

Успехи современной биологии, 2022, T. 142, № 1, стр. 37-51
Современные представления о токсинах высших грибов: безазотистые органические соединения
А. А. Ховпачев 1, *, В. А. Башарин 1, С. В. Чепур 2, **, Д. В. Цой 2, И. М. Иванов 2, С. В. Волобуев 3, ***, М. А. Юдин 2, А. С. Никифоров 2, Л. Б. Калинина 3
1 Военно-медицинская академия им. С.М. Кирова
Санкт-Петербург, Россия
2 Государственный научно-исследовательский испытательный институт военной медицины
Санкт-Петербург, Россия
3 Ботанический институт им. В.Л. Комарова РАН
Санкт-Петербург, Россия
* E-mail: vtmz@vmeda.org
** E-mail: gniiivm_2@mail.ru
*** E-mail: rbo.fungi@binran.ru
Поступила в редакцию 06.03.2021
После доработки 09.07.2021
Принята к публикации 25.07.2021
- EDN: BQCWGY
- DOI: 10.31857/S0042132421050045
Аннотация
В продолжение обсуждения токсинов высших грибов рассмотрены их ядовитые безазотистые органические соединения. Обсуждены проблемы их изучения и систематизации. Проанализированы современные представления о токсических процессах, индуцируемых ядовитыми терфенилами, сесквитерпенами, тритерпенами, трихотеценами и олигоизопреноидами. Представлены некоторые недавно открытые цитотоксические соединения подобной структуры. Сделан вывод о том, что системное изучение токсинов безазотистой структуры востребовано, в то время как данная область исследований остается малоосвоенной.
ВВЕДЕНИЕ
Вследствие того, что высшие грибы (макромицеты) испытывают постоянную угрозу со стороны микофагов, одна из ключевых стратегий их защиты реализована в производстве токсинов. Тяжелые последствия интоксикаций их плодовыми телами, содержащими в качестве главных ядовитых компонентов безазотистые органические соединения, определяют актуальность их исследований для современной биологии и медицины (Медицинская токсикология …, 2012; Sterner, Anke, 1995; Yokoyama, Gonmori, 2009; Beuhler, 2016; Yin et al., 2019).
Тем не менее в российской (Магазов и др., 2019) и зарубежной (Sterner, Anke, 1995; Yin et al., 2019) токсинологии изучение безазотистых ядовитых соединений не относят к числу приоритетных исследований. Как следствие, остаются не раскрытыми многие механизмы их действия, объясняющие проявления детерминированных ими расстройств. Фундаментальная проблема токсичности веществ подобной структуры требует системного изучения (McMorris et al., 2003; Schüffler, 2018). К медицинским аспектам этой проблемы относят совершенствование токсикологической помощи при интоксикациях макромицетами, или мицетизмах (шифры МКБ-10 T 40.9, Т 62.0) (Медицинская токсикология…, 2012; Beuhler, 2016). Вместе с тем, за минувшее десятилетие описаны новые ядовитые вещества безазотистой структуры, изучены ранее неизвестные токсидромы (White et al., 2019). В этой связи в продолжение обсуждения токсинов макромицетов назревает необходимость обобщения сведений о структурах и путях реализации эффектов их ядовитых безазотистых органических соединений.
РОЛЬ БЕЗАЗОТИСТЫХ ОРГАНИЧЕСКИХ СОЕДИНЕНИЙ ВЫСШИХ ГРИБОВ В ФОРМИРОВАНИИ ПАТОЛОГИЧЕСКИХ ПРОЦЕССОВ
Безазотистые токсины макромицетов представлены среди ароматических и терпеновых структур. Традиционно считают, что основная причина интоксикаций содержащими их грибами обусловлена пищевой значимостью внешне сходных с ними съедобных видов (Мусселиус, Рык, 2002). Кроме этого, грибы – ценный источник лекарственного сырья (Abraham, 2011; Gill, 2003).
Терфенилы
Терфенилы – это углеводороды, структурно содержащие бис-фенилзамещенный ароматический цикл, либо циклохинон (терфенилхиноны) (Li et al., 2017). В природе о-терфенилы неизвестны, м-производные встречаются в ограниченном количестве, а п-терфенильные соединения в настоящий момент выявлены только среди лишайников и грибов. В этой связи, п-терфенилы уникальны в своем роде. Вместе с тем, терфенилы макромицетов представляют обширный класс пигментов: лейкомелон представителей рода Inonotus P. Karst., гироцианин Гиропоруса синеющего (Gyroporus cyanescens (Bull.) Quél.), флебиаруброн Пунктулярии щетинисто-зональной (Punctularia strigosozonata (Schwein.) P.H.B. Talbot) и т.д. (Zhou, Liu, 2010), аурантрихолон Трихоломы золотисто-оранжевой (Tricholoma aurantium (Schaeff.) Ricken) и т.д. В целом, рассматриваемые пигменты малоядовиты для млекопитающих: наибольшую среди них среднесмертельную дозу (ЛД50) наблюдают у терферола (1) – 160 мг/кг, мыши в/бр (рис. 1) (The Merck index …, 2006). Однако, среди терфенилов макромицетов токсикологическим значением обладают полипоровая (2), вульпиновая (3) и усталиновая (4) кислоты.
Рис. 1.
Химическая структура ядовитых терфенилов и терфенилоподобных соединений макромицетов (адаптировано по: Sano et al., 2002; Schüffler, 2018).
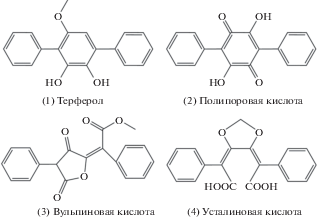
С отравлениями полипоровой кислотой (4С по White et al., 2019), содержащейся в базидиомах Гапалопилуса красноватого (Hapalopilus rutilans (Pers.) Murrill), связывают всего 7 случаев интоксикации в Германии и Франции, произошедшие в период с 1986 г. по 2013 г. (Villa et al., 2013). Плодовые тела H. rutilans отличает необычайно высокое содержание данного соединения – до 40% от сухой массы (Bauer et al., 1990). Пигмент часто встречают и в других афиллофороидных грибах, а также лишайниках, но в гораздо меньших количествах (Gill, 2003; Li et al., 2017).
Все известные интоксикации H. rutilans вызваны его ошибочной идентификацией в качестве сходной с ним и потребляемой в пищу в этих краях Фистулины печеночной (Fistulina hepatica (Schaeff.) With.) (Villa et al., 2013). Интоксикации сопровождаются отделением фиолетовой мочи в течение нескольких дней, что неудивительно, поскольку в зависимости от pH полипоровая кислота приобретает оттенки от красного до темно-коричневого цвета. Латентный период до проявления гастроинтестинального синдрома составляет около 12 ч. Далее, к концу 1–2 сут возникают диплопия, разнонаправленный нистагм, оглушение и зрительные галлюцинации. Неврологические нарушения проходят к 4 сут (White et al., 2019). Неспецифические изменения регуляции витальных функций, признаки отека мозга и диффузная лабильность волновой активности на спонтанной электроэнцефалограмме возвращаются к референсным показателям только на 7–10 сут (Kraft et al., 1998). Лабораторно интоксикации сопровождаются легким и умеренным цитолитическим синдромом, метаболическим ацидозом, гипокалий- и гипокальцийемией и явлениями почечного повреждения: повышением сывороточных концентраций мочевины, креатинина и альбуминурией (Bauer et al., 1990; Villa et al., 2013).
Подобные изменения выявляют при внутривенных (в/в), внутрибрюшинных (в/бр) и внутрижелудочных (в/ж) интоксикациях грызунов. Токсичность per os полипоровой кислоты для мышей относительно невысока – ЛД50 = 800 мг/кг. При этом нейродепрессивные эффекты возникают уже при ее введении в дозах более 100 мг/кг и проявляются снижением мышечной силы, локомоторной активности, нарушениями темнового рефлекса и координации движений (Kraft et al., 1998).
По механизму действия полипоровую кислоту относят к ингибиторам Src-тирозин-протеинкиназы (КФ 2.7.10.2), дигидрооротатдегидрогеназы (КФ 1.3.5.2). В отношении дигидрооротатдегидрогеназы ее полуингибирующая концентрация (IC50) составляет 0.5 ммоль/л (Burton, Cain, 1959; Kraft et al., 1998). Помимо этого, известно, что она проявляет свойства агентов, разобщающих окислительное фосфорилирование. Соединение обладает способностью переносить Н+ через мембраны дыхательной цепи, минуя протонные каналы, а также может встраиваться в кристы митохондрий, делая их проницаемыми для Н+. В результате ликвидируется протонный градиент, что ведет к увеличению скорости окисления, однако фосфорилирования АДФ не происходит. Вследствие разобщения этих процессов возникает энергодефицит (Abo-Khatwa et al., 1996; Chen, Liu, 2017).
Ранее способность полипоровой кислоты влиять на обмен пуриновых оснований посредством ингибирования перечисленных ферментов рассматривали как цитостатическую в рамках онкотерапии лейкемии (Burton, Cain, 1959). Однако в отличие от аналогичного по основному механизму цитостатика – бреквинара, полипоровая кислота в дозе 100 мг/сут не оказывает у грызунов противовоспалительного действия на модели окзазолон-индуцированного отека. Наряду с отсутствием избирательности в отношении опухолевых клеток, это послужило основанием для прекращения дальнейших исследований, посвященных внедрению этого соединения в клиническую практику (Jafee et al., 1993).
Полипоровая кислота приводит нервные клетки к гибели вследствие апоптоза. Показано, что в ходе 2 сут инкубации с линиями глии, нейронов и феохромацитомы PC-12 она вызывает утрату их способности к адгезии и расщепление ДНК-активированными эндонуклеазами (Jafee et al., 1993). Тем не менее, считают, что механизм нейротоксичности полипорового терфенилхинона, опосредующий неврологические и психические изменения в ходе интоксикации, носит функциональный характер и изучен слабо (Villa et al., 2013).
Терфенил – вульпиновую кислоту – выявляют в плодовых телах Пульвероболета Равенеля (Pulveroboletus ravenelii (Berk. ex M.A. Curtis) Murrill), встречаемого в странах Северной Америки (Stephenson, Rundel, 1979; Duncan et al., 2003). Вульпиновую кислоту – ЛД50 = 178 мг/кг, мыши в/бр, также относят к группе лишайниковых, поскольку ее накапливают лишайники родов Letharia и Vulpicida. Издавна пастухи использовали их ядовитые свойства для истребления нападающих на стада хищников, закладывая лишайники в тушки животных (Вишневский, 2019; Abo-Khatwa et al., 1996). К сходным по структуре, но менее токсичным соединениям, относят хризопетраровую, лепрариновую и ризокарповую кислоты. По всей видимости, они выполняют функцию пропеллентов, подавляющих рост конкурентов лишайников за субстрат (Li et al., 2017).
Ядовитые производные пигмента флебиаруброна – усталиновую кислоту (4) и ее метаболиты, относят к основным токсинам Рядовки опаленной (Tricholoma ustale (Fr.) P. Kumm.) (Sano et al., 2002; Sawayama et al., 2006). Свежие базидиомы T. ustale в среднем содержат 4 мкг/г усталиновой кислоты и при попадании в желудок приводят к выраженному резиноидному, или желудочно-кишечному синдрому со спазмами, рвотой и диареей (5 группа по White et al., 2019). Это один из трех видов грибов, наиболее часто вызывающих мицетизмы в Японии. Так, в период с 1989 г. по 2010 г. было зарегистрировано в общей сложности 347 пострадавших (Beuhler, 2016; Yoshioka et al., 2020). В условиях резорбтивного действия токсины T. ustale способны оказывать нейро- и кардиотоксические эффекты. Основные механизмы токсичности усталиновой кислоты опосредованы ингибированием Na+/K+-АТФазы. Концентрации, наполовину ингибирующие работу белкового переносчика в нейронах и энтероцитах свиньи, составляют 5.2 и 0.77 ммоль/л соответственно (Sano et al., 2002).
Латентный период между приемом в пищу T. ustale и возникновением резиноидного синдрома составляет от 30 мин до 3 ч. При тяжелых интоксикациях возникают неврологические нарушения (Yoshioka et al., 2020). Аналогично, при внутрибрюшинном введении ЛД50 у грызунов в дополнение к абдоминальному синдрому развиваются тремор, судороги. Смерть животных происходит в результате остановки дыхательных движений на высоте судорог (Sano et al., 2002).
Несмотря на то, что цитотоксические свойства терфенилов начали изучать уже в середине прошлого века, на настоящий момент отношения структура–активность данной группы соединений весьма расплывчаты (Li et al., 2017). Тем не менее, за последние 20 лет были проанализированы механизмы действия для некоторых из них. Как и полипоровая кислота, атроментин Свинушки толстой (Tapinella atrotomentosa (Batsch) Šutara (= Paxillus atrotomentosus (Batsch) Fr.) индуцирует процессинг каспазы-3 в клетках лейкемии человека (Kim, Lee, 2009). Телефоровая кислота, пигмент Омфалота подиллюдового (Omphalotus subilludens (Murrill) H.E. Bigelow) и Полиозеллуса сложного (Polyozellus multiplex (Underw.) Murrill), проявляет свойства ингибитора пролилолигопептидазы (КФ 3.4.21.26) нейронов человека и позиционируется в качестве терапевтического средства при болезни Альцгеймера (Ju-Yeon et al., 1999). Встречаемый среди афиллофороидных грибов терферол ингибирует циклическую аденозин-3,5-монофосфатфосфодиэстеразу (КФ 3.1.4.53) (Li et al., 2017).
Пироны
В отечественных рекомендациях по лечению интоксикаций макромицетами (Мусселиус, Рык, 2002; Клинические рекомендации…, 2012) упомянут стерил-пироновый психодислептический синдром со зрительными галлюцинациями. В качестве ассоциированных с ним макромицетов приводят Гимнопилус Юноны (Gymnopilus junonius (Fr.) P.D. Orton), Г. великолепный (G. spectabilis (Weinm.) A.H. Smith.), Строфарию сине-зеленую (Stropharia aeruginosa (Curtis) Quel.), Псилоцибе увенчанную (Psilocybe coronilla (Bull.) Noordel (= Stropharia coronilla (Bull.) Quel.), Чешуйчатку чешуйчатую (Pholiota squarrosa (Vahl) P. Kumm.). Среди представленных видов псилоцин был достоверно выявлен только в плодовых телах G. spectabilis (Hatfield, Valdes, 1978).
Данное состояние в руководствах ассоциируют с меконовой кислотой (5) – соединением класса пиронов (рис. 2). Этимология названия синдрома также подразумевает участие в токсическом процессе стирилзамещенных пиронов, или стирилпиронов (styrylpyrones).
Рис. 2.
Химическая структура пиронов макромицетов (5–7) и строение психоактивных кава-лактонов кустарника Piper methysticum (8–10) (адаптировано по: Lee, Yun, 2011).

Меконовая кислота в качестве меконата морфия выделена Ф. Сертюнером из Мака снотворного (Papaver somniferum L., 1753). С начала прошлого века кислоту используют в качестве аналитического маркера на присутствие алкалоидов опия в биологических жидкостях: с хлорным железом дает глубоко-красное окрашивание. Начиная с этого периода, соединению приписывают слабое наркотическое действие, что в настоящее время признают ошибочным суждением (Fairbairn, Steele, 1981). Считают, что роль меконовой кислоты, наряду с фумаровой, янтарной и другими первичными метаболитами, может состоять в образовании растворимых солей с алкалоидами. По-видимому, вследствие этого ее находят в различных биологических источниках, в том числе съедобных грибах (Schüffler, 2018). В то же время, меконовую кислоту активно используют в косметологической промышленности как ингибитор тирозиназы, способствующий осветлению кожи (Kletskov et al., 2018).
Брейди с соавт. показано наличие стирилпиронов бис-норянгонина (6) и гиспидина (7) в плодовых телах G. spectabilis (Hatfield, Brady, 1969) и Чешуйчатке чешуйчатожирной (Pholiota squarrosoadiposa J.E. Lange) (рис. 2) (Brady, Benedict, 1972). Двигаясь в русле общего развития идей о соединениях пиронового ряда растительного происхождения, авторами высказано предположение, что выявленные стирилпироны также обладают и галлюциногенными свойствами. Ссылаясь на эти исследования, в ряде работ данные соединения отождествляют с психоактивными стирилпиронами Перца опьяняющего (Piper methysticum Forst F., 1786) (Вишневский, 2019). Последние также известны как кава-лактоны: название кустарника P. methysticum “kava” c полинезийского – горькое растение.
Однако гиспидин, бис-норянгонин и еще 44 стирилпирона со структурами иноскавина, интерофунгина, феллигридина и даваллиалактона к настоящему времени выявлены среди широкого списка дереворазрушающих грибов, которые не обладают галлюциногенными свойствами. Вместе с тем, среди индивидуальных соединений пиронов грибного происхождения описаны только антиоксидантная, гипогликемическая и цитотоксическая активности (Zhou, Liu, 2010; Lee, Yun, 2011).
Кава-лактоны перца действительно обладают психотропными свойствами. Опьяняющие эффекты корня растения связывают с 18 соединениями, из которых до 96% экстрагируемой безазотистой массы приходится на каваин (8), янгонин (9), метистицин (10) и 3 их производных (White, 2018). В отличие от известных стирилпиронов грибов, все психогенные кава-лактоны P. methysticum содержат метокси-группу в 4 положении гетороцикла, которая представляет их общую черту (Lee, Yun, 2011). Таким образом, отсутствие галлюциногенных свойств среди известных пиронов макромицетов можно объяснить тем, что структурно они не идентичны кава-лактонам P. methysticum.
Тем не менее, стирилпироны грибов и кава-лактоны объединяет и много общего в строении. Логичен вопрос: есть ли сходство в проявлениях описанного мицетизма и токсического действия кавы, опосредованное общими чертами структур их биоактивных соединений? С этой позиции необходимо более детальное рассмотрение известных механизмов действия и наблюдаемых эффектов кава-лактонов.
Потребление напитка на основе порошка корня растения вызывает обратимую анестезию рта и кожи, эйфорию, седативный эффект, миорелаксацию, атаксию и ступорозный сон. Вместе с тем, в отчетах о токсическом действии P. methysticum на зрительных галлюцинациях, которые приводят в качестве основного дифференциально-диагностического критерия рассматриваемого мицетизма, внимание не акцентируют, либо не упоминают (White, 2018). Таким образом, описание грибного стерил-пиронового синдрома мало соответствует состоянию, наблюдаемому в ходе токсического действия P. methysticum.
По мнению Шульгина (Shulgin, 1973), существует некоторая неопределенность и противоречивость в отношении ниши кава-лактонов среди психотомиметиков, которая до сих пор не разрешена. Так, в исследованиях in vitro и in vivo выявлен довольно широкий спектр их центральных эффектов: усиление связывания лиганда с рецептором γ-аминомасляной кислоты (ГАМК) типа A, ингибирование потенциал-зависимых Na+- и Ca2+-каналов, снижение обратного захвата дофамина и норадреналина и усиление ацетилхолиновой нейротрансмиссии, возможно, из-за ингибирования ацетилхолинэстеразы (Sarris et al., 2011). По всей видимости, они могут модулировать серотониновую и каннабиноидную рецепторные системы. Важно отметить, что антагонист бензодиазепинового сайта ГАМК-рецепторов – флумазенил не ингибирует эффекты кава-лактонов, что исключает их работу через этот сайт. Аналогичным образом, µ-антагонист налоксон не влияет на биологическое действие кава-лактонов, предполагая, что опиоидные рецепторы не участвуют в реализации их эффектов (Sarris et al., 2011; White, 2018). Помимо этого, кава-лактоны оказывают выраженный депрессивный эффект в отношении активности цитохрома P450 и слабое ингибирование моноаминоксидазы B (Anke, Razman, 2004).
В ходе клинических испытаний кава-лактонов в качестве препаратов с анксиолитическим действием показано, что они обладают высоким риском фармакологического взаимодействия (White, 2018). Однако клинические исследования претерпели неудачу именно по причине того, что кава-лактоны P. methysticum обладают гепатотоксичностью (Sarris et al., 2011). Цитотоксические эффекты выявляют у гиспидина и бис-норянгонина (Li et al., 2017).
Таким образом, к общим чертам пиронов растительного и грибного происхождения относят только их цитотоксичность. Наряду с этим, в настоящее время отсутствуют данные, подтверждающие психоактивные свойства как меконовой кислоты, так и стирилпиронов макромицетов. В рецензируемых зарубежных изданиях стерил-пироновый синдром среди мицетизмов не упоминают, а клинические наблюдения отсутствуют. Суммируя различные аргументы, выделение среди интоксикаций психоактивными грибами данной нозологии в отечественных источниках дискутабельно и остается на уровне необоснованных допущений.
Терпеновые соединения
Терпеновые соединения составляют обширную и разнообразную группу ненасыщенных углеводородов – производных изопрена (2-метилбутадиен-1,3). Их, совместно со стероидами, относят к неомыляемым липидам. Структурно терпеновые соединения делят на димеры (монотерпены), тримеры (сесквитерпены) или полимеры (политерпены) изопрена, и их кислородные производные – терпеноиды: спирты, альдегиды и кетоны. Монотерпены из макромицетов, в отличие от растений, не выделены. Грибы продуцируют токсичные метаболиты с изопреновыми включениями, которые иногда рассматривают отдельно от терпенов. Так, производные лизергиновой кислоты содержат присоединенные к индолу триптамина изопреновые звенья, а меротерпеноиды – сесквитерпеноидный фрагмент совместно с поликетидом (Abraham, 2011).
Считают, что макромицеты способны вырабатывать некоторые соединения подобной структуры в ходе реализации антифидантной защиты, запускаемой после механического повреждения. Формально такой подход к продукции вторичных метаболитов, которые не участвуют в процессах ауторепарации, может соответствовать процессу токсинообразования. По такому принципу в базидиомах накапливаются сесквитерпены группы лактарана: бленнины A и D Рыжика соснового (Lactarius deliciosus (L.) Gray), лактароруфин и фурандиол Груздя горького (L. rufus (Scop.) Fr.) (Chen, Liu, 2017).
В настоящее время идентифицировано более 200 цитотоксичных терпенов макромицетов, ядовитые свойства которых значимы для человека в различной степени. Цитотоксичность одних служит препятствием дальнейших разработок в качестве антибактериальных и антифунгицидных средств. Примерами могут служить кринипеллины Кринипеллиса шероховатого (Crinipellis scabella (Alb. et Schwein.) Murrill (= Crinipellis stipitaria (Fr.) Pat.)), гемимицины Гемимицены кукушечной (Hemimycena cucullata (Pers.) Singer), стриатины Бокальчика полосатого (Cyathus striatus (Huds.) Willd.). Так, токсичность последних при внутрибрюшинном введении мышам варьирует от 60 до 110 мг/кг. К основным ограничениям применения других соединений данного класса в рамках химиотерапии относят индуцируемые ими специальные формы токсического процесса. Примером могут служить проонкогенные в тесте Эймса цитотоксиканты маразминовая кислота (ЛД50 = = 15–30 мг/кг, мыши в/бр) и мерлидиалы, первоначально выделенные из Беоспоры мышехвостой (Baeospora myosura (Fr.) Singer) и Флебии дрожащей (Phlebia tremellosa (Schrad.) Nakasone ex Burds.) соответственно (Sterner, Anke, 1995). Значимые для клинических токсикологов терпены грибов представлены в группах олигоизопреноидов, сескви- и тритерпенов, а также трихотеценов.
Олигоизопреноиды
В рассмотренных выше G. spectabilis и G. junonius, которые вызывают зрительные галлюцинации, наличие псилоцибина было подтверждено только в базидиомах, собранных в Европе и Северной Америке (Hatfield, Valdes, 1978). Вместе с тем, в азиатских популяциях макромицетов (вид G. junonius широко известен в Японии как “оhwaraitake” – гриб большого смеха) триптамины не выявляют (Kayano et al., 2014; Lee et al., 2020).
Установлено, что G. spectabilis и G. junonius содержат в качестве минорных компонентов 3 вида олигоизопреноидов: гимнопилины (11), гимнопренолы (12) и гимнопилены (13). Структурно перечисленные соединения можно охарактеризовать как ациклические полиоксигенированные непредельные жирные кислоты и спирты, содержащие от 9 до 12 изопреновых звеньев (рис. 3). Хотя известные механизмы действия олигоизопреноидов Gymnopilus на нервную систему млекопитающих не могут полностью объяснить возникновение неконтролируемого смеха, галлюциногенное действие азиатских популяций ассоциируют именно с ними (Findlay, He, 1991; Kim et al., 2012).
Рис. 3.
Химическая структура олигоизопреноидов (11–13) и механизм образования иллюдинами белково-нуклеиновых аддуктов на примере процесса алкилирования иллюдина С (14) (адаптировано по: Yin et al., 2019).

Ранее, с этими соединениями связывали горький вкус представителей рода Gymnopilus. В дальнейшем оказалось, что гимнопилины способны преодолевать гематоэнцефалический барьер и проявлять цитотоксические свойства в отношении нервных клеток. При оценке посттетанического ответа доказана их способность к деполяризации двигательных нейронов переднего рога спинного мозга (Tanaka et al., 1993). Выявлено, что гимнопилины в концентрации IC50 = 30 мкмоль/л активируют везикулярный транспорт Ca2+ через систему Gq–фосфолипаза C. В мотонейронах первого порядка, мантийных глиоцитах и леммоцитах спинального ганглия крыс этот механизм приводит к повышению концентрации иона Ca2+ в цитоплазме (Miyazaki et al., 2012). В данном исследовании также отмечено, что олигоизопреноиды активируют цАМФ-зависимый каскад. На клетках феохромоцитомы РС12 показано, что помимо мобилизации внутриклеточного пула Ca2+, гимнопилины ингибируют работу никотинчувствительных ацетилхолиновых рецепторов в ответ на введение полных агонистов – IC50 = 15 мкмоль/л. При этом в зависимости от структуры активность соединений с m = 2 увеличивается в следующем порядке: n = 7 < n = 6 < n = 5 (Kayano et al., 2014).
Помимо центрального действия, олигоизопреноиды обладают выраженными гемодинамическими эффектами. Установлено, что на изолированных препаратах аорты крыс сумма гимнопиллинов в концентрациях более 0.1 мг/кг оказывает вазодилатирующее действие. На организменном уровне (анестезированные крысы) они проявляют положительный инотропный и отрицательный хронотропный эффекты. Кроме этого, их внутривенное введение в дозах 5 и 50 мг/кг повышает среднее артериальное давление в 1.4 и 1.7 раз соответственно. С учетом кинетики и содержания токсина в G. spectabilis – 5 мг/г сухой массы, полагают, что его уровни в плазме будут достигать гипертензивных концентраций при потреблении человеком массой 60 кг 30 г высушенных плодовых тел (Nishio et al., 2012).
Таким образом, к настоящему времени показано, что гимнопилины и родственные им соединения обладают центральным н-холинолитическим действием и гипертензивным эффектом. Тем не менее, выясненных механизмов недостаточно для полноценного суждения о психодислептическом процессе, который ассоциирован с псилоцибин-негативными популяциями Gymnopilus. Поэтому их дальнейшее изучение остается открытой проблемой (Miyazaki et al., 2012).
Сесквитерпены
В странах Океании, Юго-Восточной Азии и Северной Америки к значимым в токсикологическом отношении относят несколько видов рода Omphalotus: Омфалот иллюдовый (Omphalotus illudens (Schwein.) Bresinsky et Besl), О. гуепиниобразный (O. guepiniiformis (Berk.) Neda) и О. японский (O. japonicus (Kawam.) Kirchm. et O.K. Mill.) (French, Garrettson, 1988; Aoki et al., 2020; Dai et al., 2020). Интересно отметить, что перечисленные макромицеты обладают биолюменисцентными свойствами, поэтому в англоязычных изданиях в качестве их обиходного названия используют оборот “jack-o-lantern mushroom” – блуждающий огонек. Резиноидный синдром вследствие интоксикации данными видами ассоциирован с производными иллюдиновой кислоты: иллюдинами, иллюданами, протоиллюданами и иллюдаланами (Yokoyama, Gonmori, 2009; Beuhler, 2016). По всей видимости, обитающий в наших причерноморских краях О. масличный (O. olearius (DC.) Singer) также содержит токсины подобного рода.
Несмотря на относительно доброкачественное течение описанных интоксикаций, сеcквитерпены иллюдинового типа Omphalotus проявляют выраженную цитотоксичность в условиях резорбции. Полулетальная доза для крыс наиболее ядовитого среди них – иллюдина С (14) составляет 15.6 мг/кг при в/в и 30 мг/кг при в/бр введении (рис. 3). Цитотоксичность производных иллюдовой кислоты опосредована их алкилирующими свойствами при взаимодействии с ДНК и белками. Так, иллюдины М, S и C проявляют апоптотическую активность в отношении клеток рака легкого A549 – IC50 = 0.22, 0.069, 0.36 мкмоль/л соответственно (McMorris et al., 2003).
Механизмы резорбтивного действия иллюдинов изучены достаточно подробно в связи с тем, что в течение достаточно продолжительного времени их исследовали в рамках химиотерапевтических средств. Так, при помощи модификации иллюдина S получен препарат ирофульвен. В ходе клинических испытаний ирофульвен в отношении злокачественных новообразований яичников и немелкоклеточного рака легких оказался более эффективен в сравнении с такими известными соединениями как митомицин, цисплатин и паклитаксел (McMorris et al., 1990).
Иллюдины в организме млекопитающих подвержены реакции токсификации в ходе окисления простагландинредуктазой PTGR1 (КФ 1.3.1.48) и НАДФ-зависимой алкенальной оксидоредуктазой (КФ 1.3.1.74). Образуемый активный метаболит легко реагирует в бесклеточных системах при физиологическом диапазоне pH с аденином и гуанином ДНК, тиоловыми нуклеофилами, такими как серосодержащие пептиды и амины. Среди особенностей иллюдинов следует обратить внимание на наличие двух различных по реакционным предпочтениям участков. Наличие этих центров может приводить к связыванию одной молекулой токсина нуклеотида и белка в непосредственной близости (Le et al., 2019). При этом внутримолекулярная перегруппировка электронов способствует образованию двойных нуклеин-белковых аддуктов, которые отличает большая стабильность в сравнении с соединениями токсина с одной из молекул (рис. 3).
В процессе алкилирования серосодержащих молекул высока вероятность ингибирования тиолсодержащих ферментов. Считают, что алкогольдегидрогеназа, глутатионредуктаза и рибонуклеозидтрифосфатредуктаза наиболее чувствительны к иллюдинам (Le et al., 2019). Заслуживает внимания нарушение токсинами работы алкогольдегидрогеназы I типа, которое ведет к снижению синтеза оксида азота (II). Так, иллюдин С ингибирует в активированных бактериальным липополисахаридом макрофагах продукцию NO со значением EC50 = 31.4 мкмоль/л, что отчасти объясняет его противовоспалительные эффекты (Chen, Liu, 2017).
Образуя карбонильные аддукты с сульфидными группами цистеина и глутатиона, иллюдины также выступают в качестве прооксидантов. Перечисленные биологически активные молекулы, наряду с ингибируемыми ферментными системами, играют ключевую роль в поддержании антиоксидантного и кислотно-основного равновесия в тканях (McMorris et al., 1990). Для некоторых разветвленных иллюдинов и иллюданов этот процесс можно рассматривать как токсификацию, поскольку в ходе него образуется α-, β-ненасыщенный кетон. Последний обладает высокой реакционной способностью и ведет к вторичному повреждению (Yin et al., 2019).
За последние 10 лет список сеcквитерпенов на основе иллюдиновой кислоты был расширен за пределы рода Omphalotus в рамках концепции “drug discovery”. Они выявлены среди макромицетов рода Coprinopsis: иллюдины I, I2 и J2 – из культур Копринопсиса епископального (Coprinopsis episcopalis (P.D. Orton) Redhead, Vilgalys et Moncalvo), копринастатин и илюдан-илюдановые димеры бовистолы – из плодовых тел К. Строссмайера (C. strossmayeri (Schulzer) Redhead, Vilgalys et Moncalvo), дигидрогранулоинден – из культур К. пепельного (C. cinerea (Schaeff.) Redhead, Vilgalys et Moncalvo) (Aoki et al., 2020; Dai et al., 2020). Терпены иллюданового типа – эхинолактоны выделяют из культуральной жидкости афиллофороидного гриба Эхинодентиеллума японского (Echinodontiellum japonicum (Imazeki) S.H. He et Nakasone) (Chen, Liu, 2017). По всей видимости, токсикологическое значение перечисленных видов ограничено отсутствием у них пищевой ценности и своеобразным габитусом.
Тритерпены
Тритерпены и их производные состоят из 6 изопреновых единиц и представлены моно-, ди-, три-, тетра- и пентациклическими углеводородами. К настоящему времени из высших грибов выделены соединения со скелетами ланостана, эргостана, кукурбитана и сапонацеолида. Ядовитые полициклические тритерпены со скелетом ланостана встречают среди макромицетов рода Hypholoma и Hebeloma, а сапонацеолида – среди представителей рода Tricholoma (рис. 4).
Рис. 4.
Химическая структура ядовитых тритерпенов со скелетами ланостана (15–20) и сапонацеолида (21, 22) (адаптировано по: Fujimoto et al., 1992; Yin et al., 2014).

Отравления Опенком серно-желтым (Hypholoma fasciculare (Huds.) P. Kumm.) и Ложноопенком кирпично-красным (H. lateritium (Schaeff.) P. Kumm.) приводят к обезвоживанию вследствие интенсивного гастроинтестинального синдрома. Помимо этого, их токсическое действие сопровождается парезами мышц и нарушениями зрения (Shi et al., 2011; Chuluunbaatar et al., 2019). Полагают, что токсины перечисленных видов могут быть одной из значимых причин гибели пациентов, которым был поставлен диагноз резиноидного синдрома (Yokoyama, Gonmori, 2009; Beuhler, 2016). В этой связи H. fasciculare и H. lateritium относят к смертельно ядовитым видам (Бадалян, 1998)
Токсичность сухого остатка водно-метанольного экстракта H. fasciculare составляет ЛД50 = 500 мг/кг, мыши в/бр (Suzuki et al., 1983), по другим данным – ЛД50 = 243.3 мг/кг, ЛД100 = 250 мг/кг, мыши в/бр (Бадалян, 1998). При этом наблюдают дозо-зависимые гастроинтестинальные, гемолитические и нейротоксические эффекты с различными латентными периодами. В пределах базидиом токсическими свойствами обладают ланостановые тритерпены фасцикулолы, сесквитерпен со скелетом кариофиллана наематолон, порообразующий протеин фасцикуларелизин (35.5 кДа). Производные фасцикулола и кариофиллана также обнаруживают в составе базидиом H. lateritium (Chuluunbaatar et al., 2019). В связи с тем, что фасцикуларелизин (ЛД50 = 0.5 мг/кг, крысы в/в) подвержен в желудке протеолизу и термолабилен, а полулетальная доза нематолона значительно превышает 225 мг/кг (мыши в/бр), ядовитые свойства перечисленных видов связывают исключительно с фасцикулолами (Sterner, Anke, 1995).
С момента первой идентификации в макромицетах тритерпенов со структурой фасцикулолов (Ikeda et al., 1977) их список был значительно расширен. В настоящем среди Hypholoma известны фасцикулолы A–М, фасцикуловые кислоты A–C, фасцикулароны А, B (Abraham, 2011; Yin et al., 2019). Вместе с тем, список ядовитых фасцикулолов остается прежним. Он ограничен изомерами E (15) – ЛД50 = 168 мг/кг, и F (16) – ЛД50 = 30–50 мг/кг, мыши в/бр (Suzuki et al., 1983), а также минорными изоформами H и I, которые проявляют цитотоксичные свойства в отношении клеток глиомы U87 и нефроцитов (Shi et al., 2011). В ходе моделирования интоксикаций гибель животных от фасцикулолов Е и F возникает в результате паралича дыхательной мускулатуры. При этом наблюдают дозозависимые гастроинтестинальные, гемолитические и нейротоксические эффекты с различными латентными периодами. Также в ходе морфологических наблюдений отмечают выраженные дистрофические изменения в печени и почках (Suzuki et al., 1983).
Помимо рассматриваемых токсинов, остальные фасцикулолы, фасцикулароны и фасцикуловые кислоты малоядовиты в отношении млекопитающих. Им приписывают роль ингибиторов роста растений. В этой связи остается актуальным вопрос выяснения механизма действия и ключевых структурных особенностей токсичных фасцикулолов E и F, придающих ядовитые свойства только им среди целой группы тритерпенов (Chen, Liu, 2017).
Среди ланостановых тритерпенов со структурой гебевинозида к ядовитым относят изоформы I (17), HS-A (18), -B (19) и -C (20). Токсичные гебевинозиды выделяют из базидиом Гебеломы винософиллумной (Hebeloma vinosophyllum Hongo) (Fujimoto et al., 1986) и Г. раздетой (H. spoliatum (Fr.) Gillet). Так, H. spoliatum содержит HS-A, -B и -C в количестве 1.1, 0.71 и 0.02% от сухой массы соответственно (Fujimoto et al., 1992). Помимо этого, HS-A присутствуют в Г. клейкой (H. crustuliniforme (Bull.) Quél.) и Г. горчичной (H. sinapizans (Paulet) Gillet), которые встречают и в нашей стране (Yin et al., 2019). Как правило, токсическое действие перечисленных видов ассоциировано только с резиноидным синдромом с коротким латентным периодом. Однако в ходе интоксикаций с летальным исходом описаны судороги с исходом в кому (Price, 1927; White et al., 2019).
Гебевинозиды содержат углеводный компонент и гликозидную связь, необходимые для их нейротоксичности. Поэтому их иногда рассматривают как гликозиды макромицетов (Chen, Liu, 2017). Экспериментально показано, что токсичность гебевинозидов I, HS-A, -B и -C варьирует мало и составляет ЛД50 = 100 мг/кг, мыши в/бр. Полагают, что механизм их действия опосредован нарушением возбудимости тканей за счет ингибирования Na+/K+-переносчика. Интоксикации веществами в полулетальных дозах сопровождаются депрессией мышечной силы и развитием атонической комы. Также соединения вызывают расслабление изолированной тонкой кишки, сокращенной путем обработки ацетилхолином или хлористым барием (Fujimoto et al., 1992).
Тритерпены позиционируют в качестве одной из возможных причин рабдомиолиза, наблюдаемого при отравлениях некоторыми грибами. Вследствие интоксикаций макромицетами выделяют ранний (3А) и отсроченный (3В) рабдомиолиз (White et al., 2019). В качестве этиологического фактора раннего рабдомиолиза, описанного на территории Азии, приводят Russula subnigricans Hongo, токсины которой в настоящее время точно не определены. Возможной причиной могут быть циклопропенкарбоновая кислота (ЛД50 = 2.5 мг/кг, мыши в/бр), которую встречают в базидиомах в виде циклопропилацетилкарнитина, и хлорированные фениловые эфиры руссофелины A–F (Yin et al., 2019).
В результате работы, выполненной группой китайских исследователей, была выявлена причина рабдомиолиза на европейской территории континента. В ходе исследования из неполярной фракции экстракта Рядовки землистой (Tricholoma terreum (Shaeff.) P. Kumm.) выделены шестнадцать индивидуальных соединений: терреолиды A–F и сапонацеолиды H–P. Среди них токсичностью обладают сапонацеолиды B (21) и М (22) – ЛД50 = 88.3 и 63.7 мг/кг, мыши в/бр соответственно. Содержание изоформ B и М в сухой массе базидиом T. terreum составляет 414 и 184 мг/кг соответственно, а количество всей суммы сапонацеолидов достигает 1.3 г/кг. Гибель животных при введении веществ в полулетальных дозах сопряжена с судорогами, морфологически наблюдают острое повреждение почек на фоне массивного рабдомиолиза (Yin et al., 2014).
Однако не у всех исследователей однозначное мнение относительно полученных результатов. Аргументируя тем, что T. terreum издавна относили к съедобным видам, считают, что модель интоксикации не отражает действительности, поскольку в эксперименте был использован термически не обработанный экстракт (Davoli et al., 2016). Вместе с тем, известно, что потребление сырых плодовых тел даже съедобных видов способно приводить к повышенным значениям креатинкиназы (Nieminen et al., 2005). Также Даволи с соавт. (Davoli et al., 2016) выражают сомнение насчет стабильности рассматриваемых тритерпенов. В ответ на указанные возражения в отношении результатов исследования, авторы последнего настаивают на валидности полученных данных (Yin et al., 2016).
Кроме этого, высказано предположение, что базидиомы T. terreum могли быть неверно идентифицированы. Вид T. terreum крайне вариабелен и обладает большой экологической пластичностью. Описано несколько видов рядовок, сходных с T. terreum, но отличающихся друг от друга наличием или отсутствием частного покрывала и различиями в строении пилеипеллиса: Рядовка байковая (Tricholoma gausapatum (Fr.) Quél.), Р. белоземлистая (T. leucoterreum Mariotto et Turetta) и Рядовка мышиная (T. myomyces (Pers.) J.E. Lange). Европейские микологи (Christensen, Heilmann-Clausen, 2013) считают, что вышеперечисленные таксоны составляют один вид. Вместе с тем, ядовитые свойства представителей, считавшихся ранее самостоятельными видами, могут различаться.
После выхода в свет работ Бедри с соавт. (Bedry et al., 2001; Bedry, Gromb, 2009), рабдомиолиз-ассоциированным видом также считают Р. зеленую (T. equestre (L.) P. Kumm. (= Tricholoma auratum Gillet)). В работах представлена серия клинических наблюдений, ассоциированных с T. equestre и произошедших в Аквитании (Франция) начиная с 1992 г. Серия включает 12 случаев, 3 из которых – с летальным исходом. Также с T. equestre связывают 3 случая в Польше (Chodorowski et al., 2003; Klimaszyk, Rzymski, 2018).
Ассоциируемые с T. equestre интоксикации сопровождает латентный период от 1 до 3 сут. Далее выявляют повышенную утомляемость, мышечную слабость и миалгии в области бедер. Слабость усиливается в течение последующих 4 сут, возникают скованность в ногах, миоглобинурия. В половине случаев токсическому действию T. equestre сопутствуют эритема лица, тошнота, которая не доходит до рвоты, и обильное потоотделение. Уровень креатинкиназы в плазме достигает 200 тыс. МЕ/л. В большинстве наблюдений разрешение симптомокомплекса происходит в течение 15 сут. Предвестниками неблагоприятного течения интоксикации среди погибших пациентов выступают нарастание одышки, лихорадки, почечной и сердечной недостаточности. Неблагоприятное течение интоксикации сопровождается гиперкалийемией и острым миокардитом (Chodorowski et al., 2003; Bedry, Gromb, 2009).
Опровергающим аргументом можно считать то, что в исследовании на людях никаких побочных эффектов T. equestre не обнаруживают (Klimaszyk, Rzymski, 2018). К достоинствам исследования следует отнести использование идентифицированных с помощью молекулярных инструментов образцов. Однако в данной работе токсичность оценивали после однократного потребления. В то же время, описанные случаи интоксикаций возникали вследствие нескольких эпизодов употребления в пищу в течение как минимум нескольких дней подряд.
На данный момент в нашей стране пока не выявлено ни одного случая рабдомиолиза, связанного с T. equestre. В этой связи не акцентируют внимание на ее ядовитые свойства и не исключают из съедобных видов (Вишневский, 2019).
С учетом четко отграниченных территорий, на которых возникают данные интоксикации, то есть эндемичности, не могут быть полностью исключены гипотезы идиосинкразических реакций, генетической предрасположенности и ядовитых паразитов грибов. Предполагают, что ответственные за рабдомиолиз плодовые тела могли быть заражены паразитирующими на грибах миксомицетами (микромицетами), которые продуцируют цитохалазин B – токсин, способный вызывать поражение мышечных волокон (Klimaszyk, Rzymski, 2018).
Трихотецены
Трихотецены традиционно считают токсинами микромицетов, однако их встречают и среди высших грибов. Исторически интерес к изучению трихотеценов предопределен их значением в качестве контаминантов пищи (Медицинская токсикология…, 2012), строительных материалов и систем кондиционирования воздуха (Smoragiewicz et al., 1993). Оказываемый трихотеценовыми микотоксинами токсический эффект сложен и зависит от конкретных точек приложения механизмов действия индивидуальных соединений, попавших в организм человека (Shifrin, Anderson, 1999; Wu et al., 2017).
Среди базидиомицетов трихотецены в качестве минорных метаболитов выявляют в плодовых телах G. junonius. Однако самостоятельное токсикологическое значение им не отводят (Lee et al., 2020). Токсикологически значимым продуцентом трихотеценов среди аскомицетов считают Подострему оленероговую (Podostroma cornu-damae (Pat.) Boedijn (≡ Trichoderma cornu-damae (Pat.) Z.X. Zhu et W.Y. Zhuang) сем. Hypocreaceae). Летальные отравления стромами этого вида описаны на территориях Австралии, Китая, Кореи, Японии и о. Ява (Saikaw et al., 2001; Kim et al., 2016; Choe et al., 2018). Вследствие выраженности токсического эффекта, некоторые исследователи считают P. cornu-damae самым ядовитым видом среди известных макромицетов (Beuhler, 2016).
Интоксикации стромами P. cornu-damae классифицируют как токсическое действие трихотеценов (4D). Этот вид считают достаточно редким в ареале обитания и хорошо дифференцируемым относительно представителей с пищевой ценностью. Вследствие этого, отравления ими встречают редко. Например, в Японии с 1983 г. по 2008 г. описано всего 13 случаев с 3 смертельными исходами (Lee et al., 2018). Токсическое действие P. cornu-damae ассоциируют с ошибочным сбором их стром вместо видов рода Cordyceps или примордиев плодовых тел Ganoderma lucidum, потребляемых населением в лечебных целях. Известны случаи криминальных отравлений (White et al., 2019).
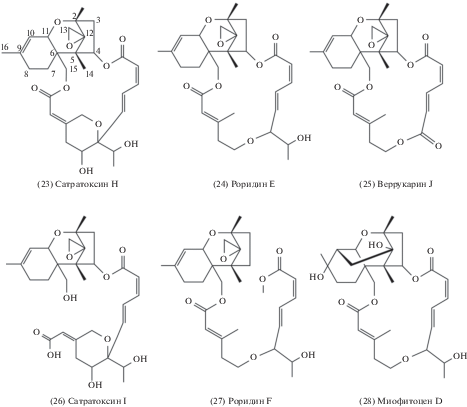
Выявляемые в стромах P. cornu-damae трихотецены проявляют выраженные ядовитые свойства: сатратоксин H (23) – ЛД50 = 0.5–1.23 мг/кг, крысы в/бр; роридин E (24) – ЛД50=10 мг/кг, мыши в/бр; ЛД50 = 55 мг/кг, мыши п/о; ЛД50 = 17.3 мг/кг – кролик п/о; веррукарин J (25) – ЛД50=0.5–0.7 мг/кг, мыши в/бр (The Merck index…, 2006; McCormick et al., 2011). Из чистых культур этого аскомицета также выделяют менее токсичные сатратоксин I (26), роридин F (27) и миофитоцен D (28) (Lee et al., 2018).
Токсины данного класса относят к поликетидам (Shifrin, Anderson, 1999), также их классифицируют как соединения сесквитерпенового ряда (Lee et al., 2020). Токсины P. cornu-damae относят к трихотеценам типа D, которые характеризует наличие эфирного макроцикла С4–С15 относительно 12,13-эпокситрихотец-9-енового ядра (рис. 5). Как правило, соединения подобной структуры более токсичны в сравнении с простыми трихотеценами типа А–С (McCormick et al., 2011). Необходимыми условиями их токсичности считают наличие двойной связи между атомами C9–C10, эпоксида при С12–С13-положении, а также макроцикла. Так, гидроксилирование C12 снижает токсичность ридина E более чем в 1000 раз, а незамкнутый макроцикл роридина F и сатратоксина I делает их неактивными в отношении линий раковых клеток (Zhu et al., 2020).
Рис. 5.
Химическая структура трихотеценов P. cornu-damae (адаптировано по: Choe et al., 2018; Lee et al., 2018).
Клинически отравления P. cornu-damae дублируют проявления микотоксикозов по типу алейкии. Описывают раздражающее действие аском на слизистую ротовой полости (Kim et al., 2016). Латентный период до начальных проявлений со стороны пищеварительной системы варьирует от 1 до 3 ч: отмечают тошноту, рвоту, диарею, повышенную температуру тела и обезвоживание (White et al., 2019). При отсроченной госпитализации последнее перетекает в гипотензию, олигурию и синдромы нарушенного сознания. Со стороны кожных покровов наблюдают мелкие кровоизлияния и петехиальную сыпь, пластинчатую десквамацию кожи ладоней, стоп, лица и алопецию. Тяжелые степени интоксикации характеризуют поражения всех органов и систем (Mogi et al., 2003).
Ключевые лабораторные изменения сопряжены с поражением костного мозга, нарушениями энергообмена, метаболизма, органоспецифической токсичности индивидуальных соединений. В ходе диагностических мероприятий наблюдают панцитопению, повышение трансаминаз, азотемию, изменения кислотно-основного и водно-электролитного балансов (Jang et al., 2013). Описаны случаи агранулоцитоза, рефрактерного к лечению гранулоцитарным колониестимулирующим фактором (Mogi et al., 2003). У пациентов в течение недели прогрессируют нарушения свертывающей системы крови, иммунитета – снижается устойчивость к инфекционным осложнениям. Вследствие этого они подвержены массивным кровотечениям и сепсису (White et al., 2019).
Описанные летальные исходы опосредованы множественной органной дисфункцией на фоне инфекционных и тромботических осложнений. В ходе аутопсии выявляют синдром диссеминированного внутрисосудистого свертывания, атрофию нервной ткани, жировое перерождение печени, почек, сердца (Saikaw et al., 2001; Mogi et al., 2003). Примечателен случай гибели пожилого мужчины вследствие геморрагического некроза легкого после потребления отваренных плодовых тел (Jang et al., 2013). Аналогичные проявления описаны при “синдроме больного здания” вследствие ингаляции трихотеценовых микотоксинов (Smoragiewicz et al., 1993). По всей видимости, в данном случае помимо перорального присутствовал ингаляционный путь поступления, который мог быть реализован в ходе термической обработки ядовитого гриба.
При проведении биологических проб очищенных субстанций роридина E, веррукарина J и сатратоксина H на грызунах обнаруживают изменения, аналогичные клиническим проявлениям. Течение интоксикации у животных характеризуют кахексия и геморрагический синдром. В месте введения токсинов возникают воспаление и раздражение. Как при внутрижелудочном, так и парентеральном введении у крыс и цыплят возникают эрозивно-язвенные изменения тонкого и толстого кишечника. Наблюдают транслокацию за пределы энтерогематического и лимфатического барьеров в норме непроникающих 14С-маннитола и условно-патогенной микрофлоры (Wu et al., 2017).
При интраназальной аппликации рассматриваемых соединений морфологически обнаруживают гибель обонятельных окончаний. При введении веществ в дозе ЛД50 прослеживают массивную гибель нейронов, в большей степени мозжечка (McCormick et al., 2011). Со стороны кроветворной системы основными мишенями выступают гематопоэтические предшественники и иммунокомпетентные клетки. Показано, что обнаруживаемые в плазме продукты расщепления ДНК происходят преимущественно из белых ростков крови и костного мозга. В результате выраженности повреждений пролиферирующих клеток костного мозга, селезенки, тимуса и лимфатических узлов трихотецены описывают как соединения с “радиомиметическим” эффектом (Smoragiewicz et al., 1993).
Трихотецены относят к ингибиторам синтеза белка и алкилирующим агентам. В частности, токсины типа D нарушают процесс инициирования трансляции, препятствуя образованию пептидной связи в пептидилтрансферазном центре 60S субъединицы рибосом (Shifrin, Anderson, 1999). Токсины оказывают множественное алкилирующее действие, образуя аддукты с ДНК, РНК и белками. Так, связывание тиоловых групп белков происходит преимущественно за счет 12,13-эпоксида в составе их ядра (McCormick et al., 2011). Поскольку введение различных органических меркаптанов не снижает их цитотоксичности, считают, что они проявляют избирательность в отношении лишь некоторых дисульфидных групп белков. Так как трихотецены не ингибируют ДНК- и РНК-полимеразы in vitro, подавление сборки нуклеиновых полимеров считают процессом вторичным в отношении снижения экспорта функционально-активных протеинов. Снижение ядерного экспорта белков и матричной РНК индуцирует через митоген-активируемую протеинкиназу опосредованные каспаза-3 и -9 апоптоз и аутофагию. Также развитию клеточной гибели способствует выраженный прооксидантный эффект токсинов, который, инициируя повреждение биологических мембран и ферментных систем, истощает клеточный ресурс репарации (Wu et al., 2017).
Вследствие перечисленных биохимических изменений также возникают снижение интенсивности всех видов обмена и специальные формы токсического процесса. Тем не менее, предпосылки для фатальных осложнений создает токсическое действие трихотеценов в отношении органов гемопоэза и иммунитета.
В зависимости от дозы, трихотецены могут быть как иммунодепрессантами, так и иммуностимуляторами. Высокие дозы токсинов приводят к быстрому началу вакуолизации лейкоцитов, в то время как низкие – способствуют экспрессии набора цитокинов in vitro и in vivo. Ряд авторов связывают это с дозозависимой направленностью модуляции работы трихотеценами внеклеточных сигнал-регулирующих киназ ERK1 и 2 (Shifrin, Anderson, 1999; McCormick et al., 2011). Ингибирование последних препятствует активации наивных T-клеток и опосредует изменение конформации F-актина макрофагов, тем самым снижая интенсивность фагоцитоза патогенных микроорганизмов. Снижение сопротивляемости вследствие иммунотоксичности трихотеценов показано на животных в отношении целого ряда патогенных и условно-патогенных вирусов, бактерий и микромицетов (Wu et al., 2017).
ЗАКЛЮЧЕНИЕ
Рассмотренные группы цитотоксичных и галлюциногенных безазотистых органических соединений неоднородны по своему составу. Однако их объединяет то, что они – этиологический фактор мицетизмов. Анализ литературы, посвященной безазотистым токсинам макромицетов, показывает, что данная область исследований, за редким исключением, остается малоосвоенной. Велик разрыв между перечнем соединений, ассоциируемых с конкретной серией интоксикаций, и списком изученных в токсикологическом отношении веществ. Вместе с тем, роль некоторых из них в формировании патологических процессов дискутабельна и требует пересмотра.
Системное исследование этих вопросов может послужить теоретической основой в разработке методов диагностики и средств терапии, а также направлений дальнейшего совершенствования оказания медицинской помощи при мицетизмах. Помимо этого, глубокое понимание процессов, реализуемых безазотистыми токсинами, представляет прочный фундамент для исследований веществ подобного рода в качестве фармакологических агентов.
Список литературы
Бадалян С.М. Биологические особенности некоторых базидиальных макромицетов (морфология, экология и физиологическая активность): Автореф. дис. … докт. биол. наук: Ереван: Институт ботаники НАН РА, 1998. 52 с.
Вишневский М.В. Ядовитые грибы России. М.: Проспект, 2019. 448 с.
Медицинская токсикология. Национальное руководство / Ред. Е.А. Лужников. М.: ГЭОТАР-Медиа, 2012. С. 748.
Магазов Р.Ш., Степанов А.В., Чепур С.В., Савельев А.П. Токсины биологического происхождения: природа, структура, биологические функции и диагностика / Ред. Р.Ш. Магазов. Уфа: Башкирская энциклопедия, 2019. 352 с.
Мусселиус С.Г., Рык А.А. Отравления грибами. М.: ООО Демиург-АРТ, 2002. 310 с.
Abo-Khatwa A.N., Al-Robai A.A., Al-Jawhari D.A. Lichen acids as uncouplers of oxidative phosphorylation of mouse liver mitochondria // Nat. Toxins. 1996. V. 4. № 2. P. 96–102.
Abraham W.-R. Bioactive sesquiterpenes produced by fungi are they useful for humans as well // Curr. Med. Chem. 2001. V. 8. № 6. P. 583–606.
Anke J, Razman I. Pharmacokinetic and pharmacodynamics drug interactions with kava (Piper methysticum Forst. F.) // J. Ethnopharmacol. 2004. V. 93. P. 153–160.
Aoki S., Aboshi T., Shiono Y. et al. Constituents of the fruiting body of poisonous mushroom Omphalotus japonicus // Chem. Pharm. Bull. (Tokyo). 2020. V. 68. № 5. P. 436–442.
Bauer S., Langner J., Rath F.W. et al. Zur toxizitat des Pilzinhaltsstoes Polyporsaure aus Hapalopilus rutilans // Tag. Berl. Akad. L. Wiss. 1990. B. 285. S. 179–182.
Bedry R., Baudrimont I., Deffieux G. et al. Wild-mushroom intoxication as a cause of rhabdomyolysis // N. Engl. J. Med. 2001. V. 343. P. 798–802.
Bedry R., Gromb S. Les intoxications particulières de la région Aquitaine // Rev. Med. Int. 2009. V. 30. № 7. P. 640–645.
Beuhler M.C. Overview of mushroom poisoning. Ch. 1 // Critical care toxicology / Eds J. Brent, K. Burkhart, P. Dargan et al. Cham: Springer Int. Publ., 2016. P. 1–26.
Brady L.R., Benedict R.G. Occurrence of bis-noryangonin in Pholiota squarroso-adiposa // J. Pharm. Sci. 1972. V. 61. P. 318.
Burton J.F., Cain B.F. Antileukemic activity of polyporic acid // Nature. 1959. V. 184. № 4695. P. 1326–1327.
Chen H.P., Liu J.K. Secondary metabolites from higher fungi // Progress in the chemistry of organic natural products / Eds A. Kinghorn, H. Falk, S. Gibbons, J. Kobayashi. Cham: Springer Int. Publ., 2017. P. 1−201.
Chodorowski Z., Anand J.S., Grass M. Acute poisoning with Tricholoma equestre of five-year old child // Przegl. Lek. 2003. V. 60. P. 309–310.
Choe S., In S., Jeon Y. et al. Identification of trichothecene-type mycotoxins in toxic mushroom Podostroma cornu-damae and biological specimens from a fatal case by LC–QTOF/MS // Foren. Sci. Int. 2018. V. 291. P. 234–244.
Christensen M., Heilmann-Clausen J. Fungi of Northern Europe. V. 4. The genus Tricholoma. Copenhagen: Dan. Mycol. Soc., 2013. 228 p.
Chuluunbaatar B., Beni Z., Dekany M. et al. Triterpenes from the mushroom Hypholoma lateritium: isolation, structure determination and investigation in bdelloid rotifer assays // Molecules. 2019. V. 24 (2). P. 301.
Dai Q., Zhang F.L., Du J.X. et al. Illudane sesquiterpenoids from edible mushroom Agrocybe salicacola and their bioactivities // ACS Omega. 2020. V. 5. № 34. P. 21961–21967.
Davoli P., Floriani M., Assisi F. et al. Comment on “Chemical and toxicological investigations of a previously unknown poisonous European mushroom Tricholoma terreum” // Chem. Eur. J. 2016. V. 22. № 16. P. 5786–5788.
Duncan C.J., Cuendet M., Fronczek F.R. et al. Chemical and biological investigation of the fungus Pulveroboletus ravenelii // J. Nat. Prod. 2003. V. 66. № 1. P. 103–107.
Fairbairn J., Steele M. Meconic acid and alkaloids in Papaver somniferum and P. bracteatum // Planta Medica. 1981. V. 41. № 1. P. 55–60.
Findlay J.A., He Z. Minor constituents of Gymnopilus spectabilis // J. Nat. Prod. 1991. V. 54. № 1. P. 184–189.
French A.L., Garrettson L.K. Poisoning with the North American jack o’lantern mushroom, Omphalotus illudens // J. Clin. Toxicol. 1988. V. 26. № 1–2. P. 81–88.
Fujimoto H., Suzuki K., Hagiwara H. et al. New toxic meta-bolites from a mushroom Hebeloma vinosophyllum. I. Structures of hebevinosides I, II, III, IV and V // Chem. Pharm. Bull. 1986. V. 34. № 1. P. 88–99.
Fujimoto H., Takano Y., Yamazaki M. et al. Isolation, identification and pharmacological studies on 3 toxic metabolites from a mushroom Hebeloma spoliatum // Chem. Pharm. Bull. 1992. V. 40. № 4. P. 869–872.
Gill M. Pigments of fungi (Macromycetes) // Nat. Prod. Rep. 2003. V. 20. № 6. P. 615–639.
Hatfield G.M., Brady L.R. Occurrence of bis-noryangonin in Gymnopilus spectabilis // J. Pharm. Sci. 1969. V. 58. № 10. P. 1298–1299.
Hatfield G.M., Valdes L.J. The occurrence of psilocybin in Gymnopilus species // Lloydia. 1978. V. 41. № 2. P. 140–144.
Ikeda M., Sato Y., Izawa M. et al. Isolation and structure of fasciculol A, a new plant growth inhibitor from Neamatoloma fasciculare // Agric. Biol. Chem. 1977. V. 41. № 8. P. 1539–1541.
Jafee B.D., Jones E.A., Loveless S.E., Chen S.F. The unique immunosuppressive activity of brequinar sodium // Transpl. Proc. 1993. V. 25. № 2. P. 19–22.
Jang J., Kim C.H., Yoo J.J. et al. An elderly man with fatal respiratory failure after eating a poisonous mushroom Podostroma cornu-damae // Tubercul. Resp. Dis. 2013. V. 75. № 6. P. 264–268.
Ju-Yeon K., Rhee I.-K., Lee K.-B. et al. Thelephoric acid and kynapcin-9 in mushroom Polyozellus multiflex inhibit prolyl endopeptidase in vitro // J. Microbiol. Biotechnol. 1999. V. 9. № 6. P. 798–803.
Kayano T., Kitamura N., Miyazaki S. et al. Gymnopilins, a product of a hallucinogenic mushroom, inhibit the nicotinic acetylcholine receptor // Toxicon. 2014. V. 81. P. 23–31.
Kim H.N., Do H.H., Seo J.S. et al. Two cases of incidental Podostroma cornu-damae poisoning // Clin. Exp. Emerg. Med. 2016. V. 30. № 3. P. 186–189.
Kim J.H., Lee C.H. Atromentin-induced apoptosis in human leukemia U937 cells // J. Microbiol. Biotechnol. 2009. V. 19. № 9. P. 946–950.
Kim K., Choi S., Lee K. Gymnopilin K: a new cytotoxic gymnopilin from Gymnopilus spectabilis // J. Antibiot. 2012. V. 65. P. 135–137.
Kletskov A.V., Potkin V.I., Kolesnik I.A. et al. Synthesis and biological activity of novel comenic acid derivatives containing isoxazole and isothiazole moieties // Nat. Prod. Comm. 2018. V. 13. № 11. P. 1507–1510.
Klimaszyk P., Rzymski P. The yellow knight fights back: toxicological, epidemiological, and survey studies defend edibility of Tricholoma equestre // Toxins (Basel). 2018. V. 10. № 11. P. 468.
Kraft J., Bauer S., Keilhoff G. et al. Biological effects of the dihydroorotate dehydrogenase inhibitor polyporic acid, a toxic constituent of the mushroom Hapalopilus rutilans, in rats and humans // Arch. Toxicol. 1998. V. 72. P. 711–721.
Le P., Nodwell M.B., Eirich J. et al. A chemical proteomic analysis of illudin-interacting proteins // Chemistry. 2019. V. 25. № 54. P. 12644–12651.
Lee I.K., Yun B.S. Styrylpyrone-class compounds from medicinal fungi Phellinus and Inonotus spp., and their medicinal importance // J. Antibiot. 2011. V. 64. P. 349–359.
Lee S., Ryoo R., Choi J.H. et al. Trichothecene and tremulane sesquiterpenes from a hallucinogenic mushroom Gymnopilus junonius and their cytotoxicity // Arch. Pharm. Res. 2020. V. 43. P. 214–223.
Lee S.R., Seok S., Ryoo R. et al. Macrocyclic trichothecene mycotoxins from a deadly poisonous mushroom, Podostroma cornu-damae // J. Nat. Prod. 2018. V. 82. № 1. P. 122–128.
Li W., Li X.-B., Lou H.-X. Structural and biological diversity of natural p-terphenyls // J. Asian Nat. Prod. Res. 2017. V. 20. № 1. P. 1–13.
McCormick S.P., Stanley A.M., Stover N.A. et al. Tricho-thecenes: from simple to complex mycotoxins // Toxins. 2011. V. 3. № 7. P. 802–814.
McMorris T.C., Cong Q., Kelner M.J. Structure-activity relationship studies of illudins: analogues possessing a spiro-cyclobutane ring // J. Org. Chem. 2003. V. 68. № 25. P. 9648–9653.
McMorris T.C., Kelner M.J., Wang W. et al. On the mechanism of toxicity of illudins: the role of glutathione // Chem. Res. Toxicol. 1990. V. 3. № 6. P. 574–579.
Miyazaki S., Kitamura N., Nishio A. et al. Gymnopilin – a substance produced by the hallucinogenic mushroom, Gymnopilus junonius – mobilizes intracellular Ca2+ in dorsal root ganglion cells // Biomed. Res. 2012. V. 33. № 2. P. 111–118.
Mogi K., Takeshita H., Yasuda T. et al. Case report: food poisoning to death by Podostroma cornu-damae, its case history and autopsy findings // Acta Criminol. Med. Leg. Jpn. 2003. V. 69. P. 14–20.
Nieminen P., Mustonen A.-M., Kirsi M. Increased plasma creatine kinase activities triggered by edible wild mushrooms // Food Chem. Tox. 2005. V. 43. № 1. P. 133–138.
Nishio A., Kitamura N., Tanaka S. et al. Multiple effects of gymnopilin on circulatory system of the rat // Biol. Pharm. Bull. 2012. V. 35. № 8. P. 1300–1305.
Price H.W. Mushroom poisoning due to Hebeloma crustuliniforme // Arch. Dis. Child. 1927. V. 34. № 3. P. 441.
Saikaw Y., Okamoto H., Inui T. et al. Toxic principles of a poisonous mushroom Podostroma cornu-damae // Tetrahedron. 2001. V. 57. № 39. P. 8277–8281.
Sano Y., Sayama K., Arimoto Y. et al. Ustalic acid as a toxin and related compounds from the mushroom Tricholoma ustale // Chem. Comm. 2002. V. 13. P. 1384–1385.
arris J., La Porte I., Schweitzer I. Kava: a comprehensive review of efficacy, safety, and psychopharmacology // Aust. N. Z. J. Psych. 2011. V. 45. P. 27–35
Sawayama Y., Tsujimoto T., Sugino K. et al. Syntheses of naturally occurring terphenyls and related compounds // Biosci. Biotech. Biochem. 2006. V. 70. № 12. P. 2998–3003.
Schüffler A. Secondary metabolites of basidiomycetes // The physiology and genetics. 2nd Edition / Eds T. Anke, A. Schüffler. N.Y.: Springer Int. Publ., 2018. P. 231–275.
Shi X.W., Li X.J., Gao J.M. et al. Fasciculols H and I, two lanostane derivatives from Chinese mushroom Naematoloma fasciculare // Chem. Biodiv. 2011. V. 8. № 10. P. 1864–1870.
Shifrin V.I., Anderson P. Trichothecene mycotoxins trigger a ribotoxic stress response that activates c-jun n-terminal kinase and p38 mitogen-activated protein kinase and induces apoptosis // J. Biol. Chem. 1999. V. 274. № 20. P. 13985–13992.
Shulgin A.T. The narcotic pepper – the chemistry and pharmacology of 'Piper methysticum' and related species // Bull. Narc. 1973. V. 25. № 2. P. 59–74.
Smoragiewicz W., Cossette B., Boutard A. et al. Tricho-thecene mycotoxins in the dust of ventilation systems in office buildings // Intern. Arch. Occup. Environ. Health. 1993. V. 65. № 2. P. 113–117.
Stephenson N.L., Rundel P.W. Quantitative variation and the ecological role of vulpinic acid and atranorin in thallus of Letharia vulpina // Biochem. Syst. Ecol. 1979. V. 7. P. 263–267.
Sterner O., Anke H. Toxic terpenoids isolated from higher fungi // Czech. Mycol. 1995. V. 4. № 8. P. 39–52.
Suzuki K., Fujimoto H., Yamazaki M. The toxic principles of Naematoloma fasciculare // Chem. Pharm. Bull. 1983. V. 31. № 6. P. 2176–2178.
Tanaka M., Hashimoto K., Okuno T. et al. Neurotoxic oligoisoprenoids of the hallucinogenic mushroom, Gymnopilus spectabilis // Phytochem. 1993. V. 34. № 3. P. 661–664.
The Merck index. An encyclopedia of chemicals, drugs, and biologicals. 14th Edition / Eds M.J. O’Neil, P.E. Heckelman, C.B. Koch, K.J. Roman. Hoboken, N.J.: Merck, John Wiley & Sons, Inc., 2006. 2564 p.
Villa A.F., Saviuc P., Langrand J. et al. Tender nesting polypore (Hapalopilus rutilans) poisoning: report of two cases // Clin. Toxicol. 2013. V. 51. № 8. P. 798–800.
White C.M. The pharmacology, pharmacokinetics, efficacy and adverse events associated with kava // J. Clin. Pharmacol. 2018. V. 58. № 11. P. 1396–1405.
White J., Weinstein S.A., De Haro L. et al. Mushroom poisoning: a proposed new clinical classification // Toxicon. 2019. V. 157. P. 53–65.
Wu Q., Wang X., Nepovimova E. et al. Trichothecenes: immunomodulatory effects, mechanisms, and anti-cancer potential // Arch. Toxicol. 2017. V. 91. № 12. P. 3737–3785.
Yin X., Feng T., Li Z.H. et al. Response to the “Comment on chemical and toxicological investigations of a previously unknown poisonous European mushroom Tricholoma terreum” // Chem. Eur. J. 2016. V. 22. № 16. P. 5789–5792.
Yin X., Feng T., Shang J.H. et al. Chemical and toxicological investigations of a previously unknown poisonous European mushroom Tricholoma terreum // Chem. Eur. J. 2014. V. 20. № 23. P. 7001–7009.
Yin X., Yang, A., Jin-Ming G. Mushroom toxins: chemistry and toxicology // J. Agric. Food Chem. 2019. V. 18. № 67. P. 5053–5071.
Yokoyama K., Gonmori K. Increase of poisoning by tropical mushrooms in Japan in recent years // Chudoku Kenkyu. 2009. V. 22. № 3. P. 240–248.
Yoshioka N., Hayakawa I., Minatani T. et al. Quantitative analysis of the Tricholoma ustale-derived toxin, ustalic acid, in mushroom and food samples by LC-MS/MS // Forensic Sci. Int. 2020. V. 317. P. 110554.
Zhou Z.Y., Liu J.K. Pigments of fungi (macromycetes) // Nat. Prod. Rep. 2010. V. 27. № 11. P. 1531–1570.
Zhu M., Cen Y., Ye W. et al. Recent advances on macrocyclic trichothecenes, their bioactivities and biosynthetic pathway // Toxins (Basel). 2020. V. 12. № 6. P. 417.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии