

Российский физиологический журнал им. И.М. Сеченова, 2023, T. 109, № 3, стр. 348-365
Нейробиологические эффекты комбинированного действия антиортостатического вывешивания, квазихронического гамма-излучения и ионов углерода у крыс
А. А. Перевезенцев 1, К. Б. Лебедева-Георгиевская 1, О. С. Кузнецова 1, В. С. Кудрин 1, 2, В. А. Пикалов 3, А. С. Штемберг 1, *
1 Институт медико-биологических проблем РАН
Москва, Россия
2 Научно-исследовательский институт фармакологии им. В.В. Закусова РАМН
Москва, Россия
3 Институт физики высоких энергий им. А.А. Логунова Национального
исследовательского центра “Курчатовский институт”
Протвино, Россия
* E-mail: andrei_shtemberg@mail.ru
Поступила в редакцию 29.06.2022
После доработки 06.02.2023
Принята к публикации 07.02.2023
- EDN: FSSYAL
- DOI: 10.31857/S086981392303007X
Аннотация
Исследовали нейробиологические эффекты физических факторов, моделирующих в наземном эксперименте условия межпланетного космического полета: гипогравитации (модель антиортостатического вывешивания) и ионизирующих излучений (квазихроническое гамма-облучение и облучение головы животных ионами углерода 12С) с учетом типологических характеристик экспериментальных животных (самцов крыс линии Long Evans). Были изучены изменения в поведении животных, ЭЭГ и нейрохимических процессах в структурах мозга, ответственных за когнитивные и эмоциональные процессы (фронтальная кора, таламус, гипоталамус). Показаны различия в двигательной и ориентировочно-исследовательской активности, а также когнитивных функциях животных с преобладанием возбуждения или торможения под действием изученных факторов, в основе которых лежат различия в балансе основных тормозного и возбуждающего нейромедиаторов (ГАМК и глутамата). В частности, установлено, что крысы с преобладанием возбуждения быстрее обучаются, но хуже сохраняют навыки по сравнению с крысами преимущественно тормозного типа. Обнаружены изменения соотношения основных ритмов ЭЭГ в результате экспериментальных воздействий.
Одной из основных проблем при подготовке межпланетных миссий является радиационный барьер – опасность воздействия галактических космических лучей, в состав которых входят тяжелые ионы высоких энергий, вплоть до 1020 МэВ. При этом на первый план выходят возможные нарушения в центральной нервной системе (ЦНС) – так называемый эргономический риск, обусловленный возможными нарушениями операторской деятельности космонавтов и связанный с угрозой их жизни непосредственно в ходе полета. Существенные нарушения функций ЦНС при воздействии тяжелых ионов показаны в целом ряде работ [1–4]. Весьма важной и в то же время наименее изученной проблемой являются нейробиологические эффекты комбинированного действия ионизирующих излучений и нерадиационных факторов космического полета, важнейшим из которых является микрогравитация. В наших предыдущих работах были изучены основные нейробиологические эффекты комбинированного действия моделируемой микрогравитации и различных видов ионизирующих излучений, входящих в состав галактических космических лучей, на разных уровнях организации ЦНС [5–8]. В частности, было показано, что при изучаемых воздействиях страдает в первую очередь эмоционально-мотивационная сфера, в основе чего лежит угнетение дофаминергической системы мозга; имеет место зависимость степени нарушений от сроков воздействия. Методологической основой этих работ было моделирование комбинированного действия длительного синхронного воздействия гамма-облучения и невесомости с помощью антиортостатического вывешивания (АнОВ) с последующим облучением головы животных протонами высоких энергий и/или ионами углерода 12С. Кроме того, как показано в ряде работ [6, 9, 11], важную роль в модификации нейробиологических эффектов воздействия факторов космического полета играют индивидуальные и типологические особенности высшей нервной деятельности (ВНД). В нашем эксперименте с обезьянами было убедительно показано, что типологические характеристики ВНД могут играть определяющую роль в характере нарушений когнитивных функций, что диктует необходимость учета этих характеристик при отборе космонавтов для межпланетных полетов [7]. Исходя из этого, задачей наших исследований было изучение отдаленных эффектов комбинированного действия АнОВ и ионизирующих излучений на интегративном и нейрохимическом уровнях, а также роли типологических особенностей ВНД в модификации этих эффектов.
Настоящая работа является продолжением этих исследований. В ней мы применили несколько иные характеристики длительного гамма-облучения животных, приблизив его к хроническому.
МЕТОДЫ ИССЛЕДОВАНИЯ
Экспериментальные животные
Работа проводилась на самцах крыс линии Long Evans из собственного вивария Института медико-биологических проблем, на дату начала эксперимента возраст животных составлял 1.5–2 мес., а масса тела 150–180 г. Всего в эксперименте было использовано 22 животных: по 6 животных в двух экспериментальных группах и по 5 животных в двух контрольных группах. Содержание животных осуществлялось в стандартных домашних клетках (ГОСТ 33216-2014) размерами 40 × 60 × 20 см (одна группа – одна клетка) при температуре 23°С на протяжении всего эксперимента, световой день составлял 12 ч (искусственное освещение) с 9:00 до 21:00, крысы имели свободный доступ к воде и корму (синтетический полнорационный комбикорм для грызунов ПК 120-1_45, ГОСТ 33215-2014, МУ 1.2.2520-09), подстил по ГОСТ 33215-2014, п. 6.8, аналогично предыдущим исследованиям [5, 6]. Выбор линии крыс обусловлен применением сложных поведенческих методик, в частности, Y-лабиринта, в котором линейные крысы Wistar дают низкие результаты. Перед началом эксперимента все животные проходили трехдневное приучение к рукам (хэндлинг).
Экспериментальные воздействия
Факторы дальнего космического полета моделировались в две фазы: 7-суточное АнОВ синхронно с гамма-облучением, затем, на следующий день, отдельно воздействие тяжелых ионов.
При создании АнОВ использовали индивидуальные металлические секции размером 42 × 42 × 40 см, скомпонованные в трехъярусный стеллаж (15 секций). Животных вывешивали за основание хвоста под углом 30°–40° с расчетом снятия статической нагрузки с задних конечностей. При этом крыс крепили с помощью специальных карабинов, надевающихся на металлический стержень так, чтобы они могли свободно перемещаться в пределах клетки. Таким образом создавался антиортостаз, вызывающий перераспределение жидкостей в организме, и снималась статическая нагрузка с задних конечностей, что воспроизводит основные эффекты микрогравитации. Данная методика – принятая экспериментальная наземная модель невесомости для мелких лабораторных животных, описанная более подробно в [15, 16]. Общий вид отдельного отсека с вывешенной крысой представлен на рис. 1.

Стеллажи с животными были размещены в помещении для облучения: стеллаж с крысами, подвергавшимися воздействию АнОВ и γ-излучения – в зоне облучения на расстоянии 4 м от источника излучения, с таким расчетом, чтобы облучение всех крыс было равномерным; крысы контрольных групп размещались в том же помещении в домашних клетках, но с тыльной стороны источника.
Для облучения использовали установку ГОБО-60 с источником 137Cs (72 г-экв. Ra). Мощность дозы составила 0.7 сГр/ч. В данном эксперименте было проведено квазихроническое облучение в течение всех 7 дней вывешивания, что приближает условия проведения эксперимента к реальным условиям космического полета. Доза гамма-облучения составила 1.2 Гр. После окончания гамма-облучения и снятия крыс с вывешивания было проведено однократное облучение головы животных ионами углерода 12С с энергией 455 МэВ/н в дозе 0.8 Гр на ускорителе У-70 на базе Института физики высоких энергий (Протвино).
Выбор режима облучения основывался на следующем: суммарная доза 2.0 Гр для крыс (исходя из их видовой радиочувствительности) примерно соответствует расчетной дозе, которая может быть получена космонавтами в межпланетном полете Земля–Марс–Земля. Снижение суммарной дозы относительно более ранних работ [5] было обусловлено большей эффективностью квазихронического облучения по сравнению с фракционированным. Общая схема эксперимента дана на рис. 2.

Методы исследования поведения животных
В целом эксперимент продолжался на протяжении 8 мес. До воздействия модельных факторов животные были типологизированы по особенностям ВНД и разбиты на группы. На следующий день после завершения воздействий начато обучение в Y-лабиринте, продолжавшееся 5 дней. Через месяц после воздействия начат основной этап поведенческого тестирования, в который вошли тесты “Открытое поле” (ОП), “Приподнятый крестообразный лабиринт” (ПКЛ), “Водный лабиринт Морриса” (ВЛМ). Наряду с этим ежемесячно проверялся выработанный в Y-лабиринте сложный рефлекс активного избегания. Спустя 6 месяцев после воздействий были сняты электроэнцефалограммы, далее крысы были умерщвлены для взятия проб на нейрохимические исследования.
Типологизация животных
Типологизация животных проводилась до начала основного эксперимента. Исходная выборка из 100 животных была протестирована в течение 2 недель (партиями, по 2 последовательных дня на каждое животное) с целью отбора однородных по типу ВНД животных – возбудимых (группы А и К) и тормозных (группы Е и С) соответственно. Для типологизации животных использовали модифицированную методику избегания закрытого темного пространства при крике раздражаемой особи (методика “эмоционального резонанса”) по Симонову [12]. Показана успешность применения этой методики для определения типологических характеристик животных [13, 14].
Тестирование проводилось в течение 2 дней по 5 мин (300 с) на каждое животное. Изначально крысу высаживали в дальний от входа в темный отсек угол светлого отсека. Регистрировали количество входов в темный отсек и общее время, проведенное в темном отсеке. Эти параметры имеют обратную зависимость. Животные, показавшие максимальную активность, считаются возбудимыми, минимальную – тормозными. При этом контрольные группы (К и С) подбирались таким образом, чтобы средние показатели в тесте совпадали с опытными (А и Е соответственно). Те животные из исходной выборки в 100 крыс, которые показали промежуточные результаты, выбраковывались.
Тестирование в “открытом поле”
Использовалась стандартная пластиковая установка (приказ Минздрава РФ № 281 от 30.04.2013) диаметром 90 см, пол которой размечен квадратами со стороной 10 см. В каждой “клеточке” посредине имеется отверстие диаметром 3 см. Продолжительность эксперимента составляла 5 мин. Животное аккуратно высаживалось в центр. Регистрировали двигательную активность (число пересечений линий) раздельно за первую и вторую половины времени тестирования, ориентировочно-исследовательскую активность (число заглядываний в отверстия, число стоек с опорой и без опоры), эмоциональность (число актов дефекации), а также время выхода в центральную часть поля и число замираний (freezing).
Тестирование в приподнятом крестообразном лабиринте (ПКЛ)
Приподнятый крестообразный лабиринт является общепринятым тестом для оценки тревожности экспериментальных животных. Критерием тревожности служит количество выходов в открытый отсек лабиринта и время пребывания в нем. Использовалась стандартная крестообразная установка размером 1.5 м, два противоположных рукава которой закрыты по бокам черными стенками высотой 50 см. Крыса первоначально высаживалась в темный рукав. Тест проводился на протяжении 2 мин, в ходе которых регистрировались число перебежек между темными рукавами, латентный период выхода в светлый рукав, количество выходов и общее время, проведенное в светлом рукаве.
Выработка дифференцировочного условного рефлекса (УР) активного избегания (УРАИ) в Y-образном лабиринте
Лабиринт представлял собой три рукава (отсека) размером 25 (длина) × 20 (ширина) × 25 (высота) см из белого непрозрачного пластика, соединенных под углом 120 градусов. В каждом отсеке были установлены: управляемый светодиодный светильник, обеспечивающий освещенность 80 лк/см2, управляемый источник звука различной частоты и электродный пол, позволяющий стимулировать крысу импульсами с частотой 4–5 Гц энергией 0.004 Дж. Параметры выработки условного рефлекса были следующими: 6 с – время изолированного действия условного сигнала; 30 с – время сочетания условного сигнала и электроболевого подкрепления; 10 с – пауза между сочетаниями. Регистрировали число реакций избегания (переходов по условному сигналу в правый отсек лабиринта); число реакций избавления (число переходов в правый отсек при сочетании условного сигнала и электроболевого подкрепления); число ошибок (переходов в левый отсек лабиринта); число отказов и число межсигнальных реакций. Проводилось по 15 сеансов обучения в течение 4 дней.
Водный тест Морриса
Водный тест (лабиринт) Морриса предназначен для исследования пространственной памяти и ориентации животных. Подробно методика была описана в предыдущих публикациях. Использовалась стандартная установка диаметром 150 см, расположенная в полузатемненной комнате с освещением по периметру, что исключает наличие ярко выраженных ориентиров. Животное высаживалось каждый раз с одной точки и должно было отыскать скрытую под водой платформу, также расположенную в постоянном месте. Температура воды составляла 20°С, воздуха – 23°C. Испытание записывалось на видео для последующей обработки в программе Bonsai (Open Ephys, США), определяющей временные характеристики прохождения теста, скорость, манеру поведения крысы.
Методы электроэнцефалографических исследований
Для анализа возможных электрофизиологических изменений в работе головного мозга крыс через 6 мес. после облучения, по достижении массы тела 350–400 г, проводили операцию для вживления интракраниальных энцефалографических электродов. Животное закреплялось в стереотаксической установке, операция происходила под общим наркозом (хлоралгидрат 350 мг/кг, концентрация 85 мг/мл). Голова крысы скальпировалась, после чего происходила очистка и сушка черепа. Электроды, представляющие собой позолоченные винты диаметром 1.5 мм и длиной 1.0 мм, закреплялись внутри кости с помощью пластмассы холодной быстрой полимеризации (протакрил). Расположение электродов: в области префронтальной коры – 2, в области моторной коры – 1, в области лимбической коры – 1; референтный и земляной электроды располагались в затылочной кости.
Спустя 7 дней после операции была произведена регистрация электроэнцефалограммы с помощью чипа Neurologger (TSE), запись проходила в свободном поведении в домашней клетке в течение одного часа, все животные записывались в течение одного дня, частота оцифровки 565 Герц.
Всего было прооперировано и записано 15 животных, в связи с техническими особенностями работы чипов Neurologger пригодными для описания оказались 7 записей, 3 контрольных и 4 экспериментальных животных. Анализировались данные, полученные для префронтальной коры как наиболее важной для реализации высших когнитивных функций.
Для оценки достоверности результатов был применен непараметрический критерий Уилкоксона.
Методы нейрохимических исследований
Для проведения нейрохимических исследований контрольных и подвергнутых экспериментальным воздействиям крыс декапитировали через 7 мес. после воздействий и выделяли следующие структуры мозга: префронтальную кору, гипоталамус, таламус. Структуры мозга замораживали в жидком азоте и взвешивали. Выделенные структуры гомогенизировали при 4°С в стеклянном гомогенизаторе с тефлоновым пестиком (0.2 мм) при скорости вращения пестика 3000 об./мин. В качестве среды гомогенизации и выделения использовали 0.1 Н НСlО4 с добавлением внутреннего стандарта ДОБА (3,4-диоксибензиламин) – вещество катехоламиновой природы, но не встречающегося в нативной ткани, в концентрации – 0.5 нмоль/мл. Cтруктуры мозга гомогенизировали в 20 объемах среды выделения. Пробы центрифугировали при 4°С, 10 000 g в течение 15 мин. Супернатант использовался в дальнейшем для определения моноаминов и их метаболитов.
Концентрацию моноаминов и их метаболитов определяли с помощью метода высокоэффективной жидкостной хроматографии (ионпарная хроматография) с электрохимической детекцией на хроматографе LC-304T (BAS, WestLafayette, США) с инжектором Rheodyne 7125, петля для нанесения образцов – 20 мкл. Изучаемые вещества разделяли на обращенно-фазной колонке ReproSil-Pur, ODS-3, 4 × 100 мм, 3 мкм (Dr.Majsch GMBH, “Элсико”, Москва). Насос PM-80 (BAS, США), скорость подвижной фазы 1.0 мл/мин, при давлении 200 атм. Мобильная фаза: 0.1 M цитратно-фосфатный буфер, содержащий 1.1 мM октансульфоновой кислоты, 0.1 мM ЭДТА и 9% ацетонитрила (pН 3.0). Скорость протока составляла 1 мл/мин. Измерение проводили с помощью электрохимического детектора LC-4B (BAS, США) на стеклоугольном электроде (+0.85 V) против электрода сравнения Ag/AgCl. Регистрация образцов проводилась с применением аппаратно-программного комплекса МУЛЬТИХРОМ 1.5 (АМПЕРСЕНД). Все использованные для анализа реактивы были высокой степени чистоты: “ос. ч.”, “х. ч.” или “analytical grade”. Для калибровки хроматографа использовались смеси рабочих стандартов определяемых веществ в концентрации 500 пмоль/мл. Величины концентрации моноаминов в опытных образцах рассчитывали методом “внутреннего стандарта”, исходя из отношений площади пиков в стандартной смеси и в образце. Определяли содержание ГАМК, глутамата, норадреналина (НА), дофамина (ДА) и его метаболитов – 3,4-диоксифенилуксусной кислоты (ДОФУК) и гомованилиновой кислоты (ГВК), 3-метокситирамина (3-МТ), серотонина (5-окситриптамина, 5-ОТ) и его метаболита – 5-гидроксииндолуксусной кислоты (5-ОИУК) в 3 структурах мозга: префронтальная кора, гипоталамус и таламус.
Результаты нейрохимических исследований выражали в виде средних величин ± стандартная ошибка среднего. Полученные результаты анализировали с помощью односторонней ANOVA и posthoc теста Дункана.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Результаты типологизации животных
В первый день среднее время в темном отсеке составило 254 с (стандартное отклонение 36.5 с, медианное время – 266 с) с разбросом от 172 до 291 с при среднем количестве заходов в темный отсек 7.5.
На второй день общая тревожность крыс снизилась и среднее время составило 248 с (стандартное отклонение 45 с, но при этом медианное – 259 с) с разбросом от 134 до 299 с, что демонстрирует расхождения в поведении индивидуумов. Среднее количество входов составило 6.04.
Для третьего и четвертого дней (вторая партия крыс) цифры составили соответственно 266 и 272 с – среднее время, 7 и 4.5 – количество входов. Разброс в третий день был от 233 до 292 с, в четвертый – от 204 до 299 с.
Для дальнейшей работы из общего массива были отобраны четыре группы животных:
– группа А (“Альтруисты” по терминологии Симонова, 6 животных) – наименьшее время в темном отсеке, наибольшее число входов, соответственно: возбудимые, высокоэмоциональные, с повышенной тревожностью;
– группа Е (“Эгоисты”, 6 животных) – наибольшее время в темном отсеке, наименьшее число входов: преимущественно тормозные, низкоэмоциональные, с пониженной тревожностью;
– группа С (контроль – эгоисты, 5 животных) – подобраны таким образом, чтобы примерно соответствовать группе E;
– группа К (контроль – альтруисты, 5 животных) – аналогично группе А.
Измеренные и усредненные параметры по группам (по второму дню, как более показательному) даны в табл. 1; можно видеть, что крысы групп E и C практически не реагируют на крик раздражаемой особи, тогда как крысы групп А и К реагируют достаточно активно, совершая до 14 (в среднем 8.8 и 7.6 соответственно) кратковременных выходов из темного отсека.
Исследование поведенческих реакций крыс
Результаты тестирования животных в “открытом поле” в целом не выявили значимых различий в поведении крыс разных типологических групп. Различия между экспериментальными группами и контролем выражались лишь в виде тенденций, подтверждающих, однако, выводы предыдущих исследований. Они проявились в более высоких показателях двигательной активности крыс с высокой возбудимостью по сравнению с преимущественно тормозными животными и контролем.
По показателям латентного периода выхода в центр “открытого поля” и времени пребывания в нем существенных различий между экспериментальными группами получено не было, однако по времени пребывания в центре показатели крыс обеих экспериментальных групп были существенно ниже контрольных, что может свидетельствовать о повышении тревожности в результате экспериментальных воздействий.
Исследование тревожности животных в приподнятом крестообразном лабиринте
Тестирование крыс всех 4 групп было проведено через месяц после экспериментальных воздействий по стандартной методике приподнятого крестообразного лабиринта продолжительностью 5 мин.
Полученные результаты в целом укладываются в выдвинутую нами гипотезу о различиях в поведении крыс исследуемых групп с противоположными характеристиками ВНД. Экспериментальные крысы с повышенной возбудимостью (группа А) продемонстрировали самое низкое число выходов в открытый отсек ПКЛ по сравнению с животными с преобладанием торможения и контролем (p < 0.05, тест Левена) (рис. 3), что свидетельствует о сохраняющейся повышенной тревожности через месяц после экспериментальных воздействий.
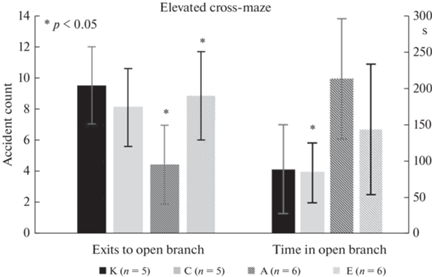
Несколько неожиданными результатами явились более высокие (на грани достоверного, p ~ 0.06) показатели общего времени, проведенного в светлом отсеке, для обеих экспериментальных групп по сравнению с контролем. Предположительно, это может быть связано с угнетением исполнительных механизмов ориентировочной реакции в гипоталамусе и ретикулярной формации в результате облучения [11].
Таким образом, тестирование в ПКЛ в целом выявило тенденции, подтверждающие результаты исследования поведения животных в “открытом поле”. Экспериментальные воздействия приводят к повышению тревожности, различия между крысами разных типологических групп отчетливо выявляются в данном тесте. Как было показано ранее, эти различия, как между экспериментальными и контрольными животными, так и между крысами разных типологических групп, сохраняются в течение длительного времени после экспериментальных воздействий [6, 7].
Исследование когнитивных функций животных в Y-образном лабиринте
Исходный уровень обучения крыс составлял 35% правильных переходов. Целью исследования было закрепление либо утрата приобретенного рефлекса при редко повторяющемся тестировании.
В данном эксперименте тестирование повторялось после первоначального обучения по одному дню с интервалом в месяц, по 15 сеансов для каждого животного.
График, отражающий общую динамику обучения по группам, приведен на рис. 4. Для наглядности из данных исключены животные (по одному в каждой группе), продемонстрировавшие худшую обучаемость.

Можно видеть, что возбудимые крысы группы А за первый этап (пять дней последовательно) теста достигли уровня обучения 37%, превзойдя животных остальных групп.
При последующих тестированиях (один раз в месяц) крысы группы А не только медленнее обучались, но и демонстрировали частичную утрату выработанного УРАИ; аналогичный результат показала группа К. После 3-го месяца редкого (раз в месяц) повторения теста выработанный УРАИ начинает ослабевать у всех групп, особенно хорошо это заметно при измерении времени избегания. Тормозные облученные крысы группы Е, однако, демонстрировали практически линейное улучшение показателей вплоть до уровня обучения 40%, в итоге продемонстрировав наилучшие показатели среди всех животных. Облученные крысы из групп А и Е проходили исходное обучение наравне с контролем, однако последующие тесты выполняли лучше контрольных, что, по-видимому, свидетельствует о большей пластичности нервных процессов в период восстановления после экспериментальных воздействий.
В целом очевидно, что повторение теста с интервалом в месяц плохо укрепляет исходно приобретенный УРАИ, тормозные крысы некоторое время сохраняют уровень обученности, тогда как более возбудимые животные могут утрачивать навык.
Особо следует отметить “шестой день” теста, т.е. повтор через два месяца после воздействий. Данный день характеризовался крайне неблагоприятными метеорологическими условиями: если накануне атмосферное давление составляло 736 мм рт. ст., то к утру следующего дня оно поднялось до 753 мм рт. ст., скорость подъема достигала 1 мм рт. ст. в час (данные метеоархива rp5.ru). По-видимому, такое колебание погоды оказало сильное угнетающее действие на животных, однако следует констатировать, что группа E опять-таки оказалась наиболее устойчивой и к этому воздействию.
Сходные результаты наблюдались в ранее проводившихся тестах УРАИ с “челночной камерой” [6, 9]. По-видимому, такие результаты свидетельствуют во-первых, о более высокой пластичности нервных процессов у крыс группы А в сравнении с группой Е, а во-вторых – о влиянии в долгосрочной перспективе регенерационных процессов в нервной ткани у облученных крыс (в сравнении с необлученными С и К).
Ввиду задействования в методике УРАИ элементов пространственного ориентирования и, как следствие, заметной схожести методик УРАИ и ВЛМ в части задействования гиппокампа, полученные результаты оказались ожидаемо схожи. Следует, однако, напомнить, что испытания по методике УРАИ проводились на протяжении 4 месяцев (с ноября по февраль) и отражают процессы восстановления ЦНС после воздействий, тогда как тест ВЛМ был выполнен фактически единоразово, в феврале, и больше отражает установившееся состояние ЦНС и отдаленные последствия воздействий. Результаты теста Морриса приведены на рис. 5.

Энцефалографические исследования
Обработка записей ЭЭГ производилась в программах Powergraph 3, EDFbrowser, Excel 10 и Statistica 8. Для улучшения качества сигнала были применены фильтры низких частот от 1 до 2 Гц.
Для визуального сравнения всех часовых записей строились спектральные плотности в программе EDFbrowser. Далее анализировались отрезки без артефактов длительностью 20–30 с. По данным отрезкам строились спектры мощности, определялась частота и амплитуда наибольшего пика. Значения амплитуды и частот спектра сохранялись для программы Excel. Для оценки достоверности результатов был применен непараметрический критерий Уилкоксона и критерий Стьюдента.
У всех животных вне зависимости от группы наблюдалась визуально нормальная картина ЭЭГ. При визуальном анализе записей никаких отклонений на ЭЭГ обнаружено не было, абсансная активность отсутствовала. Визуальный анализ спектральных плотностей показал уменьшение доли альфа-ритма относительно всего времени записи.
Было проведено сравнение средних амплитуд и частот по записям, очищенным от артефактов; различий в зависимости от типологических особенностей ВНД животных выявлено не было, поэтому дальнейший анализ проводили, сравнивая показатели контроля и всех крыс, подвергшихся экспериментальным воздействиям.
Исходя из проведенного анализа, наблюдали значимое (p < 0.05, t тест Стьюдента) уменьшение средней амплитуды и средней частоты ритма ЭЭГ у облученных крыс по сравнению с контрольными животными.
Для оценки изменения ритмов были посчитаны процентные соотношения вклада основных ритмов в электрическую активность мозга животных. Данные представлены на рис. 6.
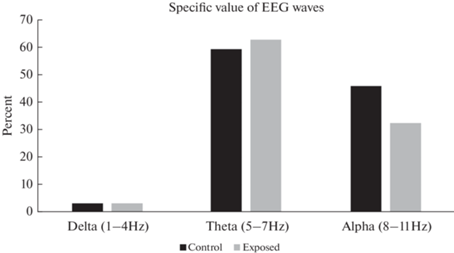
Исходя из проведенного анализа, вклад дельта- и тета-ритмов несущественно увеличился, в то время как вклад альфа-ритма значительно снизился. Таким образом, после экспериментальных воздействий происходило ослабление альфа-ритма, наблюдаемое и при анализе спектральной плотности.
Нейрохимические исследования
В результате исследований в таламусе и фронтальной коре крыс экспериментальных групп были обнаружены существенные (p < 0.05, t-тест Левена) различия в концентрациях ГВК и соотношении ГВК/ДА, представленные на рис. 7 и 8.
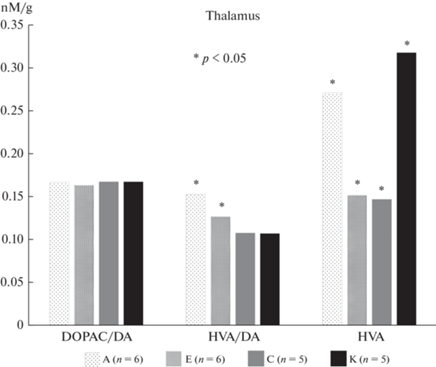
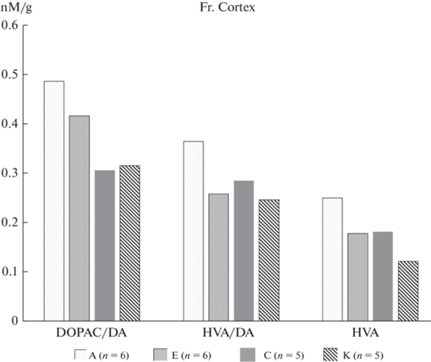
Наиболее заметные различия между типологическими группами обнаружены в таламусе, который осуществляет передачу нервных импульсов коре в ответ на различные воздействия. Концентрация ГВК у крыс с преобладанием возбуждения (группа А) почти в 2 раза превышала таковую у крыс преимущественно тормозного типа (группа Е), что согласуется с результатами поведенческих исследований. Изменения затрагивали преимущественно дофаминергическую систему. Во фронтальной коре сохранялась та же тенденция, хотя и в значительной мере менее выраженная.
На данном этапе исследований мы также оценили соотношение основных тормозных и возбуждающих нейромедиаторов у крыс разных типологических групп после экспериментальных воздействий. Как и следовало ожидать, во фронтальной коре концентрация ГАМК – основного тормозного нейромедиатора оказалась выше у крыс с преобладанием торможения (группа Е) (рис. 9), однако без статистически достоверного различия. Статистически достоверно (p < 0.05, t-тест Левена) концентрация ГАМК ниже у облученных животных по сравнению с экспериментальными. В то же время, в концентрации глутамата – основного возбуждающего нейромедиатора не было выявлено существенных различий между крысами разных типологических групп, однако было зарегистрировано существенное различие по этому показателю между соответствующими контрольными типологическими группами. Можно предположить, что сохраняющийся в течение длительного времени возбуждающий эффект экспериментальных воздействий нивелировал разницу между группами.
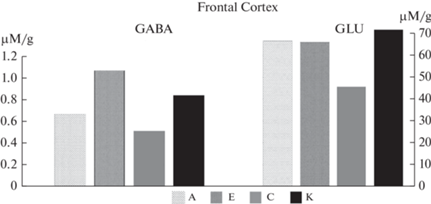
В таламусе и гипоталамусе существенных различий по этим показателям выявлено не было.
Ранее значительные изменения в концентрации моноаминов и их метаболитов были показаны в префронтальной коре при декапитации животных после предпринятых воздействий, в частности, после облучения крыс ионами углерода 12C [13], но не в отдаленный период [14]. Аналогичные изменения происходили у крыс линии WAG/Raj на ранних этапах формирования абсансной эпилепсии, а также в ответ на фармакологическое воздействие – введение медапара, через 1 сутки после облучения протонами высоких энергий как на пролете [15], так и в пике Брэгга [16].
Высокая чувствительность и реактивность как префронтальной, так и фронтальной коры относительно других исследованных структур мозга является фундаментальным свойством этой структуры. Повышенная реактивность этих структур к стрессу показана в ряде работ, в частности, в работе [17]. Медиальная префронтальная кора крыс является одним из важнейших узлов системы быстрого обучения, который извлекает недавние и отдаленные воспоминания [18]. Наблюдали быстрое усиление функционального сопряжения латеральной префронтальной коры, базальных ганглиев (хвостатого ядра – caudate nucleus и неостриатума – putamen) и орбитофронтальной коры, а также между латеральной префронтальной корой и премоторной корой [19]. Обнаружена быстрая реакция дорзо-латеральной верхней префронтальной коры к кодированию новых стимулов в ходе задач, требующих высокого уровня рабочей памяти [20].
Обнаружены также клеточные механизмы, способствующие реализации быстрых реакций нейронов префронтальной коры. Взаимодействие между ингибирующими быстрыми спайковыми интернейронами и возбуждающими пирамидальными нейронами крыс облегчает реализацию фундаментальных свойств корковых сетей. Ключевой функцией быстрых спайковых интернейронов является обеспечение быстрого торможения в локальных сетях сенсорной и моторной коры и обработки входной информации из таламуса в кору. Эти результаты свидетельствуют о наличии различных льготных подсетей или локальных сетей между быстрыми спайковыми интернейронами и пирамидальными клетками префронтальной коры крыс, которые могут быть специфическими для этой области коры [21]. Выявлена быстрая ДА-ергическая модуляция содержания кальция и быстрых потенциалов в дендритах пирамидных нейронов префронтальной коры крыс [18]. В префронтальной коре ДА-ергическая модуляция реализуется менее чем за 0.5 с, в то время как в других структурах эта реакция занимает несколько секунд.
Помимо того, что префронтальная кора формирует и контролирует эмоциональные и мотивационные состояния [22, 23], она играет ключевую роль в когнитивных процессах [24]. В то же время фронтальная кора отвечает за инициацию и контроль произвольных движений. Если у человека когнитивные процессы можно определить как приобретение знаний и опыта и их дальнейшее использование, то у животных это приобретение опыта и его дальнейшее использование. Еще одной из основных функций фронтальной и префронтальной коры при реализации поведения животными является принятие решения и выбор действия [25 ] , также связанное с когнитивной деятельностью.
Таким образом, основной реакцией моноаминергических систем на примененные нами воздействия можно считать изменение активности дофаминергической системы, которая является одной из основных в процессе обучения. В то же время, изменение концентраций основных тормозного и возбуждающего нейромедиаторов вполне согласуется с поведенческими исследованиями и объясняет нейрохимические механизмы различий в поведении между животными разных типологических групп. Животные с изначально более низким метаболизмом дофамина и преобладанием ГАМК (“Эгоисты”) оказываются менее подверженными влиянию модельных факторов полета.
Список литературы
Rabin BM, Joseph JA, Shukitt-Hale B (2004) Heavy particle irradiation, neurochemistry and behavior: threshold, dose-response curves and recovery of function. Advances in Space Res 33: 1330–1333. https://doi.org/10.1016/j.asr.2003.09.051
Rabin BM, Joseph JA, Shukitt-Hale B (2005) A longitudinal study of operant responding in rats irradiated when 2 months old. Radiat Res 164: 552–555. https://doi.org/10.1667/rr3349.1
Britten RA, Davis LK, Johnson AM, Keeny S, Siegel A, Sanford LD, Singletary SJ, Lonart G (2012) Low (2 cGy) doses of 1 GeV/u 56Fe-particle radiation loads to a persistent reduction in the spatial learning ability of rats. Radiat Res 177: 146–151. https://doi.org/10.1667/rr2637.1
Machida M, Lonart G, Britten A (2010) Low (60 cGy) doses of 56Fe HZE-particle radiation lead to a persistent reduction in the glutamatergic readily releasable pool in rathippocampal synaptosomes. Radiat Res 174: 618–623. https://doi.org/10.1667/RR1988.1
Лебедева-Георгиевская КБ, Кохан ВС, Шуртакова АК, Перевезенцев АА, Кудрин ВС, Штемберг АС, Базян АС (2019) Нейробиологические эффект комбинированного воздействия антиортостатического вывешивания и ионизирующих излучений различного качества. Нейрохимия 3: 254–264. [Lebedeva-Georgievskaya KB, Kohan VS, Shurtakova AK, Perevezentsev AA, Kudrin VS, Shtemberg AS, Bazyan AS (2019) Neurobiological effects of combined exposure to anti-orthostatic hanging and ionizing radiation of various quality. Neurochemistry 3: 254–264. (In Russ)]. https://doi.org/10.1134/S1027813319030105
Штемберг АС, Перевезенцев АА, Лебедева-Георгиевская КБ, Митрофанова ОВ, Кудрин ВС, Базян АС (2020) Роль типологических особенностей высшей нервной деятельности в нейробиологических эффектах комбинированного действия антиортостатического вывешивания, γ-излучения, протонов и ионов углерода 12С. Радиац биол Радиоэкол 1: 51–62. [SHtemberg AS, Perevezentsev AA, Lebedeva-Georgievskaya KB, Mitrofanova OV, Kudrin VS, Bazyan AS (2020) The role of typological features of higher nervous activity in the neurobiological effects of the combined action of anti-orthostatic hanging, gamma radiation, protons and carbon ions 12C. Radiac Biol Radioecol 1: 51–62. (In Russ)]. https://doi.org/10.31857/S0869803120010130
Семагин ВН, Зухарь АВ, Куликов МА (1988) Тип нервной системы, стрессоустойчивость и репродуктивная функция. М. Наука. [Semagin VN, Zuhar’ AV, Kulikov MA (1988) Type of nervous system, stress resistance and reproductive function. M. Nauka. (In Russ)].
Штемберг АС (1987) Роль индивидуальных типологических особенностей высшей нервной деятельности в формировании и радиационной устойчивости упроченных двигательно-оборонительных условных рефлексов у крыс. Изв АН СССР сер Биол (4): 547–557. [Shtemberg AS (1987) The role of individual typological features of higher nervous activity in the formation and radiation stability of strengthened motor-defensive conditioned reflexes in rats. Izv AN USSR ser Biol (4): 547–557. (In Russ)].
Симонов ПВ (1976) Условные реакции эмоционального резонанса у крыс. В кн:Нейрофизиологический подход к анализу внутривидового поведения. М. Наука. [Simonov PV (1976) Conditional reactions of emotional resonance in rats. In: Neurophysiological approach to the analysis of intraspecific behavior. M. Nauka. (In Russ)].
Айрапетянц МГ, Хоничева НМ, Мехедова АЯ, Ильяна Вильяр X (1980) Реакции на умеренные функциональные нагрузки у крыс с индивидуальными особенностями поведения. Журн высш нервн деят 5: 994. [Ajrapetyanc MG, Honicheva NM, Mekhedova AYA, Il’yana Vil’yar X (1980) Reactions to moderate functional loads in rats with individual behavioral characteristics. J Higher Nerv Activity 5: 994 (In Russ)].
Хоничева НМ, Ильяна Вильяр Х (1981) Характер поведения в ситуации избегания как критерий оценки типологических особенностей крыс. Журн высш нервн деят 5: 975–983. [Honicheva NM, Il’yana Vil’yar H (1981) The nature of behavior in a situation of avoidance as a criterion for assessing the typological characteristics of rats. J Higher Nerv Activity 5: 975–983.]
Логановский КН (2000) Невроические и психопатологические синдромы в отдаленном периоде воздействия ионизирующих излучений. Журн неврол психиатр им СС Корсакова 4: 15–21. [Loganovskij KN (2000) Neurotic and psychopathological syndromes in the long-term period of exposure to ionizing radiation. J Neurol Psychiatr named after SS Korsakov 4: 15–21. (In Russ)].
Штемберг АС, Базян АС, Лебедева-Георгиевская КБ, Матвеева МИ, Кудрин ВС, Кохан ВС (2013) Влияние облучения протонами высокой энергии на поведение крыс и его нейрохимические механизмы. Авиакосм эколог мед 6: 54–60. [SHtemberg AS, Bazyan AS, Lebedeva-Georgievskaya KB, Matveeva MI, Kudrin VS, Kohan VS (2013) The effect of high-energy proton irradiation on rat behavior and its neurochemical mechanisms. Aerospace Ecol Med 6: 54–60. (In Russ)].
Штемберг АС, Кохан ВС, Кудрин ВС, Матвеева МИ, Лебедева-Георгиевская КБ, Тимошенко ГН, Молоканов АГ, Красавин ЕА, Наркевич ВБ, Клодт ПМ, Базян АС (2015) Воздействие высокоэнергетических протонов в пике Брэгга на поведение крыс и обмен моноаминов в некоторых структурах мозга. Нейрохимия 1: 78–85. [SHtemberg AS, Kohan VS, Kudrin VS, Matveeva MI, Lebedeva-Georgievskaya KB, Timoshenko GN, Molokanov AG, Krasavin EA, Narkevich VB, Klodt PM, Bazyan AS (2015) The effect of high-energy protons at Bragg peak on rat behavior and monoamine metabolism in some brain structures. Neurochemistry 1: 78–85. (In Russ)]. https://doi.org/10.7868/S1027813315010100
McGregor IS (1991) Contrasting effects of stress on medial and sulcal prefrontal cortex self-stimulation. Brain Res Bull 27(2): 225–229.
Ruge H, Wolfensteller U (2013) Functional integration processes underlying the instruction-based learning of novel goal-directed behaviors. Neuroimage 68: 162–172. https://doi.org/10.1016/j.neuroimage.2012.12.003
Tulving E (1972) Episodic and semantic memory. In: Organization of Memory. E Tulving, W Donaldson (Eds). New York. Acad. 381–403.
King D, Zigmond MJ, Finlay JM (1997) Effects of dopamine depletion in the medial prefrontal cortex on the stress-induced increases in extracellular dopamine in the nucleus accumbens and shell. Neurology 1: 141–153.
Zaitsev AV, Lewis DA (2013) Functional properties and short-term dynamics of unidirectional and reciprocal synaptic connections between layer 2/3 pyramidal cells and fast-spiking interneurons in juvenile rat prefrontal cortex. Eur J Neurosci 38 (7): 2988–2998. https://doi.org/10.1111/ejn.12294
Маркель АЛ (1981) К оценке основных характеристик поведения крыс в тесте “открытого поля”. Журн высш нервн деят 2: 301–307. [Markel’ AL (1981) To assess the main characteristics of the behavior of rats in the “open field” test. J Higher Activity 2: 301–307. (In Russ)].
Zhou WL, Antic SD (2012) Rapid dopaminergic and GABAergic modulation of calcium and voltage transients in dendrites of prefrontal cortex pyramidal neurons. J Physiol 590 (16): 3891–3911. https://doi.org/10.1113/jphysiol.2011.227157
Белокопытова КВ, Белов ОВ, Кудрин ВС, Наркевич ВБ, Клодт ПМ, Базян АС, Красавин ЕА, Тимошенко ГН (2015) Распределение моноаминов и их метаболитов в структурах головного мозга крыс в поздние сроки после облучения ионами 12C. Нейрохимия 3: 243–251. [ Belokopytova KV, Belov OV, Kudrin VS, Narkevich VB, Klodt PM, Bazyan AS, Krasavin EA, Timoshenko GN (2015) Distribution of monoamines and their metabolites in rat brain structures in the late stages after irradiation with 12C ions. Neurochemistry 3: 243–251. (In Russ)]. https://doi.org/10.7868/S1027813315030024
Bechara A, Damasio AR (2005) The Somatic Marker Hypothesis: A neural theory of economic decision. Games Econom Behav 52(2): 336–372. https://doi.org/10.1016/j.geb.2004.06.010
Дополнительные материалы отсутствуют.
Инструменты
Российский физиологический журнал им. И.М. Сеченова