia. Russ J Physiol 107(8): 927–954. (In Russ)]. https://doi.org/10.31857/S0869813921080070
Vaiman EE, Shnayder NA, Khasanova AK, Strelnik AI, Gayduk AJ, Al-Zamil M, Sapronova MR, Zhukova NG, Smirnova DA, Nasyrova RF (2022) Pathophysiological Mechanisms of Antipsychotic-Induced Parkinsonism. Biomedicines 10(8): 2010. https://doi.org/10.1007/s40120-018-0105-0
Дроздов АЛ, Демченко ЕМ, Эйяд А, Неруш ОП (2011) Влияние психотропных лекарственных средств на спонтанную поведенческую активность белых крыс. Вiсник Днiпропетр унiвер. Бiологiя. Медицина 2(1): 47–53. [Drozdov AL, Demchenko EM, Ejjyad A, Nerush OP (2011) Influence of psychotropic drugs on spontaneous behavioral activity of albino rats. Visn Dnipropetr Univ Ser Biol Med 2(1): 47–53. (In Russ)]. https://doi.org/10.15421/021108
Białoń M, Wąsik A (2022) Advantages and Limitations of Animal Schizophrenia Models. Int J Mol Sci 23(11): 5968. https://doi.org/10.3390/ijms23115968
Tecott LH, Nestler EJ (2004) Neurobehavioral assessment in the information age. Nat Neurosci 7(5): 462–466. https://doi.org/10.1038/nn1225
Дробижев МЮ, Овчинников АА (2012) Зачем нужна современная классификация антипсихотиков? Рос мед журн 20(29): 1449–1457. [Drobizhev MYu, Ovchinnikov AA (2012) Why a contemporary classification of antipsychotics is needed? Rus Med J 20(29): 1449–1457. (In Russ)].
Irving CB, Adams CE, Lawrie S (2013) Haloperidol versus placebo for schizophrenia. Cochrane Database Syst Rev (11): CD003082. https://doi.org/10.1002/14651858.CD003082.pub3
Prut L, Belzung C (2003) The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: a review. Eur J Pharmacol 463(1–3): 3–33. https://doi.org/10.1016/S0014-2999(03)01272-X
Sysoev YI, Shits DD, Puchik MM, Prikhodko VA, Idiyatullin RD, Kotelnikova AA, Okovityi SV (2022) Use of Naïve Bayes Classifier to Assess the Effects of Antipsychotic Agents on Brain Electrical Activity Parameters in Rats. J Evol Biochem Phys 58: 1130–1141. https://doi.org/10.1134/S0022093022040160
Carola V, D’Olimpio F, Brunamonti E, Mangia F, Renzi P (2002) Evaluation of the elevated plus-maze and open-field tests for the assessment of anxiety-related behaviour in inbred mice. Behav Brain Res 134(1–2): 49–57. https://doi.org/10.1016/s0166-4328(01)00452-1
Wahlsten D (2011) Mouse behavioral testing. How to use mice in behavioral neuroscience. Acad Press. Cambridge (MA).
Zhai S, Shen W, Graves SM, Surmeier DJ (2019) Dopaminergic modulation of striatal function and Parkinson’s disease. J Neural Transm (Vienna) 126(4): 411–422. https://doi.org/10.1007/s00702-019-01997-y
Звездочкина НВ, Муранова ЛН, Андрианов ВВ, Архипова СС, Гайнутдинов ХЛ, Голубев АИ, Плещинский ИН (2004) Локомоторные реакции и возбудимость нейронов в условиях блокады дофамина галоперидолом у беспозвоночных и позвоночных животных. Рос физиол журн им ИМ Сеченова 90(11): 1381–1392. [Zvezdochkina NV, Muranova LN, Andrianov VV, Arkhipova SS, Gaynutdinov KhL, Golubev AI, Pleshchinskiy IN (2004) Locomotor reactions and excitability of neurons during the blockade of dopamine by haloperidol in vertebrate and invertebrate animals. Russ J Physiol 90(11): 1381–1392. (In Russ)].
Strange PG (2008) Antipsychotic drug action: antagonism, inverse agonism or partial agonism. Trends Pharmacol Sci 29(6): 314–321. https://doi.org/10.1016/j.tips.2008.03.009
Alves JA, Boerner BC, Laplagne DA (2016) Flexible Coupling of Respiration and Vocalizations with Locomotion and Head Movements in the Freely Behaving Rat. Neural Plast 2016:4065073. https://doi.org/10.1155/2016/4065073
Bouët V, Borel L, Harlay F, Gahéry Y, Lacour M (2004) Kinematics of treadmill locomotion in rats conceived, born, and reared in a hypergravity field (2 g). Adaptation to 1 g. Behav Brain Res 150(1–2): 207–216. https://doi.org/10.1016/S0166-4328(03)00258-4
Шишелова АЮ, Алиев РР, Раевский ВВ (2015) Ранний сенсорный опыт определяет разнообразие исследовательского поведения в зрелом возрасте. Экспер психол 8(1): 73–84. [Shishelova AYu, Aliev RR, Raevsky VV (2015) Early sensory experience determines variety of exploratory behavior in adult age. Exp Psychol (Russia) 8(1): 73–84. (In Russ)].
de Souza Caetano KA, de Oliveira AR, Brandão ML (2013) Dopamine D2 receptors modulate the expression of contextual conditioned fear: role of the ventral tegmental area and the basolateral amygdala. Behav Pharmacol 24(4): 264–274. https://doi.org/10.1097/FBP.0b013e32836356c4
Amtage J, Schmidt WJ (2003) Context-dependent catalepsy intensification is due to classical conditioning and sensitization. Behav Pharmacol 14(7): 563–567. https://doi.org/10.1097/00008877-200311000-00009
Дополнительные материалы отсутствуют.
Инструменты
Российский физиологический журнал им. И.М. Сеченова