

Успехи современной биологии, 2021, T. 141, № 4, стр. 390-398
Цифровой механизм приема-передачи акустической информации у птиц
Б. М. Звонов *
Институт проблем экологии и эволюции им. А.Н. Северцова РАН
Москва, Россия
* E-mail: Zvonovbm@gmail.com
Поступила в редакцию 08.01.2021
После доработки 22.01.2021
Принята к публикации 22.01.2021
Аннотация
Изучены пути приема и передачи акустического сигнала у птиц. Для этого проанализированы механизмы акустической изоляции у птиц, изучено построение сигналов бедствия и тревоги у различных видов птиц, рассмотрена изменчивость сигнала для обеспечения индивидуальности в границах видового сигнала. Выяснено, что все механизмы обеспечиваются временно́й дискретностью сигнала, который эволюционно исходит из одночастотного сигнала, что показано с помощью методики денервации сиринкса. Изучение нервной активности первичных слуховых ядер птиц показало, что слуховая система птиц адаптирована к анализу строгой временной дискретности акустических сигналов. Для подтверждения функциональности временной дискретности акустических сигналов в жизни птиц создана компьютерная программа цифрового электронно-акустического синтезатора репеллентных сигналов для управления поведением птиц, основанная на принципах построения естественных сигналов бедствия птиц. В результате стало возможным создание универсального сигнала для всех видов птиц, а также комбинаций сигналов и их случайных воспроизведений во времени, чтобы птицы не привыкали к синтезированному сигналу репеллента.
ВВЕДЕНИЕ
Использование акустической информации в жизни птиц так же необходимо как питание и размножение. Именно акустическая сигнализация обеспечивает птицам изоляцию при образовании пары, выкармливании птенцов и их воспитании, обеспечивая защиту своей территории от видовых соперников (Звонов, 2020). Акустическая сигнализация обеспечивает безопасность, когда птицы издают тревожные сигналы и сигналы бедствия не только для своего вида, но и для других видов.
Чтобы выяснить, какие способы передачи акустической информации используют птицы в своей жизнедеятельности, обратимся к созданию акустического изолирующего механизма во внутривидовом общении. Рекламирующая видовая песня птиц является маркером для фиксации территориального участка многих видов птиц. Ее значение настолько велико, что во весь период от откладки яиц парой до полного выращивания птенцов песня самца маркирует их территориальный участок и оберегает птиц своего вида от нежелательного проникновения соседей. Такое определяющее действие акустического сигнала заставляет исследовать механизм образования элементов сигнала, которые отличают песню данного самца от соседей, оставаясь при этом в пределах видового сигнала.
МАТЕРИАЛЫ И МЕТОДЫ
На рис. 1 приведены осциллограммы рекламирующих песен двух самцов-соседей пеночек-теньковок. Несложный подсчет длительности отдельного периода в сериях первой и второй особи показывает достоверную разницу у обоих самцов в течение всего репродуктивного периода, и она оказалась константой для каждой особи Т1 = 0.27 с, Т2 = 0.32 с. Разница в длительностях составляет всего 0.05 с, то есть 50 мс. И это является определяющим маркером отдельного самца при территориальной близости к другому самцу. В дальнейшем мы должны будем убедиться, способна или нет слуховая система птиц анализировать столь малые различия в длительности интервалов между импульсами.
Рис. 1.
Рекламирующие песни двух самцов пеночек-теньковок Phylloscopus collybitus. Верхняя последовательность – самец № 1, нижняя – самец № 2.

Далее обратимся к анализу временны́х различий в рекламирующих песнях других видов. Анализ песен двух особей соловьиной широкохвостки Cettia cetti (рис. 2), записанных с интервалом в четыре дня в течение 20 дней, показал полное совпадение как в частотных параметрах, так и во временны́х на протяжении всего срока наблюдения. Это говорит о стабильности физических параметров песни особи. Здесь же следует отметить, что в тревожных сигналах этих особей (рис. 2, 5, 10) временно́й рисунок меняется в сторону учащения и подобия, что говорит об универсальности этого сигнала как для видового, так и для межвидового общения птиц.
Рис. 2.
Видовые песни двух особей (а) и (б) соловьиной широкохвостки Cettia cetti, записанные с интервалом в 4 дня; 5, 10 – их тревожные сигналы.

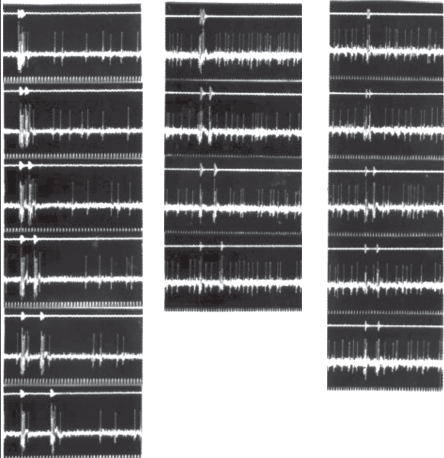
На рис. 3 приведены рекламирующие песни 10 особей широкохвостки, чтобы показать, как отдельные особи изменяют рисунок своей песни во временны́х параметрах, оставаясь узнаваемыми как вид.

Теперь обратимся к организации тревожных сигналов у различных видов птиц.
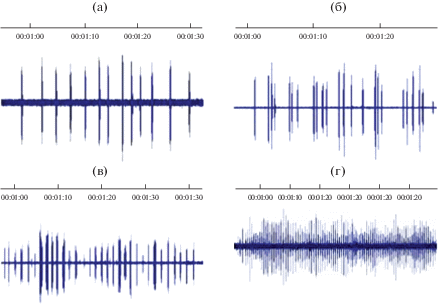
На рис. 4 приведены осциллограммы тревожных сигналов и сигналов бедствия птенцов из различных семейств. Привлекает внимание тот факт, что все сигналы построены по одному принципу – короткие, часто повторяющиеся импульсы, которые легко преобразовать в цифровой механизм. Именно такое построение сигнала является естественным для птенца и в то же время наиболее дальнобойным для привлечения родителей.
Рис. 4.
Осциллограммы сигналов тревоги и бедствия птенцов некоторых видов птиц. (а) – сигнал тревоги 10-дневного птенца полярной крачки Sterna paradisaea; (б) – слеток большого баклана Phalacrocorax carbo; (в) – 2-недельный птенец рыжей цапли Ardea purpurea; (г) – недельный птенец тундряной куропатки Lagopus mutus.
Необходимо выяснить, какую структуру имел первоначально акустический сигнал птиц, который затем преобразовался в ритмический, чтобы обеспечивать надежную дистанционную связь как на внутривидовом, так и межвидовом уровне. Рассмотрим акустическую коммуникацию речной крачки Sterna hirundo (рис. 5).
Рис. 5.
Сигнализация речной крачки в норме и после денервации сиринкса: а–д – индивидуальные различия призывного сигнала самок, е – сигнал самки при посадке к гнезду, ж – сигнал самца на противника, з – сигнал самки на самца с кормом, и – сигнал самки перед спариванием, к – сигнал самки после спаривания, л – призывный сигнал самки, м – призывный сигнал самки при денервации сиринкса справа, н – призывный сигнал самки при денервации сиринкса слева, о – призывный сигнал самки при двусторонней денервации сиринкса (по: Звонов, 1986).
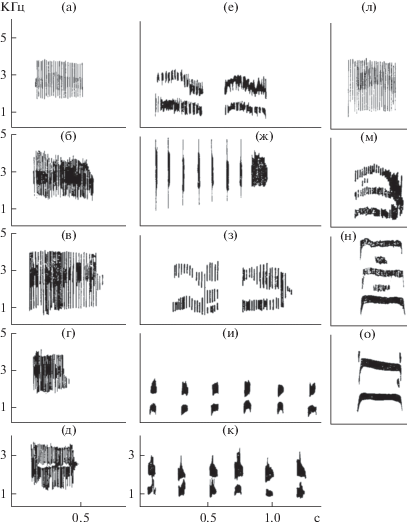
В левой колонке (а, б, в, г, д) приведены сонограммы призывных сигналов пяти особей речной крачки. Все сигналы построены на ритмической основе и индивидуальны. Были записаны коммуникативные сигналы одной самки и ее самца в различных ситуациях: е – сигнал самки при посадке к гнезду, ж – сигнал самца на противника, з – сигнал самки на самца с кормом, и – сигнал самки перед спариванием, к – сигнал самки после спаривания, л – призывный сигнал самки. Все многообразие сигналов построено на использовании различных временны́х построений, поддающихся цифровой интерпретации. Затем проводились исследования с использованием методики денервации сиринкса с целью выяснения влияния нервной регуляции на структуру сигнала. При двусторонней денервации (рис. 5о), когда нервная регуляция отсутствует, сигнал теряет свою временну́ю дискретность и предстает таким, каким он был первоначально. Именно на его основе в процессе эволюции строились сложные коммуникативные акустические сигналы (Звонов, 2011). Следует отметить, что, когда самка, сидя на гнезде, издавала призывный сигнал после денервации, самец ее не узнавал и игнорировал.
Теперь обратимся к возможностям слуховой системы птиц воспринимать и обрабатывать акустическую информацию. Мы проанализировали обработку акустической информации первичными слуховыми ядрами птиц – ангулярным (n. angularis), магноцеллюлярным (n. magnocellularis) и ламинарным (n. laminaris). Для этого применялась методика отведения нейронной активности отдельных нейронов первичных слуховых ядер. После отведения производился морфоконтроль для определения локализации кончика микроэлектрода в том или ином ядре.
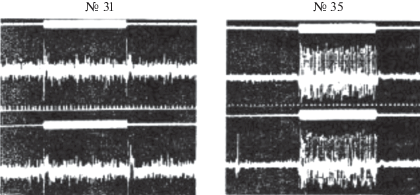
Прямым доказательством кодирования длительности отдельных элементов и слогов является наличие в первичных слуховых ядрах двух типов нейронов: тонических и фазических, способных с большой степенью точности кодировать информацию о длительности сигналов. Были найдены как фазические, так и тонические нейроны. На рис. 6 приведены фазический нейрон № 31 и тонический – № 35.
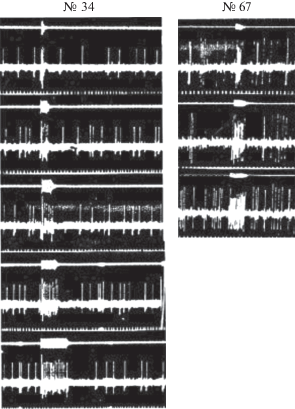
Точно так же тонические нейроны №№ 34 и 67 следят за изменением длительности стимула (рис. 7).
Не менее важно отметить “способность” нейронов первичных слуховых ядер птиц иметь быстрые циклы восстановления, чтобы следить за изменением длительности пауз между импульсами (рис. 8).
Это очень важно в тех случаях, когда индивидуальные различия определяются длительностью пауз между отдельными элементами и слогами песни.
До определенных частот (более 1000 Гц) отдельные нейроны разряжаются синхронно с частотой заполнения сигнала. Следовательно, при поступлении дискретного (ритмического) сигнала, состоящего из отдельных импульсов с частотой 1000 Гц и более нижние уровни слухового анализатора “способны” анализировать частотное разрешение длительностью 1 мс (Konishi, 1969). Именно такие сверхмалые интервалы между импульсами в сериях, например теньковок, воникают при образовании индивидуальных особенностей песен. Заметим, что слуховая система человека по этому показателю на порядок хуже.
Для подтверждения прямого воздействия импульсных сигналов с нужной временной структурой, был создан цифровой электронно-акустический синтезатор репеллентных сигналов птиц, на который был выдан патент (Звонов, Терсков, 2000).
За основу были взяты импульсные сигналы, из которых построены сигналы бедствия птиц (рис. 4). Именно импульсные сигналы с их крутыми фронтами, обеспечивают наиболее широкий частотный диапазон, так необходимый для обеспечения использования синтезатора для любых видов птиц.
Импульсная последовательность формируется следующим образом (рис. 9). Отдельная акустическая серия состоит из ряда импульсов, длительность которых изменяется от сеанса к сеансу в пределах наиболее эффективного репеллентного воздействия. По такому же принципу изменяется и интервал между импульсами. Количество импульсов в серии также изменяется для достижения максимального отпугивающего эффекта. Следующей, третьей переменной, является длительность серии. И, наконец, четвертая переменная – периодичность воспроизведения серий. Комбинация этих переменных в пределах наибольшего репеллентного эффекта обеспечивает устойчивое непривыкание к акустическому сигналу. За счет разработанной нами компьютерной программы (Электронно-акустический синтезатор репеллентных сигналов птиц) достигается создание нескольких сотен комбинаций, которые выбираются по случайному закону.

РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Только в совокупности изучения акустической сигнализации птиц и возможностей ее слуховой системы анализировать тонкости изменчивости сигнала в зависимости от жизненной необходимости можно понять акустическую коммуникацию птиц.
О значении временны́х параметров в передаче акустической информации имеются данные (Звонов, 1989, 2004, 2005, 2008; Jouventin, 1972; Bergmann, 1973; Lemon, 1974; Sossinka, Bohner, 1980) для многих видов птиц. Большинство из этих видов имеют несложную, компактную песню с ярко выраженными видовыми признаками. Наиболее сложен вопрос с видами, у которых невозможно выделить сколько-нибудь определенной территориальной видовой песни, и тем более с видами пересмешниками и имитаторами, включающими в свой репертуар заимствованные от других видов элементы. Так, у черной каменки не обнаружены песни, общие для всех особей даже одной локальной популяции (Костина, Панов, 1981). В большинстве случаев варианты, общие для двух или нескольких особей, являются имитациями сигналов других видов птиц и весьма просты по своей структуре. Однако авторам удалось установить, что выявленные различия между песнями рассмотренных трех форм данного вида опистолика (opistholeuca), капистрата (capistrata) и пиката (picata) затрагивают не структуру индивидуальных песен, а тип организации песенной последовательности. Такая организация видового сигнала, безусловно, является более сложной формой ритмической организации, и не исключено, что это одна из стадий эволюционного процесса, затрагивающего становление строгой структурной организации видового сигнала. Очевидно, при такой организации сигнала имеется большой простор для создания индивидуальных различий в ритмической организации. Найденная определенная песенная последовательность позволяет говорить и об определенной форме цифрового кода.
Имеющиеся работы по географической изменчивости песен птиц (Синежук, Кречмар, 2010; Podos, Warren, 2007) показывают, что подобная изменчивость носит дискретный характер и проявляется в форме диалектов.
А как же поступает в таком случае соловей, чья песня построена из сложных перепевов и слогов? Е.Н. Панов отмечает (1983), что песня соловья обладает явной и четкой ритмической структурой, которая держится на принципе комбинирования множества контрастирующих друг с другом звуков в разнообразные, но все же частично предсказуемые конструкции. Слагающие эту песню сигналы сами по себе не несут никакого смыслового значения. Для тех потребностей общения, которые обслуживает песня птиц, вполне достаточно, чтобы все самцы данного вида пользовались общей ритмической и частотной схемой пения, в то время как исходные элементы, воплощающие эту схему, могут быть достаточно различны у разных особей. Таким образом, даже в столь сложном сигнале, как песня соловья, можно выделить вполне определенную закономерность, основанную на ритмической структуре, которая и несет видовую и индивидуальную информацию. И как каждая ритмическая структура укладывается в определенный цифровой код.
Другой вопрос, зачем соловью так сложно строить свою песню, когда, казалось бы, все это можно сделать проще, как, например, в сигнализации коростеля (Грабовский, 1985), который достаточно просто строит свою акустическую коммуникацию, ограничиваясь односложными сигналами, но варьируя при этом длительностью секвенций. Соловей же обладает развитой нижней гортанью – сиринксом, который дает возможность создавать сложные частотно-модулированные звуки. Однако, так же как у коростеля, все они затем строятся в секвенции, представляющие собой один из вариантов ритмической организации. И такое положение оказывается вполне оправданным, так как имеющиеся на сегодняшний день работы (Tretzel, 1970; Guttinger, 1972; Winderle, 1979; Brenowitz, 1982) показывают, что при наследовании песни наиболее четко закреплена генетически именно их ритмическая структура.
Так как аналогичная картина проявляется у насекомых (Жантиев, Дубровин, 1974; Alexander, 1960), амфибий (Thompson, Martof, 1957; Littlejohn, Main, 1959; Littlejohn, Michaud, 1959; Blair, 1964), млекопитающих (Воронцов и др., 1969; Miller, 1944), хотелось бы считать это правило одним из основных правил биоакустической дивергенции у животных вообще, независимо от их систематической принадлежности.
Анализ полученных данных свидетельствует о большом разнообразии акустических сигналов, используемых птицами для целей дивергенции (смещение частотного диапазона, изменение временны́х характеристик отдельных элементов и слогов песни, использование амплитудной и частотной модуляции, изменение количественного соотношения слогов). Необходимо лишь заметить, что при всем разнообразии средств для создания акустического изолирующего механизма основными являются различия во временны́х параметрах построения песни или отдельных ее компонентов.
Определив, какую роль в кодировании акустической информации у птиц играют временны́е параметры сигнала, важно выяснить возможность обработки этих параметров на различных уровнях акустического анализатора у птиц.
Исследование периферического отдела слухового анализатора показывает (Konishi, 1970; Moller, 1972), что уже на уровне улитки осуществляется частотный анализ. Причем границы спектра частот, воспринимаемых улиткой, превосходят границы спектра частот, который данный вид использует в видовом общении. Такая возможность периферического отдела вполне оправдана, так как с одной стороны, вид должен воспринимать звуки своих соседей, которые могут нести информацию о возможной опасности, а с другой – слуховая система должна идти несколько впереди звукоиздающей системы в своем развитии, чтобы обеспечить возможность восприятия изменяющихся в эволюционном процессе звуковых сигналов своего вида.
Таким образом, мы видим, что функциональные характеристики слуховой системы как на уровне периферических, так и на уровне центральных ее отделов обнаруживают хорошее соответствие параметрам брачных сигналов, имеющим опознавательное значение.
Певчие птицы обучаются анализировать и издавать сигналы, использующиеся во внутривидовой коммуникации, а процессы обучения связаны с целостностью слуховых путей головного мозга. Пластичность обучения птиц отражается в аналитических процессах слуховых нейронов. И обучение слуховой системы направлено на строгую фиксацию как длительности отдельных посылок, так и интервалов между ними.
ЗАКЛЮЧЕНИЕ
Известно, что акустический сигнал, издаваемый птицей для передачи информации, имеет аналоговою форму. При этом его структура изменчива в зависимости от жизненной ситуации (тревога, охрана территории, связь с птенцом, коммуникация с партнером и т. д.). И эта изменчивость структуры определяется подвижностью временны́х параметров отдельных слогов сигнала и интервалов между ними за счет влияния нервной регуляции, что создает неограниченные возможности информационного многообразия. В то же время частотный диапазон сигнала определяется возможностями морфологических структур звукоиздающего аппарата птиц – у мелких воробьиных он располагается в более высокочастотном спектре, а крупных птиц – в низкочастотном. Такой аналоговый сигнал уже на первичном уровне обработки нейронными сетями слуховых ядер преобразуется в цифровую форму, т.к. разряды нейронов имеют импульсную структуру, и дальнейший ход обработки информации строится по цифровой форме в виде “0–1”.
Электронно-акустический синтезатор позволяет генерировать сигналы с крутыми фронтами, что обеспечивает (по закону Фурье) перекрытие всего спектра частот, к которым адаптирована слуховая система птиц – от 100 до 20000 гц. В-арьирование временными параметрами акустического сигнала создает множественность комбинаций и их случайное воспроизведение во времени, чтобы птицы не привыкали к синтезированному сигналу репеллента.
Список литературы
Воронцов Н.П., Ляпунова Е.А., Загоруйко Н.Г. Сравнительная кариология и становление изолирующих механизмов в роде Marmota // Зоол. журн. 1969. Т. 48. Вып. 3. С. 67–74.
Грабовский И.И. Акустическая сигнализация и коммуникация в локальном поселении коростелей // Зоол. журн. 1985. Т. 62. Вып. 2. С. 314–319.
Жантиев Р.Д., Дубровин Н.Н. Звуковые сигналы сверчков (Orthoptera, Oeacanthidae, Gryllidae) // Зоол. журн. 1974. Т. 53. Вып. 3. С. 91–98.
Звонов Б.М. Цифровой код в передаче акустической информации у птиц // Изв. АН СССР. Сер. биол. 1989. № 50. С. 50–57.
Звонов Б.М. Акустический образ чечевицы обыкновенной (Carpodacus erythrinus (Pall.) // Изв. РАН. Сер. биол. 2004. № 5. С. 561–565.
Звонов Б.М. Механизм передачи акустической информации у птиц // Изв. РАН. Сер. биол. 2005. № 2. С. 189–200.
Звонов Б.М. Акустический образ ремеза (Remiz pendulinus (L.) // Вестн. международной общественной академии экологической безопасности и природопользования (МОАЭБП). 2008. Вып. № 3 (10). С. 74–82.
Звонов Б.М. Пути развития акустических сигналов у птиц // Успехи соврем. биол. 2011. Т. 131. №3. 326–334.
Звонов Б.М. Механизм акустической изоляции у птиц на примере теньковки Phylloscopus collybitus // Рус. орнитол. журн. 2020. Т. 29. Экспресс-вып. 1996. С. 5263–5271.
Звонов Б.М., Терсков Н.Г. Способ синтезирования репеллентных сигналов животных и устройство для его осуществления. 2000. Патент № 2147178 от 10.04.2000 г.
Костина Г.Н., Панов Е.Н. Индивидуальная и географическая изменчивость песни у черной каменки O-enanthe picata // Зоол. журн. 1981. Т. 60. Вып. 9. С. 164–172.
Панов Е.H. Знаки, символы, языки. 2-е изд., доп. М.: Знание, 1983. 240 с.
Синежук Е.Б., Кречмар Е.А. Географическая изменчивость демонстративной песни обыкновенной чечевицы (Carpodacus erythrinus) // Зоол. журн. 2010. Т. 89. № 3. С. 1–14.
Alexander R.D. Sound communication in Ortoptera and C-icadidae // Animal sound and communication / Eds W.E. Lanyon, W.N. Tavolga. AIBS Bull. 1960. V. 7. P. 217–229.
Bergmann H-H. Die Imitationsleistung einer Mischsanger-Dorngras-Mucke (Sylvia communis) Ein Beitrag zum Problem Angeborener und Erworbener Gesangsmerkmale // J. Ornithol. 1973. V. 114. № 3. P. 317–332.
Blair W.F. Isolating mechanisms and interspecies interaction in Anuran amphibian // Quart. Rew. Biol. 1964. V. 39. P. 164–177.
Brenowitz E.A. The active space of red winged blackbird song // J. Comp. Physiol. 1982. V. 147. № 4. P. 511–529.
Guttinger H.R. Elementwahl und Strophenaufban in der Gesaugsentwicklung einiger Papageiamadinen-Arten (Gattung; Erythrura, Familie: Estrididae) // Z. Tierpsychol. 1972. V. 31. № 1. P. 68–86.
Jouventin P. Un nouveau systeme de reconnaisanse acoustique chez les oiseaux // Behaviour. 1972. V. 43. № 1–4. P. 176–191.
Konishi M. Time resolution by single auditory neurons in birds // Nature. 1969. V. 222. № 5. P. 193–202.
Konishi M. Hearing, single-unit analysis end vocalization in songbirds // Science. 1970. V. 166. P. 46–73.
Littlejohn M., Main A. Call structure in two genera of australian burrowing // Copeia. 1959. № 3. P. 86–91.
Littlejohn M., Michaud T. Mating call discrimination by females of Stretcher’s chorus frog Pseudacris streckeri // Texas J. Sci. 1959. V. 11. P. 301–320.
Lemon R.E. Song dialects song matching and species recognition by cardinals Richmondena cardinalis // Ibis. 1974. V. 116. № 4. P. 545–557.
Miller A.H. Specific differences in the notes chipmunks // J. Mammal. 1944. V. 25. P. 32–45.
Moller A.R. Coding of sounds in lower levels of the auditory system // Quart. Rev. Biophys. 1972. V. 5. № 1. P. 59–71.
Podos J., Warren P. The evolution of geographic vatiation in birdsong // Adv. Stud. Behav. 2007. V. 37. P. 403–458.
Sossinka R., Bohner J. Song types in the zebra finch Poepbila guttata castanotis // Z. Tierpsychol. 1980. V. 53. № 2. P. 123–132.
Tretzel S. Akustisches. Lerner und Rhythmusangleichung bei Schamadrosseln (Copsychus malabaricus Scop.) // Zool. Anz.-Supplementad. 1970. V. 33. P. 290–302.
Thompson E., Martof B. A comparison of the physical characteristics of frog calls (Pseudacris) // Physiol. Zool. 1957. V. 30. P. 67–81.
Winderle J.M. Components of song used for species recognition in the common yellowthroat // Anim. Behav. 1979. V. 27. № 4. P. 982–989.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии