

Успехи современной биологии, 2019, T. 139, № 6, стр. 594-612
Экологический эксперимент в базовых и прогнозных биогеоценологических исследованиях
Э. Г. Коломыц *
Институт экологии Волжского бассейна РАН
Тольятти, Россия
* E-mail: egk2000@mail.ru
Поступила в редакцию 13.06.2019
После доработки 12.07.2019
Принята к публикации 15.07.2019
Аннотация
Предложена авторская парадигма экспериментальной биогеоценологии, где управление операционной системой осуществляется с помощью эмпирико-статистических моделей пространственной организации лесных экосистем топологического уровня. Описаны стратегия топологического ландшафтно-экологического анализа, идеология и методы геоэкологического прогнозирования. Представленные базовые и прогнозные модели, раскрывают механизмы формирования топоэкологических систем, их естественную и антропогенную динамику. По известным сценариям глобальных изменений климата описаны закономерности положительной и отрицательной регуляции углеродного цикла лесными биогеоценозами в различных зонально-региональных условиях Волжского бассейна. Экспериментальный экологический анализ проведен на конкретных примерах, с применением методов базовой эмпирической имитации расчетных прогнозируемых ситуаций.
ВВЕДЕНИЕ
Экологический эксперимент включает пять последовательных стадий: гипотезу, планирование, реализацию, статистический анализ и интерпретацию (Хелберт, 2008а). При этом первостепенное значение имеют планирование эксперимента и особенности его реализации; они в равной степени определяют обоснованность исследования и его итоги. Статистический же анализ должен “… увеличить четкость, выразительность и объективность, с которыми результаты представляются и интерпретируются” (там же, с. 12).
В биогеоценологии проведение экологического эксперимента связывается обычно с изучением на комплексных стационарах различных природных режимов (геофизических, геохимических, фитоценотических, почвенных и др.) для весьма ограниченного набора природных комплексов (Сочава, 1969; Крауклис, 1979; Грин и др., 1980; Беручашвили, 1990; Экосистемы …, 2004 и др.). В отличие от лабораторных экспериментов, исследователь здесь “… изучает, по существу, опыты, которые ставит сама природа” (Сукачев, 1975, с. 466). Такие исследования можно отнести к категории управляемых (манипулятивных) экспериментов, необходимыми составляющими которых являются повторность воздействия и контроль; последний отслеживает дрейф во времени и регулирует условия, в которых проводится эксперимент (Хелберт, 2008а).
Однако многолетние режимные наблюдения за геокомпонентами и геопотоками даже в небольшом числе локальных гео(эко)систем на нескольких стационарах одновременно требуют участия значительного персонала в течение длительного времени. Кроме того, известная полимасштабность ландшафтной организации (Хорошев, 2016) предполагает, чтобы стационарная модель представляла собой систему вложенных друг в друга пунктов режимных наблюдений, ключевых участков, трансект и окружения самого стационара (Крауклис, 1979), что весьма усложняет организацию экспериментальных работ.
Задача оказывается гораздо более сложной, когда объектом исследования становятся не отдельные биогеоценологические единицы, а разномасштабная система локальных и тем более региональных природных комплексов с определенным типологическим разнообразием. “Зачастую эти крупномасштабные управляемые эксперименты лишены повторности воздействия …”, однако можно “…. максимизировать ценность эксперимента, контролируя в нем возможно большее количество различных переменных” (Хелберт, 2008б, с. 64–65). Именно в таком направлении должен осуществляться, по-видимому, массовый ландшафтно-экологический эксперимент, что и было осуществлено нами на практике. В настоящем сообщении излагаются результаты многолетнего поиска автора в той области экспериментальной лесной биогеоценологии, которая основана на изучении механизмов пространственной организации гео(эко)систем топологического уровня и использует эргодические свойства этих систем для прогнозных построений их климатогенной динамики.
СТРАТЕГИЯ ЛАНДШАФТНО-ЭКОЛОГИЧЕСКОГО ЭКСПЕРИМЕНТА
Изложим стратегию ландшафтно-экологического эксперимента применительно к разработке научно-методической основы геосистемного мониторинга глобальных изменений для зоны перехода от леса к степи, где лесные сообщества находятся изначально в неустойчивом состоянии. Стратегия должна содержать в себе обоснование путей и методов установления параметров структурно-функциональной организации лесных экосистем юга бореального пояса, наиболее чувствительных к глобальным и региональным климатическим изменениям, которые могут быть использованы для получения региональных и локальных биогеографических и ландшафтных прогнозов в свете предстоящих антропогенных изменений климата.
При поиске рабочих вариантов стратегии решаются, по крайней мере, три взаимосвязанные задачи. Во-первых, следует ответить на вопрос: “существует ли пространственная или временнáя изменчивость описанных объектов и каковы ее структурные особенности?” (Шитиков и др., 2008, с. 213). В данном случае необходимо установить причинные механизмы территориальной организации и функционирования лесных экосистем на зональном экотоне леса и степи. Эти механизмы могут рассматриваться как передаточные звенья локального распространения и трансформации глобально-региональных климатических сигналов. Во-вторых, следует выявить пути прохождения и способы преобразования этих сигналов по системе межкомпонентных и межкомплексных ландшафтных связей с построением соответствующих эмпирических моделей. В-третьих, нужно определить основные параметры структурно-функциональной организации лесных экосистем локального уровня, которые могут быть рекомендованы в качестве объектов регионального и глобального мониторинга биосферы, конечной целью которого являются, как известно (Израэль, 1984; Герасимов, 1985), биоэкологический и геосистемный прогнозы.
Ключевой раздел исследования – разработка количественных методов оценки устойчивости лесных гео(эко)систем к геофизическим воздействиям по параметрам состояния их почвенно-фитоценотического блока. С этой целью необходимо построить эмпирические модели структуры и функционирования лесных природных комплексов, находящихся в критических условиях переходов от леса к степи. Затем по функциональным моделям следует оценить замкнутость круговорота первичного органического вещества в лесных биогеоценозах как фактор устойчивости биосферы на локальном уровне ее организации. Наконец, основываясь на характеристиках устойчивости, предстоит выявить мозаику локальных реакций на глобальные воздействия с помощью прогнозных экспериментов с моделями ландшафтных связей при заданных параметрах ожидаемых изменений регионального климата.
Ландшафтно-экологический эксперимент можно представить в виде определенного алгоритма с построением соответствующих дискретных эмпирико-статистических моделей по определению (Розенберг, 1984). Прослеживая по каналам связей те или иные сдвиги экологических ниш одного признака (явления) по градиенту другого (фактора), исследователь устанавливает систему передаточных функций, осуществляющих распространение внешних вещественно-энергетических сигналов по сети межкомпонентных связей. На основе этого, а также исходя из установленных ранее закономерностей абиотической ординации самих биогеоценозов, выявляется мозаика их цепных реакций на воздействия климатической системы (Коломыц, 2008).
Толчком к преобразованию гео(эко)систем при новом климатическом сигнале служит обострение межкомплексных взаимодействий (Злотин, 1987), выводящее отдельные биогеоценозы или даже весь биогеоценотический покров данного региона из состояния прежнего геотопического типа устойчивости. Основной движущей силой экосистемных перестроек становятся конкурентные отношения экологических ниш растительных сообществ (Свирежев, Логофет, 1978) в пространстве новых гидротермических полей. Благодаря конкуренции происходит экологическая диверсификация – разделение ниш, благоприятствующее развитию более сложных биотических образований (Pianka, 1978). Эволюция осуществляется миграционными процессами в растительном покрове (Васильев, 1946), поэтому экосистемы переходят в миграционный тип устойчивости, с поглощением одних биогеоценозов (редуцентов) другими (абсорбентами), обладающими более устойчивыми климатическими нишами (Коломыц, 2008). В целом фитобиота из фактора, ранее стабилизирующего экосистему (Сочава, 1978), превращается в фактор динамико-преобразующий (Симонов, 1982). Такова исходная идеологическая предпосылка, положенная нами в основу разработки методов ландшафтно-экологических прогнозов.
Рассмотрим более подробно некоторые аспекты экспериментального биогеоценологического анализа с приведением конкретных примеров.
КРУПНОМАСШТАБНАЯ ЛАНДШАФТНО-ЭКОЛОГИЧЕСКАЯ СЪЕМКА
В построении модели гео(эко)системы как сложно организованного многокомпонентного образования “… особенно важен принцип полноты, так как обилие одновременно протекающих обменных процессов требует очень большого количества эмпирических данных” (Ляпунов, 1981, с. 16). Началом экспериментальных биогеоценологических исследований служит осуществление однократной крупномасштабной ландшафтно-экологической съемки на экспериментальных полигонах (Коломыц, 1995, 2008). Съемка позволяет в достаточно короткие сроки полевых работ получить массовую экспресс-информацию о максимально возможном разнообразии структурных и функциональных характеристик топоэкосистем в данном регионе. В этих материалах “представителями” основных вещественно-энергетических процессов выступают их выходные дискретные характеристики. Так, анализ локальных биотических потоков проводится по результатам “работы” отдельных звеньев малого биологического круговорота – различным живым и мертвым фитомассам (скелетной и зеленой, древесной, кустарниковой и травянистой, подстилочной и гумусной), которые, таким образом, рассматриваются как функциональные параметры состояния гео(эко)системы. Абиотическая группа геопотоков в лесном биогеоценозе описывается такими характеристиками, как относительные показатели радиационного баланса под пологом леса, глубина залегания грунтовых вод, температура и влажность почвы, морфологические признаки вертикального почвенного профиля, содержание типоморфных химических элементов в почве и др.
Весь спектр пробных площадей должен охватывать свойственное данному экорегиону основное разнообразие геоморфологических условий и почвенно-фитоценотических структур. Модели состояний гео(эко)систем должны включать в качестве своих элементов признаки основных представителей функциональных блоков природного комплекса. В соответствии с иерархией характе́рных времен различных геокомпонентов (Арманд, 1975) можно выделить следующие геокомпонентные признаки.
А. Независимые переменные – морфометрические характеристики мезо- и микрорельефа: абсолютная высота, экспозиция склона, угол наклона поверхности; тип местоположения и форма микрорельефа.
Б. Частично зависимые переменные – нанорельеф и почвенно-литологические характеристики: выходы на поверхность глыб коренных пород, микротеррасированность склона, покрытие почвы остроугольными валунами, щебнем и дресвой, степень смытости почвы, механический состав горизонта В почвы, степень защебненности верхних почвенных горизонтов.
В. Зависимые переменные – почвенно-растительные и микроклиматические характеристики: мощности горизонтов А0 и А1, температура, влажность и влагосодержание почвы на различных глубинах; характеристики растительного покрова (флористическая и фитоценотическая структура различных биогоризонтов, экологические группы напочвенного покрова); сырая и воздушно-сухая фитомассы травостоя; степень антропогенной дигрессии биогеоценоза и др.
Сбор фактического материала должен быть целенаправленным, пригодным для построения предполагаемых моделей, что и составляет методическую суть биогеоценологического эксперимента. При изучении естественных закономерностей территориальной организации ландшафта объектами исследований должны быть по возможности климаксные, то есть коренные (или близкие к ним), гео(эко)системы, находящиеся в относительно равновесном состоянии. Для оценок же того или иного антропогенного сдвига в ландшафтной структуре изучаемой территории необходимо получить информацию по геосистемам-аналогам, которые подвержены трансформации в разной степени, но находятся в сходных зонально-региональных биоклиматических условиях, на одних и тех же формах микрорельефа и принадлежат к бассейну водотока одного порядка.
При сборе фактического материала необходимо также соблюдение следующих условий.
1. Каждая географическая точка (пробная площадь), характеризующая некоторый гомогенный ареал (выдел ландшафтной фации – биогеоценоз), описывается множеством признаков, фиксируемых одновременно и привязанных к одному и тому же локальному местоположению и к определенной форме микрорельефа (размер пробной площади в наших исследованиях составлял обычно 20 × 20 м, хотя в лесоведении принят стандартный размер 50 × 50 м).
2. Точки должны охватывать по возможности все территориальное многообразие локальных гео(эко)систем по их независимым и частично зависимым признакам в целях наиболее полного выявления “экологической амплитуды” зависимых переменных. Это обеспечивает необходимую фактологическую базу для последующего анализа пространственной сопряженности геокомпонентов и взаимоотношений между природными комплексами.
3. Все признаки должны быть “сквозными”, то есть присутствовать в той или иной мере во всех точках и иметь минимальное число нулевых значений. Тем самым достигается попарная сопоставимость всех точек между собой по всему набору геокомпонентных признаков.
4. Для построения моделей все признаки необходимо выразить численными показателями либо упорядоченными качественными характеристиками, оцененными в баллах. Число градаций (классов) признака не должно быть чрезмерно большим. Опыт статистического анализа показывает (Коломыц, 1995), что при числе пробных площадей менее 100 и при современной точности полевых наблюдений и измерений следует выделять не более 5–7 градаций.
5. Согласно представлениям о принципиально вероятной природе ландшафтных закономерностей (Арманд и др., 1969), исходный эмпирический материал должен быть массовым, пригодным для статистической обработки. Количество пробных площадей должно быть соразмерно числу фиксируемых в поле признаков (Арманд, 1975). Только в этом случае статистический объем выборки будет отражать генеральную совокупность вариаций каждого признака по данной территории. В наших исследованиях на каждом экспериментальном полигоне было описано от 40–50 до 70 точек, при съеме информации на каждой из них по 60-ти параметрам состояния лесного биогеоценоза.
6. При однократной ландшафтно-экологической съемке исследуемые признаки характеризуют “мгновенное” состояние каждого геокомпонента и в своей совокупности дают некоторый пространственно-временной срез природного комплекса с общей траектории его разночастотных изменений (прежде всего, сезонных и многолетних), что обеспечивает сравнимость данных по всем пробным площадям. В связи с этим весь цикл описаний и измерений производится в макси-мально сжатые сроки, в течение которых такие динамичные показатели, как температура и влажность средних и нижних слоев почвы, а также состав и масса травянистой растительности, не испытывают существенных изменений. Более того, с помощью наблюдений на контрольной площадке значения этих признаков приводятся к некоторому единому сроку. Не фиксируются такие высокочастотные признаки, как температура и влажность приземного слоя воздуха.
В наших исследованиях множество структурно-функциональных признаков биогеоценоза, фиксируемое на каждой пробной площади, привязывалось к одному и тому же локальному типу местоположений. Все геокомпонентные признаки являются “сквозными”, то есть взяты по всей совокупности биогеоценозов. Тем самым достигается возможность, во-первых, находить пространственные связи между различными геокомпонентными признаками, а во-вторых, – проводить сопряженный анализ самих топоэкосистем по тому или иному набору геокомпонентов.
Сбор фактического материала на каждой пробной площади и его первичная обработка проводятся в соответствии с известными теоретическими положениями биогеоценологии и методическими руководствами (Родин, Базилевич, 1965; Молчанов, 1971; Смирнов, 1971; Сукачев, 1972; Программа и методика …, 1974; Перельман, 1975; Методы …, 1978; Анучин, 1982; Биологическая …, 1982; Титлянова, 1983; Беручашвили, 1990; Загреев и др., 1992; Швиденко и др., 2008 и др.).
Весь комплекс измеренных и рассчитанных структурно-функциональных признаков, используемых для моделирования, разбивается на шесть геокомпонентных блоков: 1) гидрогеоморфологический; 2) почвенный структурный 3) фитоценотический структурный; 4) фитоценотический функциональный; 5) ландшафтно-геофизический; 6) ландшафтно-геохимический. В наших исследованиях использованы 83 геокомпонентных признака (Коломыц, 2008).
КАТЕНАРНАЯ ОРГАНИЗАЦИЯ БИОГЕОЦЕНОЗОВ И РЕГИОНАЛЬНАЯ СИСТЕМА ЛОКАЛИЗОВАННОЙ ПРИРОДНОЙ ЗОНАЛЬНОСТИ
Наиболее объективные показатели структуры и функционирования природных комплексов выявляются при учете их катенарной организации (Тишков, 2005; Gerrard, 1981), то есть при анализе ландшафтных сопряжений того или иного таксономического ранга. Согласно ландшафтно-геохимической классификации (Глазовская, 1988) выделяются семь типов локальных местоположений (МП) (рис. 1). Они образуют определенную катенарную систему, которая включает следующие звенья (в направлении от водораздела к местному базису эрозии): элювиальное (Э), трансэлювиальное (ТЭ), транзитное (Т), трансаккумулятивное (ТА), аккумулятивное (А), супераквальное (СА) и элювиально-аккумулятивное (Э–А). Соответствующие этим звеньям формы микрорельефа (геотопы) характеризует геоморфологическую основу элементарного природного комплекса – биогеоценоза (Сукачев, 1972), или ландшафтной фации (Сочава, 1978).
Рис. 1.
Катенарная система расположения основных типов местоположений (геотопов) (по Глазовская, 1988). Перенос влаги и вещества в почве и рыхлых отложениях: 1 – в вертикальном направлении; 2 – вдоль склона; 3 – литодинамические и воздушные потоки; 4 – уровень грунтовых вод; 5 – водоем. СА, А, ТА, … – обозначения типов местоположений (см. в тексте).
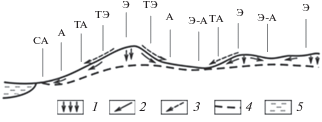
Элювиальные МП на равнине относятся к категории плакоров, по определению (Высоцкий, 1960), которые в максимальной степени отвечают зонально-региональной норме данной территории и, таким образом, являются “внутриландшафтным ядром” ее физико-географического фона, согласно (Сочава, 1978; Крауклис, 1979). Плакоры служат своего рода “нулем” отсчета для оценок как настоящих, так и возможных будущих факторально-динамических отклонений топоэкосистем от их зонально-региональной нормы (Коломыц, 2008). Тем самым упорядочиваются представления о самих природно-территориальных структурах и ожидаемых тенденциях их климатогенных преобразований.
Местные геоморфологические и эдафические факторы преломляют данный климатический фон и создают соответствующие факторально-динамические ряды локальных гео(эко)систем (Крауклис, 1979). Эти ряды состоят из плакоров и множества экстразональных, по (Спрыгин, 1986), природных комплексов, которые служат “представителями” на данной территории различных типов географической среды, нередко весьма удаленных.
По информационным моделям осуществляется абиотическая ординация топоэкосистем. Прежде всего, она проводится в ряду “литоморфность–гидроморфность”, согласно (Колесников, 1956), то есть по ведущему комплексному фактору дифференциации локального экологического пространства. На этой основе устанавливается система экологических ниш почвенно-растительных комплексов в пространстве параметров абиотической среды, что позволяет находить их исходные геотопически устойчивые состояния (Коломыц, 2009). Каждый факторально-динамический ряд представляет собой региональную систему локализованной природной зональности. Она служит топологической моделью эколого-географического пространства, которая способна описывать механизмы реакции локальных экосистем на региональные климатические изменения.
В качестве примера такой модели приведем подтаежный низменно- и возвышенно-равнинный биом Приокско-Террасного заповедника (ПТЗ), расположенного на трансконтинентальном бореальном экотоне Евразии (Коломыц, 2003). Биом представлен хвойными и сосново-широколиственными лесами и их мелколиственными дериватами (табл. 1). Зафиксированное нами при ландшафтно-экологической съемке состояние лесных сообществ заповедника является результатом примерно 100-летнего процесса лесовосстановительных смен после длительного периода антропогенного обезлесения, поэтому все леса заповедника в той или иной степени производны (Оценка и сохранение …, 2000). Псевдокоренными можно считать эвритопные сосново-дубово-липово-еловые леса (группа биогеоценозов (бгц) 3, табл. 1, рис. 2а), занимающие широкий диапазон типов местоположений – от элювиального до трансаккумулятивного.
Таблица 1.
Группы биогеоценозов (ландшафтных фаций) территории Приокско-Террасного заповедника и их условные обозначения (Экспликация к рис. 2 и 3).
| Краткая характеристика | Символ |
|---|---|
| 1. Ксероморфные и мезоксероморфные сосновые и сосново-березовые леса, с осиной и липой, элювиальные и трансэлювиальные, кустарничково-зеленомошные и вейниково-разнотравные |  |
| 2. Мезогидроморфные еловые, сосново-еловые и елово-сосновые леса, элювиальные и трансэлювиальные, кислично-зеленомошные и кустарничково-разнотравные |  |
| 3. Мезоморфные и ксеромезоморфные сосново-липово-дубовые и сосново-липовые леса, с широким диапазоном местоположений (от элювиального до трансаккумулятивного), разнотравные и широкотравные |  |
| 4. Мезоморфные и мезоксероморфные липово-березовые леса, с осиной, а также березняки и осинники, с дубом, липой и елью, трансэлювиальные и транзитные |  |
| 5. Мезогидроморфные ельники, а также сосново-еловые и елово-сосновые леса, транзитные и трансаккумулятивные, черничные зеленомошно- и кислично-разнотравные |  |
| 6. Гидроморфные хвойные и мелколиственные заболоченные леса (сосняки, ельники, березняки, черноольшаники) элювиально-аккумулятивных и супераквальных местоположений |  |
 Условные обозначения к символам групп биогеоценозов. Древостои-доминанты:
Условные обозначения к символам групп биогеоценозов. Древностои-доминанты: а – сосна; б – ель; в – дуб; г – липа, вяз; д – береза, осина; е – ольха черная. Почвообразующие породы: ж – пески; и – супеси и легкие суглинки; к – средние и тяжелые суглинки; л – супесчано-суглинистая морена с валунами; м – карбонатные породы (известняки, доломиты).
Условные обозначения к символам групп биогеоценозов. Древостои-доминанты:
Условные обозначения к символам групп биогеоценозов. Древностои-доминанты: а – сосна; б – ель; в – дуб; г – липа, вяз; д – береза, осина; е – ольха черная. Почвообразующие породы: ж – пески; и – супеси и легкие суглинки; к – средние и тяжелые суглинки; л – супесчано-суглинистая морена с валунами; м – карбонатные породы (известняки, доломиты).
Рис. 2.
Экорегион Приокско-Террасного заповедника. Экологические ниши групп биогеоценозов (а, в) и их зональных групп (б, г) в пространствах типов местоположения и эдафического увлажнения. Условные обозначения: 1 – экологический доминант; 2 – “размытая” часть экологической ниши; 3 – траектория, соединяющая экологические доминанты; 4 – пространство экологической ниши; 5 – анклав; 6 – направление возможной трансгрессии данного явления от его экологического доминанта; 7 – область спорадического распространения явления в пределах данных градаций фактора. К(А;В) – нормированный коэффициент межкомпонентных связей, по (Пузаченко, Скулкин, 1981). Зональные группы биогеоценозов: БТ – бореальная таежная; ББ – бореальная боровая; Н/Б – неморально-бореальная; Б/Н – бореально-неморальная; Н – неморальная. Экологические группы биогеоценозов (по эдафическому увлажнению): К – ксерофитная; МК – мезоксерофитная; М – мезофитная; МГ – мезогигрофитная; Г – гигрофитная. Обозначения групп биогеоценозов см. в табл. 1. Типы локальных местоположений см. в тексте.
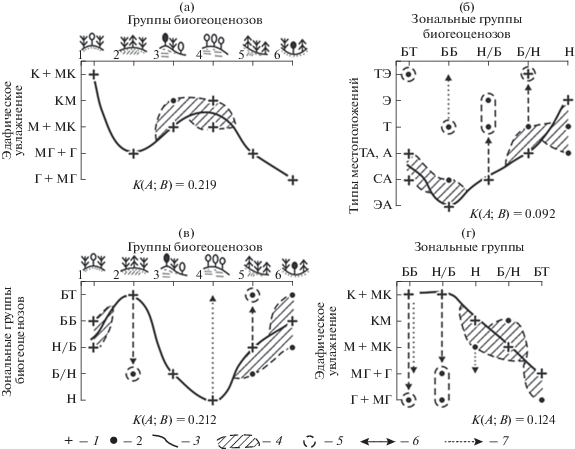
Установлена зональная принадлежность различных групп биогеоценозов в данном экорегионе. Несмотря на то, что современные лесные сообщества заповедника вторичны, все они уже приобрели вполне отчетливые зональные черты (табл. 1, рис. 2в). Последние достаточно четко распределены под влиянием четырех абиотических факторов-эдификаторов: 1) типа местоположения (рис. 2б), 2) глубин залегания известняков и 3) грунтовых вод, 4) поверхностного увлажнения эдафотопа (рис. 2а, 2г). Мезоморфные и ксеромезоморфные сосново-липово-дубовые леса (группа бгц 3) как представители подтаежной зоны Среднеокского бассейна относятся по экологическому доминанту к зонально-климаксной бореально-неморальной группе. Мезогидроморфные сосново-еловые леса плоских междуречий (группа бгц 2), а также их гидроморфные варианты в речных долинах и плоских западинах (группа 6) с близким залеганием моренных суглинков образуют экстразональную группу бореально-таежного литоэдафического климакса лесов, по классификации (Одум, 1975).
Наконец, липово-березовые леса с дубом и осиной (группа бгц 4) как производные от коренных дубо-липняков образуют противоположный, неморальный катенарный “полюс” экстразонального литоэдафического климакса. Данная зональная группа биогеоценозов сформировалась в условиях наиболее близкого залегания известняка и умеренных глубин грунтовых вод, создающих в целом ксеромезоморфное эдафическое увлажнение (рис. 2а).
Полизональность локальных экосистем правомерно рассматривать в качестве формы их реакции на глобально-региональные изменения климата (Коломыц, 2006), поэтому региональные системы-модели локальной полизональности могут имитировать основные направления и масштабы локальных экосистемных перестроек. Эти модели позволяют также оценить, насколько функционально-структурные преобразования топоэкосистем и соответствующие сдвиги природных границ способны преобразовать весь зонально-региональный фон данной территории под действием того или иного климатического тренда.
Итак, в пределах каждого экорегиона мы можем построить гидротермический ряд (или несколько рядов) плакорно-экстразональных биогеоценозов в направлении изменения их термо- или гидроэдафотопов, представляя этот ряд (на основе свойства пространственно-временно́го самоподобия операционной системы) как некоторый аналог соответствующих фоновых климатических изменений (тренда). Далее, анализируя упорядоченные системы локальной зональности, адекватные вектору прогнозируемых изменений климата, мы получаем возможность имитировать основные направления и масштабы наиболее вероятных экосистемных перестроек в рассматриваемом регионе. Пространственные последовательности заменяются на временны́е и по векторному спектру топологической полизональности намечаются соответствующие цепочки местных биогеоценотических переходов. Создается общий сценарий реакции локальных экосистем на те или иные климатические сдвиги. Таков механизм локальной эмпирической имитации регионального биоклиматического тренда, лежащей в основе методики ландшафтно-экологического прогнозирования.
ГИДРОЭДАФИЧЕСКИЕ УСЛОВИЯ КРИТИЧЕСКИХ СОСТОЯНИЙ ЛЕСНЫХ ЭКОСИСТЕМ
Для равнинных бореальных и суббореальных ландшафтов эмпирически установлено, что основной пропускной канал связей региональных и локальных экосистем с фоновой климатической системой проходит через летнее влагосодержание почвы, которое, с одной стороны, служит достаточно надежным геофизическим индикатором состояния экосистем, а с другой – является наиболее мощным экологическим фактором, предопределяющем их территориальную организацию (Коломыц, 2003, 2008, 2010). Для основного водосбора Волжского бассейна по данным 120 агрометеостанций и 5 воднобалансовых станций найдены тесные связи июльских запасов продуктивной влаги в слоях почвы 0–20 (W-20), 0–50 (W-50) и 0–100 см (W-100) с годовым коэффициентом атмосферного увлажнения Высоцкого–Иванова (Kувл(1) = r(год)/E0, где r(год) – годовое количество осадков, а E0 – годовая испаряемость). Применительно к лесным экосистемам эти связи имеют следующий вид с соответствующими коэффициентами корреляции (R) и детерминации (R2):
(1)
$\begin{gathered} \left( {W{\text{ - }}20} \right) = {\text{ехр}}{\kern 1pt} \text{[}2.281 + 0.8068{{K}_{{{\text{увл}}}}}\left( 1 \right)]; \\ {\text{R}} = 0.83;\,\,\,\,{{{\text{R}}}^{2}} = 0.69; \\ \end{gathered} $(2)
$\begin{gathered} \left( {W{\text{ - }}50} \right) = {\text{ехр}}{\kern 1pt} \text{[}2.622 + 1.071{{K}_{{{\text{увл}}}}}\left( 1 \right)]; \\ {\text{R }} = 0.89;\,\,\,\,{{{\text{R}}}^{2}} = 0.79; \\ \end{gathered} $(3)
$\begin{gathered} \left( {W{\text{ - }}100} \right) = {\text{ехр}}{\kern 1pt} \text{[}3.225 + 1.134{{K}_{{{\text{увл}}}}}\left( 1 \right)]; \\ {\text{R }} = 0.88;\,\,\,\,{{{\text{R}}}^{2}} = 0.77.~ \\ \end{gathered} $В свою очередь, параметр Kувл(1) определяется почти исключительно среднеиюльской температурой (Коломыц, 2003), в то время как влияние температуры января незначимо. Очевидно, главный фактор климатогенной трансформации природных экосистем в регионе – термические условия вегетационного периода. С другой стороны, существует достаточно тесная зависимость распределения лесных топоэкосистем от летнего влагосодержания почвы. Так, неморальнолесные биогеоценозы Среднего Поволжья имеют информационные коэффициенты сопряженности, по (Пузаченко, Скулкин, 1981), с параметрами (W-20), (W-50) и (W-100) соответственно 0.154–0.173, 0.128–0.162 и 0.139–0.200. Это существенно превышает критический порог значимости связей – 0.070 (Коломыц, 1995).
Предпрогнозный этап ландшафтно-экологического анализа Волжского бассейна состоял в определении критических значений летнего содержания почвенной влаги, при которых совершаются переходы зональных типов растительного покрова: от смешанных лесов к широколиственным, а затем первых и вторых – к лесостепным формациям, то есть к комплексу островных лесов и луговых степей (Коломыц, Сурова, 2010). Для этой цели были использованы полученные ранее корреляционные связи зональных и подзональных границ в различных секторах бассейна с параметром Kувл(1) (Коломыц, 2003). Однако данный входной параметр не учитывает местное разнообразие сложных процессов преобразований тепла и круговорота влаги, дающих целый спектр состояний гидроэдафотопов при одних и тех же фоновых соотношениях поступающей солнечной энергии и выпадающих осадков (Варлыгин, Базилевич, 1992).
В результате этих процессов возникает определенное топологическое множество коэффициентов увлажнения, только теперь уже не атмосферного, а эдафического – Kувл(2). Для определения последних использованы представленные выше уравнения связи параметра Kувл(1) со среднеиюльскими влагозапасами в почве. Подставляя в эти формулы базовые значения запасов почвенной влаги для каждой группы лесных биогеоценозов в пределах данного экорегиона, мы получили соответствующее множество локальных коэффициентов эдафического увлажнения Kувл(2). Значения параметра Kувл(2) оказались в целом в 1.5–2 раза более высокими по сравнению с Kувл(1). Это позволяет объяснить, почему лесные сообщества могут устойчиво существовать при явном дефиците атмосферного увлажнения.
На основе корреляционных связей июльских запасов продуктивной влаги в плакорных лесных экосистемах с параметром Kувл(2) были получены два ряда критических влагозапасов Wкрит (таксономических норм и минимальных значений) для трех основных зональных рубежей на территории Среднего Поволжья: а) перехода от бореального пояса к суббореальному; б) смены широколиственных лесов типичной и южной лесостепью; в) перехода от южной лесостепи к северной степи. По определению А.Д. Арманда (1989), при критическом состоянии гео(эко)системы происходит ее качественная перестройка, включающая структурные изменения. Представленные в табл. 2 июльские критические влагозапасы почвы сняты с графиков статистических связей, по которым были получены расчетные формулы (1)–(3). Значения Wкрит характеризуют конкретные состояния гидроэдафотопов, при которых должны совершаться указанные наиболее резкие (скачкообразные) функционально-структурные изменения в геосистемах плакорного ряда, отображающих, как уже говорилось, зонально-региональный фон данной территории.
Таблица 2.
Средние и минимальные (в скобках) критические значения июльских запасов продуктивной влаги в почве, при которых происходят зональные смены плакорных растительных сообществ на территории Среднего Поволжья
| Ландшафтно-зональные условия (экорегион) | Слой почвы, см |
Уровни критических запасов продуктивной влаги в почве, мм | ||
|---|---|---|---|---|
| I – для смешанных лесов (зональный переход к широколиственным лесам) | II – для сплошного неморального лесного покрова (зональный переход к типичной и южной лесостепи) | III – для островных парковых лесов и редколесий (зональный переход к северной степи) | ||
| Подзона широколиственных лесов (Нижегородское Предволжье) | 0–20 | 46 (37) | 40 (32) | 30 (24) |
| 0–50 | 108 (87) | 92 (74) | 62 (48) | |
| 0–100 | 215 (180) | 185 (147) | 122 (102) | |
| Подзона типичной лесостепи (Присурье) |
0–20 | 42 (34) | 38 (29) | 29 (23) |
| 0–50 | 96 (78) | 85 (70) | 57 (43) | |
| 0–100 | 190 (150) | 170 (132) | 110 (87) | |
| Подзона южной лесостепи (Самарская Лука) | 0–20 | 38 (32) | 36 (30) | 28 (22) |
| 0–50 | 84 (70) | 79 (65) | 52 (38) | |
| 0–100 | 170 (130) | 155 (120) | 100 (75) | |
Обращает на себя внимание сравнительно узкий диапазон критических значений летнего влагосодержания почвы между первым и вторым уровнями, то есть между северной и южной границами неморальнолесной подзоны. Это говорит о том, что при интенсивном развитии термоаридного тренда процесс остепнения и последующего распада лесного покрова может охватить в равной степени как широколиственные, так и смешанные формации, создавая не только мелколиственно-дубравную, но и хвойную типичную лесостепь.
ИДЕОЛОГИЧЕСКИЕ ОСНОВЫ ПРОГНОЗНОГО ЭКОЛОГИЧЕСКОГО ЭКСПЕРИМЕНТА
Стратегия ландшафтно-экологического прогноза состоит в первоначальной идентификации природно-территориальных единиц определенным значениям базовых климатических условий и в последующей количественной оценке наиболее вероятных преобразований этих объектов в соответствии с ожидаемыми изменениями климата на заданные сроки. При этом, в отличие от известных отечественных и зарубежных подходов (Пузаченко, Скулкин, 1981; Кобак и др., 2002; Emanuel et al., 1985; Leemans, 1992 и др.), предполагается многозначный характер трансформации экосистем при фиксированном значении геофизического тренда, когда новое их состояние может иметь черты не одного, а нескольких состояний, существующих на начальный момент времени.
Выдвигаемый нами основной принцип ландшафтно-экологического прогнозирования гласит: климатически обусловленное функциональное преобразование одной гео(эко)системы в другую тем значительнее, чем меньше была степень пересечения их климатических ниш в начальном состоянии и чем больше окажется величина пересечения ниш после сближения систем по данному геофизическому признаку. Этот принцип соответствует одному из основных положений экологии сообществ. Обостренная конкуренция между популяциями приводит к трансформации самого сообщества в том направлении, которое отвечает новому состоянию среды, согласно закону конкурентного исключения Гаузе (Розенберг и др., 1999), при условии, что популяции-иммигранты по своей конкурентоспособности доминируют над местными популяциями.
Экспериментальный характер методики ландшафтно-экологического прогнозирования (Коломыц, 2008) состоит, прежде всего, в том, что в расчетных моделях будущие экологические ситуации воспроизводятся с помощью их эмпирической имитации пространственно-распределенными параметрами базовых экологических ниш изучаемых объектов. Сам прогноз является по существу управляемым экспериментом, по определению (Хелберт, 2008б). Исследователь задает входные параметры в данную операционную систему и получает на выходе картину прогнозируемых структурных и функциональных состояний изучаемых объектов в данной статистической выборке с выявлением новых вневыборочных объектов. В нашем прогнозном анализе в экспериментальный методический ансамбль вошли: 1) локальная эмпирическая имитация регионального биоклиматического тренда по моделям бинарной гидро- и термоэдафической ординации лесных топогеосистем ( Коломыц, 2009; Коломыц, Шарая, 2012); 2) использование свойства полизональности локальных геосистем как способа их реакции на глобальные изменения климата (Коломыц, 2006).
ПРОЦЕДУРА БИОГЕОЦЕНОЛОГИЧЕСКОГО ПРОГНОЗИРОВАНИЯ
Вероятностный ландшафтно-экологический прогноз осуществляется с помощью операций с современными и предполагаемыми гидротермическими нишами экосистем (Коломыц, 2008; Залиханов и др., 2010). Для этого каждая ниша должна быть представлена некоторым “размытым” (дескриптивным) множеством в виде вектора-строки (или вектора-столбца). Составляющими вектора служат нормированные частные коэффициенты связи, по (Пузаченко, Скулкин, 1981), которые рассматриваются как “весовые” коэффициенты.
Приведем алгоритм прогнозно-экологических расчетов, проведенных нами для лесных биогеоценозов различных экорегионов Волжского бассейна.
I. Предварительное установление зонально-региональных связей июльских запасов продуктивной почвенной влаги с коэффициентом увлажнения. Для этого использовались данные 120 агрометеостанций Волжского бассейна и его окружения, а также пяти воднобалансовых станций (для перехода от сельскохозяйственных полей к лесопокрытым площадям).
II. Расчеты изменения летних запасов влаги в слоях почвы 0–20, 0–50 и 0–100 см различных топоэкосистем на конец каждого прогнозируемого этапа (1985–2050; 2050–2100; 2100–2200; 2200–2300 гг.). По найденным экспоненциальным зависимостям влагосодержания почвы различного мехсостава от средней июльской температуры воздуха (Коломыц, 2008) были рассчитаны (по каждому экспериментальному полигону) базовые и прогнозные значения зонально-региональной (фоновой) нормы почвенных влагозапасов. Далее, для каждого из указанных слоев почвы и каждого прогнозного срока вычислялся коэффициент как отношение фоновых запасов влаги этого срока к фоновым же запасам базового периода. Наконец, умножением данного коэффициента на конкретные влагозапасы базового периода по всем пробным площадям полигона были получены прогнозируемые значения влагозапасов на этих площадях.
III. Локальный ландшафтно-экологический прогноз: оценка вероятностей и скоростей взаимных переходов в системе биогеоценозов, согласно установленным трендам климатогенных изменений летних запасов почвенной влаги. Под переходом понимается такая смена функционального, а затем и структурного состояния данного объекта, при которой объект принимает (с той или иной долей вероятности) черты других объектов-прообразов благодаря тому, что его гидроэдафическая ниша при своем изменении все более пересекается с нишами этих объектов. Вероятности переходов одних биогеоценозов в другие, а также их стабилизации (устойчивости к данному климатическому сигналу) представлены на графоаналитических моделях (рис. 3).
Рис. 3.
Экорегион Приокско-Террасного заповедника. Орграфы функциональных ландшафтно-экологических переходов, согласно прогнозно-климатической модели E GISS, (а) – 2050 г., (б) – 2200 г., между группами фаций (биогеоценозов), имеющих различную зональную принадлежность и находящихся на различных уровнях базового функционирования. Вероятности переходов: 1 – 0.10 и менее; 2 – 0.11–0.20; 3 – 0.21–0.30; 4 – 0.31–0.40; 5 – 0.41–0.50; 6 – 0.51–0.60; 7 – 0.61–0.70; 8 – 0.71–0.80. Обозначения групп биогеоценозов см. в табл. 1. Южнотаежная группа биогеоценозов является вневыборочной (экстразональной).
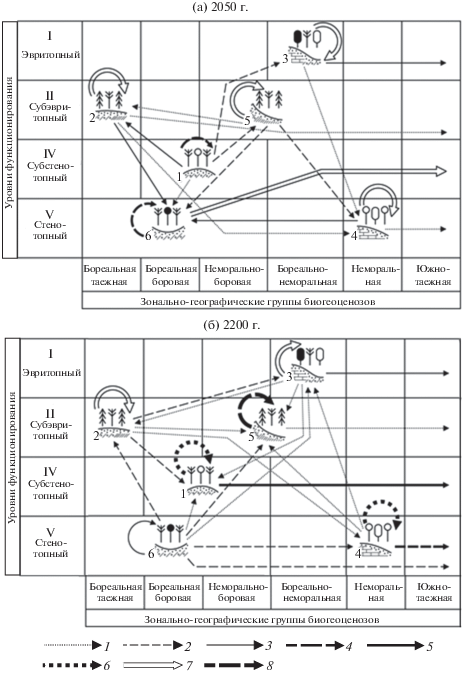
Сама процедура прогнозирования уже излагалась нами достаточно подробно (Коломыц, 2003, 2008; Залиханов и др., 2010). Отметим две важные особенности прогноза. Во-первых, учитывается вероятность “остаточных” переходов того или иного объекта в другие прообразы-объекты, отсутствующие в данной операционной системе (выборке), то есть во вневыборочные объекты. Для маргинальных лесных экосистем типичной и южной лесостепи эти переходы оказываются доминирующими и даже единственно возможными. Во-вторых, на основе мер функционального изоморфизма топоэкосистем, характеризующих степень их близости по основным параметрам биологического круговорота (Коломыц, 2008), устанавливаются наиболее предпочтительные переходы, с минимальными проявлениями нелинейности систем (Зейдис и др., 2001), что позволяет упорядочить всю картину траекторий экосистемных трансформаций.
Расчеты скоростей ландшафтно-экологических переходов проводятся по методам теории марковских цепей (Harbaugh, Bonham-Carter, 1970; Jeffers, 1978). Вычисляются относительные и абсолютные скорости переходов, время поглощения редуцентов абсорбентами и другие параметры, позволяющие более разносторонне представить ту или иную прогнозируемую ситуацию.
Для высокогорий Центрального Кавказа прогнозные расчеты были аналогичны, с той лишь разницей, что в их основе лежат базовые эмпирические связи высокогорных лесов и лугов с летними температурами почвы на глубинах 20, 30 и 40 см (Залиханов и др., 2010; Коломыц, Шарая, 2012).
Следует отметить важное методическое ограничение, свойственное всем статичным пространственным эмпирико-статистическим моделям (Lischke et al., 1998). Приоритетным объектом ландшафтно-экологического прогнозирования здесь является функциональная релаксация как первоочередная реакция гео(эко)систем на внешнее воздействие (Коломыц, 2008). При ожидаемых глобальных изменениях климата на ближайшие десятилетия речь может идти в первую очередь о функциональном, а не о структурно-морфологическом экологическом прогнозе. Представленные на прогнозных моделях вероятности переходов одних гео(эко)систем в другие указывают на то, в каких ландшафтно-экологических условиях будут функционировать эти системы.
РЕЗУЛЬТАТЫ БИОГЕОЦЕНОТИЧЕСКОГО ПРОГНОЗА
Для примера снова взят Приокско-Террасный заповедник, а климатический прогноз – по новейшей глобальной модели E GISS (Hansen et al., 2007). Модель дает для данного региона холодно-гумидный тренд до середины XXI столетия с последующей слабо выраженной термоаридизацией климата. Базовым считается период метеонаблюдений 1881–1985 гг., с конца которого, собственно, и начались глобальные антропогенные изменения климата (Climate change …, 1996).
На этапе регионального похолодания к 2050 г. ожидается общая достаточно высокая устойчивость почти всех биогеоценозов заповедника (мера стабилизации составляет более 60%), однако намечается определенная тенденция (до 25%) трансформации ксероморфных ТА зеленомошно-вейниковых сосновых боров в соседние Э мезогидроморфные кислично-разнотравные ельники и елово-сосновые леса. Последние же, в свою очередь, начнут преобразовываться с еще большей интенсивностью (около 30%) в гидроморфные ЭА хвойно-мелколиственные кустарничково-сфагновые сообщества междуречных понижений (рис. 3а). В том же направлении изменятся (на 27%) и более дренированные мезоморфные Т и ТЭ дубово-липово-березовые разно- и широкотравные леса. Более того, возникнут явные признаки трансгрессии в данный подтаежный экорегион южнотаежных сообществ. Последние будут проникать главным образом по речным долинам и междуречным западинам, замещая более чем на 65% упомянутые выше переувлажненные леса. Вторая, менее интенсивная, волна трансгрессии (до 30%) охватит умеренно дренированные водоразделы и склоны с мезофильными сосново-липово-дубовыми лесами. Определенные южнотаежные признаки (до 6–9%) проявятся также и в остальных биогеоценотических группах лесов данного экорегиона.
Таким образом, даже относительно слабый холодно-аридный сигнал, со сменой годового коэффициента увлажнения с 1.52 до 1.78–1.79, способен вызвать весьма заметный, хотя и достаточно локализованный, процесс распространения лесных и лесоболотных ассоциаций южной тайги вплоть до южной границы подтаежной зоны.
К 2100 г., когда по модели E GISS условия атмосферного увлажнения приблизятся к современным (Kувл = 1.50), заметно усилится стабилизация биогеоценотических групп со слабо выраженными отклонениями от базовых состояний. В дальнейшем к концу XXII столетия в экорегионе начнет развиваться термоаридный климатический тренд (Kувл = 1.36) и наметятся биогеоценотические переходы, обратные вышеописанным. Так, заболоченные А и СА хвойно-мелколиственные леса будут на 14–15% замещаться мезогидроморфными Э и ТЭ ельниками и сосняками, которые, в свою очередь, с такой же скоростью начнут трансформироваться в ксероморфные березово-сосновые леса на аналогичных местоположениях (рис. 3б). А в ареалы последних произойдет значительное внедрение (почти на 48%) уже неморальных северо-лесостепных сообществ. К концу XXIII в. все эти процессы несколько ослабнут в связи с некоторым повышением атмосферного увлажнения.
РАСЧЕТ УГЛЕРОДНЫХ БАЛАНСОВ ЛЕСНЫХ ЭКОСИСТЕМ ПРИ ИЗМЕНЕНИЯХ КЛИМАТА
На примере лесных экосистем Волжского бассейна разработаны прогнозные модели динамики дискретных параметров биологического круговорота и углеродного баланса для двух альтернативных климатических трендов – регионального потепления и похолодания. Эти модели описывают общую будущую картину биотической регуляции углеродного цикла в системе лес–атмосфера по численным климатическим сценариям с учетом функционально-структурных перестроек биогеоценозов. Известные математические (балансовые) модели лесных экосистем описывают процессы движения органического вещества между различными трофическими уровнями (Теоретические основы …, 1983; Пулы и потоки …, 2007 и др.). Однако эти континуальные модели характеризуют круговороты углерода с нераспределенными в пространстве параметрами – либо на глобальном уровне, либо для отдельных зональных типов биомов. На региональном и тем более локальном уровнях необходимо учитывать существенную ограничивающую роль дискретных геоморфологических и гидроэдафических факторов.
Нами рассматривались следующие дискретные параметры малого биологического круговорота: 1) BS – общая скелетная масса древостоя, подроста и подлеска; BV – общая зеленая масса всех ярусов лесного сообщества; BR – общая масса корней: WD – мертвая древесно-кустарниковая фитомасса; ML – масса лесной подстилки (опад + + отпад + мор); HU – масса лабильного гумуса в гор. А1 (примерно в верхнем 20-сантиметровом слое почвы). Динамика указанных фитомасс (их положительные либо, наоборот, отрицательные изменения в прогнозируемом интервале времени), по существу, и есть тот механизм, который осуществляет биотическую регуляцию углеродного цикла.
Для оценок изменений содержания углерода в различных биотических компонентах нами применен известный лесоводственный метод (Морозов, 1949; Цельникер, 2006), основанный на динамике перечисленных фитомасс. Изменение (∆С) массы потока углерода в системе почва–растение–атмосфера можно представить в следующем виде (Кобак, 1988):
где ∆С(Fa) – поток СО2 над растительным покровом; Rm – выделение СО2 в результате жизнедеятельности почвенных и наземных сапротрофов (преимущественно бактерий и грибов), разлагающих гумус, лесную подстилку и скелетный детрит; NPP – чистая первичная продукция биогеоценоза (надземная + корневая). Используя дискретные показатели малого биологического круговорота, приведенное выше уравнение запишем в развернутой форме:(5)
$\begin{gathered} \Delta {\text{С}}(Fa) = \Delta {\text{С}}(WD) + \Delta {\text{С}}(ML) + \Delta {\text{С}}(HU) \\ --\,\,\Delta {\text{С}}(BS)--\Delta {\text{С}}(BV)--\Delta {\text{С}}(BR). \\ \end{gathered} $Здесь представлены углеродные пулы, соответствующие упомянутым выше фитомассам. По данному балансовому уравнению рассчитываются возможные изменения потоков углерода между почвенно-растительным покровом и атмосферой в различные прогнозируемые сроки (по заданным глобальным климатическим моделям) для каждой группы лесных сообществ во всех рассматриваемых региональных экосистемах. Каждый член правой части уравнения (5) может иметь как положительные, так и отрицательные значения. При положительных величинах первые три слагаемые дают прибавку эмиссии СО2 из почвенно-растительного покрова в атмосферу, а вторые – убыль этого потока. В данном случае пул мортмассы выступает в качестве дополнительного источника углерода, а живая фитомасса играет роль его стока (депонирования). При отрицательных же значениях указанных параметров картина обратная: в редуцированной ветви биологического круговорота поступление диоксида углерода в атмосферу сокращается, а автотрофный биогенез, снижающий свою интенсивность, потребляет меньшее количество СО2, компенсируя тем самым возникший дефицит углекислого газа в атмосфере.
В итоге складывается общий баланс изменений углеродного обмена между биогеоценозами и атмосферой [±∆С(Fa)], который должен показать, поглощает ли данная группа лесных биогеоценозов дополнительное количество СО2 из атмосферы в результате обусловленных глобальным потеплением сдвигов в биологическом круговороте или, наоборот, становится источником его дополнительных выбросов. В первом случае осуществляется отрицательная обратная связь, направленная на выполнение принципа Ле Шателье по стабилизации или даже ослаблению первичного термоаридного климатического сигнала, а во втором – положительная обратная связь, которая ведет к усилению парникового эффекта атмосферы, а, следовательно, и самого потепления.
При похолодании те же слагаемые углеродного баланса играют прямо противоположную роль, если оценивать их вклад в ослабление этого похолодания. При отрицательных значениях суммы (ΔС(BS) + ΔС(BV) + ΔС(BR)) и положительных (ΔС(ML) + ΔС(WD) + ΔС(HU)) можно говорить о позитивной биотической регуляции холодного климатического сигнала, направленной на его ослабление. В противном случае эта регуляция окажется негативной, то есть усиливающей процесс похолодания.
МЕХАНИЗМЫ РЕГУЛЯЦИИ УГЛЕРОДНОГО ЦИКЛА В ЛЕСНЫХ БИОГЕОЦЕНОЗАХ
Для выявления регулирующей роли лесов в углеродном цикле необходим длительно устойчивый гидротермический сигнал. Механизмы этой регуляции нами рассмотрены на основе прогнозируемых изменений биологического круговорота в лесных биогеоценозах при заданных сценариях климатического прогноза до 2150–2200 гг. по двум глобальным моделям (табл. 3): а) экстремальной HadCM3, версия А2 (Pope et al., 2000) и б) более умеренной E GISS (Hansen et al., 2007). Как и большинство известных моделей семейства AOGCMs, первая модель предсказывает термоаридный климатический тренд по всей Средней полосе Русской равнины. Экологический прогноз по этой модели позволяет оценить реакцию лесных экосистем на такие глобальные климатические изменения, которые могут превышать порог выносливости основных лесообразующих пород и привести к распаду на обширных пространствах не только неморальных, но и бореальных лесов. Вторая же модель достаточно реалистично отображает столетнюю климатическую динамику в пределах Европейского континента. В своем последнем варианте (Schmidt et al., 2006) она дает более сложную картину: холодно-аридный тренд до середины XXI в. с последующей слабо выраженной термоаридизацией климата.
Таблица 3.
Отклонения основных климатических параметров от их базовых значений, согласно прогнозно-климатическим моделям HadCM3 и E GISS*
| Полигоны (биомы) | Kувл базовый |
Модель HadCM3, 2150 г. (экстремальный термоаридный тренд) | Модель E GISS, 2050 г. (слабо выраженный холодно-гумидный тренд) |
||||||
|---|---|---|---|---|---|---|---|---|---|
| Δtянв | Δtиюл | Δrгод | Кувл | Δtянв | Δtиюл | Δrгод | Кувл | ||
| Жигули | 0.95 | +6.9 | +8.9 | +24 | 0.33 | –1.0 | –0.5 | –60 | 0.92 |
| Присурье | 1.07 | +7.2 | +7.0 | +21 | 0.45 | –1.0 | –1.0 | –71 | 1.15 |
| Приокско-Террасный заповедник | 0.52 | +6.5 | +5.1 | –31 | 0.74 | –1.3 | –1.9 | –71 | 1.79 |
Прогнозная климатическая модель HadCM3 дает повышение к концу XXI в. среднеиюльской температуры в Среднем Поволжье на 5.5–7.0°C. Это говорит о том, что к концу столетия весь юг бореального пояса на Русской равнине окажется в суббореальных климатических условиях. На этом фоне годовое количество осадков почти не изменится, либо увеличится незначительно. Прибавка осадков не сможет нейтрализовать рост температуры, поэтому коэффициент увлажнения будет весьма заметно снижаться. Таким образом, по всему Волжскому бассейну будет развиваться термоаридный климатический тренд. Начнется прогрессирующее летнее иссушение почвы – всеобщее явление, ожидаемое при глобальном потеплении на юге умеренного пояса практически всеми прогнозно-климатическими моделями (Climate change …, 1996). Противоположную картину следует ожидать в первые десятилетия при реализации холодно-аридного климатического тренда по модели E GISS (табл. 3).
Аналитическое прогнозное моделирование позволило получить весьма разнообразную картину прогнозируемых климатогенных сдвигов в функционировании лесных биогеоценозов Волжского бассейна и изменений их углеродного бюджета, даже при одном и том же климатическом сигнале. Прежде всего, установлено, что за достаточно большие интервалы времени максимальное по абсолютным значениям изменение испытывает не детритогенез, а автотрофный биогенез. При этом львиную долю в сдвигах продуктивности составляет скелетная древесно-кустарниковая фитомасса BS и главным образом масса древесины BW (рис. 2 и 3). Эта закономерность характерна для всех рассматриваемых лесных экосистем и при любом климатическом сигнале.
При развитии резко выраженного термоаридного климатического тренда по модели HadCM3 за грядущий 100–150-летний период следует ожидать следующие сдвиги в углеродном цикле (рис. 4а, табл. 4).
Рис. 4.
Прогнозируемые изменения содержания углерода в различных пулах биогеоценозов Приокско-Террасного заповедника на 2150 г., согласно климатическим моделям HadCM3 (а) и на 2050 г. – по модели E GISS (б). Обозначения групп биогеоценозов см. в табл. 1.
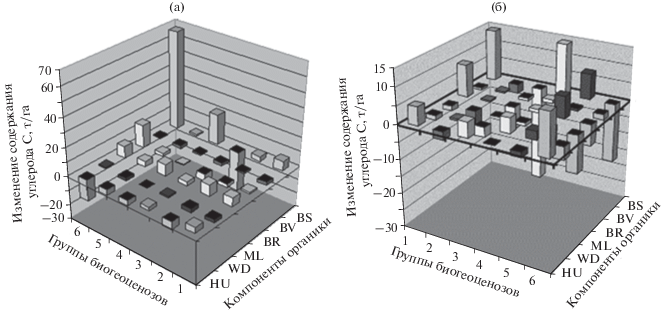
Таблица 4.
Прогнозируемые, согласно глобальным климатическим моделям, суммарные изменения содержания углерода (т/га) по отношению к базовому периоду в плакорных лесных биогеоценозах различных регионов Среднего и Верхнего Поволжья
| Региональная экосистема (экспериментальный полигон, по Коломыц, 2008) |
Базовые значения |
Изменения в прогнозные сроки, гг. | |||
|---|---|---|---|---|---|
| модель HadCM3 | модель E GISS | ||||
| 2050 | 2150 | 2050 | 2200 | ||
| Южная лесостепь, на границе с северной степью (Жигули) | 216.36 | –90.81 | –100.47 | –16.67 | –49.0 |
| Средняя (типичная) лесостепь (Присурье) | 221.90 | –40.75 | –32.08 | –49.00 | –35.45 |
| Неморальнолесная подзона, у ее северной границы (Зеленый Город) | 206.92 | 36.37 | 63.34 | –34.02 | –8.11 |
| Подтаежная зона, у ее южной границы (Приокско-Террасный заповедник) | 243.43 | –32.84 | –31.45 | –33.08 | –28.47 |
| У северной границы неморальнолесной подзоны, низменно-зандровое полесье (Выкса) | 247.41 | –31.36 | –29.56 | –31.36 | –14.48 |
| Южная полоса подтаежной зоны в Низменном Заволжье (Керженец) | 144.93 | –28.25 | –31.68 | –12.03 | –20.84 |
1. Содержание углерода, законсервированного в скелетной фитомассе лесов Волжского бассейна, может измениться на ±(20–40)% от базовой величины, что указывает на весьма существенную регулирующую роль, которую должна сыграть продуктивность южнотаежных, смешанных и неморальных лесов региона в их углеродном обмене с атмосферой. Глобальное потепление вызовет в одних случаях резкое падение продуктивности, в других – столь же значительное ее увеличение, что обрисует весьма противоречивую картину биотической регуляции углеродного цикла. К 2100 г. изменения содержания углерода в BS составит от –25–40 т/га в южной лесостепи до +85–105 т/га в хвойно-широколиственных лесах.
2. Наиболее сильно сократится продуктивность маргинальных лесных сообществ на зональном экотоне леса и степи (семигумидная экосистема Жигулевских гор) – прежде всего в плакорных липняках и дубравах эрозионно-суглинистых возвышенностей, а также в лесах малых долин (табл. 4). Их роль как стока углерода из атмосферы резко снизится. Этот процесс будет сопровождаться существенным ускорением разложения скелетного дебриса WD и гумуса HU, хотя масса лесной подстилки ML увеличится из-за возрастающего дефицита осадков. В целом детритная ветвь метаболизма здесь будет способствовать дополнительному накоплению СО2 в атмосфере и совместно с сокращением BS вызовет прогрессирующее нарушение принципа Ле Шателье на южной границе лесного пояса.
3. В подзоне типичной лесостепи (семигумидный экорегион Присурье) плакорные дубово-липовые леса возвышенных равнин до 2050 г. будут иметь отрицательный показатель ∆С(BS) = –6.2 т/га, однако в последней четверти XXI в. они начнут уже дополнительно консервировать углерод в древесно-кустарниковой фитомассе, и к 2150 г. здесь ∆С(BS) будет равен +9.4 т/га.
4. В возвышенно-равнинных смешанных лесах неморальнолесной подзоны (Нижегородское Предволжье, экорегион Зеленый Город) и на южной границе подтайги (гумидный экорегион ПТЗ) при потеплении ожидается, напротив, существенное повышение продуктивности с положительной биотической регуляцией углеродного цикла. Этот процесс проявится особенно значительно в ТЭ ксероморфных и СА гидроморфных бореальных лесах, которые сыграют роль основного стока углерода. Одновременно произойдет активизация разложения мортмассы и гумуса, что несколько снизит положительный для углеродного баланса эффект роста продуктивности лесов.
6. Процесс глобального потепления должен привести к развитию в Средней полосе Русской равнины двух взаимно противоположных феноменальных явлений биотической регуляции углеродного цикла. Крайний юг лесной зоны, в том числе зональный экотон леса и степи в средневолжской части равнины, окажется ареной значительного дополнительного накопления углерода в атмосфере (рис. 3, табл. 4). Соответственно расстроится сбалансированность углеродного цикла как стабилизирующего механизма состояния наземных экосистем. Система биогеоценоз–атмосфера в этих зональных условиях будет функционировать с положительной обратной связью, что вызовет местное усиление парникового эффекта и ускорит общий процесс деградации и исчезновения маргинальных лесов в южной и типичной лесостепи.
Основную положительную регуляцию углеродного цикла при глобальном потеплении, направленную на снижение парникового эффекта атмосферы, будут выполнять коренные широколиственно-темнохвойные леса и вторичные липово-березовые и мелколиственные сообщества возвышенных равнин в неморальнолесной подзоне и в южной полосе подтаежной зоны. Второстепенную роль сыграют некоторые группы подтаежных и неморальных сосновых лесов на низменных зандрах. В типичной лесостепи существенную позитивную роль сыграют елово-сосновые и сосново-широколиственные леса возвышенных равнин, а также переувлажненные леса речных долин. По-видимому, это единственные лесные формации, которые смогут оказать определенную поддержку устойчивости континентальной биосферы при глобальном потеплении. Регулирующая роль теневых широколиственных лесов Поволжья будет негативной.
Перейдем к рассмотрению экологических последствий холодно-гумидного тренда, который ожидается моделью E GISS на период до 2050 г. и притом в сравнительно слабо выраженной тенденции (табл. 3). К данному прогнозному сроку в южной и типичной лесостепи (экорегионы Жигули и Присурье) произойдет небольшое понижение зимних и летних температур при существенном сокращении осадков, особенно в холодный период. Этот климатический тренд создаст свои зональные контрасты в биотической регуляции углеродного цикла. В Жигулевском массиве в подавляющем большинстве топоэкосистем проявится отрицательный углеродный баланс, правда, не столь феноменальный, как при термоаридном тренде по модели HadCM3 (табл. 4). Особенно пострадают мезогидроморфные теневые широколиственные леса средних и нижних частей крутых склонов (∆С(Fa) = –36.64 т/га). Среднее значение ∆С(Fa) в данном экорегионе составит –9.45 т/га. Это будет вызвано значительным снижением скелетной массы древостоев и кустарников (ср. ∆С(Fa) = –7.53 т/га), а также дополнительной эмиссией СО2 в результате активизации процессов разложения дебриса, гумуса и лесной подстилки. Следовательно, деградирующие маргинальные леса южной лесостепи будут осуществлять положительную регуляцию углеродного цикла, направленную на смягчение процесса похолодания. В подзоне типичной лесостепи при холодно-гумидном тренде можно ожидать уже отрицательную регуляцию углеродного цикла, направленную на усиление процесса похолодания. Ярким примером служит экорегион Присурье (табл. 4).
Смешанные леса более гумидного Приокско-Террасного заповедника обнаруживают большую пестроту в своих функциональных реакциях на прогнозируемый холодно-гумидный тренд (рис. 4б). В плакорных мезофильных сосново-липово-дубовых лесах произойдет значительная потеря углерода (более 33 т/га), и это будет вызвано, главным образом, ускоренным разложением лесной подстилки. Такой же, хотя и менее интенсивный, процесс охватит заболоченные хвойно-лиственные леса речных долин. Дополнительная эмиссия СО2 из этих лесных экосистем в атмосферу должна будет, по-видимому, смягчать общий холодный климатический тренд.
Две другие, более ксероморфные, группы биогеоценозов ПТЗ: водораздельные березово-сосновые и липово-березовые, с осиной, леса на песках и элювии карбонатных пород – должны отличаться почти столь же значительной отрицательной регуляцией углеродного цикла в период похолодания (∆С(Fa) ≈ 19.5–25.5 т/га). Здесь решающую роль сыграет, по-видимому, возрастающее накопление гумуса в почве по причине ослабления его минерализации в условиях более прохладных и сухих летних сезонов.
На 2150 г. (уже начавшегося потепления) по модели E GISS обрисовывается примерно та же картина локальных контрастов углеродного баланса, которую дает на тот же срок сценарная модель HadCM3. Это касается как Присурской типичной лесостепи, так и смешанных лесов Южного Верхневолжья.
Интересно сопоставить между собой, с одной стороны, углеродные балансы плакорных лесных экосистем, отображающих, как известно, в своих функционально-структурных параметрах зонально-региональный фон данной территории, а с другой – средневзвешенные для той же территории балансы (табл. 5). Разница впечатляет. В двух примерах из трех приведенных лесные сообщества плакорного ряда давали значения ∆Fa, резко отличные от обобщенных для данного района значений не только по величине, но и по знаку. В Присурье и ПТЗ плакорные леса должны осуществлять весьма существенную отрицательную регуляцию углеродного цикла, между тем как в целом по каждому их этих регионов прогнозируется положительная регуляция, правда довольно слабая. Экстразональные лесные сообщества, образующие целый спектр локальных отклонений от зонально-региональной нормы и занимающие более 70–75% площади, в обоих случаях дают результаты, с лихвой перекрывающие углеродный баланс плакорных лесов.
Таблица 5.
Плакорное (А) и средневзвешенное по площади (Б) базовое содержание органического углерода в почвенно-растительном покрове (т/га) и его изменения на особо охраняемых природных территориях Среднего и Верхнего Поволжья в соответствии с прогнозно-климатическими моделями
| а) Южная лесостепь. Национальный природный парк “Самарская Лука” (Жигули) | |||||
| Варианты осреднения | Базовый период | Модель HadCM3 | Модель E GISS | ||
| 2075 г. потепление | 2150 г. потепление | 2050 г. похолодание | 2200 г. потепление | ||
| А | 216.36 | –90.81 [─] (42.0) | –100.47 [─] (46.4) | –15.20 [+] (7.0) | –55.88 [─] (22.8) |
| Б | 193.7 | –73.87 [─] (38.1) | –82.60 [─] (42.6) | –10.81 [+] (5.6) | –44.23 [─] (22.8) |
| б) Типичная лесостепь. Национальный природный парк “Чаваш Вармане” (Присурье) | |||||
| Варианты осреднения | Базовый период | Модель HadCM3 | Модель E GISS | ||
| 2075 г. потепление | 2150 г. потепление | 2050 г. похолодание | 2200 г. потепление | ||
| А | 221.9 | –40.75 [─] (18.4) | –32.08 [─] (14.5) | –61.03 [+] (27.5) | –32.51 [─] (14.7) |
| Б | 167.4 | 24.3 [+] (14.5) | 34.75 [+] (20.8) | 16.14 [─] (9.6) | 37.41 [+] (22.3) |
| в) Южная подтайга. Приокско-Террасный биосферный заповедник | |||||
| Варианты осреднения | Базовый период | Модель HadCM3 | Модель E GISS | ||
| 2075 г. потепление | 2150 г. потепление | 2050 г. похолодание | 2200 г. потепление | ||
| А | 243.43 | –32.84 [─] (13.5) | –31.45 [─] (12.9) | –44.88 [+] (18.4) | –27.82 [─] (11.4) |
| Б | 200.49 | 9.02 [+] (4.5) | 10.65 [+] (5.3) | –9.52 [+] (4.7) | 6.16 [+] (3.1) |
Налицо убедительное доказательство неправомерности экстрополяции модельных сценариев биологического круговорота и углеродного баланса, которые получаются при имитационном моделировании “типичных биомов” (Пулы и потоки …, 2007; Formam, 1995) на всю территорию природной зоны/подзоны или крупного региона. Для каждого структурного подразделения той или иной зонально-региональной единицы необходимо вносить поправки, обусловленные местными закономерностями катенарной организации гео(эко)систем, и это может привести к совершенно иным результатам (Тишков, 2005), что и продемонстрировано проведенным нами опытом эмпирико-статистического моделирования.
ЗАКЛЮЧЕНИЕ
В изложенной авторской парадигме экспериментальной биогеоценологии управление операционной системой осуществляется с помощью эмпирико-статистических моделей пространственной организации лесных экосистем топологического уровня через свойства их пространственно-временно́го самоподобия. Полученный массив научно-методических разработок представлен таким образом, чтобы обогатить системный биогеоценологический анализ моделями базовых и прогнозируемых состояний топоэкосистем как целостных природно-территориальных образований, в духе учения В.Н. Сукачева о биогеоценозах. Можно надеяться, что результаты данного научного поиска будут способствовать развитию прогнозного направления в лесной биогеоценологии.
Список литературы
Анучин Н.П. Лесная таксация. М.: Лесная промышленность, 1982.
Арманд А.Д. Информационные модели природных комплексов. М.: Наука, 1975. 126 с.
Арманд А.Д. Общие представления // Экосистемы в критических состояниях. М.: Наука, 1989. С. 10–41.
Арманд Д.Л., Преображенский В.С., Арманд А.Д. Природные комплексы и современные методы их изучения // Изв. АН СССР. Сер. географ. 1969. № 5. С. 5–16.
Беручашвили Н.Л. Геофизика ландшафта. М.: Наука, 1990. 287 с.
Биологическая продуктивность лесов Поволжья / Ред. С.Э. Вомперский. М.: Hayкa, 1982. 282 c.
Варлыгин Д.Л., Базилевич Н.И. Связи продукции зональных растительных формаций Мира с некоторыми параметрами климата // Изв. РАН. Сер. географ. 1992. № 1. С. 23–32.
Васильев В.Н. Закономерности процесса смен растительности // Мат. по истории флоры и растительности СССР. М.–Л.: Изд-во АН СССР, 1946. Вып. II. С. 365–403.
Высоцкий Г.Н. Избранные труды. М.: Сельхозгиз, 1960. 435 с.
Герасимов И.П. Экологические проблемы в прошлой, настоящей и будущей географии мира. М.: Наука, 1985. 247 с.
Глазовская М.А. Геохимия природных и антропогенных ландшафтов СССР. М.: Высш. школа, 1988. 328 с.
Грин А.М., Ананьева Л.М., Куликов Ю.Н., Утехин В.Д. Сравнительные исследования структуры и функционирования природных и природно-технических экосистем // Структура, динамика и развитие ландшафтов. М.: Наука, 1980. С. 82–100.
Загреев В.В., Сухих В.И., Шывиденко А.З. и др. Общесоюзные нормативы для таксации лесов. М.: Колос, 1992. 495 с.
Залиханов М.Ч., Коломыц Э.Г., Шарая Л.С. и др. Высокогорная геоэкология в моделях. М.: Наука, 2010. 487 с.
Зейдис И.М., Кружалин В.И., Симонов Ю.Г. и др. Общие свойства динамики геосистем // Вестн. МГУ. Сер. 5. География. 2001. № 4. С. 3–8.
Злотин Р.И. Экологические проблемы биоты и устойчивость геосистем // Изв. АН СССР. Сер. географ. 1987. № 6. С.45–51.
Израэль Ю.А. Экология и контроль состояния природной среды. Л.: Гидрометеоиздат, 1979. 375 с.
Кобак К.И. Биотические компоненты углеродного цикла. Л.: Гидрометеоиздат, 1988. 247с.
Кобак К.И., Кондрашёва Н.Ю, Турчинович И.Е. Влияние изменений климата на природную зональность и экосистемы России // Изменения климата и их последствия. СПб.: Наука, 2002. С. 205–210.
Колесников Б.П. Кедровые леса Дальнего Востока / Тр. Дальневост. филиала АН СССР. Т. 2 (4). М.–Л.: Изд-во АН СССР, 1956. 263 с.
Коломыц Э.Г. Организация и устойчивость хвойнолесных экосистем на бореальном экотоне Русской равнины // Изв. РАН. Сер. географ. 1995. № 3. С. 37–51.
Коломыц Э.Г. Региональная модель глобальных изменений природной среды. М.: Наука, 2003. 371 с.
Коломыц Э.Г. Полизональность локальных геосистем как реакция на глобальные изменения климата // Изв. РАН. Серия геогр. 2006. № 2. С. 35–47.
Коломыц Э.Г. Локальные механизмы глобальных изменений природных экосистем. М.: Наука, 2008. 427 с.
Коломыц Э.Г. Лесные экосистемы Волжского бассейна в условиях глобального потепления // Экология. 2009. № 1. С. 3–15.
Коломыц Э.Г. Локальные коэффициенты увлажнения и их значение для экологических прогнозов // Изв. РАН. Серия геогр. 2010. № 4. С. 69–80.
Коломыц Э.Г., Сурова Н.А. Прогноз влияния глобального потепления на ресурсы почвенной влаги в маргинальных лесах Среднего Поволжья // Водные ресурсы. 2010. Т. 37. № 1. С. 89–101.
Коломыц Э.Г., Шарая Л.С. Прогнозное моделирование высокогорных экосистем в свете глобальных изменений климата // Экология. 2012. № 1. С. 3–15.
Крауклис А.А. Проблемы экспериментального ландшафтоведения. Новосибирск: Наука, 1979. 232 с.
Ляпунов А.А. О математическом моделировании в проблеме “Человек и биосфера” // Моделирование биогеоценотических процессов. М.: Наука, 1981. С. 5–29.
Методы изучения биологического круговорота в различных природных зонах / Ред. Н.И. Базилевич, А.А. Титлянова, В.В. Смирнов и др. М.: Наука, 1978. 183 с.
Молчанов А.А. Продуктивность органической массы в лесах различных зон. М.: Наука, 1971. 275 с.
Морозов Г.Ф. Учение о лесе. Изд. 7-е. М.–Л.: Гослесбумиздат, 1949. 368 с.
Одум Ю. Основы экологии. Пер. с англ. М.: Мир, 1975. 740 с.
Оценка и сохранение биоразнообразия лесного покрова в заповедниках Европейской России / Ред. Л.Б. Заугольнова. М.: Центр по проблемам экологии и продуктивности лесов РАН, 2000. 196 с.
Перельман А.И. Геохимия ландшафта. М.: Высшая школа, 1975. 341 с.
Программа и методика биогеоценологических исследований / Ред. Н.В. Дылис. М.: Наука, 1974. 403 с.
Пузаченко Ю.Г., Скулкин В.С. Структура растительности лесной зоны СССР: системный анализ. М.: Наука, 1981. 275 с.
Пулы и потоки углерода в наземных экосистемах России / Ред. Г.А. Заварзин. М.: Наука, 2007. 315 с.
Родин Л.Е., Базилевич Н.И. Динамика органического вещества и биологический круговорот зольных элементов и азота в основных типах растительности Земного Шара. М.–Л.: Наука, 1965. 253 с.
Розенберг Г.С. Модели в фитоценологии. М.: Наука, 1984. 265 с.
Розенберг Г.С., Мозговой Д.П., Гелашвили Д.Б. Экология. Элементы теоретических конструкций современной экологии. Самара: Самар. науч. центр РАН, 1999. 396 с.
Свирежев Ю.М., Логофет Д.О. Устойчивость биологических сообществ. М.: Наука, 1978. 352 с.
Симонов Ю.Г. Основные свойства объектов географического прогнозирования и способы их формализованного описания // Проблемы регионального географического прогноза. М.: Наука, 1982. С. 112–193.
Смирнов В.В. Органическая масса в некоторых лесных фитоценозах Европейской части СССР. М.: Наука, 1971. 362 с
Сочава В.Б. Экспериментальные географические исследования и освоение тайги // Информац. бюллетень по комплекс. освоению таежных территорий. Иркутск: Ин-т географии Сибири и Дальнего Востока СО АН СССР. 1969. Вып. 4. С. 5–14.
Сочава В.Б. Введение в учение о геосистемах. Новосибирск: Наука, 1978. 319 с.
Спрыгин И.И. Материалы к познанию растительности Среднего Поволжья // Научное наследство. Т. 11. М.: Наука, 1986. 512 с.
Сукачев В.Н. Избранные труды. Т. 1. Основы лесной типологии и биогеоценологии. Л.: Hayкa, 1972. 418 c.
Сукачев В.Н. Избранные труды. Том 3. Проблемы фитоценологии. Л.: Наука, 1975. 543 c.
Теоретические основы и опыт экологического мониторинга / Ред. В.Е. Соколов, Н.И. Базилевич. М.: Наука, 1983. 253 с.
Титлянова А.А. Методология и методы оценки чистой первичной продукции и построение баланса химических элементов в экосистемах // Теоретические основы и опыт экологического мониторинга. М.: Наука, 1983. С. 63–76.
Тишков А.А. Биосферные функции природных экосистем России. М.: Наука, 2005. 309 c.
Хелберт С.Х. Мнимые повторности и планирование экологических полевых экспериментов // Проблемы экологического эксперимента (планирование и анализ наблюдений). Тольятти: Ин-т экологии Волж. бассейна РАН, 2008а. С. 9–45.
Хелберт С.Х. Неверное истолкование мнимых повторностей и сопутствующие проблемы: ответ Л. Оксанену // Проблемы экологического эксперимента (планирование и анализ наблюдений). Тольятти: Ин-т экологии Волж. бассейна РАН, 2008б. С. 61–65.
Хорошев А.В. Полимасштабная организация географического ландшафта. М.: Изд-во КМК, 2016. 416 с.
Цельникер Ю.Л. Газообмен СО2 в лесных биогеоценозах / Идеи биогеоценологии в лесоведении и лесоразведении. М.: Наука, 2006. С. 213–229.
Швиденко А.З., Щепаченко Д.Г., Нильссон С., Болуй Ю.И. Таблицы и модели хода роста и продуктивности насаждений основных лесообразующих пород Северной Евразии (нормативно-справочные материалы). М.: Междунар. ин-т прикладного системного анализа, 2008. 885 с.
Шитиков В.К., Розенберг Г.С., Крамаренко С.С., Якимов В.Н. Современные подходы к статистическому анализу экспериментальных данных // Проблемы экологического эксперимента (планирование и анализ наблюдений). Тольятти: Ин-т экологии Волжского бассейна РАН, 2008. С. 212–250.
Экосистемы Теллермановского леса / Ред. В.В. Осипов. М.: Ин-т лесоведения РАН, 2004. 340 с.
Climate change 1995. The science of climatic change / Eds J.T. Houghton, L.G. Meira Filho, A. Callander et al. Cambridge, UK: Cambridge Univ. Press, 1996. 572 p.
Formam R.T.T. Land mosaics: the ecology of landscapes and regions. Cambridge: Cambridge Univ. Press, 1995. 605 p.
Emanuel W.R., Shugart H.H., Stevenson M.R. Climatic changes and the boreal-scale distribution of terrestrial ecosystem complexes // Climat. Change. 1985. №7. P. 29–43.
Gerrard A.J. Soil and lanforms. An integration of geomorphology and pedology. London–Boston–Sydney: George Allen & Unwin, 1981. 208 p.
Hansen J., Sato M., Ruedy R. et al. Climate simulations for 1880–2003 with GISS model E // Climate Dynamics. 2007. V. 29. P. 661–696.
Harbaugh J.W., Bonham-Carter G. Computer simulation in geology. New York–London–Sydney–Toronto: Willey-Interscience, 1970. 319 p.
Jeffers J.N.R. An introduction to systems analysis: with ecological applications. London: Edward Arnold, 1978. 198 p.
Leemans R. Modelling ecological and agricultural impacts of global change on a global scale // J. Sci. Indust. Res. 1992. V. 51. P. 709–724.
Lischke H., Guisan A., Fischlin A., Bugmann H. Vegetation response to climate change in the Alps: modeling studies // Views from the Alps: regional perspectives on climate change. Cambridge, Massachusetts, USA: MIT Press, 1998. P. 309–350.
Odum Eu. P. Fundamentals of ecology. Third edition. Philadelphia–London–Toronto: W.B. Saunders Company, 1971. 740 p.
Pianka E.R. Evolutionary ecology. Second edition. New York–Hagerstown–San Francisco–London: Harper and Row, Publishers, 1978. 399 p.
Pope V.D., Gallani M.L., Rowntree P.R., Stratton R.A. The impact of new physical parametrizations in Hadley Centre climate model – HadCM3 // Climate Dynamics. 2000. V. 16. P. 123–146.
Schmidt G.A., Ruegy R., Hansen J.E. et al. Present day atmospheric simulations using GISS Model E: comparison to in-situ, satellite and reanalysis data // J. Climate. 2006. V. 19. P. 153 –192.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии