

Успехи современной биологии, 2022, T. 142, № 1, стр. 52-69
Повреждения, адаптации, стратегии древесных видов в условиях техногенеза: структурно-функциональные уровни реализации адаптивного потенциала
Р. В. Уразгильдин 1, *, А. Ю. Кулагин 1
1 Уфимский Институт биологии – обособленное структурное подразделение Уфимского федерального исследовательского центра РАН
Уфа, Россия
* E-mail: urv@anrb.ru
Поступила в редакцию 05.08.2021
После доработки 30.08.2021
Принята к публикации 30.08.2021
- EDN: QYTXAZ
- DOI: 10.31857/S0042132422010082
Аннотация
Обзор посвящен систематизации работ в области адаптивных реакций древесных растений к техногенезу на основных иерархических структурно-функциональных уровнях организации и в области адаптивных стратегий. Рассмотрено влияние различных типов промышленного загрязнения на микро-, макроморфологию и физиологические реакции хвои и листьев, радиальный прирост стволовой древесины, корневые системы, жизненное состояние древостоев, а также рассмотрены вопросы определения адаптивных реакций и адаптивных стратегий древесных видов к техногенезу. Показана относительная независимость адаптивных реакций в пределах органов, выделены специфические и неспецифические реакции, обсуждаются причины данных поливариантных реакций, лежащие в основе адаптивного потенциала, принципа множественного обеспечения биологически необходимых функций для сохранения гомеостаза, толерантности видов к техногенезу. Рассмотрены вопросы устойчивости биосистем к техногенезу, в основе которой лежат адаптивные реакции, протекающие на всех уровнях организации живого – от цитогенетического до экосистемного. Промышленное загрязнение, как новый для растений в историческом плане фактор, вызывает необходимость разработки вопросов, связанных с адаптивными стратегиями видов к техногенезу, в основе которых лежат адаптивный потенциал, изменчивость, устойчивость и экологическая пластичность видов.
ВВЕДЕНИЕ
Микро- и макроморфология хвои и листьев, их физиологические показатели (водный обмен и пигментный комплекс), радиальный прирост стволовой древесины, состояние и развитие корневых систем, жизненное состояние древостоев очень часто используются для мониторинга и индикации повреждения лесных экосистем промышленными загрязнителями. Анализ литературных данных, посвященных адаптациям растений к техногенезу, позволил выделить ряд наиболее общих специфических адаптивных реакций, принятых в дендроэкологии как “классические” при усилении степени промышленного загрязнения:
– уменьшение линейных размеров, площади и массы листа/хвои, размеров устьиц, толщины губчатой паренхимы; увеличение устьичной плотности, плотности жилкования, толщины листа и всех тканей (кроме губчатой паренхимы) и др.;
– уменьшение интенсивности транспирации (как показатель уменьшения вентилируемости внутренних полостей листа для уменьшения проникновения токсикантов) при кратковременной и незначительной фумигации токсикантами, а при хронической фумигации – значительное усиление интенсивности транспирации (потеря возможности контролировать транспирационный процесс); увеличение относительного содержания воды в листе (как результат уменьшения интенсивности транспирации и увеличения количества физиологически связанной воды); уменьшение дефицита водного насыщения листа (как результат уменьшения интенсивности транспирации и увеличения относительного содержания воды);
– уменьшение содержания хлорофилла а и компенсаторное увеличение хлорофилла b и каротиноидов; уменьшение соотношения хлорофилл а/хлорофилл b и увеличение соотношения (хлорофилл а + хлорофилл b)/каротиноиды;
– уменьшение величины радиального прироста, потеря его чувствительности к климатическим сигналам, перераспределение долей ранней и поздней древесины в общем приросте, изменение длительности онтогенетических периодов и нарушение цикличности прироста, появление повышенного количества ложных годичных колец или их выпадение, ускоренное начало старения древостоев и др;
– подавление роста и развития корневых систем;
– ухудшение жизненного состояния древостоев.
Таким образом, наиболее общей и частой реакцией для всех видов в дендроэкологии считается усиление ксероморфности вегетативных органов, подавление физиологических процессов, роста, развития и жизненного состояния деревьев. С другой стороны, различные промышленные загрязнители по-разному влияют на древесные виды и ответные реакции могут быть как специфическими, так и не специфическими и нейтральными, что будет показано в данном обзоре. Такое многообразие и неоднородность адаптивных реакций выступают основой для разработки положений об адаптивном потенциале и адаптивных стратегиях древесных видов к техногенезу, что будет рассмотрено в заключительной части обзора.
АДАПТИВНЫЕ РЕАКЦИИ
Каждая биосистема для существования в природе должна противопоставить воздействию внешней среды достаточное разнообразие соответствующих реакций. Принцип множественного обеспечения биологически необходимых функций является чрезвычайно важным для увеличения уровня гомеостаза (Маслов, 1980). Чем большее число механизмов адаптации используется растением одновременно на самых разных уровнях, тем более устойчив организм к действию токсических ингредиентов (Тарабрин, 1984). Не каждый стресс обязательно негативен для деревьев, но может вместо этого вызвать повышенную устойчивость к стрессу. Кратковременная стрессовая реакция может не совпадать с длительным изменением жизненного состояния деревьев, поэтому изменения должны интерпретироваться с долгосрочной точки зрения (Kozlowski, Pallardi, 2002).
Адаптивные реакции, возникающие в ответ на действие факторов окружающей среды, характеризуют изменчивость метаболизма и устойчивость растений в экстремальных условиях. Адаптационные процессы протекают на всех уровнях организации живого – от цитогенетического до экосистемного. На уровне организма механизмы адаптации, свойственные клетке, дополняются новыми, отражающими взаимодействие органов в целом растении. В процессе жизнедеятельности биометрические параметры испытывают колебания, определяемые как внутренними, так и внешними факторами. Регуляторные гомеостатические механизмы обеспечивают низкий уровень этих колебаний. При воздействиях, превышающих границы толерантной зоны, в биосистеме развивается комплекс физиологических и биохимических изменений. Комплекс изменений, происходящих в живой системе, отражает ее переход из состояния, поддерживаемого гомеостатическими механизмами, в новое квазистационарное стрессовое состояние. Биотипическая, возрастная и модификационная неоднородность популяций выступает в качестве адаптивного полиморфизма на разных уровнях организации. Важнейшими механизмами являются изменения популяционной структуры вида, в результате которых большее представительство в популяции приобретают особи, обладающие наибольшей резистентностью к действию конкретного токсического фактора (Кулагин, 1985; Веселова и др., 1993; Пахомова, 1995).
Вопросам изучения адаптаций растений, в том числе и древесных, к техногенному загрязнению посвящена обширная литература. Выделен целый комплекс адаптаций к техногенным воздействиям, начиная с биохимических, клеточных, анатомических и далее до экосистемных (Кулагин, 1985). Древесные растения имеют специальные механизмы для преодоления повреждений, вызванных стрессами окружающей среды. Но ситуация осложняется, когда возникают множественные стрессы. Механизмы, принятые против одного фактора стресса, не обязательно аналогичны механизмам противодействия множественным стрессам. Реакция растения может отличаться, когда два или более стресса возникают последовательно или одновременно (Chelli-Chaaboimi, 2013). Биотические и абиотические стрессы влияют на рост, развитие и продуктивность растений. Чтобы справиться с этими стрессами, растения “разрабатывают” определенные эффективные стратегии, которые позволяют им адаптироваться – избежать или терпеть стрессы. Такие стратегии адаптации находятся на морфологическом, анатомическом, биохимическом и молекулярном уровнях (Devi et al., 2017).
При воздействии экологических стрессов древесные растения должны перераспределять энергию таким образом, чтобы обеспечить адаптацию к стрессу, но также поддерживать рост и производительность. Для достижения этих жизненно важных целей растение реагирует активацией многих метаболических процессов, контролирующих фотосинтез, ионный гомеостаз и сигнализацию гормонов растений, которые могут изменять экспрессию генов. Эти реакции обычно экспрессируются как на фенотипическом, так и на генотипическом уровне (Skirycz, Inzé, 2010). В результате на различных структурно-функциональных уровнях организации растительного организма возникает множество адаптивных реакций, направленность которых – специфические реакции, неспецифические, нейтральные (отсутствие реакции) – зависит от адаптивного потенциала растения. Далее проведем анализ публикаций, посвященных адаптивным реакциям древесных растений на основных структурно-функциональных уровнях иерархической организации с выделением направленности этих реакций.
Влияние техногенеза на макро- и микроморфологию хвои/листа
Основная масса исследований указывает на значительное уменьшение макроморфологических параметров ассимиляционного аппарата древесных растений в условиях техногенеза. Так, серия публикаций указывает на проявление тех или иных изменений, как то уменьшение длины, ширины, площади, массы ассимиляционных органов, их количества на побегах (в частности плотности охвоения побегов и возраста хвои) длины побегов и др. Например, показано, что в условиях высокого автотрафика г. Кветта (Пакистан) наблюдается значительное снижение длины, ширины, площади листа и длины черешка у Elaeagnus angustifolia L., Eucalyptus tereticornis L., Ficus carica L., Fraxinus excelsior L., Melia azadirach L., Morus alba L., Morus nigra L., Fistacia vera L., Prunus armeniaca L., Punica granatum L., Robinia pseudoacacia L., Rosa indica L. и Vitis vinifera L., причем значительные различия выявлены в зависимости от сезона: максимальное снижение обнаружено летом, среднее осенью, а наименьшее – весной. По мере старения растений уменьшение морфологических параметров возрастает (Leghari, Asrar, 2013). В условиях атмосферного загрязнения Иркутско-Черемховского промышленного центра нарушение питания почвенными элементами (содержание S, F, Fe, Al, Pb, Cd, Hg, N, Ca, Mg в хвое увеличивается, а содержание P, K, Mn уменьшается) приводит к снижению параметров роста Pinus sylvestris L. (длины и массы хвои, длины побегов, количества хвоинок на побегах), а впоследствии приводит к системному нарушению функционирования организма растений (Mikhailova et al., 2017). С использованием камер для фумигации выявлено, что с увеличением концентрации O3 у Populus nigra L. и P. euramericana L. значительно снижается фотосинтетическая биомасса из-за образования меньшего количества новых листьев, уменьшения их размера и преждевременного их опадения (Schreuder et al., 2001). Аналогичные результаты показаны во многих отечественных и зарубежных публикациях (Баканов, 1997; Неверова, Колмогорова, 2002; Сухарева, Лукина, 2004; Бойко, 2005; Кулагин, Зайцев, 2008; Сейдафаров, 2009; Лукина, 2011; Бикмуллин, 2012; Аминева, 2016; Василевская, Лебедевич, 2016; Донцов и др., 2016; Василевская, Сидорчук, 2018; Jahan, Iqbal, 1992; Seyyednejad et al., 2009b, 2011; Seyyednejad, Koochak, 2013; Aribal et al., 2016; Qadir et al., 2016; Swain et al., 2016; Areington et al., 2017; Rostunov et al., 2017; Allahnouri et al., 2018; Chaudhary, Rathore, 2019; Uka, Belford, 2020). Также показано, что степень угнетения морфологических параметров зависит от периода вегетации (Лукина, 2011; Василевская, Сидорчук, 2018), времени года (Leghari, Asrar, 2013; Areington et al., 2017), стадии дегрессии древостоя (Сухарева, Лукина, 2004), а в некоторых случаях наблюдается нелинейная изменчивость морфологических параметров и интенсивный преждевременный листопад (Сухарева, Лукина, 2004; Бикмуллин, 2012).
Но в то же время, у многих древесных видов в условиях промышленного загрязнения в отношении морфологических параметров могут наблюдаться неспецифические адаптивные реакции, то есть увеличение всех или только некоторых морфологических параметров, причем данный факт, как правило, зависит от типа промышленного загрязнения. Например, показано достоверное удлинение годичных побегов у Pecea obovata Ledeb., Sorbus sibirica Hedl., Betula pendula Roth в условиях г. Кемерово (Легощина, 2016), Salix caprea L., Sorbus aucuparia L., Malus baccata (L.) Borkh., Populus balsamifera L. и Betula pendula Roth в условиях г. Ижевска (Бухарина и др., 2007а), Pinus sylvestris L. в условиях Казанского промышленного центра (Бикмуллин, 2012) и др. (Турмухаметова, 2005; Зайцев, Кулагин, 2006; Скотников, 2007; Кулагин, Зайцев, 2008; Аминева, 2016). Также отмечается увеличение длины, ширины, площади, массы ассимиляционных органов на примере Picea obovata Ledeb. в зоне загрязнения Сыктывкарского лесопромышленного комплекса (Тужилкина, Плюснина, 2014), Tilia cordata Mill. в условиях высоких концентраций металлов в почвах индустриальной территории провинции Нижняя Силезия (Польша) (Kosiba, 2008), Albizia lebbeck Benth. в условиях загрязнения от нефтехимических компаний в высокотемпературных районах Ирана (Seyyednejad et al., 2009a) и др. (Зайцев, Кулагин, 2006; Скотников, 2007; Бикмуллин, 2012; Jochner et al., 2015).
Довольно часто исследователи сталкиваются вообще с отсутствием реакций древесных видов на техногенное воздействие. Например, в древостоях Picea abies (L.) H. Karst. в зонах действия промышленных выбросов предприятий Московской области не установлено достоверных различий с контролем по длине, толщине хвои и ее массе в абсолютно сухом состоянии (Баканов, 1997), не выявлено отличий по длине хвои Picea obovata Ledeb. в магистральных посадках г. Красноярск (Донцов и др., 2016), отсутствуют изменения по массе листьев Tilia cordata Mill. в условиях углеводородного загрязнения Уфимского промышленного центра (Аминева, 2016), не обнаружено достоверных различий по длине одно- и двулетних побегов Pinus sylvestris L. в условиях Казанского промышленного центра (Бикмуллин, 2012) и др. (Тужилкина, Плюснина, 2014; McDermot et al., 2020).
В отношении микроморфологических характеристик ассимиляционного аппарата древесных растений в условиях техногенеза также наблюдается поливариантность адаптивных реакций. Многие исследователи указывают на значительную ксероморфизацию параметров, как, например, увеличение устьичной полности, но снижение устьичного индекса, размеров устьиц и устьичной щели (длина, ширина и площадь), количества эпидермальных клеток на единицу площади листа; увеличение толщины листа и толщины тканей листа, но уменьшение размеров клеток тканей листа и др. на примере: Lagerstroemia indica L., Callistemon citrinus (Curtis) Skeels в промышленной долине Катманду в центральной части Непала (Shrestha et al., 2021), Fraxinus excelsior L., Amaranthus viridis L., Cynodon dactylon L., Chenopodium album L., Robinia pseudoacacia L., Sophora mollis (Royle) Baker в столичном г. Кветта (Пакистан) (Laghari1 et al., 2015), Eugenia uniflora L. вдоль главной городской дороги в Рио-де-Жанейро (Бразилия) (Bezerra et al., 2020), Mangifera indica L., Albizia procera (Roxb.) Benth. и Swietenia mahagoni (L.) Jacq. в придорожных древостоях гг. Газипур и Мименсингх (Бангладеш) (Mitu et al., 2019), Fraxinus rotundifolia Mill. вдоль дороги с высоким трафиком в провинции Хамедан (Иран) (Allahnouri et al., 2018), Robinia pseudoacacia L., Ulmus minor “Umbraculifera” в условиях сильного автотранспортного загрязнения воздуха в Тегеране (Иран) (Rashidi et al., 2012; Abbasi et al., 2018). Во многих случаях указывается на значительное увеличение количества закупоренных устьиц (Laghari1 et al., 2015; Mitu et al., 2019; Uka, Belford, 2020; Shrestha et al., 2021).
Однако, у многих древесных видов могут наблюдаться неспецифические адаптивные реакции микроморфологических параметров в условиях промышленного загрязнения, направленные на ослабление ксероморфизации: снижение устьичной плотности, количества вспомогательных клеток, но увеличение размеров устьиц и устичных щелей, числа и размеров эпидермальных клеток и трихом; снижение толщины и площади поперечного сечения хвои/листа и толщины тканей, но увеличение размеров клеток тканей и др. Такие изменения можно наблюдать на примере: Callistemon citrinus (Curtis) Skeels, Lagerstroemia indica L. в промышленной долине Катманду (Непал) (Shrestha et al., 2021), Picea obovata Ledeb. в зоне загрязнения Сыктывкарского лесопромышленного комплекса (Тужилкина, Плюснина, 2014), Ficus platyphylla Del., Mangifera indica L., Polyalthia longifolia Sonn., Terminalia cattapa L. в метрополисе Кумаси (Гана) (Uka, Belford, 2020), Morus alba L. вдоль дороги с высоким трафиком движения в провинции Хамедан (Иран) (Allahnouri et al., 2018), Cinnamomum camphora (L.) J. Presl, Lawsonia inermis L. и Bougainvillea spectabilis Willd. в промышленном районе города Джидда (Саудовская Аравия) (Shaheen et al., 2016), Platanus orientalis L. в условиях автотранспортного загрязнения в мегаполисе Мешхед (Иран) (Pourkhabbaz et al., 2010).
Нередки случаи отсутствия реакций микроморфологических параметров на техногенное воздействие. Сравнение листьев деревьев Acer rubrum L. в лесах небольшого (Ньюарк) и крупного (Филадельфия) городов показало, что существенных различий в толщине листьев, верхнего эпидермиса, губчатой паренхимы и палисадной паренхимы не выявлено. Аналогично, на абаксиальной поверхности листьев не обнаружено различий в устьичной плотности, однако размеры устьиц (длина и ширина) значительно больше у кленов в лесах Филадельфии (McDermot et al., 2020). У листьев Eugenia uniflora L. вдоль главной городской дороги в Рио-де-Жанейро (Бразилия) площадь и удельная площадь листа, масса сырого и сухого листа, толщина и плотность сырого листа, толщина палисадной, губчатой паренхимы и абаксиального эпидермиса, суккулентность существенно не различались между загрязнением и контролем (Bezerra et al., 2020). Исследования листьев Platanus orientalis L. в условиях автотранспортного загрязнения в мегаполисе Мешхед (Иран) не выявили анатомических нарушений, длина, ширина и площадь проекции клеток палисадной паренхимы и губчатой паренхимы не различаются между загрязнением и контролем, однако верхняя кутикула значительно уменьшается (Pourkhabbaz et al., 2010).
Наибольший интерес вызывает относительная независимость адаптивных реакций в пределах одного органа: лист и хвоя, являясь едиными целостными органами, следуя логике, должны проявлять к одному и тому же стрессовому фактору однонаправленные адаптивные реакции, однако во многих исследованиях мы одновременно видим разнонаправленные специфические и неспецифические реакции в пределах одного органа. Анатомические исследования в индустриальном центре г. Карачи (Пакистан) выявили видоспецифические адаптивные перестройки в тканях листьев: у Ficus bengalensis L. уменьшаются верхняя кутикула, верхняя столбчатая паренхима и губчатая паренхима, но увеличиваются верхний эпидермис, нижняя столбчатая паренхима и нижний эпидермис, а нижняя кутикула остается без изменений; у Eucalyptus sp. уменьшаются верхняя кутикула, верхний эпидермис, верхняя столбчатая паренхима, губчатая паренхима и нижний эпидермис, но увеличиваются верхний и нижний гиподермис, нижняя столбчатая паренхима и нижняя кутикула; у Guaiacum officinale L. уменьшаются все анатомические параметры. Таким образом, больше всего страдают клетки столбчатой и губчатой паренхимы – они становятся “сплющенными” (Jahan, Iqbal, 1992). Аналогичные результаты показаны и другими авторами (Бикмуллин, 2012; Тужилкина, Плюснина, 2014; Донцов и др., 2016; Dineva, 2004, 2005, 2006; Pourkhabbaz et al., 2010; Rashidi et al., 2012; Shaheen et al., 2016; Allahnouri et al., 2018; Bezerra et al., 2020; Shrestha et al., 2021). Также обращает на себя внимание то, что во многих случаях одни морфологические параметры изменяются значительно и достоверно (или недостоверно), а другие – незначительно и недостоверно (Неверова, Колмогорова, 2002; Сухарева, Лукина, 2004; Легощина, 2016; Jahan, Iqbal, 1992; Dineva, 2004, 2005, 2006; Abbasi et al., 2018; Uka, Belford, 2020).
Влияние техногенеза на физиологию ассимиляционного аппарата
Водный обмен и пигментный комплекс ассимиляционного аппарата древесных растений характеризуются значительной чувствительностью к стрессовым факторам окружающей среды, поэтому многие исследования указывают на проявление специфических адаптивных реакций физиологических процессов в условиях техногенеза. Листья деревьев Lagerstroemia speciosa L., растущих вдоль обочины Национального шоссе в г. Дехрадун (Индия), характеризуются низкими скоростью ассимиляции углерода, интенсивностью транспирации, устьичной проводимостью и устьичным сопротивлением, но при этом наблюдается повышенная эффективность водопользования, а повышение дефицита давления пара и концентрации пролина в тканях листьев являются индикаторами стресса (Singh et al., 2017). Фумигация листьев Acer saccharinum L. раздельно Cd2+ и SO2 показала, что чистый фотосинтез и транспирация значительно снижаются в обоих случаях, а совместное воздействие токсикантов снижает их еще гораздо больше. Диффузионные сопротивления листьев к переносу диоксида углерода и водяного пара увеличиваются как с увеличением концентрации Cd2+, так и SO2 (Lamoreaux, Chaney 1978). Показано, что фумигация O3 значительно уменьшает углы контакта капель с листьями (увеличивает их смачиваемость) и увеличивает потери воды и проводимость водяного пара у древесных видов Populus nigra L., P. euramericana L. сорт Robusta и Pseudotsuga menziesii (Mirb.) Franco, что приводит к повреждению листьев и преждевременному их опаданию (Schreuder et al., 2001). У клена остролистного в условиях нефтехимического загрязнения Уфимского промышленного центра наблюдается снижение концентрации хлорофиллов a и b и каротиноидов, причем наиболее чувствительными к загрязнению являются хлорофилл b и каротиноиды (Васильева, 2011), а реакция пигментов ивы белой зависит от периода вегетации: до середины вегетации наблюдаются незначительные и недостоверные уменьшение содержания хлорофилла а, увеличение содержания хлорофилла b, и значительное и достоверное компенсаторное увеличение содержания каротиноидов; в середине вегетации происходит резкий скачок увеличения содержания всех пигментов; в конце вегетации концентрация всех пигментов понижается (Ахмадуллин, 2014). Высокие уровни загрязнения SO2, NO2 и взвешенными веществами в городских условиях значительно снижают скорость фотосинтеза у Platanus occidentalis L. (Сеул, Южная Корея) и у Erythrina orientalis Murr. (Макати и Кезоне, Филиппины), но при этом значительно повышают активность антиоксидантных ферментов – аскорбатпероксидазы и глутатионредуктазы, что является компенсаторным механизмом для минимизации ущерба от стресса (Woo, Je, 2006; Woo et al., 2007). Аналогичные результаты прослеживаются в ряде работ (Masuch et al., 1992; Chapla, Kamalakar, 2004; Paolettia, Grulke, 2005; Prajapati, Tripathi, 2008; Gao et al., 2016; Mikhailova et al., 2017; Rostunov et al., 2017; Duan et al., 2019).
Однако, вне зависимости от древесного вида и типа промышленного загрязнения в отношении физиологических параметров могут наблюдаться неспецифические реакции. На отвалах угольной промышленности Кузбасса у хвои Pinus sylvestris L. выявлено незначительное повышение водоудерживающей способности, снижение суточных потерь воды и значительное возрастание водного дефицита. В динамике вегетационного периода наиболее ярко эти тенденции проявляются в июле (Цандекова, Колмогорова, 2016). Для Acer platanoides L., Populus tremula L., P. nigra L. и P. balsamifera L. в условиях нефтехимического загрязнения Уфимского промышленного центра показано, что листья характеризуются высоким относительным содержанием воды и низким дефицитом водного насыщения, у всех видов наблюдается усиление интенсивности транспирации и нарушение суточного хода данного параметра (Уразгильдин, Кужлева, 2003; Васильева, 2011). В условиях избыточных концентраций металлов в почвах, прилегающих к заводам в г. Сосновец (Польша), показано пониженное относительное содержание воды в листьях Robinia pseudoacacia L., причем с усилением загрязнения оно значительно снижается. Данная тенденция сохраняется на протяжении всей вегетации (Skrynetska et al., 2018). Загрязнение воздушного бассейна г. Калининград приводит к значительному увеличению уровня зеленых пигментов, каротиноидов и антоциановых пигментов у деревьев Sorbus aucuparia L., Tilia cordata Mill., Acer platanoides L. и Picea pungens Engelm. У всех видов выявлена прямая корреляционная зависимость между количеством зеленых пигментов и аскорбиновой кислотой и обратная – между содержанием хлорофиллов a и b и каротиноидов (Майдебура, 2006). Показано увеличение относительного содержания хлорофилла в листьях Brachylaena discolor DC. на участках, загрязненных диоксидом серы в промышленном районе Южного Дурбанского бассейна (Южная Африка), причем этот показатель значительно коррелирует с сезонным изменением концентраций SO2 в приземном слое и в листьях (Areington et al., 2017). Более высокие концентрации хлорофиллов a, b и каротиноидов наблюдаются в листьях Callistemon citrinus (Curtis) Skeels возле нефтехимических заводов в промышленном районе г. Махшахр (Иран) (Seyyednejad et al., 2009a). Ряд других исследований показывает аналогичные результаты (Тужилкина, 2009; Тужилкина, Плюснина, 2014; Баландайкин, 2014; Freer-Smith, Taylor, 1992; Burkhardt et al., 2001; Matysiak, 2001).
Также может наблюдаться и отсутствие физиологических реакций на техногенное воздействие. Последовательное влияние имитированного кислотного дождя (рН 3.0 в течение 1 мес.) и засухи (20 дн.) на водный обмен саженцев Pinus radiata D. Don показало, что кислотные дожди не влияют ни на водный потенциал хвои, ни на относительное содержание воды, но отмечается заметное влияние на транспирацию. Последовавшая засуха оказала большое влияние на потенциал воды, относительное содержание воды, проницаемость мембраны и скорость транспирации. Следовательно, кислотные дожди, не оказывая значительного прямого воздействия, изменяют толерантность этого вида к засухе и повышают чувствительность к обезвоживанию (Mena-Petite et al., 1999). Вариационный анализ показал, что атмосферные осадки и концентрация NO в воздухе не оказывают существенного влияния на скорость транспирации городских деревьев Ginkgo biloba L., Aesculus chinensis Bunge, Magnolia liliiflora Desr., Robinia pseudoacacia L., Pinus tabuliformis Carr., Cedrus deodara (Roxb.) G. Don, Castanea mollissima Blume в Пекине (Китай), а температура (воздуха, почвы и общая радиация), влажность (дефицит давления пара, относительная влажность воздуха и влажность почвы) и загрязнители (NO2, O3, SO2 и PM2.5) значительно влияют на транспирацию деревьев (как суточную, так и почасовую) в течение всего года. Атмосферное загрязнение, характеризуемое концентрациями отдельных загрязнителей, является лишь дополнительным незначительным фактором стресса в городской среде, а условия микроклимата являются определяющими факторами, регулирующими характер транспирации деревьев (Wang et al., 2011, 2012, 2014).
Особый интерес вызывают публикации, в которых показана относительная независимость физиологических адаптивных реакций в пределах одного органа, когда в листьях и хвое одновременно наблюдаются разнонаправленные специфические, неспецифические и нейтральные реакции, несмотря на единство и целостность этих органов. В условиях города Пскова отмечено уменьшение содержания общей воды в листьях Betula pendula Roth., Tilia cordata L., Syringa vulgaris L. и Populus nigra L., при этом наблюдаются видоспецифические перераспределения во фракционном составе – уменьшается количество свободной воды, но увеличивается количество связанной. У всех видов при усилении загрязнения снижается водоудерживающая способность и степень суккулентности, но усиливается интенсивность транспирации и дефицит водного насыщения (Хмелевская, 2008). В условиях загрязнения Сыктывкарского лесопромышленного комплекса показано, что на контрольном и загрязненном участках влагоемкость, оводненность и водный потенциал хвои Picea obovata Ledeb. практически одинаковы, но в условиях загрязнения при возрастании устьичной проводимости наблюдается незначительное повышение водного дефицита и значительное снижение интенсивности транспирации и фотосинтеза (Сенькина, 2017). В условиях загрязнения городской среды г. Краков (Польша) показано, что в период май–июль при усилении загрязнения содержание воды в листьях Tilia cordata Mill и Populus × canadensis Moench сорт Robusta значительно понижается, но в период август–ноябрь реакция меняется на противоположную, и оводненность листьев возрастает у обоих видов, причем к ноябрю разница между загрязнением и контролем становится значительной, и особенно ярко это проявляется у тополя (Klamerus-Iwan et al., 2018). В условиях нефтехимического загрязнения Уфимского промышленного центра показаны видоспецифические реакции пигментного комплекса основных лесообразователей: увеличение содержания всех пигментов – Betula pendula Roth.; уменьшение содержания всех пигментов – Larix sukaczewii Dyl.; увеличение содержания хлорофиллов и уменьшение содержания каротиноидов – Pinus sylvestris L., Tilia cordata Mill; уменьшение содержания хлорофиллов и увеличение содержания каротиноидов – Picea obovata Ledeb., Quercus robur L. У лиственных видов в течение вегетации происходит снижение содержания всех пигментов, в то время как у хвойных наблюдается снижение их содержания до июля и существенное увеличение к концу вегетации. У липы в условиях загрязнения резко возрастает доля хлорофилла а в составе фотосинтезирующих пигментов, остальные виды характеризуются стабильностью пигментного состава в соотношении “Хл а/Хл b” на протяжении всего вегетационного периода. Соотношение (Хл а + + Хл b)/Каротиноиды показывает, что у хвойных видов увеличивается доля хлорофиллов а и b на фоне уменьшения доли каротиноидов, а у лиственных лесообразователей, напротив, увеличивается доля каротиноидов на фоне уменьшения долей хлорофиллов а и b (Уразгильдин и др., 2016; Urazgil’din et al., 2018). Промышленное загрязнение в Паневнике (Польша) вызывает снижение содержания хлорофильных пигментов в хвое текущего года генерации Pinus strubus L. и P. nigra Arnd., но незначительное увеличение у P. silvestris L. и Pseudotsuga menziesii Franco. При этом у всех четырех видов наблюдается значительное возрастание содержания феофитина. В то же время в хвое второго года генерации всех четырех видов все пигменты показывают рост относительно контроля, но в разной степени интенсивности (Gowin, Goral, 1977). Подобные результаты прослеживаются в ряде других работ (Бухарина и др., 2007б; Уразгильдин, Сейдафаров, 2007; Бухарина, Двоеглазова, 2010; Сунцова и др., 2011; Ахмадуллин, Зайцев, 2013а; Зиятдинова и др., 2013; Аминева и др., 2016; Донцов и др., 2016; Prusty et al., 2005; More, Chaubal, 2017; Mukherjee, Agrawal, 2018).
Следует отметить, что широкое распространение получила методика вычисления индекса толерантности видов к промышленному загрязнению. Однако, анализ публикаций, использующих его для оценки устойчивости растений к загрязнению, показывает значительную противоречивость получаемых данных как при сравнении с результатами других авторов, так и в пределах собственных исследований. Например, индексы толерантности одного и того же растения в пределах одного региона с одним типом загрязнения у разных авторов могут меняться на противоположные, а в других случаях по отношению к одному и тому же источнику загрязнения показаны значительные перепады по мере приближения от контроля к источнику, что не дает возможности установить четкую тенденцию (Joshi, Swami, 2007, 2009; Tripathi, Gautam, 2007; Lakshmi et al., 2008; Liu, Ding, 2008; Agbaire, 2009; Agbaire, Esiefarienrhe, 2009; Das, Prasad, 2010; Jyothi, Jaya, 2010; Meerabai et al., 2012; Thambavani, Sabitha, 2012; Qadir et al., 2016). Следовательно, несмотря на высокую чувствительность физиологических параметров к промышленному загрязнению, изменение их количественных характеристик в значительной степени зависит от адаптивного потенциала растения как к уровню, так и к типу загрязнения.
Влияние техногенеза на дендрохронологию древесных растений
Анализ публикаций показывает, что как правило наблюдается однозначное уменьшение величины радиального прироста древесных видов в ответ на промышленное загрязнение (Стравинскене, 1987; Таранков, Матвеев, 1994; Демьянов и др., 1996; Неверова, 2002; Кучеров, Мулдашев, 2003; Бойко, 2005; Сейдафаров, 2009; Матвеев, Акулов, 2012; Аминева и др., 2014; Рыбаков, 2016; Ярмишко и др., 2017; Кладько, Бенькова, 2018; Woodwell, Miller, 1963; Danek, 2007; Kuang et al., 2008; Song et al., 2009; Pärn, Mandre, 2011; Wertz, 2012; Godek et al., 2015; Sensuła et al., 2015, 2017; Barniak, Krąpiec, 2016; Łuszczyńska et al., 2018; Perone et al., 2018; Rutkiewicz, Malik, 2018; Schekalev et al., 2020), потеря его чувствительности к климатическим сигналам (Кучеров, Мулдашев, 2003; Аминева и др., 2014; Уразгильдин и др., 2018; Sensuła et al., 2015, 2017;), перераспределение долей ранней и поздней древесины в общем приросте (Мусаев, 1995; Глазун, 1998; Неверова, 2002; Чжан и др., 2011; Матвеев, Акулов, 2012; Белов, 2017; Skuterud et al., 1994), изменение длительности онтогенетических периодов и нарушение цикличности прироста (Таранков, Матвеев, 1994; Николаева, Савчук, 2009; Аминева и др., 2014; Уразгильдин и др., 2018), появление повышенного количества ложных годичных колец или их выпадение (Пастернак и др., 1989; Радиационное…, 1990; Кучеров, Мулдашев, 2003; Barniak, Krąpiec, 2016), ускоренное начало старения древостоев (Кладько, Бенькова, 2018; Уразгильдин и др., 2018), зависимость величины прироста от расстояния между древостоями и источниками эмиссии и от ландшафта местности (Неверова, 2002; Матвеев, Акулов, 2012; Danek, 2007; Sensuła et al., 2015, 2017; Barniak, Krąpiec, 2016; Perone et al., 2018; Schekalev et al., 2020), а также тесная связь между уменьшением ширины древесных колец и содержанием в годичных кольцах металлов и различных микроэлементов (Мюльгаузен, Панкратова, 2016; Рыбаков, 2016; Song et al., 2009; Perone et al., 2018), восстановление прироста после снижения объемов выбросов загрязняющих веществ (Борисова, 2009; Ярмишко и др., 2017; Danek, 2007; Pärn, Mandre, 2011; Stravinskiene et al., 2013; Sensuła et al., 2015, 2017; Barniak, Krąpiec, 2016; Łuszczyńska et al., 2018).
Однако, нефтехимическое, радиационное и некоторые типы смешанных загрязнений способны стимулировать увеличение радиального прироста, при этом эффекты видоспецифичны и зависят от возраста и жизненного состояния деревьев (Мусаев, 1995; Глазун, 1998; Чжан и др., 2011; Щетинкин, Щетинкина, 2014; Белов, 2017; Уразгильдин и др., 2018; Chandorkar, Demgler, 1987; Skuterud et al., 1994; Stravinskiene et al., 2013). Предполагается, что углеводороды в составе нефтехимического загрязнения могут действовать в качестве “внекорневой подкормки” на хвойные древесные виды, поскольку являются химическими аналогами естественного почвенного гумуса (Уразгильдин и др., 2018). В настоящее время имеющиеся данные не позволяют сформировать единое мнение о влиянии хронического облучения на рост растений: одни исследователи указывают на ингибирование роста, другие отвергают какие-либо особенности в его действии, а третьи указывают на возможность позитивного действия относительно невысоких доз радиации (Белов, 2017). Так, с одной стороны, в сосняках юга Нечерноземной зоны европейской части России отмечено негативное влияние даже сравнительно малых доз радиации на радиальный прирост на следующий год после аварии на ЧАЭС (Скок, 2005). С другой стороны, утверждается, что хроническая радиация при МЭД до 1.5 мР/ч не оказывает сильного ингибирующего действия на ростовые процессы, а зачастую вызывает их стимуляцию (Глазун, 1998).
Влияние техногенеза на корневые системы древесных растений
Промышленные эксгалаты изменяют, прежде всего, физико-химические свойства почвы, вызывая увеличение ее кислотности (Baker et al., 1975) или, наоборот, подщелачивание (Nyborg et al., 1975), нередко в условиях техногенеза имеет место уменьшение содержания в почве минеральных веществ (Paavilainen, 1967; Safford, 1976) и изменение микробиологического состава ризосферы (Newman, Hart, 2006). Наиболее чувствительными как к естественным, так и к антропогенным факторам, являются поглощающие корни, тем не менее, загрязнение вызывает, как правило, существенное уменьшение корненасыщенности почвы всеми фракциями корней, как в условиях промышленных центров (Мусаев, 1995; Ярмишко, 1997; Веселкин, 1999; Кулагин, Зайцев, 2008; Helmisaari et al., 1999; Puhe, 2003; Albrechtová et al., 2017), так и в опытах с искусственным внесением токсикантов в среду (Kahle, 1993; Lux et al., 2004, 2011; Cudlín et al., 2007; Li et al., 2020). Интересным является адаптивный механизм, когда при общем угнетении корневых систем или угнетении только отдельных корневых фракций наблюдается перераспределение во фракционном составе в пользу увеличения долей тех или иных корневых фракций (Кулагин, Зайцев, 2008; Сейдафаров, 2009; Васильева и др., 2011; Яшин и др., 2012; Ахмадуллин, Зайцев, 2013б; Аминева, 2016), а также эффект “избегания” корневыми системами наиболее загрязненных слоев почвенного профиля (Ярмишко, 1997; Веселкин, 1999; Зайцев и др., 2019). В одних случаях наблюдается интенсивное накопление токсикантов в корнях (Brunner et al., 2008), а в других случаях – активное выделение корневых экссудатов, препятствующих проникновению токсикантов в растение (Wang et al., 2020).
Как и в случае с радиальным приростом, нефтехимическое загрязнение способно стимулировать увеличение корненасыщенности почвы у хвойных древесных видов (Зайцев, 2000; Скотников, 2007), однако для лиственных видов данный эффект носит видоспецифический характер (Сейдафаров, 2009; Васильева и др., 2011; Яшин и др., 2012; Ахмадуллин, Зайцев, 2013б; Аминева, 2016). Но радиоактивное загрязнение наносит более значительное повреждение корневым системам, чем радиальному приросту (Мусаев, 1995).
Влияние техногенеза на жизненное состояние древостоев
В отличие от морфологии и физиологии хвои и листьев, морфологии корневых систем и дендрохронологических характеристик, показатели жизненного состояния древостоев не проявляют адаптивный полиморфизм, неспецифические реакции отсутствуют и наблюдается однозначное ухудшение жизненного состояния древостоев вне зависимости от древесного вида и типа загрязнения (Уразгильдин, 1998; Бойко, 2005; Колмогорова, 2005; Зайцев, Кулагин, 2006; Скотников, 2007; Ковылина и др., 2008; Кулагин, Зайцев, 2008; Сейдафаров, 2009; Кулагин и др., 2010; Васильева, 2011; Радостева, 2011; Ахмадуллин, 2014; Аминева, 2016; Зайцев и др., 2017; Armolaitis, 1998; Šrámek, 1998; Vacek et al., 2013; Jakovljević et al., 2019). Однако степень ухудшения во многом зависит от древесного вида и типа загрязнения. Жизненное состояние древостоев является наиболее точным и понятным показателем, характеризующим влияние техногенеза на древостои, поскольку в методическом плане является комплексной оценкой как габитуальных, так и лесотаксационных параметров всего фитоценоза. Следует отметить стремительное развитие методического аппарата оценки жизненного состояния древостоев и переход от упрощенных схем только визуальной оценки к интегрированию качественных и количественных параметров древостоев.
АДАПТИВНЫЕ СТРАТЕГИИ
Как видно из данного обзора, полиморфизм адаптивных реакций древесных видов на техногенный стресс на каждом структурно-функциональном иерархическом уровне довольно обширен, даже в пределах одного органа. Встает закономерный вопрос: как же оценить всю эту сумму разнонаправленных реакций, как на их основе определить адаптивную стратегию вида к конкретному типу загрязнения и какой адаптивный потенциал лежит в основе реализации этой стратегии?
Промышленное загрязнение, как новый для растений в историческом плане фактор, вызывает необходимость разработки вопросов, связанных с адаптивными стратегиями видов к техногенезу, в основе которых лежит адаптивный потенциал видов. Адаптивный потенциал древесных растений складывается из изменчивости, экологической пластичности и устойчивости на различных структурно-функциональных уровнях организации. Обеспечение гомеостаза растений при развитии в условиях техногенеза сопровождается неизбежными флуктуациями показателей на различных уровнях организации, при этом величина максимальных флуктуаций определяет экологическую пластичность, а минимальные величины флуктуаций служат показателем устойчивости вида (Кулагин, 2006). Таким образом, адаптивная стратегия представляет собой реализованную часть адаптивного потенциала вида и характеризуется широким адаптивным полиморфизмом, проявляющимся комплексом специфических и неспецифических адаптивных реакций (а также степенью их изменчивости и степенью взаимозависимости между ними), протекающих на всех уровнях организации живого – от цитогенетического до экосистемного.
Разработка вопроса адаптивных стратегий к техногенезу уходит корнями к динамическому развитию экологических стратегий Раменского–Грайма (Раменский, 1938; Grime, 1979), в основу которых положено “поведение” видов в отношении широкого спектра эколого-ценотических ситуаций. В поступательном развитии данного направления лежит множество работ, предлагавших как подходы к классификации систем адаптивности растений, так типы и подтипы стратегий (Серебрякова, 1971; Уиттекер, 1980; Работнов, 1992; Усманов и др., 2001; Миркин, Наумова, 2012; MacLeod, 1884 по Hermy, Stieperaete, 1985; MacArthur, Wilson, 1967). Однако, эколого-ценотические адаптивные стратегии не дают ответов на вопросы каковы адаптивные стратегии видов к новому для них в историческом плане техногенному фактору при естественно сложившемся и уже привычном для них фитоценотическом факторе.
Это дало толчок к развитию нового направления в области дендроэкологии – адаптивные стратегии растительных организмов к техногенезу. Наиболее яркий подход к систематизации этого вопроса, но не для отдельного вида, а для совокупности разных видов растений, предложен В.К. Жировым с соавт. (2006). Авторами рассматривается вопрос о взаимодействии структур различных уровней организации растительного организма при формировании адаптивных реакций к промышленному загрязнению. Ими предложено степень независимости или согласованности отдельных структур иерархий в условиях, далеких от экологического оптимума, определять пассивной или активной стратегией адаптивного ответа организма и надорганизменных структур. При оценке изменчивости структур различных иерархий в градиенте техногенного воздействия в аспекте вариаций уровня их интегрированности ими предложено три типа реакций: 1) активизация процессов энергообмена при снижении целостности на всех уровнях; 2) снижение уровня энергообмена при возрастании целостности структур всех уровней и падении их устойчивости; 3) дальнейшее падение уровня энергообмена при снижении целостности структур всех иерархий.
Анализ совокупности современных подходов к определению и выявлению адаптивных стратегий к техногенному фактору позволил разработать авторский подход к данному вопросу, основанный на материалах собственных многолетних исследований (Уразгильдин, 2009). Представляется целесообразным рассматривать адаптивную стратегию древесного вида к техногенному фактору как степень согласованности относительно независимых (разнонаправленных) адаптивных реакций (биохимических, физиологических, анатомических, морфологических, габитуальных и т.д.), совокупно проявляющихся на всех иерархических структурно-функциональных уровнях организации древесных растений популяции. Исходя из современных представлений об адаптивных реакциях организма, стрессе и толерантности, предлагается различать следующие адаптивные реакции древесных растений к действию техногенного фактора:
– стрессовые – при низком адаптивном потенциале адаптивные реакции направлены на активное ограничение влияния стрессового фактора, затрачивая значительные энергетические ресурсы;
– умеренно-стрессовые – незначительное или недостоверное проявление адаптивных реакций, направленных на ограничение влияния стрессового фактора;
– нейтральные – отсутствие реакций при влиянии стрессового фактора;
– умеренно-толерантные – возрастающий адаптивный потенциал позволяет незначительно или недостоверно усиливать процессы роста и развития, несмотря на давление стрессового фактора;
– толерантные – высокий адаптивный потенциал позволяет быть невосприимчивым к стрессовому фактору, не расходовать энергию на ограничение его влияния и без вреда усиливать процессы роста и развития.
Отнесение того или иного рассматриваемого параметра к конкретной адаптивной реакции принимается на основе степени и достоверности его изменения под действием стрессирующего фактора. Итоговая адаптивная стратегия определяется как мода совокупности адаптивных реакций, а количественная оценка степени согласованности адаптивных реакций проводится с использованием коэффициентов меры разнообразия: в пределах иерархических уровней используется коэффициент равномерности, а между иерархическими уровнями – среднеквадратическое отклонение. Чем больше иерархических уровней организации будет вовлечено в оценку, тем больше будут выражены мода, коэффициент меры разнообразия и адаптивная стратегия древесного вида. В целом, чем толерантнее адаптивная стратегия вида, тем выше его адаптивный потенциал и успешнее его существование в условиях стрессирующего фактора.
Разработанные методические подходы позволили оценить адаптивные стратегии и адаптивный потенциал лесообразователей Предуралья к нефтехимическому загрязнению на примере Уфимского промышленного центра: сосна, лиственница и дуб характеризуются “толерантной” адаптивной стратегией и высоким адаптивным потенциалом, липа характеризуется “стрессовой” адаптивной стратегией и низким адаптивным потенциалом, ель и береза характеризуются “нейтральной” адаптивной стратегией и средним адаптивным потенциалом. Для каждого древесного вида показана относительная независимость адаптивных реакций на каждом иерархическом структурно-функциональном уровне и относительная независимость адаптивных реакций между иерархическими уровнями. У всех видов наибольшей степенью согласованности адаптивных реакций в пределах иерархических уровней характеризуются показатели жизненного состояния древостоев и морфология корневой системы, наименьшей – параметры водного обмена хвои/листьев. По степени согласованности адаптивных реакций между иерархическими уровнями древесные виды образуют ряд уменьшения: береза > липа > лиственница > ель > сосна > дуб.
ЗАКЛЮЧЕНИЕ
В основе рассмотренного многообразия адаптивных реакций лежит принцип множественного обеспечения биологически необходимых функций для сохранения гомеостаза, когда биосистема для поддержания своего существования в стрессовых условиях должна противопоставить воздействию внешней среды достаточное разнообразие соответствующих реакций. Чем большее число механизмов адаптации используется растением одновременно на самых разных уровнях организации ассимиляционного аппарата, тем более устойчив организм к действию токсических ингредиентов. Кроме того, не каждый стресс обязательно негативен для деревьев, он может вместо этого вызвать повышенную устойчивость к стрессу, а кратковременная стрессовая реакция может не совпадать с длительным изменением жизненного состояния деревьев (Маслов, 1980; Тарабрин, 1984; Kozlowski, Pallardi, 2002). Таким образом, в зависимости от величины адаптивного потенциала растения в процессе приспособления включаются те механизмы, которые позволяют при наименьших энергетических затратах повысить его толерантность к стрессовому фактору, а многообразие реакций является залогом повышения устойчивости к фитотоксикантам. Результирующая схема реализации адаптивных стратегий древесных видов к техногенезу нам представляется в следующем виде (рис. 1).
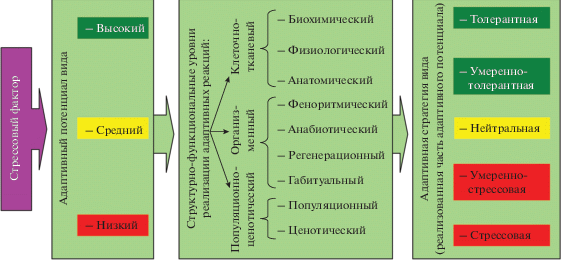
Предложенная схема гармонично укладывается в методические подходы выделения и оценки адаптивных реакций на структурно-функциональных иерархических уровнях организации древесного растения, определения на их основе адаптивной стратегии вида к техногенному фактору и опосредованной оценке адаптивного потенциала вида.
В заключение следует отметить ряд проблемных вопросов, решение которых позволит более детально оценивать влияние техногенеза на древесные растения:
– фрагментарность исследований – оценка влияния техногенеза на один определенный орган (как правило ассимиляционный аппарат вследствие его доступности и тесный контакт с окружающей средой) или ограниченную группу параметров или процессов;
– малочисленность работ с оценкой эффектов в их развитии – в динамике вегетационного периода и в многолетней динамике;
– простое описание выявленного эффекта без оценки сути адаптивной реакции и ее направленности;
– отсутствие увязки в единую адаптивную схему множества выявленных эффектов;
– ошибочная подмена понятия “адаптивная стратегия” выявленными “адаптивными реакциями”. Зарубежные исследователи довольно часто оперируют понятием “адаптивная стратегия” при обсуждении реакций какого-то конкретного параметра (группы параметров) или органа в ответ на внешнее воздействие. На наш взгляд, в данных случаях корректнее использовать термин “адаптивная реакция”, так как речь идет о реакциях только на одном определенном структурно-функциональном уровне организации растительного организма, в то время как “адаптивная стратегия” подразумевает совокупное использование всех имеющихся защитно-приспособительных механизмов, складывающихся из комплекса относительно независимых адаптивных реакций на всех структурно-функциональных уровнях организации растительного организма – то есть всего растения в целом;
– необходим переход от анализа видоспецифических адаптивных реакций (на отдельных уровнях структурно-функциональной организации) к комплексной оценке адаптивных стратегий древесных видов (как единого целого организма) на тот или иной вид техногенного загрязнения с вовлечением в оценку как можно большего числа иерархических уровней организации (от морфологии и физиологии корневых систем до морфологии и физиологии хвои/листьев);
– на наш взгляд в публикациях мало уделяется внимания вопросам разработки методических подходов к оценке адаптивного потенциала растений к техногенезу и разработке классификации адаптивного потенциала. На сегодняшний день имеется определение адаптивного потенциала и его общая характеристика (высокий–средний–низкий), однако необходима детальная количественная и качественная оценка этого термина.
Список литературы
Аминева К.З. Эколого-биологическая характеристика дуба черешчатого (Quercus robur L.) в условиях техногенного загрязнения (на примере Уфимского промышленного центра): Дис. … канд. биол. наук. Тольятти: ИЭВБ РАН, 2016. 163 с.
Аминева К.З., Уразгильдин Р.В., Кулагин А.Ю. Прирост стволовой древесины дуба черешчатого (Quercus robur L.) в условиях техногенного загрязнения // Биосфера. 2014. Т. 6. № 4. С. 388–399.
Аминева К.З., Уразгильдин Р.В., Кулагин А.Ю., Денисова А.В. Вегетационная динамика водного обмена хвои ели сибирской, сосны обыкновенной, лиственницы Сукачева в условиях техногенного загрязнения Предуралья // Карельский науч. журн. 2016. Т. 5. № 1 (14). С. 81–86.
Ахмадуллин Р.Ш. Эколого-биологическая характеристика ивы белой (Salix alba L.) в условиях Уфимского промышленного центра: Дис. … канд. биол. наук. Оренбург: ГОУ ВПО ОГПУ, 2014. 175 с.
Ахмадуллин Р.Ш., Зайцев Г.А. Особенности транспирации листьев ивы белой в условиях нефтехимического загрязнения // Изв. Саратовского ун-та. Серия Химия. Биология. Экология. 2013а. Т. 13. Вып. 2. С. 53–56.
Ахмадуллин Р.Ш., Зайцев Г.А. Корненасыщенность почвы в насаждениях ивы белой (Salix alba L.) в условиях нефтехимического загрязнения Уфимского промышленного центра // Изв. Уфимского НЦ РАН. 2013б. № 3. С. 30–33.
Баканов А.В. Экологическая оценка состояния лесных насаждений с помощью методов фитоиндикации на примере Сергиево-Посадского района: Дис. … канд. биол. наук. М: МГУЛ, 1997. 198 с.
Баландайкин М.Э. Динамика и различия в концентрации основных фотосинтетических пигментов листьев березы, произрастающей в неоднородных условиях // Химия раст. сырья. 2014. № 1. С. 159–164.
Белов А.А. Динамика радиального прироста сосны обыкновенной в насаждениях Брянской области, загрязненных радионуклидами: Дис. … канд. биол. наук. Москва: ФБУ ВНИИЛМ, 2017. 158 с.
Бикмуллин Р.Х. Оценка состояния древостоев сосны обыкновенной (Pinus sylvestris L.) и березы повислой (Betula pendula Roth) на территории Казанского промышленного центра Республики Татарстан: Дис. … канд. биол. наук. Оренбург: ГОУ ВПО ОГПУ, 2012. 169 с.
Бойко А.А. Дендроэкологическая характеристика березы повислой (Betula pendula Roth.) в условиях смешанного типа загрязнения окружающей среды (Уфимский промышленный центр): Дис. … канд. биол. наук. Оренбург: ГОУ ВПО ОГПУ, 2005. 122 с.
Борисова О.В. Влияние аэротехногенного загрязнения на хвойные и лиственные древостои в Новгородской области: Дис. … канд. биол. наук. Тольятти: ИЭВБ РАН, 2009. 155 с.
Бухарина И.Л., Двоеглазова А.А. Биоэкологические особенности травянистых и древесных растений в городских насаждениях. Ижевск: Изд-во “Удмуртский университет”, 2010. 184 с.
Бухарина И.Л., Ведерников К.Е., Поварницина Т.М. К вопросу о влиянии техногенной среды на формирование и биохимический состав годичного побега древесных растений // Вестн. ИжГТУ. 2007а. № 2. С. 145–148.
Бухарина И.Л., Поварницина Т.М., Ведерников К.Е. Эколого-биологические особенности древесных растений в урбанизированной среде. Ижевск: ФГОУ ВПО Ижевская ГСХА, 2007б. 216 с.
Василевская Н.В., Лебедевич И.Е. Воздействие техногенного загрязнения г. Мурманска на рост и развитие Sorbus gorodkovii Pojark // Естеств. и техн. науки. 2016. № 4 (94). С. 23–28.
Василевская Н.В. Сидорчук А.В. Воздействие промышленного загрязнения комбината “Печенганикель” на динамику роста Sorbus gorodkovii Pojark (Мурманская область) // Уч. записки ПетрГУ. 2018. № 3 (172). С. 28–35.
Васильева К.А. Эколого-биологические особенности клена остролистного (Acer platanoides L.) в условиях техногенного загрязнения: Дис. … канд. биол. наук. Уфа: ИБ УНЦ РАН, 2011. 160 с.
Васильева К.А., Зайцев Г.А., Кулагин А.Ю. Особенности строения корневых систем клена остролистного (Acer platanoides L.) в условиях нефтехимического загрязнения // Вестн. Удмуртского ун-та. Серия 6: Биология. Науки о Земле. 2011. Вып. 2. С. 55–60.
Веселкин Д.В. Освоение почвы корнями хвойных при загрязнении тяжелыми металлами // Тез. докл. науч. конф. “Б.П. Колесников – выдающийся отечественный лесовод и эколог. К 90-летию со дня рождения”. Екатеринбург: [б.и.], 1999. С. 19.
Веселова Т.В., Веселовский В.А., Чернавский Д.С. Стресс у растений (биофизический подход). М.: Издательство МГУ, 1993. 144 с.
Глазун И.Н. Изменчивость хвойных растений в радиоактивно загрязненных насаждениях Брянского округа зоны широколиственных лесов: Дис. … канд. с.-х. наук. Брянск: БГИТА, 1998. 161 с.
Демьянов В.А., Китсинг Л.И., Ярмишко В.Т. Влияние промышленного загрязнения на радиальный прирост Larix Gmelinii (Pinaceae) // Изв. РАН. Серия биол. 1996. № 4. С. 490–494.
Донцов А.С., Сунцова Л.Н., Иншаков Е.М. Оценка состояния окружающей среды г. Красноярска по состоянию фотосинтетического аппарата ели сибирской // Хвойные бореальной зоны. 2016. Т. 37. № 5–6. С. 246–250.
Жиров В.К., Хаитбаев А.Х., Говорова А.Ф., Гонтарь О.Б. Взаимодействия структур различных уровней организации и адаптационные стратегии растений // Вестн. МГТУ. 2006. Т. 9. № 5. С. 725–728.
Зайцев Г.А. Особенности формирования корневых систем сосны обыкновенной (Pinus sylvestris L.) и лиственницы Сукачева (Larix sukaczewii Dyl.) в техногенных условиях Предуралья (Уфимский промышленный центр): Дис. … канд. биол. наук. Уфа: БГУ, 2000. 137 с.
Зайцев Г.А., Кулагин А.Ю. Сосна обыкновенная и нефтехимическое загрязнение. Дендроэкологическая характеристика, адаптивный потенциал и использование. М.: Наука, 2006. 124 с.
Зайцев Г.А., Дубровина О.А., Логвинов К.В. Корневая система березы повислой в условиях загрязнения // ЭКОБИОТЕХ 2019 / Мат. VI Всерос. конф. с междунар. участием. 2019. С. 140–143.
Зайцев Г.А., Кулагин А.Ю., Уразгильдин Р.В. и др. Относительное жизненное состояние древесных насаждений в условиях промышленного загрязнения // Изв. Уфимского НЦ РАН. 2017. № 2. С. 63–68.
Зиятдинова К.З., Уразгильдин Р.В., Денисова А.В. Водный обмен листьев дуба черешчатого (Quercus robur L.) в условиях техногенного загрязнения окружающей среды // Вестн. ЧелГУ. 2013. № 7. Вып. 2. С 181–184.
Кладько Ю.В., Бенькова В.Е. Радиальный рост древесных видов в условиях высокой антропогенной нагрузки г. Красноярска // Сиб. лесн. журн. 2018. № 4. С. 49–57.
Ковылина О.П., Зарубина И.А., Ковылин А.Н. Оценка жизненного состояния сосны обыкновенной в зоне техногенного загрязнения // Хвойные бореальной зоны. 2008. Т. 25. № 3–4. С. 284–289.
Колмогорова Е.Ю. Видовое разнообразие и жизненное состояние древесных и кустарниковых растений в зеленых насаждениях города Кемерово: Дис. … канд. биол. наук. Томск: ТГУ, 2005. 163 с.
Кулагин А.А. Реализация адаптивного потенциала древесных растений в экстремальных лесорастительных условиях: Дис. … докт. биол. наук. Тольятти. ИЭВБ РАН, 2006. 430 с.
Кулагин А.А., Зайцев Г.А. Лиственница Сукачева в экстремальных лесорастительных условиях Южного Урала. М.: Наука, 2008. 171 с.
Кулагин А.Ю., Гиниятуллин Р.Х., Уразгильдин Р.В. Средостабилизирующая роль лесных насаждений в условиях Стерлитамаксткого промышленного центра. Уфа: Гилем, 2010. 108 с.
Кулагин Ю.З. Индустриальная дендроэкология и прогнозирование. М.: Наука, 1985. 118 с.
Кучеров С.Е., Мулдашев А.А. Радиальный прирост сосны обыкновенной в районе Карабашского медеплавильного комбината // Лесоведение. 2003. № 2. С. 43–49.
Легощина О.М. Оценка ростовых процессов у древесных растений в условиях преобладающего влияния выбросов промзоны г. Кемерово // Бюл. науки и практики. 2016. № 5. С. 14–19.
Лукина Ю.М. Влияние техногенного загрязнения комбината “Североникель” на рост и развитие древесных растений: на примере Betula Czerepanovii Orlova: Дис. … канд. биол. наук. Петрозаводск: ПГУ, 2011. 177 с.
Майдебура И.С. Влияние загрязнения воздушного бассейна города Калининграда на анатомо-морфологические особенности и биохимические показатели древесных растений: Дис. … канд. биол. наук. Калининград: РГУ им. И. Канта, 2006. 146 с.
Маслов С.П. Ограничения возможностей гомеостаза мультифункциональностью и главные пути его обхода // Уровни организации биологических систем. М.: Наука, 1980. С. 8–19.
Матвеев С.М., Акулов В.В. Динамика состояния сосны обыкновенной (Pinus sylvestris L.) вдоль пригородных автотрасс г. Воронежа 1991–2007 гг. // Вестн. ТГУ. 2012. № 364. С. 212–218.
Миркин Б.М., Наумова Л.Г. Современное состояние основных концепций науки о растительности. Уфа: АН РБ, Гилем, 2012. 488 с.
Мусаев Е.К. Реакция прироста и структуры годичных колец сосны (Pinus sylvestris L.) на радиоактивное воздействие в районе Чернобыльской АЭС: Дис. … канд. биол. наук. Красноярск: Институт леса им. В.Н. Сукачева СО РАН, 1995. 175 с.
Мюльгаузен Д.С., Панкратова Л.А. Влияние аэротехногенного загрязнения на радиальный прирост сосны обыкновенной на Кольском севере // Вестн. СПбГУ. Науки о Земле. 2016. Вып. 4. С. 124–133.
Неверова О.А. Морфометрическая и дендрохронологическая диагностика состояния древесных насаждений как способ индикации загрязнения урбанизированной среды // Успехи соврем. естествознания. Биол. науки. 2002. № 1. С. 57–64.
Неверова О.А., Колмогорова Е.Ю. Ксерофитизация листьев древесных растений как показатель загрязнения атмосферного воздуха (на примере г. Кемерово) // Лесн. хозяйство. 2002. № 3. С. 29–33.
Николаева С.А., Савчук Д.А. Комплексный подход и методика реконструкции роста и развития деревьев и лесных сообществ // Вестн. ТГУ. Биология. 2009. № 2 (6). С. 111–125.
Пастернак П.С., Молотков П.И., Кучма Н.Д., Подкур П.П. Лесоводственно-экологические последствия загрязнения лесов аварийными выбросами // Чернобыль-88 / Докл. 1 Всесоюзн. науч.-техн. совещ. по итогам ликвидации последствий аварии на Чернобыльской АЭС. Чернобыль, 1989. Т. 3. Ч. 2. С. 36–60.
Пахомова В.М. Основные положения современной теории стресса и неспецифический адаптационный синдром у растений // Цитология. 1995. № 1 (2). С. 66–91.
Работнов Т.А. Фитоценология. 3-е изд. М.: МГУ, 1992. 350 с.
Радиационное воздействие на хвойные леса в районе аварии на Чернобыльской АЭС / Ред. Г.М. Козубов, А.И. Таскаев. Сыктывкар: Коми НЦ РАН, 1990. 136 с.
Радостева Э.Р. Эколого-биологическая характеристика насаждений сосны обыкновенной (Pinus sylvestris L.) и березы повислой (Betula pendula Roth) при лесной рекультивации отвалов горнодобывающей промышленности (Республика Башкортостан): Дис. … канд. биол. наук. Уфа: ИБ УНЦ РАН, 2011. 190 с.
Раменский Л.Г. Введение в комплексное почвенно-ботаническое исследование земель. М.: Сельхозгиз, 1938. 620 с.
Рыбаков Д.С. Биогеохимическая оценка экологического риска на примере Pinus sylvestris L. // Принципы экол. 2016. № 2. С. 70–83.
Сейдафаров Р.А. Эколого-биологические особенности липы мелколистной (Tilia cordata Mill.) в условиях техногенного загрязнения (на примере Уфимского промышленного центра): Дис. … канд. биол. наук. Уфа: ИБ УНЦ РАН, 2009. 204 с.
Сенькина С.Н. Показатели водообмена хвои Picea obovata (Pinaceae) в условиях аэротехногенного загрязнения (Республика Коми) // Раст. ресурсы. 2017. Т. 53 (2). С. 255–264.
Серебрякова Т.И. Морфогенез побегов и эволюция жизненных форм. М.: Наука, 1971. 359 с.
Скок А.В. Изменчивость репродуктивных и ростовых процессов сосны обыкновенной в различных зонах хронического радиоактивного загрязнения ЧАЭС Южного Нечерноземья РФ: Дис. … канд. биол. наук. Брянск: БГУ им. акад. И.Г. Петровского, 2005. 158 с.
Скотников Д.В. Дендроэкологическая характеристика ели сибирской (Picea obovata Ledeb.) в условиях нефтехимического загрязнения (Уфимский промышленный центр): Дис. … канд. биол. наук. Уфа: ИБ УНЦ РАН, 2007. 160 с.
Стравинскене В.П. Изменение радиального прироста деревьев в зоне действия промышленного загрязнения // Лесн. хозяйство. 1987. № 5. С. 34–36.
Сунцова Л.Н., Иншаков Е.М., Козик Е.В. Оценка состояния городской среды методом фитоиндикации (на примере г. Красноярска) // Лесн. журн. 2011. № 4. С. 29–32.
Сухарева Т.А., Лукина Н.В. Химический состав и морфометрические характеристики хвои ели сибирской на Кольском полуострове в процессе деградационной сукцессии лесов // Лесоведение. 2004. № 2. С. 36–43.
Тарабрин В.П. Природа устойчивости растений к промышленным эксгалатам // Адаптация древесных растений к экстремальным условиям среды. Петрозаводск: Карельский филиал АН СССР, 1984. С. 90–97.
Таранков В.И., Матвеев С.М. Радиальный прирост древостоев сосны обыкновенной в зоне действия промышленного загрязнения // Лесн. журн. 1994. № 4. С. 48–51.
Тужилкина В.В. Реакция пигментной системы хвойных на длительное аэротехногенное загрязнение // Экология. 2009. № 4. С. 243–248.
Тужилкина В.В., Плюснина С.Н. Комплексная оценка состояния хвои Picea obovata (Pinaceae) в условиях аэротехногенного загрязнения // Раст. ресурсы. 2014. Т. 50. Вып. 4. С. 579–586.
Турмухаметова Н.В. Особенности морфогенеза побегов и феноритмов Betula pendula Roth и Tilia cordata Mill. в условиях городской среды: Дис. … канд. биол. наук. Новосибирск: ЦСБС СО РАН, 2005. 278 с.
Уиттекер Р. Сообщества и экосистемы. М.: Прогресс, 1980. 327 с.
Уразгильдин Р.В. Эколого-биологическая характеристика тополей в условиях загрязнения окружающей среды (на примере Уфимского промышленного центра): Дис. … канд. биол. наук. Уфа: БГУ, 1998. 172 с.
Уразгильдин Р.В. Классификация адаптивных стратегий древесных растений к техногенному загрязнению (на примере липы сердцевидной Tilia cordata Mill.) // Аграрная Россия. 2009. Спец. выпуск. С. 205–209.
Уразгильдин Р.В., Кужлева Н.Г. Водный режим листьев тополей в условиях промышленного загрязнения // Сб. докладов науч.-практ. конф. “Лесное образование, наука и хозяйство”, посвященной 125-летию Уфимского лесхоз-техникума. Уфа: РИО РУНМЦ МО РБ, 2003. С. 174–188.
Уразгильдин Р.В., Сейдафаров Р.А. Водный режим листьев липы мелколистной (Tilia cordata Mill.) в условиях промышленного загрязнения окружающей среды // Вестн. ОГУ. 2007. Вып. 75. С. 369–372.
Уразгильдин Р.В., Аминева К.З., Зайцев Г.А. и др. Сравнительная характеристика формирования пигментного комплекса дуба черешчатого (Quercus robur L.), липы сердцевидной (Tilia cordata Mill.) и березы повислой (Betula pendula Roth) в условиях промышленного загрязнения // Карельский науч. журн. 2016. Т. 5. № 1 (14). С. 90–94.
Уразгильдин Р.В., Полякова Г.Р., Аминева К.З. и др. Прирост стволовой древесины сосны обыкновенной, ели сибирской и лиственницы Сукачева в условиях промышленного загрязнения // Бюл. моск. общ-ва испытателей природы. Отд. Биол. 2018. Т. 123. Вып. 2. С. 45–61.
Усманов И.Ю., Рахманкулова 3.Ф., Кулагин А.Ю. Экологическая физиология растений: учебник. М.: Логос, 2001. 224 с.
Хмелевская И.А. Эколого-физиологические исследования древесных пород в г. Пскове // Вестн. ПГПУ: Серия “Естественные и физико-математические науки”. 2008. Вып. 6. С. 37–57.
Цандекова О.Л., Колмогорова Е.Ю. Особенности адаптационных перестроек хвои Pinus sylvestris L. в условиях породного отвала угольного разреза “Кедровский” // Вестн. ОГУ. 2016. № 6 (194). С. 81–85.
Чжан С.А., Рунова Е.М., Пузанова О.А., Чжан Л.А. Изменение радиального прироста сосны обыкновенной в зоне длительного воздействия промышленного загрязнения // Хвойные бореал. зоны. 2011. Т. 29. № 3–4. С. 304–308.
Щетинкин С.В., Щетинкина Н.А. Особенности динамики радиального прироста дуба черешчатого в условиях радиоактивного загрязнения лесов центральной лесостепи // Лесотехн. журн. 2014. № 3. С. 130–139.
Ярмишко В.Т. Сосна обыкновенная и атмосферное загрязнение на Европейском Севере. Санкт-Петербург: Издательство НИИ Химии СПбГУ, 1997. 210 с.
Ярмишко В.Т., Лянгузова И.В., Лянгузов А.Ю. Изменение годичного прироста стволов Pinus sylvestris (Pinaceae) при снижении аэротехногенного загрязнения // Раст. ресурсы. 2017. № 53 (4). С. 527–542.
Яшин Д.А., Зайцев Г.А., Зиятдинова К.З., Уразгильдин Р.В. Особенности строения корневых систем березы повислой в условиях нефтехимического загрязнения // Изв. Самарского НЦ РАН. 2012. Т. 14. № 1 (6). С. 1581–1583.
Abbasi S., Hosseini S., Khorasani N., Karbassi A. Responses of the morphological traits of elm (Ulmus minor “Umbraculifera”) leaves to air pollution in urban areas (a case study of Tehran metropolitan city, Iran) // Appl. Ecol. Environ. Res. 2018. № 16 (4). P. 4955–4968.
Agbaire P.O. Air pollution tolerance indices (APTI) of some plants around Erhoike-Kokori oil exploration site of Delta State, Nigeria // Int. J. Phys. Sci. 2009. V. 4. P. 366–368.
Agbaire P.O., Esiefarienrhe E. Air pollution tolerance indices (apti) of some plants around Otorogun Gas Plant in Delta State, Nigeria // J. Appl. Sci. Environ. Manag. 2009. V. 13. Iss. 1. P. 11–14.
Albrechtová P., Mauer O., Gebauer R. et al. A comparative study of above- and below-ground parameters of healthy and declining young Norway spruce trees in a mountain area affected by air pollution // Scand. J. Forest Res. 2017. V. 32. Iss. 6. P. 481–487.
Allahnouri M., Ghasemi A.F., Pazhouhan I. Traffic effects on leaf macro- and micromorphological traits // Folia Oecologica. 2018. V. 45. № 2. P. 92–101.
Areington C.A., Varghese B., Sershen N. The utility of biochemical, physiological and morphological biomarkers of leaf sulfate levels in establishing Brachylaena discolor leaves as a bioindicator of SO2 pollution // Plant Physiol. Biochem. 2017. V. 118. P. 295–305.
Aribal L.G., Llamos E.J.N., Bruno A.G.T., Medina M.A. Comparative leaf morphometrics of two urban tree species: an assessment to air pollution impacts // J. Biodiv. Environ. Sci. 2016. V. 9. № 1. P. 106–115.
Armolaitis K. Nitrogen pollution on the local scale in Lithuania: vitality of forest ecosystems // Environ. Poll. 1998. V. 102. Iss. 1. Suppl. 1. P. 55–60.
Baker D.E., Chesin L. Chemical monitoring of soil for environmental quality animal and health // Adv. Agronom. 1975. V. 27. P. 306–366.
Barniak J., Krąpiec M. The tree-ring method of estimation of the effect of industrial pollution on pine (Pinus sylvestris L.) tree stands in the northern part of the Sandomierz basin (SE Poland) // Water Air Soil Poll. 2016. V. 227. Iss. 5. P. 166.
Bezerra L.A., Callado C.H., Cunha M.D. Does an urban environment affect leaf structure of Eugenia uniflora L. (Myrtaceae)? // Acta Botan. Brasil. 2020. № 34 (2). P. 266–276.
Brunner I., Luster J., Günthardt-Goerg M.S., Frey B. Heavy metal accumulation and phytostabilisation potential of tree fine roots in a contaminated soil // Environ. Poll. 2008. V. 152. P. 559–568.
Burkhardt J., Kaiser H., Kappen L., Goldbach H.E. The possible role of aerosols on stomatal conductivity for water vapour // Basic Appl. Ecol. 2001. V. 2. P. 351–364.
Chandorkar K.R., Demgler N.G. Effect of low level continuous gamma irradiation on vascular cambium activity in scotch pine Pinus sylvestris L. // Environ. Experim. Botan. 1987. V. 27. № 2. P. 165–175.
Chapla J., Kamalakar J.A. Metabolic responses of tropical trees to ozone pollution // J. Environ. Biol. 2004. № 25. P. 287–290.
Chaudhary I.J., Rathore D. Dust pollution: its removal and effect on foliage physiology of urban trees // Sustain. Cit. Soc. 2019. V. 51. Article 101696.
Chelli-Chaaboimi A. Mechanisms and adaptation of plants to environmental stress: a case of woody species // Physiological mechanisms and adaptation strategies in plants under changing environment. New York: Springer-Verlag New York Inc., 2013. P. 1–18.
Cudlín P. Kieliszewska-Rojucka B., Rudawska M. et al. Fine roots and ectomycorrhizas as indicators of environmental change // Plant Biosyst. 2007. V. 141. P. 406–425.
Danek M. The influence of industry on Scots pine stands in the south-eastern part of the Silesia-Krakow Upland (Poland) on the basis of dendrochronological analysis // Water Air Soil Poll. 2007. V. 185. P. 265–277.
Das S., Prasad P. Seasonal variation in air pollution tolerance indices and selection of plant species for industrial areas of Rourkela // Indian J. Environ. Prot. 2010. V. 30. № 12. P. 978–988.
Devi E.L., Kumar S., Singh T.B. et al. Adaptation strategies and defence mechanisms of plants during environmental stress // Medicinal plants and environmental challenges. Cham: Springer Internat. Publishing, 2017. P. 359–413.
Dineva S.B. Comparative studies of the leaf morphology and structure of white ash Fraxinus americana L. and London plane tree Platanus acerifolia Willd growing in polluted area // Dendrobiology. 2004. V. 52. P. 3–8.
Dineva S.B. Development of the leaf blades of Acer platanoides in industrially contaminated environment // Dendrobiology. 2006. V. 55. P. 25–32.
Dineva S.B. Leaf blade structure and the tolerance of Acer negundo L. (box elder) to the polluted environment // Dendrobiology. 2005. V. 53. P. 11–16.
Duan J., Fu B., Kang H., Song Z., Wei A. Response of gas-exchange characteristics and chlorophyll fluorescence to acute sulfur dioxide exposure in landscape plants // Ecotoxicol. Environ. Safety. 2019. V. 171. Iss. 30. P. 122–129.
Freer-Smith P.H., Taylor G. Comparative evaluation of the effects of gaseous pollutants, acidic deposition and mineral deficiencies on gas exchange of trees // Agricult.Ecosyst. Environ. 1992. V. 42. Iss. 3–4. P. 321–332.
Gao F., Calatayud V., García-Breijo F. et al. Effects of elevated ozone on physiological, anatomical and ultrastructural characteristics of four common urban tree species in China // Ecol. Indicat. 2016. V. 67. P. 367–379.
Godek M., Sobik M., Błaś M. et al. Tree rings as an indicator of atmospheric pollutant deposition to subalpine spruce forests in the Sudetes (Southern Poland) // Atmosph. Res. 2015. V. 151. P. 259–268.
Gowin T., Goral I. Chlorophyll and pheophytin content in needles of different age of trees growing under conditions of chronic industrial pollution // Acta Societ. Botan. Polon. 1977. V. 46. № 1. P. 151–159.
Grime J.P. Plant strategies and vegetation processes. Chichester: Wiley and Sons, 1979. 222 p.
Helmisaari H.S., Makkonen K., Olsson M. et al. Fine-root growth, mortality and heavy metal concentrations in limed and fertilized Pinus silvestris (L.) stands in the vicinity of a Cu-Ni smelter in SW Finland // Plant Soil. 1999. V. 209. P. 193–200.
Jahan S., Iqbal M. Z. Morphological and anatomical studies of leaves of different plants affected by motor vehicles exhaust // J. Islam. Acad. Sci. 1992. V. 5. № 1. P. 21–23.
Jakovljević T., Marchetto A., Lovreškov L. et al. Assessment of atmospheric deposition and vitality indicators in mediterranean forest ecosystems // Sustainability. 2019. V. 11. Iss. 23. P. 6805.
Jochner S., Markevych I., Beck I. et al. The effects of short-and long-term air pollutants on plant phenology and leaf characteristics // Environ. Poll. 2015. V. 206. P. 382–389.
Joshi P.C., Swami A. Physiological responses of some tree species under roadside automobile pollution stress around city of Haridwar, India // Environmentalist. 2007. № 27. P. 365–374.
Joshi P.C., Swami A. Air pollution induced changes in the photosynthetic pigments of selected plant species // J. Environ. Biol. 2009. № 30. P. 295–298.
Jyothi S.J., Jaya D.S. Evaluation of air pollution tolerance index of selected plant species along roadsides in Thiruvananthapuram, Kerala // J. Environ. Biol. 2010. V. 31. P. 379–386.
Kahle H. Response of roots of trees to heavy metals // Environ. Experim. Botan. 1993. V. 33. P. 99–119.
Klamerus-Iwan A., Błońska E., Lasota J. et al. Seasonal variability of leaf water capacity and wettability under the influence of pollution in different city zones // Atmosph. Poll. Res. 2018. V. 9. Iss. 3. P. 455–463.
Kosiba P. Variability of morphometric leaf traits in small-leaved linden (Tilia cordata Mill.) under the influence of air pollution // Acta Soc. Botan. Polon. 2008. V. 77. Iss. 2. P. 125–137.
Kozlowski T.T., Pallardy S.G. Acclimation and adaptive responses of woody plants to environmental stresses // Botan. Rev. 2002. V. 68. Iss. 2. P. 270–334.
Kuang Y.W., Sun F.F., Wen D.Z. et al. Tree-ring growth patterns of Masson pine (Pinus massoniana L.) during the recent decades in the acidification Pearl River Delta of China // Forest Ecol. Manag. 2008. V. 255. P. 3534–3540.
Laghari1 S.K., Zaidi1 M.A., Razaq G. Impact of solid waste burning air pollution on some physio-anatomical characteristics of some plants // Pakistan J. Botan. 2015. V. 47 (1). P. 225–232.
Lakshmi P.S., Sravanti K.L., Srinivas N. Air pollution tolerance index of various plant species growing in industrial areas // The Ecoscan. 2008. V. 2 (2). P. 203–206.
Lamoreaux R.J., Chaney W.R. Photosynthesis and transpiration of excised silver maple leaves exposed to cadmium and sulphur dioxide // Environ. Poll. 1978. V. 17. Iss. 4. P. 259–268.
Leghari S.K., Asrar M. Effect of air pollution on the leaf morphology of common plant species of Quetta city // Pakistan J. Botan. 2013. № 45. P. 447–454.
Li P., Yin R., Shang B. et al. Interactive effects of ozone exposure and nitrogen addition on tree root traits and biomass allocation pattern: an experimental case study and a literature meta-analysis // Sci. Tot. Environ. 2020. № 710. Article № 136379.
Liu Y., Ding H. Variation in air pollution tolerance index of plants near a steel factory: implications for landscape-plant species selection for industrial areas // WSEAS Trans. Environ. Dev. 2008. V. 4. Iss. 1. P. 24–32.
Łuszczyńska K., Wistuba M., Malik I. Reductions in tree-ring widths of silver fir (Abies alba Mill.) as an indicator of air pollution in southern Poland // Environ. Socio-econ. Stud. 2018. V. 6. Iss. 3. P. 44–51.
Lux A., Martinka M., Vaculik M., White P.J. Root responses to cadmium in the rhizosphere: a review // J. Experim. Botan. 2011. V. 62. P. 21–37.
Lux A., Šottníková A., Opatrná J., Greger M. Differences in structure of adventitious roots in Salix clones with contrasting characteristics of cadmium accumulation and sensitivity // Physiol. Plantarum. 2004. V. 120. P. 537–545.
MacArthur R.H., Wilson E.D. The theory of island biogeography. New Jersey: Princeton Univ. Press, 1967. 203 p.
Hermy M., Stieperaete H. Capitalists and preoletarians (MacLeod, 1884): an early theory of plant strategies // Oikos. 1985. V. 44. № 2. P. 364–366.
Masuch G., Franz J.T., Kettrup A. Histological and biochemical differences of slightly and severely injured spruce needles of two stands in northrhine westphalia // Environ. Experim. Botan. 1992. V. 32. Iss. 2. P. 163–182.
Matysiak R. Content of carotenoids in needles of Pinus sylvestris L. growing in a polluted area // Dendrobiology. 2001. № 46. P. 39–42.
McDermot C.R., D’Amico V., Trammell T. Sensitivity of stomate size in red maple (Acer rubrum L.) trees in deciduous forests to urban conditions // Preprints. 2020. 2020040235. https://doi.org/10.20944/preprints202004.0235.v1
Meerabai G., Ramana V.C., Rasheed M. Effect of industrial pollutants on physiology of Cajanus cajan (L.) – Fabaceae // Int. J. Environ. Sci. 2012. V. 2. № 4. P. 1889–1894.
Mena-Petite A., Duñabeitia M.K., Gonzalez-Moro B. et al. Sequential effects of acidic precipitation and drought on water relations of Pinus radiata seedlings // J. Plant Physiol. 1999. V. 155. Iss. 1. P. 93–100.
Mikhailova T.A., Afanasieva L.V., Kalugina O.V. et al. Changes in nutrition and pigment complex in pine (Pinus sylvestris L.) needles under technogenic pollution in Irkutsk region, Russia // J. Forest Res. 2017. V. 22. № 6. P. 386–392.
Mitu K.J., Islam M.A., Biswas P. et al. Effects of different environmental pollutants on the anatomical features of roadside plants // Progress. Agric. 2019. № 30 (4). P. 344–351.
More R.S., Chaubal S.S. Determination of stress and comparison by estimation of chlorophyll – a, b and carotenoid contents among plants growing along Mithi river, Mumbai // Int. J. Sci. Engin. Res. 2017. V. 8. Iss. 1. P. 1–8.
Mukherjee A., Agrawal M. Use of GLM approach to assess the responses of tropical trees to urban air pollution in relation to leaf functional traits and tree characteristics // Ecotoxicol. Environ. Safety. 2018. V. 152. Iss. 15. P. 42–54.
Newman G.S., Hart S.C. Nutrient covariance between forest foliage and fine root // Forest Ecol. Manag. 2006. № 2–3. P. 136–141.
Nyborg M., Crepin J., Hocking D., Baker J. Effect of sulphur dioxide on precipitation and on the sulphur content and acidity of soils in Alberta, Canada // Proc. of the first Internat. symp. on “Acid Precipitation and the Forest Ecosystem”. Columbus: Ohio State University, 1975. P. 767–778.
Paavilainen E. The effect of fertilisation on the root systems of swamp pine stands // Folia Forest Polonica. 1967. № 31. P. 1–9.
Paolettia E., Grulke N.E. Does living in elevated CO2 ameliorate tree response to ozone? A review on stomatal responses // Environ. Poll. 2005. V. 137. P. 483–493.
Pärn H., Mandre M. Dendrochronological analysis of the growth and growth-climate relationships of conifers in the region of alkaline dust deposition // Forest Ecol. Manag. 2011. № 262. P. 88–94.
Perone A., Cocozza C., Cherubini P. et al. Oak tree-rings record spatial-temporal pollution trends from different sources in Terni (Central Italy) // Environ. Poll. 2018. V. 233. P. 278–289.
Pourkhabbaz A., Rastin N., Olbrich A. et al. Influence of environmental pollution on leaf properties of urban plane trees, Platanus orientalis L. // Bull. Environ. Contam. Toxicol. 2010. № 85. P. 251–255.
Prajapati S.K., Tripathi B.D. Seasonal variation of leaf dust accumulation and pigment content in plant species exposed to urban particulates pollution // J. Environ. Qual. 2008. V. 37. P. 865–870.
Prusty B.A.K., Mishra P.C., Azeez P.A. Dust accumulation and leaf pigment content in vegetation near the national highway at Sambalpur, Orissa, India // Ecotoxicol. Environ. Saf. 2005. V. 60. P. 228–235.
Puhe J. Growth and development of the root system of Norway spruce (Picea abies) in forest stands – a review // Forest Ecol. Manag. 2003. V. 175. P. 253–273.
Qadir S.U., Raja V., Siddiqui W.A. Morphological and biochemical changes in Azadirachta indica from coal combustion fly ash dumping site from a thermal power plant in Delhi, India // Ecotoxicol. Environ. Saf. 2016. V. 129. P. 320–328.
Rashidi F., Jalili A., Kafaki S.B. et al. Anatomical responses of leaves of Black Locust (Robinia pseudoacacia L.) to urban pollutant gases and climatic factors // Trees. 2012. № 26. P. 363–375.
Rostunov A., Konchina T., Zhestkova E. et al. The dependence of morphological and physiological indicators of the leaves of woody plants on the degree of technogenic pollution // Environment. Technology. Resources / Proc. of the 11th Internat. sci. and pract. conf. V. I. Latvia: Rezekne Academy of Technologies, 2017. P. 235–239.
Rutkiewicz P., Malik I. Spruce tree-ring reductions in relation to air pollution and human diseases a case study from Southern Poland // Environ. Socio-Econ. Stud. 2018. V. 6. Iss. 2. P. 22–28.
Safford L.O. Seasonal variation in the growth and nutrient content of yellow-birch replacement roots // Plant Soil. 1976. V. 44. № 2. P. 439–444.
Schekalev R.V., Martynyuk А.А., Melekhov V.I. Variability properties of Pinus sylvestris L. wood in growing stock under technogenic impact // Изв. вузов. Лесн. журн. 2020. № 4. С. 113–122.
Schreuder M., van Hove L.W.A., Brewer C.A. Ozone exposure affects leaf wettability and tree water balance // New Phytol. 2001. V. 152 (3). P. 443–454.
Sensuła B., Opała M., Wilczýnski S., Pawełczyk S. Long- and short-term incremental response of Pinus sylvestris L. from industrial area nearby steelworks in Silesian Upland, Poland // Dendrochronologia. 2015. V. 36. P. 1–12.
Sensuła B., Wilczyński S., Monin L. et al. Variations of tree ring width and chemical composition of wood of pine growing in the area nearby chemical factories // Geochronometria. 2017. V. 44. P. 226–239.
Seyyednejad S.M., Koochak H. Some morphological and biochemical responses due to industrial air pollution in Prosopis juliflora (Swartz) DC plant // African J. Agr. Res. 2013. V. 8 (18). P. 1968–1974.
Seyyednejad S.M., Niknejad M., Koochak H. A review of some different effects of air pollution on plants // Res. J. Environ. Sci. 2011. №. 5 (4). P. 302–309.
Seyyednejad S.M., Niknejad M., Yusefi M. Study of air pollution effects on some physiology and morphology factors of Albizia lebbeck in high temperature condition in Khuzestan // J. Plant Sci. 2009a. V. 4. P. 122–126.
Seyyednejad S.M., Niknejad M., Yusefi M. The effect of air pollution on some morphological and biochemical factors of Callistemon citrinus in petrochemical zone in South of Iran // Asian J. Plant Sci. 2009b. № 8. P. 562–565.
Shaheen A.M., Al-Toukhy A.A., Hajar A.S. Effect of air pollution on leaf traits of three tree species growing in the industrial zone of Jeddah, Saudi Arabia // Meteorol. Environ. Arid Land Agric. Sci. 2016. V. 26. № 2. P. 33–40.
Shrestha S.D., Devkota A., Jha P.K. Assessment of air pollution impact on micro-morphological and biochemical properties of Callistemon citrinus (Curtis) Skeels and Lagerstroemia indica L. // Sci. World. 2021. V. 14. № 14. P. 132–140.
Singh H., Savita, Sharma R. et al. Physiological functioning of Lagerstroemia speciosa L. under heavy roadside traffic: an approach to screen potential species for abatement of urban air pollution // Biotech. 2017. V. 7 (1). P. 61.
Skirycz A., Inzé D. More from less: plant growth under limited water // Curr. Opin. Biotechnol. 2010. V. 21. P. 197–203.
Skrynetska I., Ciepał R., Kandziora-Ciupa M. et al. Ecophysiological responses to environmental pollution of selected plant species in an industrial urban area // Int. J. Environ. Res. 2018. V. 12. P. 255–267.
Skuterud L., Goltsova N.I., Naeumann R. et al. Histological changes in Pinus sylvestris L. in the proximal-zone around the Chernobyl Power // Plant Sci. Environ. 1994. V. 157. P. 387–397.
Song Y.J., Kim Y.D., Choi K.R. Estimation of the effects of air pollutants on tree ring growth in black pines (Pinus thunbergii) // J. Ecol. Environ. 2009. V. 32. Iss. 2. P. 109–113.
Šrámek V. SO2 air pollution and forest health status in Northwestern Czech Republic // Chemosphere. 1998. V. 36. Iss. 4–5. P. 1067–1072.
Stravinskiene V., Bartkevicius E., Plausinyte E. Dendrochronological research of Scots pine (Pinus sylvestris L.) radial growth in vicinity of industrial pollution // Dendrochronologia. 2013. V. 31. P. 179–186.
Swain S., Mallick S.N., Prasad P. Effect of industrial dust deposition on photosynthetic pigment chlorophyll and growth of selected plant species in Kalunga industrial areas, Sundargarh, Odisha // Int. J. Botan. Stud. 2016. V. 1. Iss. 5. P. 1–5.
Thambavani S.D., Sabitha M. A. The spectral determination of chlorophylls a, b and total carotenoids using various solvents for tree species growing near sugar mill // Asian J. Experim. Chem. 2012. V. 7. Iss. 1. P. 5–9.
Tripathi A.K., Gautam M. Biochemical parameters of plants as indicators of air pollution // J. Environ. Biol. 2007. № 28 (1). P. 127–132.
Uka U.N., Belford E.J.D. Response of roadside tree leaves in a tropical city to automobile pollution // Notulae Sci. Biol. 2020. № 12 (3) P. 752–768.
Urazgil’din R.V., Amineva K.Z., Zaitsev G.A., Kulagin A.Yu. Comparative characteristics of pine, spruce and larch pigmental complex seasonal variability in industrial pollution conditions // Ecology and geography of plants and plant communities / The fourth Internat. sci. conf. on ecology and geography of plants and plant communities. 2018. V. 2018. P. 232–242.
Vacek S., Bílek L., Schwarz O. et al. Effect of air pollution on the health status of spruce stands // Moun. Res. Dev. 2013. V. 33. № 1. P. 40–50.
Wang H., Ouyang Z., Chen W. et al. Water, heat, and airborne pollutants effects on transpiration of urban trees // Environ. Poll. 2011. V. 159. Iss. 8–9. P. 2127–2137.
Wang H., Wang X., Zhao P. et al. Transpiration rates of urban trees, Aesculus chinensis // J. Environ. Sci. 2012. V. 24. Iss. 7. P. 1278–1287.
Wang H., Ouyang Z., Chen W. et al. Transpiration characteristics of chinese pines (Pinus tabulaeformis) in an urban environment // Designing low carbon societies in landscapes / Eds N. Nakagoshi, A. J. Mabuhay. Tokyo: Springer, 2014. P. 57–71.
Wang J., Farooq T.H., Aslam A. et al. Non-targeted metabolomics reveal the impact of phenanthrene stress on root exudates of ten urban greening tree species // Environ. Res. Available online 22 October 2020. https://doi.org/10.1016/j.envres.2020.110370
Wertz B. Dendrochronological evaluation of the impact of industrial imissions on main coniferous species in the Kielce Upland // Sylwan. 2012. V. 156. Iss. 5. P. 379–390.
Woo S.Y., Je S.M. Photosynthetic rates and antioxidant enzyme activity of Platanus occidentalis growing under two levels of air pollution along the streets of Seoul // J. Plant Biol. 2006. № 49. P. 315–319.
Woo S.Y., Lee D.K., Lee Y.K. Net photosynthetic rate, ascorbate peroxidase and glutathione reductase activities of Erythrina orientalis in polluted and non-polluted areas // Photosynthetica. 2007. № 45. P. 293–295.
Woodwell G.M., Miller L.N. Chronic gamma radiation affects the distribution of radial increment in Pinus rigida stems // Science. 1963. V. 139. № 3551. P. 222–223.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии