

Водные ресурсы, 2021, T. 48, № 2, стр. 164-172
Цветение водохранилища многолетнего регулирования стока
К. К. Эдельштейн a, Ю. С. Даценко a, b, *, В. В. Пуклаков a
a Московский государственный университет им. М.В. Ломоносова
119991 Москва, Россия
b Институт водных проблем РАН
119333 Москва, Россия
* E-mail: yuri0548@mail.ru
Поступила в редакцию 11.11.2019
После доработки 18.08.2020
Принята к публикации 25.09.2020
Аннотация
На основе многолетних наблюдений и модельных расчетов рассмотрены факторы развития фитопланктона в стратифицированном Можайском водохранилище. Для анализа изменчивости величин биомассы фитопланктона использован индекс погодной контрастности, основанный на выделении синоптических циклов температуры водной поверхности. Сравнительная оценка интенсивности цветения в разные годы проведена на основе индекса цветения воды, рассчитанного как сумма ежесуточных величин биомассы фитопланктона, полученных по модельному расчету. Проанализированы особенности развития диатомовых водорослей и цианобактерий в водохранилище в зависимости от гидрометеорологических условий и многолетние изменения индексов цветения воды и погодной контрастности.
ВВЕДЕНИЕ
В настоящей работе под цветением водохранилища авторы понимают явление интенсивного развития в нем фитопланктона, при котором биомасса водорослей в поверхностном трофогенном слое достигает величины ≥1 г/м3. В этом случае на водопроводных станциях питьевого и промышленного водоснабжения возникает необходимость интенсификации очистки воды [10]. Исследование фитопланктона на Можайском водохранилище, головном водоеме в Москворецкой системе источников водоснабжения столицы, началось в первый год заполнения его ложа водой Верхней Москвы-реки [1]. Площадь водоема 31 км2, объем 0.235 км3, длина 28 км, максимальная глубина 22 м. В межень средняя скорость стокового течения составляет несколько сантиметров в секунду [19]. Поэтому ежегодно водохранилище термически и гидрохимически стратифицировано в вегетационный сезон, что типично для слабопроточных водохранилищ.
С 1965 г. регулярные альгологические наблюдения проводятся на 5–10 вертикалях в верхнем, среднем и приплотинном участках водохранилища [12–15, 17, 21]. В России столь продолжительные наблюдения за цветением воды ведутся лишь на двух водохранилищах – Можайском и Рыбинском.
На основе имеющейся базы данных по гидрохимическим и альгологическим показателям разработана математическая модель гидроэкологического режима водохранилища.
На первом этапе ее разработки определялся генетический состав воды в расчетных отсеках. В них в результате смешения речных и водохранилищных вод создавались водные массы, положение которых в водоеме менялось в течение года, что и было представлено в виде одномерной многосекционной воднобалансовой модели СВМ (Состав Водных Масс) [7, 8, 32]. Затем эта модель была трансформирована в двухмерную в вертикальной плоскости боксовую модель термогидродинамических процессов в водохранилище – модель ТМО (тепло- и массообмен) [18], которая модернизирована в многосекционную боксовую гидрологическую модель водохранилища с экологическим блоком – модель ГМВ-МГУ, описание алгоритма и результатов верификации которой опубликованы в [2]. В экологическом блоке этой модели рассчитываются ежедневные гидрологические характеристики, гидрохимические показатели воды и биомасса водорослей разных отделов в метровых боксах 18 секций (отсеков), на которые разделяется вся толща воды Можайского водохранилища.
И по натурным данным, и по модельному расчету в Можайском водохранилище четко выделяются ежегодный весенний пик цветения преимущественно диатомовыми водорослями, следующая за ним фаза “чистой воды” и продолжительный период летне-осеннего интенсивного развития цианобактерий, что характерно для большинства водохранилищ России [26]. В отдельные годы летне-осеннее цветение в водохранилищах Москворецкой системы было настолько интенсивным, что приводило к необходимости переключать подачу воды в Москву с особенно сильно зацветшего водохранилища на водохранилища с наименьшим цветением [24].
Годовые величины рассчитанной по модели суммарной биомассы планктонных водорослей в водохранилище определяются биомассой цианобактерий (коэффициент корреляции между ними – r = 0.96, n = 55 лет, р = 0.95) [6]. Поэтому в данной работе предпринята попытка установить причину вариации интенсивности цветения воды в Можайском водохранилище цианобактериями в течение почти 60-летнего его существования (1961–2018 гг.) в зависимости от изменчивости погодных условий. Для этого использованы результаты полевых исследований и модельных расчетов по ГМВ-МГУ ежесуточных величин биомассы цианобактерий и температуры воды верхнего трофогенного слоя центрального плеса Можайского водохранилища.
МЕТОДЫ РАСЧЕТА ИНДЕКСА КОНТРАСТНОСТИ ПОГОДЫ И ИНДЕКСА ЦВЕТЕНИЯ ВОДЫ
В безледный период на водоемах умеренных широт ежегодно из-за изменчивости погоды наблюдается чередование повышений температуры водной поверхности и ее понижений. Это вызывается совокупным воздействием меняющихся величин поступающей солнечной радиации, облачности, ветра, температуры и влажности воздуха. Два 70-суточных периода автоматизированных гидрометеорологических наблюдений в центре Можайского водохранилища (с 26.06.2016 по 3.09.2016 и в те же даты 2017 г.) разделены на фазы нагревания и охлаждения поверхности воды, каждая пара которых образует погодный цикл гидроэкологического режима водоема. За фазы погодных циклов авторами принят период времени изо дня в день повышающейся или понижающейся температуры поверхностного метрового слоя воды на ≥3°С в течение ≥3 сут [4]. Это позволяет при оценке длительности фазы не учитывать пространственную неоднородность поля температуры (по многолетним наблюдениям, в пределах плеса температура ≤3°С) и пренебречь ее внутрисуточной изменчивостью при неизменной погоде. По данным регистрации температуры в 2016 г. выделено 4 погодных цикла (8 фаз), а в 2017 г. – только 2 (4 фазы). В эти же периоды впервые были проведены автоматизированные наблюдения за интенсивностью фотосинтеза фитопланктона с использованием продукцимера [3]. Регистрация содержания кислорода и температуры воды двумя оксиметрами продукцимера с дискретностью 15 мин в темном и светлом сосудах, вода в которых автоматически обновлялась через каждые 3 ч, велась в течение 70 сут на горизонте 0.5 м над затопленным руслом р. Москвы (глубина 12 м). Одновременно судовой метеостанцией непрерывно регистрировались метеохарактеристики на высоте 2 м над водной поверхностью и термокосой – температура воды на шести горизонтах этой вертикали. Интенсивность фотосинтетически активной радиации (ФАР) регистрировалась ФАР-логером на высоте 10 м над берегом. Наиболее показательно изменчивость метеохарактеристик проявлялась в повышении и понижении температуры воды поверхностного слоя. Вся совокупность выше перечисленных метеохарактеристик определяет тепловой баланс этого слоя и, следовательно, изменение температуры воды с изменением любой из этих характеристик, поэтому именно эта температура использовалась для расчета предложенного индекса погодной контрастности (ИПК) [29]:
(1)
${\text{ИПК}} = \frac{{\sum\limits_1^n {{{{\left| {\Delta T} \right|}}^{i}}} }}{{\left[ {\left( {{{T}_{{\max {\text{рп}}}}} - {{T}_{{in~\,{\text{рп}}}}}} \right) + {\text{\;}}\left( {{{T}_{{\max {\text{рп}}}}}{\text{\;}} - {{T}_{{f~{\text{рп}}}}}} \right)} \right]}},$Чем сильнее изменчивость температуры фазы и чем фаз больше в течение расчетного периода, тем контрастнее выражены чередования погодных циклов, что и характеризуется величиной ИПК.
Расчет ИПК для 70-суточных периодов 2016 и 2017 гг. показал, что в 2016 г. ИПК был в 2.2 раза больше, чем в 2017 г. При этом максимальные часовые величины валовой первичной продукции в 2016 г. оказались в 1.6 раза выше, а среднесуточные за фазу – в 1.8 раз выше, чем в 2017 г. Сравнение средних величин продукции за расчетные периоды этих лет показало, что в 2016 г. она была в 1.5 раза выше, чем в 2017 г. [29]. Такое соотношение ИПК и средней за период валовой первичной продукции свидетельствует о влиянии изменчивости погоды на продуктивность водохранилища. В оба года максимальные величины валовой первичной продукции получены в такие теплые фазы, которым предшествовало наибольшее охлаждение водной толщи плеса.
Различие погодных фаз гидроэкологического режима водохранилища проявляется в том, что в фазы охлаждения и нагревания внутренний водообмен плеса существенно различен. В фазы охлаждения воды в холодную, ветреную и пасмурную погоду возникают вихри Ленгмюра. Эта вихреобразная циркуляция перемешивает водную толщу до большой глубины и может достигать дна. Это перемешивание приводит к выносу в трофогенный слой большого количества бескислородных, но насыщенных продуктами деструкции детрита и органики в илах глубинных вод. В фазы нагревания при штилевой и безоблачной погоде в темное время суток эпилимнион перемешивается ячеистой конвекцией при охлаждении воды вплоть до восхода солнца. В это время в нем образуются цилиндрообразные ячейки Бенара. По их периферии в охлаждающемся поверхностном слое вода опускается до термоклина в слое температурного скачка, а в центре ячеек возникает восходящий ток воды к водной поверхности. Возникает так называемый малый биогидрохимический круговорот [27]. С восходящими токами из нижних слоев эпилимниона к поверхности выносится растворенный диоксид углерода (СО2) и биогенные вещества, в том числе фосфор. В первую же половину светлого времени суток биогенные вещества поглощаются водорослями в процессе фотосинтеза.
Автоматизированные наблюдения показали, что одним из факторов межгодовой изменчивости продукции цианобактерий в летне-осенний сезон разных лет могут быть частота и контрастность изменчивости погоды в сезон цветения. По данным многолетних наблюдений за температурным режимом водохранилища (1961–2001гг.), число погодных циклов в среднем за вегетационный сезон составляло 6 и варьировало от 3 до 9 в отдельные годы [4].
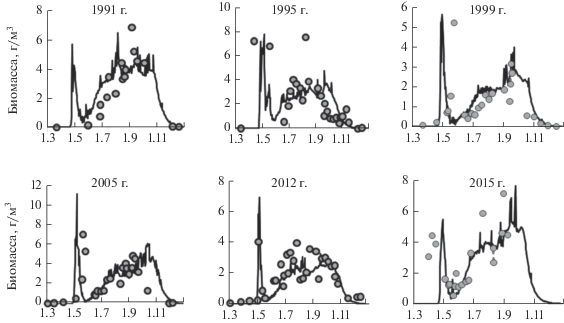
Чтобы проверить реальность гипотезы о влиянии контрастности погоды на интенсивность цветения воды в водохранилище, по модели ГМВ-МГУ выполнен расчет ежедневных величин биомассы цианобактерий и диатомовых водорослей для 58 лет существования водохранилища. Примеры модельного воспроизведения режима цветения водорослей в приплотинном плесе Можайского водохранилища в годы, близкие к средним по притоку воды летом при жаркой либо прохладной погоде, приведены на рис. 1. По результатам расчетов для каждого года был выделен сезон цветения цианобактерий, который начинался с момента достижения их биомассы 1 мг/л ранним летом и заканчивался при уменьшении этой величины до 1 мг/л осенью. Для сравнительной оценки интенсивности цветения в разные годы предложен индекс цветения воды (ИЦВ), равный сумме ежедневных величин биомассы фитопланктона за весь период их цветения [25]:
(2)
${\text{ИЦВ}} = \mathop \sum \limits_{{{\tau }_{1}}}^{{{\tau }_{2}}} B_{{{\text{phyt}}}}^{i}({\text{г\;}}{{{\text{м}}}^{{ - {\text{3}}}}}),$Рис. 1.
Рассчитанные по модели (черная линия) и наблюденные (точки) величины биомассы фитопланктона в поверхностном слое приплотинного плеса Можайского водохранилища.
Расчетный период (РП) для оценки изменчивости погоды по полученным в гидродинамическом блоке той же модели среднесуточным значениям температуры воды на горизонте 1 м начинался с момента достижения температуры поверхностного слоя воды 15°С, после которой в водоеме, как правило, формируется устойчивый слой температурного скачка. Заканчивался этот период, когда среднесуточная температура поверхностного слоя снижалась до 10°С. Весь этот период подразделялся на фазы нагревания и фазы охлаждения воды так же, как это было сделано ранее для расчета ИПК за 70-суточные периоды 2016 и 2017 гг. Для всех лет существования водохранилища для каждого расчетного периода получен интегральный индекс погодной контрастности (ИИПК):
РЕЗУЛЬТАТЫ И ОБСУЖЕНИЕ
Весеннее цветение водохранилища
Весеннее цветение Можайского водохранилища связано с обильным развитием диатомовых водорослей. Оно начинается обычно в апреле, наиболее ранняя дата начала цветения – 7 апреля, наиболее поздняя – 16 мая. Минимальная продолжительность периода этого цветения – 4, максимальная – 28 сут. По длине водохранилища четко прослеживается неодновременность начала цветения, а затем и его окончания – от верховий водохранилища к плотине с отставанием в 3–4 дня. Например, если на верхнем участке водохранилища цветение наступает 26 апреля, то в центральной части (Красновидовский плес) – 29 апреля, а в приплотинном плесе – 3 мая. Окончание цветения происходит в той же последовательности. Более раннее цветение верхних участков водохранилища связано с тем, что в них поступают биогенные вещества с речными водами, они мелководны, хорошо прогреваются и перемешиваются.
Модельные расчеты показали, что между биомассой диатомовых водорослей и факторами окружающей среды немного статистически значимых связей. Из 18 рассмотренных авторами гидрометеорологических, гидрохимических и водохозяйственных показателей, которые могли бы оказывать влияние на цветение диатомовых водорослей, высоко значимая корреляция индекса цветения диатомовых водорослей (ИЦВд) обнаружена только с коэффициентом замедленности весеннего наполнения: Кτ = τ/τ0 (τ и τ0 – длительность половодья данного года и средняя многолетняя его длительность, сут (r = 0.45, р = 0.95)). Эта связь показывает, что чем медленнее происходит весеннее наполнение водохранилища, тем выше средняя биомасса диатомовых. Связано это, вероятно, с тем, что при слабом развитии весенних процессов на водосборе большая часть весенних вод поступает в водоем в виде богатого биогенами почвенного, а не склонового стока, характерного для бурной весны. Кроме того, при длительной холодной весне в водоеме дольше поддерживаются благоприятные для холоднолюбивых диатомовых водорослей температурные условия.
Рассчитанный ход изменений биомассы диатомовых водорослей характеризуется острыми пиками. При этом ее максимальные величины имеют более тесную статистическую связь с рассматриваемыми предикторами, чем ее средние значения в поверхностном слое.
ИЦВд очень слабо коррелирует с количеством штилевых дней и рассчитанным по модели притоком фосфора и азота с водосбора водохранилища за время цветения воды диатомовыми водорослями. Со всеми остальными факторами связь ИЦВд статистически незначима. Также незначительно зависит от абиотических факторов и продолжительность периода цветения диатомовых водорослей, что еще раз подтверждает малую предсказуемость цветения воды этой группой фитопланктона.
Оценка зависимости ИЦВд от концентрации кремния не анализировалась, поскольку по многократным наблюдениям, например в 2018 г. по данным шести съемок (с февраля по сентябрь), его содержание в основной водной массе водохранилища варьировало в диапазоне 0.8–3.0 мг/л весной и летом, а в зимней водной массе увеличивалось до 6 мг/л. В другие годы содержание в воде кремния весной достигало даже 10–12 мг/л и выше.
Подсчеты ИЦВд для диатомовых водорослей в приплотинном плесе водохранилища показали исключительно большую межгодовую изменчивость этого показателя за рассматриваемый многолетний период. Коэффициент вариации ИЦВд = 37%, продолжительности периода цветения – 31%.
Если уменьшать интенсивность наполнения водохранилища увеличением сброса воды из него в весенний период, то увеличатся продольный водообмен в период половодья и вынос из водохранилища насыщающих водную толщу биогенных веществ и развивающегося фитопланктона, что может привести к ослаблению весеннего цветения.
Летне-осеннее цветение водохранилища
Индекс цветения цианобактериями (ИЦВцб) более тесно связан с большинством анализируемых факторов. Высоко значима корреляционная связь ИЦВцб со следующими факторами: четырьмя гидрологическими – объемами притока воды в половодье (r = –0.49) и во время летне-осенних паводков (r = 0.42), коэффициентом водообмена водохранилища в половодье (r = –0.37), скоростью переноса водных масс (r = 0.44); двумя метеорологическими – скоростью ветра (r = –0.40) и количеством штилевых дней (r = 0.45); четырьмя гидрохимическими – притоком с водосбора фосфора (r = 0.93) и азота (r = 0.92), поступлением из донных отложений фосфора (r = 0.36) и азота (r = 0.34); двумя водохозяйственными – соотношением объема воды в водохранилище в конце его зимней сработки и объема вод половодья (r = = 0.50), предвесенней сработкой водохранилища (r = –0.36).
Менее значима корреляционная связь ИЦВцб с еще четырьмя факторами: с поступлением солнечной радиации, температурой воды в период летнего цветения, слоем атмосферных осадков и интенсивностью наполнения водохранилища. Разнообразие факторов и их совместное влияние на экосистему водохранилища, нередко противоречивое, не позволяет объяснить значительную межгодовую изменчивость цветения в летне-осенний сезон [25].
Цветение воды в водохранилище цианобактериями многократно продолжительнее, чем весеннее цветение, и поэтому ИЦВцб составляет бóльшую величину в суммарном индексе цветения водохранилища водорослями всех отделов планктона во все годы существования водохранилища. Межгодовая изменчивость интенсивности летне-осеннего цветения, рассчитанная по результатам моделирования режима биомассы фитопланктона за 1961–2018 гг. (рис. 2), и частота чередования теплой и холодной погоды в сезон цветения этих водорослей, характеризуемая ИИПК (табл. 1), послужили основой для проверки гипотезы хорошей связи между этими двумя показателями.
Рис. 2.
Изменение от года к году ИЦВцб, ИИПК в центральном плесе Можайского водохранилища и линейный тренд ИЦВцб.
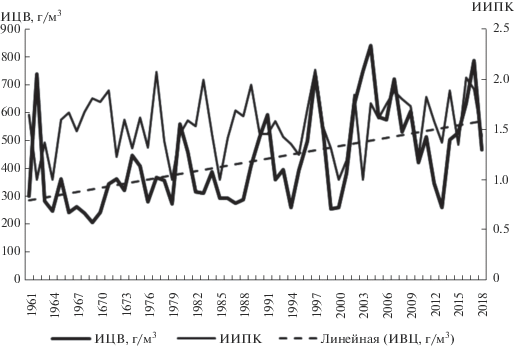
Таблица 1.
Продолжительность сезона цветения синезеленых водорослей t, сут, число погодных фаз Nф, ИИПК и ИЦВ, г/м3, в центральном плесе Можайского водохранилища (по данным модельных расчетов)
| Год | t | Nф | ИИПК | ИЦВ | Год | t | Nф | ИИПК | ИЦВ |
|---|---|---|---|---|---|---|---|---|---|
| 1961 | 144 | 6 | 1.64 | 301 | 1990 | 102 | 4 | 1.45 | 514 |
| 1962 | 133 | 2 | 1.00 | 737 | 1991 | 152 | 4 | 1.45 | 591 |
| 1963 | 174 | 4 | 1.36 | 282 | 1992 | 120 | 4 | 1.58 | 359 |
| 1964 | 124 | 2 | 1.00 | 247 | 1993 | 108 | 4 | 1.43 | 395 |
| 1965 | 125 | 10 | 1.59 | 361 | 1994 | 130 | 4 | 1.36 | 259 |
| 1966 | 97 | 6 | 1.67 | 241 | 1995 | 150 | 4 | 1.25 | 393 |
| 1967 | 132 | 4 | 1.48 | 263 | 1996 | 145 | 6 | 1.70 | 498 |
| 1968 | 119 | 4 | 1.67 | 239 | 1997 | 123 | 8 | 2.09 | 734 |
| 1969 | 108 | 4 | 1.81 | 206 | 1998 | 128 | 4 | 1.52 | 535 |
| 1970 | 132 | 6 | 1.77 | 242 | 1999 | 184 | 4 | 1.31 | 254 |
| 1971 | 129 | 6 | 1.89 | 343 | 2000 | 114 | 2 | 1.00 | 260 |
| 1972 | 163 | 4 | 1.23 | 363 | 2001 | 120 | 4 | 1.19 | 382 |
| 1973 | 116 | 6 | 1.59 | 321 | 2002 | 108 | 8 | 1.84 | 631 |
| 1974 | 134 | 4 | 1.31 | 446 | 2003 | 139 | 2 | 1.00 | 749 |
| 1975 | 136 | 6 | 1.62 | 409 | 2004 | 145 | 8 | 1.76 | 840 |
| 1976 | 91 | 4 | 1.32 | 280 | 2005 | 156 | 6 | 1.61 | 585 |
| 1977 | 110 | 8 | 2.07 | 367 | 2006 | 132 | 6 | 1.75 | 573 |
| 1978 | 100 | 4 | 1.38 | 358 | 2007 | 152 | 8 | 1.88 | 720 |
| 1979 | 133 | 2 | 1.00 | 272 | 2008 | 128 | 6 | 1.80 | 531 |
| 1980 | 121 | 4 | 1.46 | 560 | 2009 | 145 | 8 | 1.73 | 605 |
| 1981 | 153 | 6 | 1.58 | 459 | 2010 | 156 | 4 | 1.23 | 422 |
| 1982 | 110 | 6 | 1.53 | 317 | 2011 | 149 | 8 | 1.82 | 512 |
| 1983 | 96 | 6 | 1.99 | 312 | 2012 | 132 | 6 | 1.58 | 347 |
| 1984 | 137 | 8 | 1.45 | 388 | 2013 | 156 | 4 | 1.37 | 259 |
| 1985 | 118 | 2 | 1.00 | 294 | 2014 | 154 | 8 | 1.89 | 503 |
| 1986 | 120 | 6 | 2.09 | 293 | 2015 | 141 | 4 | 1.34 | 527 |
| 1987 | 133 | 6 | 1.69 | 275 | 2016 | 144 | 8 | 2.02 | 632 |
| 1988 | 148 | 6 | 1.63 | 289 | 2017 | 117 | 8 | 1.90 | 787 |
| 1989 | 134 | 10 | 2.18 | 411 | 2018 | 171 | 4 | 1.43 | 466 |
Межгодовая изменчивость и линия тренда ИЦВцб в центральном плесе (рис. 2) мало отличаются от таковых для ИЦВцб в приплотинном плесе [25], что свидетельствует о единых причинах изменчивости интенсивности цветения для большей части акватории Можайского водохранилища. Как и в приплотинном плесе, в центральном районе водохранилища в первые годы его существования диапазон изменчивости ИЦВ от года к году был в 3 раза меньше, чем в последние годы рассматриваемого многолетнего периода. Причем значения ИЦВ в наименее продуктивные годы увеличились незначительно, тогда как в наиболее продуктивные годы они возросли вдвое, что и определило хорошо выраженный положительный тренд ИЦВ в Можайском водохранилище.
Как видно из приведенных в табл. 1 данных, продолжительность летне-осеннего цветения сильно меняется: от 91 сут в 1976 г. до 184 сут в 1999 г. и в среднем составляет 132 сут. Тем не менее связь продолжительности сезона цветения и ИИПК статистически не значима (r = –0.12), что свидетельствует об асинхронности межгодовой изменчивости этих характеристик.
Весь ряд рассчитанных за 58 лет значений ИИПК и ИЦВ был разбит на два периода: 1961–1971 гг. – интенсивного переформирования ложа водохранилища в первые годы после его образования, 1972–2018 гг. – относительной стабилизации сформированного илистыми отложениями покрова дна глубинных участков.
В начальный период формирования экосистемы водохранилища особенно сильное цветение было отмечено в 1962 г. (табл. 1) – второй год затопления поймы, когда происходило активное выделение в придонный слой воды продуктов бактериального разложения органических веществ в затопленном почвенном покрове дна. Экспериментально было показано, что через год после затопления лугово-болотной почвы, отобранной со дна Можайского водохранилища, концентрация ортофосфатов в воде над ней возрастает до 7.18 мг/л [9]. В первые годы существования водохранилища происходит активная переработка абразионных участков берегов, при которой глинисто-песчаные продукты абразии доминируют при формировании донных отложений и образующийся детрит частично перекрывается минеральными веществами, что препятствует обогащению трофогенного поверхностного слоя воды биогенными продуктами разложения оседающего детрита при штормовом взмучивании [5]. В эти годы в придонных слоях очень медленно формировалась зона аноксии и не наблюдалось ее повсеместное существование. После исключения из первого периода аномального по сильному цветению 1962 г. корреляционная связь между ИИПК и ИЦВ в это десятилетие отсутствовала (рис. 3а).
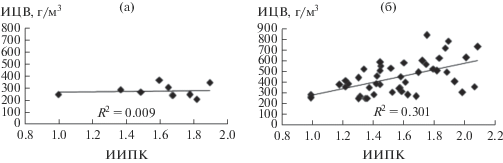
В последующий период (1972–2018 гг.) с уже сформированным покровом дна глубинных участков из илистых отложений выявленная ранее зависимость продуктивности от контрастности погоды [29] стала весьма вероятной благодаря полученной статистически высоко значимой связи ИЦВ с ИИПК (рис. 3б). Коэффициент корреляции этих индексов равен 0.55 (при n = 46 значимый rβ = 1% = 0.384 [22]). Из этого периода исключен 2003 г., ИЦВ которого не соответствует его ИИПК. Особенностью этого года было очень высокое значение ИЦВ при всего лишь двухфазной изменчивости температуры поверхностного слоя (фаза нагревания с 6 июня до 28 июля при максимальной среднесуточной температуре воды 27.20°C и последовавшая за ней фаза охлаждения до 22 октября). Двухфазными были также термические режимы в 1979, 1985 и 2000 гг., но интенсивность цветения в 2003 г. была в 2–3 раза выше, чем в эти годы (табл. 1). Причиной этого был аномально большой приток биогенных веществ, поступивших в водоем с водами многоводных июльских паводков, вследствие чего биомасса цианобактерий в июле–августе возросла до 4–6 мг/л. Температура речных вод превышала температуру трофогенного слоя, поэтому воды паводков поступали в основном в поверхностный слой, так как их плотность была наименьшая и препятствовала конвективному перемешиванию. К тому же в фазу нагревания и в августе скорость ветра ≤2 м/с при среднем значении 1.1 м/с, что исключало глубокое ветровое перемешивание поверхностных слоев воды в водоеме. В сентябре произошел всплеск цветения цианобактерий (до 12.7 мг/л в центральной части водохранилища), связанный с поступлением в водоем большого объема паводковых вод (расходы воды сентябрьского паводка в р. Москве превышали ее расходы во время весеннего половодья) и, соответственно, большого количества биогенных веществ, что продлило цветение водорослей до 22 октября. На фоне медленного осеннего охлаждения речных вод и трофогенного слоя в водохранилище конвективное перемешивание усиливалось в ветреную погоду (средняя скорость ветра в сентябре составляла 2.4 м/с при среднесуточной в некоторые сутки до 6 м/с). Это сопровождалось выносом большого количества биогенных веществ (дополнительно к их поступлению с речным притоком), накопленных в гиполимнионе за предшествующий длительный период безветренной погоды. Эти особенности погодных условий привели к тому, что 2003 г. оказался одним из наиболее продуктивных за время существования водохранилища, хотя значение его ИИПК было минимальным.
Влияние контрастности изменчивости погоды на продуктивность, по-видимому, характерна и для водоемов Европы, для которых лимнологом В. Оле установлено [31], как напоминает С.И. Кузнецов [11], что особенность структуры их биоактивности состоит в том, что вероятность цветения в них в 1.5 раза выше, чем в экваториальных озерах. Доля оседающего на дно органического вещества в структуре биоактивности озер умеренных широт – самая высокая среди озер мира, а скорость деструкции органического вещества в илах – в 1.5–2 раза превышает седиментацию легко минерализующегося бактериями детрита. Это указывает на заметный вклад внутренней биогенной нагрузки в цветение водоемов замедленного водообмена.
ВЫВОДЫ
Весеннее цветение водохранилища определяется диатомовыми водорослями, имеет непродолжительный характер (от 4 до 28 сут) и зависит, во-первых, от интенсивности наполнения водохранилища водой половодья и количества приносимых ею растворенных биогенных веществ; во-вторых – от длительности интенсивного конвективного перемешивания при штилевой погоде во время прогрева поверхностного слоя воды от 0 до 10–15°С – температуры, благоприятной для развития холодолюбивых диатомовых водорослей. Возникающий в период весеннего цветения большой биогидрохимический круговорот распространяется в водоеме повсеместно до дна пойменных участков, а нередко и до дна русловой ложбины, что интенсифицирует весеннее цветение дополнительным выносом в трофогенный слой воды, обогащенной биогенами.
Особенность слабопроточного водохранилища многолетнего регулирования стока с замедленным водообменом – большая продолжительность летне-осеннего цветения – до 180 сут. Оно вызвано цианобактериями и связано, как правило, с контрастностью погоды. Чем продолжительней фаза охлаждения, тем сильнее снижается температура поверхности воды в эту фазу; и чем больше таких фаз, тем интенсивнее вспышки цветения цианобактериями. Они возникают при смене погоды холодной фазы солнечной и штилевой погодой теплой фазы. В фазу холодной и пасмурной погоды с возникающими конвективно-ветровыми вихрями происходит смена малого биогидрохимического круговорота веществ в эпилимнионене большим круговоротом биогенных веществ вследствие разрушения стратификации и вовлечения в перемешивание вод гиполимниона, насыщенных регенерируемыми биогенными веществами в илах и в придонном анаэробном слое воды. Эту закономерность показывает статистически высоко значимая связь между интегральным индексом контрастности погоды и индексом цветения цианобактериями в почти полувековом ряду колеблющейся от года к году продуктивности слабопроточного стратифицированного в летне-осенний период водохранилища. Однако из этой закономерности бывают исключения, например двухфазный летне-осенний сезон 2003 г., отличавшийся аномально теплой и исключительно дождливой погодой с многоводными летне-осенними паводками с большим содержанием в воде биогенных веществ.
Контрастность изменчивости летне-осенней погоды особенно характерна для климата умеренных широт. Изменчивость погодных условий в значительной мере способствует формированию ярко выраженной внутрисезонной и годовой изменчивости продуктивности водохранилищ многолетнего регулирования стока, в том числе и водохранилищ системы водоснабжения г. Москвы.
Авторы благодарны М.Г. Ершовой (МГУ им. М.В. Ломоносова) за активное участие в лимнологических расчетах и анализе полученных результатов.
Список литературы
Виленкина М.Н., Сахарова М.И. Химизм воды, бактериальное население и фитопланктон Можайского водохранилища и изменения их в реке Москве (наблюдения с апреля 1960 г. по июль 1961 г.) // Учинское и Можайское водохранилища. М.: Изд-во МГУ, 1963. С. 389–406.
Гидроэкологический режим водохранилищ Подмосковья (наблюдения, диагноз, прогноз) / Под ред. К.К. Эдельштейна. М.: Перо, 2015. 286 с.
Гончаров А.В., Гречушникова М.Г., Пуклаков В.В. Новые возможности классического метода: автоматизированное определение первичной продукции и деструкции органического вещества в водоеме // Биология внутр. вод. 2018. № 4. С. 107–110.
Гречушникова М.Г. Изменчивость термического состояния Можайского водохранилища в вегетационный период. Автореф. дис. … канд. геогр. наук. М.: МГУ, 2002. М.: 23 с.
Даценко Ю.С. Эвтрофирование водохранилищ. Гидролого-гидрохимические аспекты. М.: ГЕОС, 2007. 252 с.
Даценко Ю.С., Пуклаков В.В., Эдельштейн К.К. Анализ влияния абиотических факторов на развитие фитопланктона в малопроточном стратифицированном водохранилище // Тр. КарНЦ РАН. Сер. Лимнология. 2017. № 10. С. 75–85.
Ершова М.Г. Моделирование генетической структуры Можайского водохранилища // Вод. ресурсы. 2003. Т. 30. № 2. С 142–153.
Ершова М.Г. Результаты расчета состава вод отдельных участков водохранилища с помощью балансовой модели // География и природ. ресурсы. 1984. № 2. С. 135–141.
Зайдельман Ф.Р., Гаджиев Я.М., Рожкова Л.С. Опыт моделирования процессов взаимодействия затопленных почв и грунтов с водами водохранилища // Комплексные исследования водохранилищ. М.: Изд-во МГУ, 1978. Вып. 4. С. 127–136.
Костоусова М.Н. Особенности очистки воды от фитопланктона // Вод. ресурсы. 1984. № 3. С. 97–104.
Кузнецов С.И. Микрофлора озер и ее геохимическая деятельность. Л.: Наука, 1970. 440 с.
Лебедев Ю.М., Широкова Е.Л. Некоторые факторы среды, определяющие продукцию Можайского водохранилища // Биол. науки. 1972. № 6. С. 116–118.
Левшина Н.А. Структурные особенности фитопланктона Можайского водохранилища // Комплексные исследования водохранилищ. М.: Изд-во МГУ, 1980. Вып. 5. С. 149–154.
Левшина Н.А. Фитопланктон Можайского водохранилища // Комплексные исследования водохранилищ. М.: Изд-во МГУ, 1973. Вып. 2. С. 50–55.
Левшина Н.А., Сахарова М.И. Планктон и бентос // Комплексные исследования водохранилищ. М.: Изд-во МГУ, 1979. Вып. 3. С. 262–274.
Моделирование режима фосфора в долинном водохранилище. М.: Изд-во МГУ, 1995. 80 с.
Орлеанская О.Б. Зимний фитопланктон Можайского водохранилища // Комплексные исследования водохранилищ. М.: Изд-во МГУ, 1978. Вып. 4. С. 189–191.
Пуклаков В.В. Гидрологическая модель водохранилища. М.: ГЕОС, 1999. 95 с.
Пуклаков В.В., Эдельштейн К.К. Расчеты плотностных течений в Можайском водохранилище // Метеорология и гидрология. 2001. № 5. С. 94–104.
Соколов Д.И. Снижение стока органических веществ р. Москвы в Можайском водохранилище // Современные проблемы водохранилищ и их водосборов. Т. II. Тр. Междунар. науч.-практ. конф. Пермь: ПГУ, 2013. С. 175–180.
Телитченко М.М., Цыцарин Г.В., Широкова Е.Л. Микроэлементы и “цветение” воды // Гидробиол. журн. 1970. № 6. С. 241–250.
Урбах В.Ю. Биометрические методы. М.: Наука, 1964. 415 с.
Чернега С.С., Эдельштейн К.К.Принципы гидроэкологической реконструкции долинных водохранилищ // Эколого-гидрологические проблемы изучения и использования водных ресурсов. Казань: АН РТ, 2006. С. 452–455.
Шушкевич Е.В., Карпушенко А.В., Полянин В.О., Столярова Е.А. Водоподготовка и управление качеством воды в условиях развития фитопланктона на примере москворецкого водоисточника // Водоснабжение и сан. техника. 2010. № 10. Ч. 1. С. 13–15.
Эдельштейн К.К., Пуклаков В.В., Даценко Ю.С. Экспериментально-теоретические основы диагноза и прогноза формирования цветения в водохранилищах – источниках муниципального водоснабжения // ВОДА MAGAZINE. 2017. № 4 (116). С. 34–40.
Эдельштейн К.К. Водохранилища России, экологические проблемы и пути их решения. М.: ГЕОС, 1998. 277 с.
Эдельштейн К.К. Гидрология озер и водохранилищ. М.: Перо, 2014. 398 с.
Эдельштейн К.К. Экологическое благоустройство москворецких водохранилищ // Природа. 1997. № 9. С. 92–97.
Эдельштейн К.К., Гречушникова М.Г., Гончаров А.В. Влияние изменчивости погоды на биопродуктивность водохранилища // Вестн. Московского ун-та. Сер. 5, География. 2020. № 2. С. 36–44.
Эдельштейн К.К., Пуклаков В.В., Даценко Ю.С., Ерина О.Н, Соколов Д.И. Интенсивность поступления кислорода из пресных водоемов // Вод. ресурсы. 2018. № 3. С. 309–318.
Ohle W. Beitrage zur Productionsbiologie der Gewasser // Archiv fur Hidrobiol. 1955. Suppl. V. 22 (3–4). P. 456–479.
Yershova M.G. Modelling of the Genetic Composition of Water Masses in Two Valley Reservoirs in the Volga Basin // Int. Revue gesamten Hidrobiol. V. 81. 1996. № 2. P. 293–313.
Дополнительные материалы отсутствуют.