

Успехи современной биологии, 2021, T. 141, № 5, стр. 443-456
Роль селена в жизнедеятельности растений, животных и человека
А. Ф. Титов 1, Н. М. Казнина 1, *, Т. А. Карапетян 2, Н. В. Доршакова 2, В. Н. Тарасова 2
1 Институт биологии – обособленное подразделение Федерального исследовательского
центра “Карельский научный центр РАН”
Петрозаводск, Россия
2 Петрозаводский государственный университет
Петрозаводск, Россия
* E-mail: kaznina@krc.karelia.ru
Поступила в редакцию 21.01.2021
После доработки 16.03.2021
Принята к публикации 19.03.2021
Аннотация
Представлен обзор литературных данных о роли селена в жизнедеятельности растений, животных и человека. Приведены концентрации селена в разных по составу почвах и в почвах разных регионов России. Рассмотрен вопрос о содержании селена в растениях как основном источнике его поступления в организм человека. Отмечено положительное влияние небольших доз селена на продуктивность и качество урожая ряда сельскохозяйственных культур. Указаны возможные причины дефицита селена в растениях. Отдельный раздел посвящен роли селена в жизнедеятельности животных. Показано его участие в функционировании целого ряда ферментов и белков, описаны некоторые заболевания у животных при дефиците этого микроэлемента. Особое внимание уделено влиянию селена на здоровье человека. Рассматриваются основные функции селена в организме человека, в том числе его канцеропротекторное действие. Суммированы данные о селенсодержащих белках с указанием их участия в конкретных биологических процессах. Подробно описаны заболевания, вызываемые дефицитом селена. Указаны возможные пути устранения дефицита этого элемента у человека. Названы вопросы, связанные с исследованием влияния селена на живые организмы, которые требуют изучения в ближайшее время.
ВВЕДЕНИЕ
Селен является микроэлементом, который в малых концентрациях необходим для всех живых организмов, включая человека. Несмотря на то, что в организме человека его содержание невелико, селен непосредственно участвует в выполнении большого числа жизненно важных функций в клетке. В частности, он необходим для тканевого дыхания, окислительного фосфорилирования, функционирования пентозофосфатного цикла и цикла трикарбоновых кислот, участвует в гормональном балансе щитовидной железы, а также в работе антиоксидантной системы организма (Bianco et al., 2002; Duntas, Benvenga, 2015; Preda et al., 2017). Поэтому дефицит селена отрицательно сказывается на здоровье человека. Например, обнаружено, что при недостатке этого микроэлемента увеличивается восприимчивость людей к инфекциям, в том числе к вирусу СПИД (Rayman, 2012), снижается продолжительность жизни. Дефицит селена обусловливает развитие ряда эндемических заболеваний (болезнь Кешана и Кашина–Бека), ускоряет развитие атеросклеротических процессов, повышает вероятность развития сердечно-сосудистых заболеваний и инсультов, приводит к развитию катаракты и многим формам рака, бесплодию у мужчин, а также к задержке роста у детей (Бабенко, 2001; Болотников, 2008).
По мнению ряда авторов, физиологический оптимум (диапазон безопасных поступлений) селена для человека составляет 350–400 мкг/день (Levander, 1991). Дефицит микроэлемента у взрослого человека отмечается при его поступлении в дозе 16–21 мкг/день и ниже (Yang et al., 1989). Согласно последним данным, в настоящее время дефицит селена выявлен у 1 млрд людей на земном шаре, включая и Россию (по: Аристархов и др., 2018). Основной причиной его возникновения является недостаточное поступление микроэлемента с продуктами питания, что в свою очередь обусловлено его низким содержанием в окружающей среде (Ermakov, 2001).
Одной из наиболее эффективных мер по устранению дефицита селена в организме человека считается использование биологически активных добавок к пище, однако чаще всего это не решает проблему недостатка микроэлемента в полном объеме. Более успешным решением может стать увеличение содержания селена в продуктах питания, особенно растительного происхождения, поскольку именно растения являются основными источниками этого микроэлемента и продуцентами, находящимися в начале пищевой цепи. Кроме того, в растениях он находится в форме метилированного селена (селенметионина), доступность которого в 5–10 раз выше, чем его доступность из неорганических соединений (Кашин, Шубина, 2011).
В настоящее время имеется довольно большое количество исследований, касающихся роли селена в жизнедеятельности растений, животных и человека, результаты которых нашли свое отражение в экспериментальных статьях (Серегина, 2007; Голубкина и др., 2017; Lubos et al., 2010; Misra et al., 2015 и др.), обзорах (Finley, 2005; Fajt et al., 2009; Mangiapane et al., 2014 и др.) и монографиях (Ермаков, Ковальский, 1974; Тутельян и др., 2002; Голубкина, Папазян, 2006; Вихрева и др., 2012 и др.). Тем не менее, интерес к этому микроэлементу не ослабевает, и исследования влияния дефицита селена на живые организмы продолжаются. Однако в основном эти работы отражают отрицательное влияние недостатка этого микроэлемента на организм человека. В настоящем обзоре предпринята попытка обобщения данных литературы, касающихся роли селена в жизнедеятельности растений, животных и человека.
СОДЕРЖАНИЕ СЕЛЕНА В ПОЧВЕ
В почве содержание селена колеблется в очень широких пределах и зависит главным образом от материнской породы, на которой сформированы почвы, от вида почвы и ее физико-химического состава (Ермаков, Ковальский, 1974). Повышенный уровень селена (20 мг/кг и выше) встречается довольно редко, и он характерен для почв, сформированных на материнских породах, богатых этим микроэлементом, а также для вторично засоленных почв. Наиболее широко они распространены в США, Канаде, Южной Америке, Австралии, Индии и Китае (White, 2016). Достаточный для растений уровень селена (200–300 мкг/кг) имеют лишь типичные черноземы (Голубкина и др., 2012). Большинство же почв характеризуются дефицитом этого микроэлемента (менее 100 мкг/кг) (Иванов, 1996). Например, в России в той или иной мере выраженный дефицит селена встречается в почвах 27 регионов, включая республики Бурятия, Карелия, Коми, Марий Эл, Мордовия, а также Архангельскую, Вологодскую, Ивановскую, Иркутскую, Ленинградскую, Московскую, Мурманскую, Новгородскую, Тверскую, Читинскую и Ярославскую области. Наибольший дефицит микроэлемента выявлен в Бурятии, Читинской области и Хабаровском крае (Голубкина, Папазян, 2006). Для таких территорий характерны в основном подзолистые, дерново-подзолистые или болотные почвы (Голубкина и др., 2017).
В целом, общее содержание селена в почве складывается из его неорганических соединений и органических форм, попадающих в почву вместе с умершими растениями и животными организмами. Важным источником поступления селена в почву является и антропогенный фактор, в частности, внесение селенсодержащих удобрений (Шеуджен и др., 2018) или сжигание ископаемого топлива (Сидельникова, 1999).
СЕЛЕН В РАСТЕНИЯХ
Растения представляют собой первое звено в пищевой цепи переноса селена в организм человека и его основной источник. Вместе с тем для растений селен является одним из наиболее труднодоступных химических элементов. Его низкая доступность связана с малой подвижностью и плохой растворимостью некоторых из его солей, в частности, селенитов и сульфидов селена, часто встречающихся в разных типах почв (Li et al., 2008). Лучше всего растениями поглощаются селенаты, хорошо растворимые в воде. Поглощение селенатов клетками корня из ризосферы осуществляется белками-транспортерами серы, гомологичными белкам арабидопсиса AtSULTR1.1 и AtSULTR1.2 (Sulfate transporter 1.1 и 1.2) (White, 2016).
Поглощение селена растениями зависит и от физико-химических свойств почвы (pH почвы, содержание солей, органических веществ и др.), а также от видовых (сортовых) особенностей (Ермаков, Ковальский, 1974; Вихрева, Лебедева, 2010; Шеуджен и др., 2013). По способности накапливать селен растения можно разделить на три группы (Ермаков, Ковальский, 1974). Первая – наиболее многочисленная, включает растения, которые накапливают этот элемент в небольших количествах (0.1–1.0 мг/кг сухой массы), гораздо меньших, чем его содержится в почве. Растения второй группы характеризуются способностью к накоплению селена в количествах, превышающих его концентрацию в почве. При этом содержание элемента в растениях может достигать 200 мг/кг сухой массы. К третьей группе относятся растения, способные к сверхнакоплению селена. Это так называемые гипераккумуляторы, содержание селена в которых может достигать 1000–15 000 мг/кг сухой массы. К ним, в частности, относятся представители родов Astragalus (сем. Fabaceae), Stanleya (сем. Brassicaceae), Oonopsis (сем. Asteraceae) и Xylorhiza (сем. Asteraceae). Внутри указанных трех групп содержание селена также может значительно различаться. Так, все сельскохозяйственные культуры относятся к первой группе. Однако, по данным литературы, наибольшее количество селена накапливают растения сем. Brassicaceae (в том числе, брокколи, белокочанная капуста, цветная капуста и капуста кольраби) (Wiesner-Reinhold et al., 2017) и семейства Poaceae (в частности, хлебные злаки). Из злаков ведущую позицию в качестве источника селена занимает мягкая пшеница. Продукты ее переработки обеспечивают около 50% всего селена, потребляемого жителями России. Обнаружены также внутривидовые колебания содержания селена. Например, в зерне разных сортов мягкой пшеницы содержание селена может варьировать от менее 60 до 880 мкг/г сухой массы (Голубкина, Папазян, 2006).
В растениях селен находится в виде селенатов, селенитов, аналогов серосодержащих аминокислот и селенопептидов. В транспорте этих соединений по растению участвует ряд транспортных белков, среди которых АВС-транспортеры (Byrne et al., 2010) и транспортеры серы, гомологичные AtSULTR2.1, AtSULTR2.2, AtSULTR3.5, AtSULTR4.1 и AtSULTR4.2 (White, 2016). Обладая способностью замещать серу, селен образует Se-содержащие аминокислоты (селеноцистеин SeCys и селенометионин SeMet) и белки (Голубкина и др., 2017). Обнаружено, что метилированные формы Se-содержащих аминокислот – селенометионин (SeMet), селенометилселеноцистеин (SeMe-SeCys) и дипептид гамма-глутамилселенометилселеноцистеин (γ-Glu-SeMe-SeCys) – обладают антиканцерогенным действием, что дополнительно повышает интерес к исследованиям, направленным на поиск путей увеличения содержания селена в растениях (Голубкина, Папазян, 2006; Zeng, Combs, 2008). Включение селена в метаболизм высших растений происходит тем же путем, что и включение серы (Zayed et al., 1998), поэтому важным определяющим фактором влияния селена на белковый обмен растительного организма является неспецифическое включение селена вместо серы в селеносодержащие аминокислоты (Stadlober et al., 2001; Whanger, 2002). Метаболизм селена в клетке растений, не являющихся гипераккумуляторами, представлен на рис. 1.
Рис. 1.
Метаболизм селена в растениях, не являющихся его гипераккумуляторами (по: van Hoewyk, 2013, с модификациями). GSH – глутатион; GS-Se-SG – селенодиглутатион.
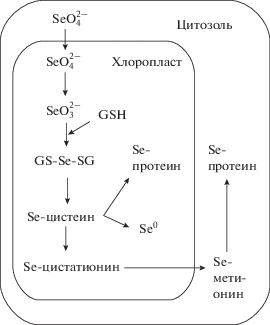
Необходимость селена для растений в настоящее время изучается. Например, обнаружено его участие в реакциях образования хлорофилла, синтеза трикарбоновых кислот, в метаболизме длинноцепочечных жирных кислот (Whanger, 2002). Однако одной из наиболее важных функций селена в растительном организме, по мнению многих авторов, считается его способность активизировать антиоксидантную защиту клеток за счет усиления активности целого ряда антиоксидантных ферментов, в том числе, супероксиддисмутазы, каталазы и глутатионпероксидазы, а также синтеза не ферментных антиоксидантов, таких как аскорбат, флавоноиды, алкалоиды и каротиноиды (Feng et al., 2013). Помимо этого, селен в форме селеноцистеина входит в состав активных центров одного из важнейших антиоксидантных ферментов – Se-зависимой глютатионпероксидазы (Аристархов и др., 2018). Полагают, что именно с участием селена в регуляции активности компонентов антиоксидантной системы связана его важная роль в защите растений от воздействия стресс-факторов (Блинохватов и др., 2001; Yao et al., 2011b; Durán et al., 2015).
Нетрудно предположить, что недостаток селена в почве должен отрицательно сказываться на жизнедеятельности растений, однако пока такого рода данных в литературе крайне мало. Известно лишь, что основным внешним признаком дефицита селена у растений является появление больших белых штрихов на листьях. Обнаружена также задержка развития растений при недостатке этого микроэлемента в почве. Большинство же из известных работ касается изучения ответной реакции растений на внесение селена в почву в тех или иных концентрациях. При этом доказано положительное влияние небольших доз микроэлемента на продуктивность и качество урожая целого ряда сельскохозяйственных культур. Например, у пшеницы добавление малых концентраций селенита натрия в субстрат приводило к увеличению урожая семян, а также вызывало повышение содержания протеина в вегетативных и репродуктивных органах (Кашин, Шубина, 2011). У обработанного селеном картофеля увеличивался урожай клубней (Прудников, 2007).
На настоящий момент механизмы стимулирующего эффекта селена на растения неизвестны, высказан лишь ряд предположений на этот счет. Так, некоторые авторы считают, что в основе этого лежит участие селена в азотном обмене (Серегина, Ниловская, 2002). Повышение же содержания азотистых соединений, обладающих гидрофильными свойствами и более высокой водоудерживающей способностью, по-видимому, может являться основным фактором, определяющим положительное влияние селена на водный обмен растений (Кашин, Шубина, 2011). На картофеле показано, что растения, обработанные низкими дозами селенита натрия, характеризовались более интенсивным дыханием, большей скоростью фотохимических реакций в хлоропластах, увеличением чистой продуктивности фотосинтеза, что обеспечивало их более интенсивный рост и большую продуктивность по сравнению с контрольными растениями (Прудников, 2007). У пшеницы разные виды обработки селенитом натрия (предпосевная, внекорневая и комплексная) приводили к повышению содержания фотосинтетических пигментов и увеличению сухой биомассы побега (Кулагина, Головацкая, 2011). Обнаружено также положительное влияние селена на скорость нетто-фотосинтеза, что, отчасти, может быть связано с повышением фотохимической активности хлоропластов (Прудников, 2007)
Выявлен также положительный эффект малых доз селена на устойчивость растений к стрессовым воздействиям. Например, при обработке растений козлятника восточного (Galega orientalis) селеном усиливался синтеза пролина, что увеличивало устойчивость растений к засолению и тепловому стрессу (Вихрева, 2001). Растения картофеля в присутствии селена оказались более устойчивы к низким (–2°С) температурам (Прудников, 2007), что, по мнению автора, связано в первую очередь с изменением гормонального баланса, в частности, с увеличением содержания ауксинов и гиббереллинов, а также снижением интенсивности процессов перекисного окисления липидов (ПОЛ), вызванных действием низкой температуры. Кроме того, обработка семян ряда сельскохозяйственных культур водными растворами селената натрия в концентрации 10–3–10–5% оказывала положительное воздействие на всхожесть и начальный рост проростков в стрессовых условиях, например: козлятника – при водном дефиците, засолении, гипертермии и гипоксии; ячменя – при фунгицидной обработке семян; пшеницы – при повышении кислотности среды (Вихрева, 2011). Это, по мнению автора, во многом обеспечивается способностью микроэлемента усиливать протеолиз за счет усиления активности протеолитических ферментов и увеличивать антиоксидантный потенциал растений.
В целом, можно заключить, что селен в малых дозах положительно влияет на жизнедеятельность растений, однако его роль в регуляции основных физиологических процессов у растений до сих пор изучена крайне слабо, а в вопросе о механизмах его влияния на растения существуют только предположения. Практически нет данных и о влиянии дефицита этого элемента на растения.
РОЛЬ СЕЛЕНА В ЖИЗНЕДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ
Роль селена для животных в основном изучена на одомашненных видах, что связано с большей практической значимостью этого вопроса. Исходя из данных литературы, потребности животных в этом микроэлементе различаются в зависимости от вида, пола, возраста и физиологического состояния функциональных систем организма (Stec et al., 2005). Для селена главным путем поступления в организм является алиментарный, при этом 90% микроэлемента поступает с пищей, преимущественно с растительной, в виде селеноцистеина (SeCys) и селенометионина (SeMet) и 10% – с водой (Степанов и др., 2012). Всасывание селена наиболее интенсивно происходит в двенад-цатиперстной кишке и в несколько меньшей степени – в тощей и подвздошной кишках, а выведение осуществляется в основном с мочой и калом или с выдыхаемым воздухом (Авцын и др., 1991). Высокие концентрации селена отмечены в печени (30% от общего содержания в организме), мышцах (30%) и почках (15%). Печень и почки – это так называемые депо селена для поддержания синтеза селенопротеинов при недостатке этого микроэлемента (Schrauzer, 2000).
Важная роль селена для животных во многом связана с тем, что он входит в состав активного центра глутатионпероксидазы в виде селеноцистеина (Holovská et al., 2003). Селеноцистеин – аналог цистеина, у которого серосодержащая тиольная группа заменена на селеносодержащую. Он имеет более низкую, чем у цистеина, константу диссоциации и более высокий восстановительный потенциал, благодаря чему селеноцистеин задействован в белках, обладающих антиоксидантной активностью (Byun, Kang, 2011). Глутатионпероксидаза – первый селеносодержащий фермент, обнаруженный в организме млекопитающих – участвует в детоксикации избыточных количеств продуктов ПОЛ и обеспечивает защиту клеток от окислительного стресса (Зайцев, Конопатов, 2004). Поскольку в составе глутатионпероксидазы находится 30–40% общего содержания селена, уровень ее активности в крови часто используется как показатель содержания этого микроэлемента у животных (Holovská et al., 2003).
Помимо глутатионпероксидазы, в организме животных имеется большое количество селенсодержащих ферментов. Среди них: глицинредуктаза, моноселенофосфат-синтетаза (катализирует синтез селенофосфата из селенида и АМФ), селеноцистеинсинтетаза, тиоредоксинредуктазы 1, 2 и 3 (катализируют NADPH-зависимое восстановление окисленного тиоредоксина в цитозоле и митохондриях) (Касумов, 1979; Зайцев, Конопатов, 2004). Селенсодержащие ферменты играют важную роль в контроле клеточного деления, апоптозе, процессах детоксикации (Кармолиев, 2005). Селен также входит в состав селенопротеинов.
В силу большой функциональной значимости селена, его дефицит приводит к различным нарушениям метаболизма, в том числе связанным с нарушениями клеточных мембран. В этом плане наиболее изучен недостаток одного из селенопротеинов – селенопротеина W, с дефицитом которого связывается развитие алиментарной мышечной дистрофии или беломышечной болезни (БМБ), которая наиболее остро проявляется у молодых особей (Radostits et al., 2000; Streeter et al., 2012). Свое название заболевание получило из-за специфического белого цвета мышц. БМБ развивается при содержании селена менее 0.1 мг/кг сухого вещества корма и имеет преимущественно очаговый, энзоотический характер. Предполагают, что это заболевание усугубляется недостатком витамина Е (Усольцева и др., 2010). В патологический процесс вовлекается миокард и поперечнополосатая мускулатура переднего и заднего пояса, реже поражаются жевательные мышцы и диафрагма. Как отмечено у ряда видов сельскохозяйственных животных, БМБ начинает развиваться вскоре после рождения – в первые две недели и последующие два–три месяца жизни. Клинические симптомы включают скованность мышц, слабость и малоподвижность (Radostits et al., 2000), изменение частоты и ритма сердцебиения (Żarczyńska et al., 2013). Так, на ЭКГ больных телят была обнаружена повышенная частота сердечных сокращений, увеличивалась амплитуда зубца Р, укорачивались интервалы PR, QT и ST (Żarczyńska et al., 2013). При развитии БМБ смертность среди молодняка может достигать 60%. Дефицит селенопротеина W возможен и у взрослых особей, но, как правило, болезнь у них протекает в скрытой форме.
У молодых животных, в частности, у ягнят и телят, при дефиците селена наблюдается дистрофия миокарда, что зачастую приводит к их гибели, у поросят отмечена алиментарная дистрофия, гепатоз, язва желудка, у жеребят – полимиозит и азотурия (Koller, Exon, 1986).
Дефицит селена и витамина Е также может быть причиной стеатита, при котором происходит дегенерация и замещение жировой ткани соединительной с отложениями кальция, что, например, было обнаружено у лошадей и ослов (Cardona, Reza, 2011; van Loon et al., 2015). В результате у животных отмечаются малоподвижность, снижение аппетита, потеря веса, лихорадка, вентральный отек и пр. У свиней дефицит селена и витамина Е вызывает так называемый синдром VESD (vitamin E/selenium deficit) (Fajt et al., 2009). Селеновая недостаточность под названием “энзоотический гепатит поросят” была впервые обнаружена и описана Е.М. Земмером еще в 1882 г.
Необходимо указать и на важную роль селена в фертильности, имплантации эмбриона, удержании плаценты, синтезе тестостерона и сперматозоидов и их подвижности. Как результат, дефицит этого микроэлемента отрицательно влияет на репродуктивные параметры и продуктивность животных. В частности, отмечено, что на селено-дефицитных территориях чаще регистрируются случаи бесплодия у коров, тогда как добавление селена в растительный корм увеличивает их плодовитость (Aréchiga, Vázquez-Flores, 1998). Кроме того, у крупного рогатого скота дефицит селена приводит к абортам (Kamada et al., 2014) и мертворождениям (Uematsu et al., 2016). Предполагают, что наиболее вероятными причинами этого являются возникающая недостаточная концентрация прогестерона, необходимая для нормального протекания беременности (Kamada et al., 2014), и формирующаяся сердечная недостаточность у плода (Underwood, Suttle, 2004).
Обнаружено также, что селен стимулирует синтез антител, активность Т-хелперов, миграцию фагоцитарных клеток и фагоцитоз (Finch, Turner, 1996). Некоторые метаболиты селена и селенопротеины (например, глутатионпероксидаза и тиоредоксинредуктаза), участвуют в иммунных и воспалительных реакциях, в антиоксидантном ответе клетки (Sordillo, 2013). Дефицит селена приводит к ослаблению иммунной защиты, обусловливает воспалительные реакции и иммунодефициты. Так, известно, что у коров при дефиците селена снижается способность нейтрофилов крови и молока нейтрализовывать патогены, что способствует развитию маститов (Sordillo, 2013; Mangiapane et al., 2014).
Таким образом, селен является элементом, который участвует во многих жизненно важных процессах в организме, влияет на метаболизм и репродуктивные свойства животных. Его дефицит приводит к развитию нарушений практически во всех органах и системах и вызывает ряд специфических заболеваний, симптоматика которых ослабевает или полностью исчезает при добавлении селена в рацион питания. Вместе с тем до настоящего времени не в полной мере изучен механизм развития болезней, связанных с недостатком селена у животных, не определены методы их дифференциальной диагностики, а также наиболее эффективные методы и формы введения селена в организм животных. Дальнейшее изучение роли селена в отдельных процессах у животных представляется весьма важным, поскольку открывает возможности для профилактического добавления этого микроэлемента с целью исключения развития патологий, обусловленных его дефицитом.
ВЛИЯНИЕ ДЕФИЦИТА СЕЛЕНА НА ОРГАНИЗМ ЧЕЛОВЕКА
Для человека главным источником селена является пища, в основном растительного происхождения, поэтому его содержание в организме во многом определяется рационом питания. “Нормы физиологических потребностей в энергии и пищевых веществах для различных групп населения Российской Федерации” определяют потребление селена для мужчин и женщин в возрасте от 18 лет и старше в 70 и 55 мкг/сут соответственно. Однако, как правило, обычный пищевой рацион человека не может обеспечить необходимую суточную потребность в этом микроэлементе. Поэтому дефицит селена испытывает довольно большой процент населения земного шара, в том числе около 80% жителей России (Тутельян и др., 2002; Drytel et al., 2013).
Отрицательное воздействие дефицита селена на организм человека объясняется большим числом разнообразных функций, которые он выполняет в клетке. В первую очередь они связаны с функционированием селенсодержащих белков: оксидоредуктаз (дейодиназ), глутатионпероксидаз, тиоредоксинредуктаз и селенопротеинов, роль и значение которых чрезвычайно многообразны, хотя до сих пор до конца не известны (табл. 1).
Таблица 1.
Селеносодержащие белки и их функции (по: Трошина и др., 2018; Ibrahim et al, 2019)
| Белок | Символ | Uniport | Функция |
|---|---|---|---|
| Глутатионпероксидаза 1 | GPX1 | P07203 | Участвует в детоксикации избытка перекиси водорода, обеспечивая сохранение про-анти-оксидантного равновесия в клетке |
| Глутатионпероксидаза 2 | GPX2 | P18283 | Участвует в апоптозе и клеточной пролиферации. Уменьшает пероксидацию в кишечнике |
| Глутатионпероксидаза 3 | GPX3 | P22352 | Уменьшает пероксидацию в крови. Участвует в защите сердечно-сосудистой системы, возможно, посредством регуляции уровня оксида азота |
| Глутатионпероксидаза 4 | GPX4 | P36969 | Участвует в антиоксидантной защите клеточных мембран. Вовлечена в процесс апоптоза, ингибирования липоксигеназ. Необходима для нормального функционирования сперматозоидов |
| Глутатионпероксидаза 5 | GPX5 | O075715 | Предположительно выполняет резервную функцию для других глутатионпероксидаз в сперме |
| Глутатионпероксидаза 6 | GPX6 | P59796 | Функция неизвестна. Предположительно участвует в обонянии |
| Глутатионпероксидаза 7 | GPX7 | Q96SL4 | Выполняет протекторную роль при раке молочной железы |
| Йодтирониндейодиназа 1 | DIO1 | P49895 | Катализирует конверсию тироксина в трийодтиронин преимущественно в щитовидной железе |
| Йодтирониндейодиназа 2 | DIO2 | Q92813 | Катализирует конверсию тироксина в трийодтиронин преимущественно вне щитовидной железы (в сердце, центральной нервной системе, гипофизе, скелетной мускулатуре, бурой жировой ткани и плаценте) |
| Йодтирониндейодиназа 3 | DIO3 | P55073 | Инактивирует тиреоидные гормоны: конверсия тироксина в реверсивный трийодтиронин и три-йодтиронина в дийодтиронин |
| Тиоредоксинредуктаза 1 | TXNRD1 | Q16881 | Уменьшает уровень цитозольного тиоредоксина, участвуя в регулировании некоторых факторов
транскрипции (NF-κВ, Ref-1, P53) и экспрессии генов, репарации ДНК, клеточном сигналинге,
апоптозе и клеточной пролиферации. Участвует в антиоксидантных процессах |
| Тиоредоксинредуктаза 2 | TXNRD2 | Q9NNW7 | Уменьшает уровень митохондриального тиоредоксина |
| Тиоредоксин-глутатионредуктаза | TXNRD3 | Q86VQ6 | Уменьшает уровень тиоредоксина. Участвует в восстановлении глутатиона |
| Селенопротеин F | SELENOF | O60613 | Возможно, участвует в фолдинге белка |
| Селенопротеин Н | SELENOH | Q8IZQ5 | Возможно, участвует в транскрипции некоторых белков. Участвует в антиоксидантной защите клеток |
| Селенопротеин I | SELENOI | Q9C0D9 | Возможно, участвует в биосинтезе фосфолипидов |
| Селенопротеин К | SELENOK | Q9Y6D0 | Участвует в кальциевом гомеостазе в иммунных клетках. Выступает как антиоксидант преимущественно в кардиомиоцитах |
| Селенопротеин М | SELENOM | Q8WWX9 | Возможно, участвует в фолдинге белка. Необходим для работы головного мозга. Низкий уровень белка определяется при болезни Альцгеймера |
| Селенопротеин N | SELENON | Q9NZV5 | Участвует в функционировании мышечной ткани |
| Селенопротеин О | SELENOO | Q9BVL4 | Возможно, участвует в регуляции окислительных процессов |
| Селенопротеин Р | SELENOP | P49908 | Основное внеклеточное депо селена. Секретируется в плазму из печени и осуществляет
транспорт селена к тканям, особенно мозга и яичка. В фагоцитах и в сосудистом эндотелии выступает в качестве антиоксиданта. Участвует в инактивации тяжелых металлов. Блокирование синтеза этого белка приводит к неврологическим проблемам и мужскому бесплодию |
| Селенопротеин R | SELENOR | P49908 | Участвует в восстановлении протеинов, поврежденных в окислительных процессах |
| Селенопротеин S | SELENOS | Q9BQE4 | Трансмембранный белок обнаружен в плазматической мембране и эндоплазматическом ретикулуме. Уменьшает окислительный стресс и регулирует внутриклеточное окислительно-восстановительное равновесие |
| Селенопротеин Т | SELENOT | P62341 | Участвует в мобилизации кальция |
| Селенопротеин V | SELENOV | P59797 | Потенциально участвует в мужской репродукции |
| Селенопротеин W | SELENOW | P63302 | Выступает как антиоксидант. Возможно, необходим для мышечного роста. Изучается его влияние на развитие онкологической патологии |
| Селенофосфатсинтаза 2 | SEPHS2 | Q99611 | Участвует в синтезе всех селенопротеинов |
Одна из наиболее важных функций селена связана с его канцеропротекторным действием, заключающимся в том, что система тиоредоксинредуктаз способствует индукции белка р53 (опухолевого супрессора) и запуску репарации ДНК (или активации апоптоза при необратимых нарушениях). При дефиците селена существенно снижается уровень метилирования ДНК, как за счет изменения доступности метильных доноров, так и за счет модулирования активности ДНК-метилтрансфераз, и вследствие этого повышается риск повреждения клеток (Свиридова и др., 2012). Кроме подавления экспрессии онкогенов, селен усиливает активность противоопухолевых клонов NK, стимулирует синтез ИЛ-1, ИЛ-2, ингибирует протеинкиназу С и обладает токсическим воздействием на клетки опухоли на разных стадиях развития патологического процесса (Schrauzer, 2000). Протеинкиназа С имеет структурную основу для взаимодействия с селеном: редокс-активные селеносоединения могут инактивировать фермент, особенно Ca2+-зависимые изоформы, реагируя с богатыми цистеином областями, присутствующими в каталитическом домене, а в некоторых случаях – с остатками цистеина в доменах под названием “цинковые пальцы”. Однако подробное изучение роли селена в канцерогенезе все еще находится на начальных этапах и требует проведения дополнительных экспериментальных исследований.
Рядом автором отмечалось, что в регионах с избыточным поступлением селена в организм человека наблюдается более низкая заболеваемость раком легких, прямой кишки, шейки матки и молочной железы, чем в регионах, где наблюдается дефицит этого элемента (Голубкина и др., 2002). В репрезентативной выборке из 13887 человек была обнаружена обратная зависимость между уровнем селена в сыворотке крови и смертностью от рака при концентрациях микроэлемента менее 130 нг/мл (Bleys et al., 2008). Очень показательны результаты внедрения государственной программы по коррекции дефицита микроэлементов и прежде всего – селенодефицита, проводимой в Финляндии, благодаря которой общая смертность от онкологических заболеваний снизилась почти в 2 раза.
Необходимо отметить важную роль селена в регуляции функций иммунной системы, обеспечении клеточного и гуморального иммунитета, модуляции фагоцитарной функции полиморфоядерных лейкоцитов (Koller, Exon, 1986). Кроме того, селен потенцирует выработку иммуноглобулинов (IgG, IgM), повышает активность Т-клеток и макрофагов (McKenzie et al., 2002). Как неспецифический иммуномодулятор селен оказывает положительный эффект при бронхиальной астме, атопических дерматитах, отдельных инфекционных заболеваниях (Кохан и др., 2009; Shaw et al., 1994). При этом его иммунотропное действие усиливается при сочетании с цинком, витаминами А, Е и С. Селен также незаменим для нормального функционирования щитовидной железы: он принимает активное участие как в метаболизме тиреоидных гормонов, так и в антиоксидантной защите (Шабалина и др., 2010; Drutel et al., 2013). Проведенные исследования продемонстрировали положительное влияние селена на течение хронического гастродуоденита, хронического и неспецифического язвенного колита, хронических гепатитов.
Недостаток поступления селена в организм человека вызывает одну из разновидностей гипомикроэлементозов, называемую гипоселенозом. Гипоселенозы наиболее вероятно развиваются у жителей, проживающих в районах с выраженным недостатком селена в почвах и продуктах питания. В частности, они были описаны у людей и животных, населяющих территорию Китая, Египта, Тайланда (Tapiero et al., 2003), а также в Центральной и Восточной Сибири (Тутельян и др., 2002). Симптоматика заболеваний, вызванных недостаточным поступлением микроэлемента в организм человека, в высокой степени разнообразна. В частности, дефицит селена ускоряет развитие атеросклероза, ишемической болезни сердца, повышает вероятность развития инфаркта миокарда и инсультов, сердечной недостаточности и аритмий. У больных с острым коронарным синдромом описано снижение содержания селена в сыворотке крови (среднее значение – 61.6 ± 1.6 мкг/л), что меньше среднего уровня этого микроэлемента у здоровых лиц (82.5 ± 1.8 мкг/л) и почти в 2 раза ниже оптимального уровня (Пятницкая и др., 2012). Определено значительное снижение уровня селена в крови больных с острым крупноочаговым и трансмуральным инфарктом миокарда, а снижение показателей элемента в плазме, по мнению авторов, может свидетельствовать о тяжести патологического процесса в миокарде и указывает на продолжающийся процесс активации свободнорадикальных реакций (Чаяло и др., 1992). Низкая концентрация селена ассоциируется и с будущей сердечно-сосудистой смертностью у пациентов с острым коронарным синдромом: у умерших от сердечных причин уровень селена был ниже по сравнению с выжившими (61.0 мкг/л против 71.5 мкг/л; р < 0.0001) (Lubos et al., 2010). Дефицит селена у пациентов с сердечной недостаточностью связан со снижением устойчивости к физической нагрузке и повышением смертности на 50%, а также с нарушением функции митохондрий in vitro в кардиомиоцитах человека, что отчасти связано с нарушениями переноса электронов по дыхательной цепи. В результате тормозится синтез АТФ, увеличивается продукция активных форм кислорода, развивается внутриклеточное окислительное повреждение (Bomer et al., 2020). Результаты исследований, проводившихся в регионах с низким потреблением селена, свидетельствуют о высоком риске сердечно-сосудистых заболеваний, особенно в случае сочетания дефицитов этого микроэлемента и витамина Е (Gavat, Voroniuc, 1999).
Умеренный дефицит селена увеличивает риск развития нефропатии, неврологических заболеваний, бесплодия у мужчин (Rayman, 2012), холестаза беременных, дисфункции щитовидной железы и гестационного диабета (Mistry et al., 2012). Тан с соавт. (Tan et al., 2001) показали, что среднее значение уровня сывороточного селена у женщин с нормально протекающей беременностью было 0.0741 ± 0.0167 мг/л, тогда как с нарушенной толерантностью к глюкозе – 0.0631 ± 0.0132 мг/л, с гестационным сахарным диабетом – 0.0635 ± 0.0120 и 0.108 ± 0.0170 мг/л у женщин фертильного возраста. Уровень селена в сыворотке крови был с большой степенью достоверности ниже у беременных с нарушенной толерантностью к глюкозе и с гестационным сахарным диабетом, чем у женщин с нормально протекающей беременностью.
Как показывают исследования, недостаток селена во время беременности может вызывать окислительный стресс, приводящий к выкидышам, преэклампсии, преждевременным родам (Mistry et al., 2012). Кроме того, дефицит селена способствует задержке внутриутробного развития плода и оказывает неблагоприятное воздействие на формирование его нервной системы. Так, у беременных женщин с диагностированным дефектом нервной трубки плода имели место достоверно более низкие уровни селена в сыворотке крови по сравнению с женщинами контрольной группы с документированным нормальным исходом у плода: 55.16 ± 11.3 мкг/л против 77.4 ± 5.5 мкг/л; р < 0.001(Cengis et al., 2004).
К состояниям, связанным с тяжелым дефицитом селена, относят болезни Кешана и Кашина–Бека. Болезнь Кешана – это ювенильная кардиомиопатия, встречающаяся в основном у детей от 2 до 10 лет и у молодых женщин репродуктивного возраста. Для нее характерны: увеличение размеров сердца с фокальными некрозами миокарда, развитие сердечной недостаточности, аритмий и тромбоэмболических осложнений. Болезнь Кашина–Бека проявляется поражением суставов, укорочением пальцев и конечностей, нарушением роста вследствие окислительного повреждения гиалинового хряща с его дегенерацией, атрофией и некрозом. Это заболевание чаще всего поражает детей в возрасте от 5 до 13 лет. В качестве важной этиологической причины рассматривается сопутствующий дефицит йода в организме и воздействие ряда токсикантов (Yao et al., 2011a).
В настоящее время доказано, что отрицательное действие дефицита селена на организм человека во многом связано с уменьшением активности селенопротеинов и, как следствие, нарушением биохимических процессов, в которых они участвуют (Pedrero, Madrid, 2009; Ruseva et al., 2013).
Таким образом, селен – это незаменимый для человека микроэлемент, принимающий участие в важнейших физиологических процессах в организме, поэтому его дефицит является причиной развития широкого спектра патологий, среди которых заболевания, занимающие ведущие позиции в структуре смертности населения развитых стран. Эссенциальность селена, крайняя неравномерность распределения его в различных регионах, отсутствие систематизированных данных о содержании селена в продукции растительного и животного происхождения, недостаток сведений о распределении селена в пищевых цепях, конечным звеном которых является человек, вызывают большой интерес с точки зрения профилактики и коррекции дефицита селена в организме человека.
ВОЗМОЖНЫЕ ПУТИ УСТРАНЕНИЯ ДЕФИЦИТА СЕЛЕНА
Среди возможных путей решения проблемы устранения дефицита селена в организме человека следует назвать: расширение ассортимента продуктов питания, обогащение готовых пищевых продуктов микроэлементами, использование биологически активных добавок, а также повышение его содержания в сельскохозяйственных растениях путем внесения в почву селеносодержащих удобрений или использования методов биофортификации.
В настоящее время именно биофортификации (обогащения) сельскохозяйственных растений селеном, как наиболее перспективной технологии, уделяется повышенное внимание. Как известно, цель биофортификации – улучшение поглощения и накопления элементов минерального питания в органах растений, используемых в пищу, путем селекции растений и генной инженерии, а также агрономическими методами (Wu et al., 2015). В случае селена можно указать и другие преимущества биофортификации, в частности, снижение уровня нитратов, повышение содержания природных антиоксидантов, увеличение устойчивости растений к различного рода стрессовым воздействиям.
На настоящий момент в промышленном масштабе выпускаются обогащенные селеном чеснок (США), томаты (Великобритания) и чай (Китай). В Финляндии для выращивания злаков применяются комплексные удобрения, обогащенные селенатом натрия (Ekholm et al., 2007). В качестве одной из стратегий биофортификации селеном предлагается использование микроорганизмов, в частности бактерий, обеспечивающих лучшее поглощение растениями целого ряда микроэлементов, в том числе селена (Durán et al., 2015). В России разработана технология получения обогащенного селеном стручкового перца. Доказана эффективность некорневой обработки селеном пшеницы, при которой его поступление происходит по безбарьерному типу (Кашин, Шубина, 2011).
Генная инженерия – еще одна стратегия для получения биообогащенных селеном пищевых продуктов, которая обычно направлена на усиление активности ферментов, участвующих в поглощении и ассимиляции селена. Возможность биофортификации селена с помощью методов генной инженерии была проверена на нескольких видах сельскохозяйственных культур, в том числе мягкой (Govasmark, Salbu, 2011) и твердой (Poblaciones et al., 2014) пшенице, рисе (Premarathna et al., 2012), чечевице (Rahman et al., 2013).
Вместе с тем увеличение содержания селена в органах растений как путем внесения селеносодержащих удобрений, так и за счет применения технологий биофортификации, не должно приводить к чрезмерному повышению его концентрации, поскольку высокие концентрации этого элемента токсичны как для растений (30 мг/кг сухой массы и более), так и для животных и человека (5 мг/день) (Finley, 2005; Cabata-Pendias, 2010).
ЗАКЛЮЧЕНИЕ
В настоящем обзоре предпринята попытка обобщить имеющиеся в литературе данные, касающиеся главным образом биологических последствий, связанных с дефицитом селена в почве, растениях, в организме животных и человека. О важной роли селена для человека начали говорить еще с 1960-х гг., когда впервые установили его роль в ответе клеток на окислительный стресс и в поддержании окислительно-восстановительного баланса клеток. До этого селен рассматривали только с точки зрения его токсичности для живых организмов. К настоящему времени важность селена для человека доказана многочисленными исследованиями и не подвергается сомнению. Помимо антиоксидантной роли, во многом это объясняется его участием в функционировании большого числа (порядка 30) белков, необходимых для регуляции клеточного роста и апоптоза, метаболизма ряда гормонов, в том числе гормонов щитовидной железы. Поэтому дефицит этого микроэлемента отрицательно сказывается на целом ряде метаболических процессов в организме человека и служит причиной развития широкого спектра патологических состояний и заболеваний. Особую значимость имеют исследования, направленные на изучение антиканцерогенного действия селена.
Недостаток селена в организме человека во многом связан с ограниченным поступлением его с пищей (рис. 2). Поскольку именно растительная пища является основным поставщиком селена для человека и животных, изучение поступления этого химического элемента в растения из почвы и накопления в органах, его роли для жизнедеятельности растений, а также влияние низких концентраций селена на физиологические процессы и продуктивность сельскохозяйственных культур являются в настоящее время особенно актуальными. Отметим, что хотя селен не считается необходимым микроэлементом для растений, многие исследования убедительно доказывают стимулирующий эффект обработки растений (или семян) этим элементом в низких концентрациях на продуктивность. Однако механизмы положительного воздействия селена на растения до сих пор остаются почти не изученными. Практически нет данных и о воздействии дефицита этого микроэлемента на жизнедеятельность растений. Вместе с тем такого рода данные чрезвычайно важны, и в первую очередь для выработки мероприятий по коррекции селенового статуса населения. Поэтому особую актуальность приобретают исследования, направленные на поиск возможных путей обогащения растений селеном (с помощью внесения селеносодержащих удобрений или использования методов биофортификации и генной инженерии), а также организация перевозок продуктов растительного происхождения из регионов с высоким содержанием селена в почвах в регионы с его низким содержанием.
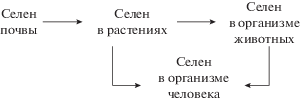
Что касается животных и человека, то по-прежнему крайне важны исследования, направленные на изучение биохимических и молекулярно-генетических механизмов, лежащих в основе различных патологических состояний и заболеваний, вызванных дефицитом селена. Эти знания необходимы не только для решения вопроса о повышении продуктивности сельскохозяйственных животных, но и, что особенно важно, для разработки новых методов, направленных на улучшение здоровья человека.
Список литературы
Авцын А.П., Жаворонков А.А., Риш М.А., Строчкова Л.С. Микроэлементозы человека: этиология, классификация, органопатология. М.: Медицина, 1991. 496 с.
Аристархов А.Н., Бусыгин А.С., Яковлева Т.А. Дефицит селена в почвах и растениях северо-восточного Нечерноземья как индикатор необходимости применения селеновых удобрений // Междунар. сельскохоз. журн. 2018. № 1 (361). С. 31–36.
Бабенко Г.А. Микроэлементозы человека: патогенез, профилактика, лечение // Микроэлементы в медицине. 2001. Т. 2. Вып. 1. С. 2–5.
Блинохватов А.Ф., Денисова Г.В., Ильин Д.Ю. и др. Селен в биосфере. Пенза: РИО ПГСХА, 2001. 324 с.
Болотников И.Ю. Экологические факторы и показатели здоровья детского населения Астраханской области // Мат. 6-й междунар. биогеохим. школы. Астрахань, 22–25 сентября 2008 г. С. 105.
Вихрева В.А. Влияние селена на рост, развитие и адаптивный потенциал козлятника восточного (Galega orientalis): Дис. … канд. биол. наук. Пенза: Пенз. ГПУ, 2001. 154 с.
Вихрева В.А., Лебедева Т.Б. Содержание селена в почвах и растениях лесостепи Среднего Поволжья // Молодой ученый. 2010. Т. 2. № 11. С. 195–198.
Вихрева В.А. Эколого-агрохимические аспекты применения селена под зерновые культуры и козлятник на черноземах лесостепи Среднего Поволжья: Автореф. дис. … докт. биол. наук. Владимир: ВлГУ, 2011. 53 с.
Вихрева В.А., Блинохватов А.А., Клейменова Т.В. Селен в жизни растений. Пенза: РИО ПГСХА, 2012. 222 с.
Голубкина Н.А., Папазян T.T. Селен в питании. Растения, животные, человек. М.: Печатный город, 2006. 269 с.
Голубкина Н.А., Скальный А.В., Соколов Я.А., Щелкунов Л.Ф. Селен в медицине и экологии. М.: КМК, 2002. 135 с.
Голубкина Н.А., Капитальчук И.П., Капитальчук М.В. Селен в почвах на разных высотных уровнях рельефа Днестровско-Прутского междуречья // Вестн. МГОУ. Сер. Естест. науки. Раздел III: География. 2012. № 1. С. 98–101.
Голубкина Н.А., Синдирева А.В., Зайцев В.Ф. Внутрирегиональная вариабельность селенового статуса населения // Юг России: экология, развитие. 2017. Т. 12. № 1. С. 107–127.
Ермаков В.В., Ковальский В.В. Биологическое значение селена. М.: Наука, 1974. 298 с.
Зайцев С.Ю., Конопатов Ю.В. Биохимия животных. Фундаментальные и клинические аспекты. CПб.: Лань, 2004. 384 с.
Иванов В.В. Экологическая геохимия элементов. Кн. 3: Редкие р-элементы. М.: Недра, 1996. 352 с.
Кармолиев Р.Х. Свободнорадикальная патология в этиопатогенезе болезней животных // Ветеринария. 2005. № 4. С. 42–47.
Касумов С.Н. Биологическое значение селена для жвачных животных. М.: ВНИИТЭИСХ, 1979. 49 с.
Кашин В.К., Шубина О.И. Биологическое действие и накопление селена в пшенице в условиях селенодефицитной биогеохимической провинции // Химия в интересах устойчивого развития. 2011. № 19. С. 151–156.
Кохан С.Т., Намоконов Е.В., Захарова О.А. и др. Влияние изменения селенового статуса на показатели цитокинов у больных с внебольничными пневмониями // Сиб. мед. журн. 2009. № 8. С. 30–32.
Кулагина Ю.М., Головацкая И.Ф. Влияние селенита натрия на рост и развитие растений пшеницы в зависимости от способа обработки // Вестн. ТГУ. Биология. 2011. № 2 (14). С. 56–64.
Прудников П.С. Влияние селена на физиолого-биохимические процессы при адаптации растений картофеля к гипотермии: Дис. … канд. биол. наук. М.: МСХА, 2007. 146 с.
Пятницкая С.В., Рудь С.С., Ковальский Ю.Г. и др. Содержание селена и состояние свободнорадикального окисления в сыворотке крови у больных с острым коронарным синдромом без стойкого подъема сегмента ST // Дальневост. мед. журн. 2012. № 2. С. 18–21.
Свиридова С.П., Кашия Ш.Р., Обухова О.А., Чучуев Е.С. Возможности эссенциального селена в онкологии // Вестн. РОНЦ им. Н.Н. Блохина РАМН. 2012. Т. 23. № 3. С. 8–9.
Серегина И.И. Влияние селена на продуктивность яровой пшеницы в зависимости от азотного питания и водообеспечения // Плодородие. 2007. № 5. С. 15–17.
Серегина И.И., Ниловская Н.Т. Биологическая роль селена в растениях // Агрохимия. 2002. № 10. С. 76–85.
Сидельникова В.Д. Геохимия селена в биосфере // Проблемы биогеохимии и геохимической экологии. М.: Наука, 1999. Т. 23. С. 81–99.
Степанов Ю.М., Белицкий В.В., Косинская С.В. Селен как микроэлемент: характеристика и значение для человека // Сучасна гастроентерологія. 2012. № 3 (65). С. 91–96.
Трошина Е.А., Сенюшкина Е.С., Терехова М.А. Роль селена в патогенезе заболеваний щитовидной железы // Клин. эксперим. тиреоидол. 2018. Т. 14. № 4. С. 192–205.
Тутельян В.А., Княжев В.А., Хотимченко С.А. и др. Селен в организме человека: метаболизм, антиоксидантные свойства, роль в канцерогенезе. M.: РАМН, 2002. 224 с.
Усольцева И.И., Гатаулина Л.Р., Гасанов А.С., Зиятдинов Р.Н. Беломышечная болезнь поросят, лечение и профилактика // Уч. записки КГАВМ им. Н.Э. Баумана. 2010. № 5. Т. 204. С. 293–296.
Чаяло П.П., Соловьев А.В., Ена Я.М. и др. Содержание церулоплазмина, фибронектина и селена в крови больных острым инфарктом миокарда // Врачебное дело. 1992. Т. 996. № 3. С. 15–17.
Шабалина Е.А., Моргунова Т.Б., Орлова С.В., Фадеев В.В. Селен и щитовидная железа // Клин. эксперим. тиреоидол. 2010. Т. 7. № 2. С. 7–18.
Шеуджен А.Х., Лебедовский И.А., Бондарева Т.Н. Биогеохимия и агрохимия селена [Эл. ресурс] // КубГАУ. 2013. № 92 (08). С. 781–798.
Шеуджен А.Х., Аканова Н.И., Хурум Х.Д. и др. Селен в черноземе выщелоченном [Эл. ресурс] // КубГАУ. 2018. № 138. С. 160–170.
Aréchiga C.F., Vázquez-Flores S., Ortiz O. et al. Effect of injection of β-carotene or vitamin E and selenium on fertility of lactating dairy cows // Theriogenology. 1998. V. 50. P. 65–76.
Bianco A.C., Salvatore D., Gereben B. et al. Biochemistry, cellular and molecular biology, and physiological roles of the iodothyronineselenodeiodinases // Endocr. Rev. 2002. V. 23 (1). P. 38–89.
Bleys J., Navas-Acien A., Guallar E. Serum selenium levels and all-cause, cancer, and cardiovascular mortality among US adult // Arch. Int. Med. 2008. V. 168 (4). P. 404–410.
Bomer N., Beverborg N.G., Hoes M.F. et al. Selenium and outcome in heart failure // Eur. J. Heart Fail. V. 2020. № 22 (8). P. 1415–1423.
Byrne S.L., Durandeau K., Nagy I., Barth S. Identification of ABC transporters from Lolium perenne L. that are regulated by toxic levels of selenium // Planta. 2010. V. 231. № 4. P. 901–911. https://doi.org/10.1007/s00425-009-1096-y
Byun B.J., Kang Y.K. Conformational preferences and pKa value of selenocysteine residue // Biopolymer. 2011. V. 95. № 5. P. 345–353. https://doi.org/10/1002/bip.21581
Cabata-Pendias A. Trase elements in soils and plant. FL: CrsPress, 2010. 548 p.
Cardona Á.J., Reza L.G. Esteatosis en un burro (Equus asinus). Primer reporte en Colombia // Rev. MVZ Córdoba. 2011. V. 16. P. 2793–2798.
Drutel A., Archambeaud F., Caron P. Selenium and the thyroid gland: more good news for clinicians // Clin. Endocrinol. (Oxf). 2013. V. 7. № 2. P. 155–164.
Durán P., Acuña J.J., Gianfreda L. et al. Endophytic selenobacteria as new inocula for selenium biofortification // Appl. Soil. Ecol. 2015. V. 96. P. 319–326.
Duntas L.H., Benvenga S. Selenium: an element for life // Endocrine. 2015. V. 48. № 3. P. 756–775.
Ekholm P., Reinivuo H., Mattila P. et al. Changes in the mineral and trace element contents of cereal, fruits and vegetables in Finland // J. Food Comp. Anal. 2007. V. 20. P. 487−495.
Ermakov V.V. Problems of extremal geochemical ecology and biogeochemical study of the biosphere // Biogeochem. Geochem. Ecol. (NPC TMG MZ RF, Moscow). 2001. P. 98–144.
Fajt Z., Svoboda M., Drábek J., Dubanský V. Selen a jeho význam pro zdravotní stav prasat – review // Veterinářství. 2009. V. 59. P. 221–224.
Finley J.W. Selenium accumulation in plant foods // Nutr. Rev. 2005. V. 63 (6). P. 196−202.
Finch J.M., Turner R.J. Effects of selenium and vitamin E on the immune responses of domestic animals // Res. Vet. Sci. 1996. V. 60. P. 97–106.
Feng R., Wei C., Tu S. The roles of selenium in protecting plants against abiotic stresses // Environ. Exp. Bot. 2013. V. 87. P. 58–68.
Gavat V., Voroniuc O. Oxidative stress and antioxidants in the diet in pathological processes at the level of the cardiovascular system // Rev. Med. Chir. Soc. Med. Nat. Iasi. I999. V. 103. № 1–2. P. 37–41.
Govasmark E., Salbu B. Translocation and re-translocation of selenium taken up from nutrient solution during vegetative growth in spring wheat // J. Sci. Food Agric. 2011. V. 91. P. 1367–1372.
Holovská K.Jr., Holovská K., Boldižárová K. et al. Antioxidant enzyme activities in liver tissue of chickens fed diets supplemented with various forms and amounts of selenium // J. Anim. Feed Sci. 2003. V. 12. P. 143–152.
Ibrahim S.A.Z., Kerkadi A., Agouni A. Selenium and health: an update on the situation in the Middle East and North Africa // Nutrients. V. 11 (7). P. 1457.
Kamada H., Nonaka I., Takenouchi N., Amari M. Effects of selenium supplementation on plasma progesterone concentrations in pregnant heifers // Anim. Sci. J. 2014. V. 85. P. 241–246.
Koller L.D., Exon J.H. The two faces of selenium – deficiency and toxicity – are similar in animals and man // Can. J. Vet. Res. 1986. V. 50. P. 297–306.
Levander O.A. Scientific rationale for the 1989 recommended dietary allowance for selenium // J. Am. Diet. Assoc.1991. V. 91 (12). P. 1572–1576.
Li H.-F., McGrath S.P., Zhao F.-J. Uptake, translocation and speciation in wheat supplied with selenate or selenite // New Phytologist. 2008. V. 178. P. 92–102.
Lubos E., Sinning C.R., Schnabel R.B. et al. Serum selenium and prognosis in cardiovascular disease: results from the AtheroGene study //Atherosclerosis. 2010. V. 209 (1). P. 271–277.
McKenzie R.C., Arthur J.R., Miller S.M. et al. Selenium and the immune system // Nutrition and immune function. UK, Wallingford: CABI Publishing, 2002. P. 239–250.
Mangiapane E., Pessione A., Pessione E. Selenium and selenoproteins: an overview on different biological systems // Curr. Protein. Pept. Sci. 2014. V. 15. P. 598–607.
Misra S., Boylan M., Selvam A. et al. Redox-active selenium compounds – from toxicity and cell death to cancer treatment // Nutrition. 2015. V. 7. P. 3536–3556.
Mistry H.D., Pipkin F.B., Redman C.W., Poston L. Selenium in reproductive health // Am. J. Obstet. Gynecol. 2012. V. 206. № 1. P. 21–30.
Pedrero Z., Madrid Y. Novel approaches for selenium speciation in foodstuffs and biological specimens: a review // Anal. Chim. Acta. 2009. V. 634. № 2. P. 135–152.
Poblaciones M.J., Rodrigo S., Santamaría O. et al. Agronomic selenium biofortification in Triticum durum under Mediterranean conditions: from grain to cooked pasta // Food Chem. 2014. V. 146. P. 378–384.
Preda C.,Vasiliu I., Mihalache L. et al. Selenium – essential antioxidant element. The example of autoimune thyroiditis // Rev. Chim. 2017. V. 68 (7). P. 1617–1621.
Radostits O.M., Arundel J.H., Gay C.C. et al. Veterinary medicine: a textbook of the diseases of cattle, sheep, pigs, goats and horses. 9th ed. London, UK: WB Saunders Company Ltd., 2000. 1877 p.
Premarathna L., McLaughlin M.J., Kirb J.K. et al. Selenate-enriched urea granules are a highly effective fertilizer for selenium biofortification of paddy rice grain // J. Agric. Food Chem. 2012. V. 60. P. 6037–6044.
Rahman M.M., Erskine W., Zaman M.S. et al. Selenium biofortification in lentil (Lens culinaris Medikus subsp. culinaris): farmers, field survey and genotype × environment effect // Food Res. Int. 2013. V. 54. P. 1596–1604.
Rayman M.P. Selenium and human health // Lancet. 2012. V. 397 (9822). P. 1256–1268.
Ruseva B., Himcheva I., Nankova D. Importance of selenoproteins for the function of the thyroid gland // Medicine. 2013. № 3. P. 60–64.
Schrauzer G.N. Selenomethionine: a review of its nutritional significance, metabolism and toxicity // J. Nutr. 2000. V. 130. P. 1653–1656.
Shaw R., Woodman K., Crane J. et al. Risk factors for asthma symptoms in Kawerau children // N Z Med. J. 1994. № 12. V. 107 (987). P. 387–391.
Sordillo L.M. Selenium-dependent regulation of oxidative stress and immunity in periparturient dairy cattle // Vet. Med. Int. Article ID 154045. 2013. https://doi.org/10.1155/2013/154045
Stadlober M., Sager M., Irgolic K.J. Effects of selenate supplemented fertilisation on the selenium level of cereals – identification and quantification of selenium compounds by HPLC-ICP-MS // Food Chem. 2001. V. 73. P. 357–366.
Streeter R.M., Divers T.J., Mittel L. et al. Selenium deficiency associations with gender, breed, serum vitamin E and creatine kinase, clinical signs and diagnoses in horses of different age groups: a retrospective examination 1996–2011 // Equine Vet. J. Suppl. 2012. V. 44. P. 31–35.
Stec A., Mochon J., Kurek L. et al. The influence of different factors on selenium levels in dairy cow herds in the Central-Eastern region of Poland // Pol. J. Vet. Sci. 2005. № 8. P. 225–229.
Tan M., Sheng L., Qian Y. et al. Changes of serum selenium in pregnant women with gestational diabetes mellitus // Biol. Trace Elem. Res. 2001. V. 83 (3). P. 7231–7237.
Tapiero H., Townsend D.M., Tew K.D. The antioxidant role of selenium and seleno-compounds // Biomed. Pharmacother. 2003. № 57 (3–4). P. 134–144.
Uematsu M., Kitahara G., Sameshima H., Osawa T. Serum selenium and liposoluble vitamins in Japanese Black cows that had stillborn calves // Vet. Med. Sci. V. 2016. № 78. P. 1501–1504.
Underwood E.J., Suttle N.F. Selenium in the mineral nutrition of livestock. 3rd ed. Wallingford, UK: CABI Publishing, 2004. P. 421–475.
van Loon G., Lefère L., Bauwens C. et al. Yellow fat disease (steatitis): description of 20 cases with emphasis on typical ultrasonographic findings // Equine Vet. J. 2015. V. 47. Suppl. V. 48. P. 19. https://doi.org/10.1111/evj.12486_43
van Hoewyk D. A tale of two toxicities: malformed selenoproteins and oxidative stress both contribute to selenium stress in plants // Ann. Bot. 2013. V. 112. P. 965–972.
White P.J. Selenium accumulation by plants // Ann. Bot. 2016. V. 117. P. 217–235.
Whanger P. D. Selenocompounds in plants and animals and their biological significance // J. Am. Coll. Nutr. 2002. V. 21. P. 223–232.https://doi.org/10.1080/07315724.2002.10719214
Wiesner-Reinhold M., Schreiner M., Baldermann S. et al. Mechanisms of selenium enrichment and measurement in brassicaceous vegetables, and their application to human health // Front. Plant Sci. 2017. V. 8. https://doi.org/10.3389/fpls.2017.01365
Wu Z., Bañuelos G.S., Lin Z.Q. et al. Biofortification and phytoremediation of selenium in China // Front. Plant Sci. 2015. V. 6. P. 136.https://doi.org/10.3389/fpls.2015.00136
Yang G., Yin S., Zhou R. et al. Studies of safe maximal daily dietary Se-intake in a seleniferous area in China. Part II: Relation between Se-intake and the manifestation of clinical signs and certain biochemical alterations in blood and urine // J. Trace Elem. Electr. Health Dis. 1989. № 3 (3). P. 123–130.
Yao Y., Pei F., Kang P. Selenium, iodine, and the relation with Kashin–Beck disease // Nutrition. 2011a. V. 27. № 11–12. P. 1095–1100.
Yao X., Chu J., He X., Ba C. Protective role of selenium in wheat seedlings subjected to enhanced UV-B radiation // Russ. J. Plant Physiol. 2011b. V. 58. P. 283–289.
Żarczyńska K., Sobiech P., Radwińska J., Rękawek W. Effects of selenium on animal health // J. Elementol. 2013. V. 18. P. 329–340.
Zayed A.M., Lytle C.M., Terry N. Accumulation and volatilization of different chemical species of selenium by plants // Planta. 1998. V. 206. P. 284–292.
Zeng H., Combs G.F.Jr. Selenium as an anticancer nutrient: roles in cell proliferation and tumor cell invasion // J. Nutr. Biochem. 2008. V. 19. P. 1–7.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии