

Зоологический журнал, 2019, T. 98, № 10, стр. 1168-1176
Новые данные о гималайском медведе (Ursus (euarctos) thibetanus G. Cuvier 1823, Carnivora, Ursidae) в плейстоцене Урала
a Институт экологии растений и животных УрО РАН
620008 Екатеринбург, Россия
b Уральский федеральный университет
620002 Екатеринбург, Россия
* E-mail: djulfa250@rambler.ru
Поступила в редакцию 04.03.2019
После доработки 25.03.2019
Принята к публикации 10.04.2019
Аннотация
Изложены новые данные об обитании гималайского медведя (Ursus thibetanus G. Cuvier) на Урале в плейстоцене. В отложениях пещеры Иманай (53°02′ с.ш., 56°26′ в.д.) обнаружены зубы I3, р4 и m1 мелкого медведя. Одонтологический и одонтометрический анализ показал, что найденные зубы принадлежат U. thibetanus. Остатки этого вида датируются микулинским (эем) межледниковьем, что подтверждает анализ сопутствующей фауны, археологического материала и радиоуглеродные даты из слоев, где найдены остатки гималайского медведя. Описаны морфологические признаки, сближающие медведя из Иманая с современными гималайскими медведями. В то же время отмечены признаки, характерные для плейстоценовых U. thibetanus.
Гималайский медведь (Ursus (Euarctos) thibetanus G. Cuvier 1823) населяет леса Южной и Юго-Восточной Азии, Китая, Кореи, Японии и Приморья (Garshelis, Steinmetz, 2016). Он питается преимущественно растительными кормами, тогда как беспозвоночные и мелкие позвоночные составляют небольшую долю в рационе (Барышников, 2007). В северных частях ареала и в горах медведи на зиму залегают в берлоги, которые устраивают в дуплах деревьев, а в горных районах – в пещере (Гептнер и др., 1967). Гималайский медведь облигатно связан с древесной растительностью, т.е. является типичным видом-дендрофилом, не столь обычным среди крупных млекопитающих. Данные о находки ископаемых остатков этого вида позволяет уверенно реконструировать лесную растительность для периода его обитания в регионе.
В плейстоцене U. thibetanus был распространен гораздо шире, чем в настоящее время. Гималайский медведь населял Кавказ и южную половину Европы и Средний Урал (Барышников, 2007). Ископаемые остатки гималайского медведя найдены на Среднем Урале (Baryshnikov, 2002) и на юге Восточной Сибири (Оводов, Филлипов, 2000; Ovodov et al., 2010). Самые ранние находки U. thibetanus в Европе относятся к раннему плиоцену (Baryshnikov, Zakharov, 2013), тогда как в Азии ископаемые остатки гималайского медведя встречаются начиная с раннего плейстоцена (Wagner et al., 2017a). Подавляющее большинство местонахождений с остатками этого вида датируются ранним–средним плейстоценом. Самые поздние находки в Европе и на Урале датируются последним (эем, микулинское, казанцевское) межледниковьем или изотопной кислородной стадией 5е (ИКС 5е), 130–120 тыс. л. н. (Барышников, 2007; Фадеева и др., 2019; Cregut-Bonnoure, 1996). Радиоуглеродная дата U. cf. thibetanus из Ботовской пещеры в Прибайкалье составляет более 41 000 лет, AA-83719 (Ovodov et al., 2010), а таковая зуба из пещеры Тетюхинская (Средний Сихотэ-Алинь) – 39 874 ± 133 лет (NSK-850, UGAMS-21786) (Косинцев и др., 2016). Не ясно, имел ли гималайский медведь такой широкий ареал на протяжении всего плейстоцена или проникал в западные районы Евразии эпизодически, вместе с распространяющейся лесной растительностью.
Цель данной работы – на основе методов одонтологического анализа описать и определить таксономическую принадлежность зубов I3, р4 и m1 мелкого медведя, найденных в отложениях пещеры Иманай (Южный Урал), и обсудить отличия плейстоценовых гималайских медведей от современных.
МАТЕРИАЛЫ И МЕТОДЫ
При раскопках пещеры Иманай на Южном Урале (53°02′ с.ш., 56°26′ в.д.), был выявлен комплекс костных остатков поздне-плейстоценовых и голоценовых млекопитающих (Гимранов и др., 2016; Gimranov, Kosintsev, 2018). Среди костных остатков найдены изолированные зубы – левый верхний третий резец (I3), левый нижний четвертый премоляр (р4) и левый нижний первый моляр (m1) принадлежащие медведю (рис. 1) и отличающиеся от остальных аналогичных остатков маленькими размерами. Зуб I3 обнаружен на глубине 15 см, р4 найден на глубине 40–50 см от поверхности, m1 на глубине 50–60 см. Коллекция костных остатков из пещеры Иманай хранится в музее Института экологии растений и животных УрО РАН, № 2284.
Рис. 1.
Зубы Ursus thibetanus из отложений пещеры Иманай: 1 – р4 sin вид сверху, 2 – р4 sin вид с буккальной стороны, 3 – I3 sin вид с лингвальной стороны, 4 – I3 sin вид с дистальной стороны, 5 – m1 sin вид сверху, 6 – m1 sin вид с лингвальной стороны.
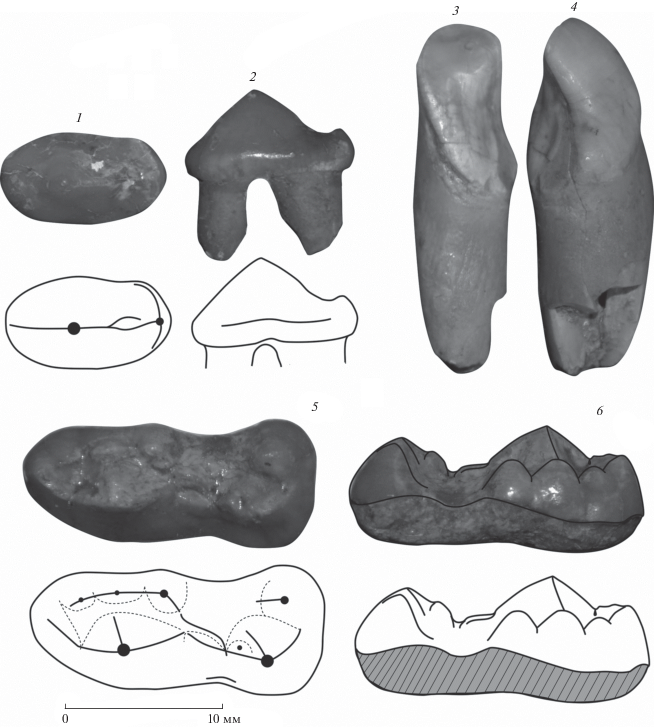
Для видовой идентификации было проведено сравнение размеров и строения коронок этих зубов с зубами современных бурого медведя (U. arctos L., 1758) с Урала (n = 25) и гималайского медведя из Приморского края (n = 35). Также было проведено сравнение с аналогичными зубами плейстоценовых бурого и гималайского медведей, медведя Денингера (U. deningeri von Reichenau 1904), кударского пещерного медведя (U. kudarensis Baryshnikov 1985), большого (U. (Spelearctos) spelaeus Rosenmuller 1799) и малого (U. (S.) savini Andrews 1922) пещерных медведей. В данной статье используется терминология для элементов коронки зуба, предложенная Рабедером (Rabeder, 1999).
РЕЗУЛЬТАТЫ
Зуб I3 значительно стерт, клыкоподобный (рис. 1), сжат с мезиального края, имеет выраженные дистальную кристу и лингвальное расширение. Длина I3 составляет 8.1 мм, ширина 7.0 мм.
Зуб р4 не стерт, коронка овальная (рис. 1), длиной 10.4 мм и шириной 5.9 мм. Соотношение длины и ширины коронки 57%. Передний корень немного больше заднего. Ребра протоконида (вид сбоку) образуют тупой угол. Протоконид не высокий, имеет переднюю и заднюю кристы, задняя криста у основания раздваивается. Буккальный цингулид отсутствует, наличие лингвального цингулида неустановленно, в связи с повреждением лингвального края основания коронки. В задней части коронки имеется бугорок, расположенный по центру у основания задней кристы.
Зуб m1 не стерт, коронка имеет срединное заужение (рис. 1). Длина зуба составляет 19.5 мм, ширина 8.6 мм. Соотношение длины и ширины коронки составляет 44%. Протоконид крупный, имеются антериальная, постериальная и внутренняя кристы. Между протоконидом и гипоконидом имеется выраженный гребень. Параконид в виде гребня, отделен от протоконида небольшой щелью. Метаконидный комплекс состоит из трех бугров, постепенно увеличивающихся в постериальном направлении. Самый крупный из них (последний) значительно уступает по размерам протокониду. Постериальная криста последнего бугра метаконидного комплекса развивается в направлении к гипокониду. Гипоконид крупный, имеет антериальную, постериальную и внутреннюю кристы. Также на антериальной кристе гипоконида имеется маленький бугорок. Энтоконид одиночный, расположен в лингва-дистальном углу коронки, вдвое меньше гипоконида, имеет только антериальную кристу. Преэнтоконидные структуры отсутствуют. Имеется плохо выраженный буккальный цингулид в зоне контакта тригонида и талонида. Другие цингулиды отсутствуют.
ОБСУЖДЕНИЕ
Зуб I3 достаточно стерт, поэтому можно провести только сравнение размеров (табл. 1). Значения длины и ширины резца из пещеры Иманай не попадают в пределы изменчивости I3 пещерных медведей, а также плейстоценового и современного бурого медведя (табл. 1). По размерам изучаемый зуб попадает в пределы изменчивости I3 современного U. thibetanus.
Таблица 1.
Размеры I3 ископаемых и современных медведей
| Промер | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| L | 13.5–19.5 n = 36 |
18.5–23.5 n = 3 |
12.4–16.7 n = 21 |
8.7–12.8 n = 26 |
8.8–13.1 n = 25 |
7.7–10.5 n = 26 |
8.2 |
| B | 10.5–15.6 n = 36 |
15.0–15.5 n = 2 |
12.6–18.2 n = 21 |
9.5–13.0 n = 21 |
7.8–11.5 n = 25 |
6.4–8.5 n = 26 |
6.9 |
Примечания. Выборки разных представителей Ursidae: 1 – U. deningeri, Herkova jama, Словения (Rabeder et al., 2010); 2 – U. savini, Краснодар, Северный Кавказ (Борисяк, 1932); 3 – U. spelaeus, Одесса, Украина (Барышников, 1987); 4 – плейстоценовый U. arctos, Аппенинский п-ов (Torres, 1988); 5 – современный U. arctos, Урал (наши данные); 6 – современный U. thibetanus, Приморье (наши данные); 7 –U. thibetanus, Иманай, Урал (n = 1).
Зуб p4 из Иманая по размерам коронки значительно меньше размеров этого зуба у пещерных медведей (U. deningeri, U. kudarensis, U. spelaeus). Зуб р4 из пещеры Иманай ненамного превосходит наименьшую длину р4 малого пещерного медведя (U. savini) из Кизеловской пещеры на Среднем Урале (табл. 2). Он меньше минимальных размеров зуба у позднеплейстоценового U. arctos и близок к минимальным размерам р4 современного U. arctos. Его размеры совпадают с размерами зубов ископаемых и современных гималайских медведей (табл. 3). U. thibetanus из Симай, Мауэр (один экземпляр), Перпиньян и Азых по размерам p4 находятся несколько обособленно от современной выборки. Зуб р4 из Мауэр (один экземпляр), Перпиньян и Азых крупнее р4 современных гималайских медведей. Другие ископаемые находки (Мауэр (один экземпляр), Гайтан, Чжоукоудянь 1), в том числе и р4 из пещеры Иманай находится в пределах области изменчивости современных гималайских медведей (рис. 2).
Таблица 2.
Размеры р4 и m1 ископаемых и современных медведей
| Зуб | Промер | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|---|
| р4 | L | 12.4–15.7 n = 21 |
12.5–16.7 n = 20 |
10.1–15.4 n = 17 |
11.9–19.7 n = 55 |
12.3–17.2 n = 10 |
10.1–15.4 n = 50 |
10.4 |
| B | 7.6–9.9 n = 21 |
6.9–10.2 n = 20 |
7.3–10.6 n = 10 |
8.6–12.8 n = 55 |
6.7–9.2 n = 10 |
5.6–8.1 n = 50 |
5.9 | |
| m1 | L | 24.5–31.1 n = 20 |
25.3–32.7 n = 55 |
24.2–27.2 n = 14 |
26.8–34.3 n = 51 |
23.6–31.1 n = 27 |
20.5–26.4 n = 50 |
19.5 |
| B | 11.7–14.5 n = 20 |
12.1–15.8 n = 76 |
11.6–14.1 n = 14 |
11.7–17.2 n = 51 |
11.4–14.7 n = 27 |
9.4–13.4 n = 50 |
8.6 |
Примечания. Выборки разных представителей Ursidae: 1 – U. deningeri, Mosbach, Германия; 2 – U. kudarensis, Кударо 1, Грузия; 3 – U. savini, Кизел, Средний Урал; 4 – U. spelaeus, Zoolithen, Германия; 5 – плейстоценовый U. arctos, Taubach, Германия; 6 – современный U. a. arctos, Европа; 7 – Ursus thibetanus, Иманай, Урал. Все размерные данные, за исключением данных медведя из Иманая, взяты из работы Г.Ф. Барышникова (2007).
Таблица 3.
Размеры р4 и m1 ископаемых и современных Ursus thibetanus
| Выборка | № | p4 | m1 | ||
|---|---|---|---|---|---|
| L | B | L | B | ||
| Ископаемые U. thibetanus |
1 | 11.3 | 6.3 | 20.7 | 9.0 |
| 1 | 11.5 | 7.7 | 20.5 | 10.4 | |
| 2 | 12.1 | 6.9 | 20.4 | 9.1 | |
| 3 | 8.9 | 5.9 | 21.6 | 10.5 | |
| 4 | 10.3 | 18.3 | 8.1 | ||
| 5 | 10.0 | 6.2 | 20.9 | 10.0 | |
| 6 | 10.4 | 6.9 | 20.3 | 10.8 | |
| 7 | 11.7 | 6.0 | 21.0 | 10.0 | |
| 8 | 10.4 | 5.9 | 19.5 | 8.6 | |
| Современные U. thibetanus |
9 | 10.0 (8.3–11.7) n = 46 |
5.7 (4.5–6.6) n = 46 |
20.2 (17.4–22.6) n = 47 |
8.9 (7.6–10.4) n = 47 |
| 10 | 10.4 (8.6–12.5) n = 35 |
5.8 (4.7–6.6) n = 35 |
20.3 (17.5–22.0) n = 21 |
8.8 (8.0–9.5) n = 21 |
|
| Ископаемые U. thibetanus |
11 | 19.4 | 9.3 | ||
| 12 | 19.6 | 9.1 | |||
| 13 | 18.8 | 8.7 | |||
| 14 | – | 10.8 | |||
| 15 | 19.7 | 9 | |||
| 15 | 17.3 | 7.5 | |||
| 15 | 17.2 | 7.8 | |||
| 15 | 20.3 | 9.2 | |||
Примечания. Выборки U. thibetanus: 1 – Германия, Mauer (Барышников, 2007); 2 – Франция, Perpignan (Барышников, 2007); 3 – Франция, Cimay (Барышников, 2007); 4 – Франция, Achenheim (Dehm, 1943); 5 – Албания, Gajtan (Fistani, Crégut-Bonnoure, 1993); 6 – Азербайджан, Азых (Барышников, 2007); 7 – Китай, Чжоукоудянь 1 (Pei, 1934); 8 – Россия, Иманай; 9 – современные материковые подвиды Ursus thibetanus (Барышников, 2007); 10 – современные Ursus thibetanus из Приморья (наши данные); 11 – Япония, Miyata (Барышников, 2007); 12 – Грузия, Кударо (Baryshnikov, 2010); 13 – Австрия, Laaerberg (Baryshnikov, 2010); 14 – Хорватия, Šandalja I (Wagner, 2017a); 15 – Россия, Иркутская обл., Ботовская пещера (Оводов, Филлипов, 2000).
Рис. 2.
Соотношение длины (L) и ширины (B) р4 ископаемых и современных U. thibetanus. 1 – Германия, Mauer (Барышников, 2007); 2 – Франция, Perpignan (Барышников, 2007); 3 – Франция, Cimay (Барышников, 2007); 4 – Албания, Gajtan (Fistani, Crégut-Bonnoure, 1993); 5 – Азербайджан, Азых (Барышников, 2007; Wagner et al., 2017); 6 – Китай, Чжоукоудянь 1 (Pei, 1934); 7 – Россия, Иманай. Точками обозначены особи U. thibetanus из современных популяций (наши данные).
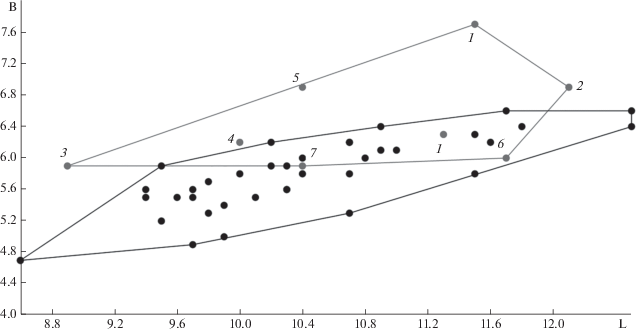
По строению коронки р4 из пещеры Иманай значительно отличается от p4 пещерных медведей, которым характерно наличие дополнительных бугорков, особенно многочисленных у малого пещерного медведя (Борисяк, 1932; Барышников, 2007). Значительно отличается морфология коронки p4 из описанного выше местонахождения и от плейстоценовых и современных бурых медведей (Гимранов, 2018), которые имеют метаконид и выраженную большую кристу (или гребень). У U. arctos протоконид высокий, его ребра образуют острый угол. У U. thibetanus протоконид низкий и его ребра образуют тупой угол, что видно на зубе из пещеры Иманай (рис. 1). Форма протоконида (вид сбоку) позволяет четко дифференцировать гималайского и бурого медведей. Строение коронки р4 из Иманая достаточно сходно с современным гималайским медведем. Единственным признак, который не встречен нами в изученной серии современных U. thibetanus, но который имеется у р4 из Иманая, является бугорок, расположенный по центру у основания задней кристы.
Зуб m1 из Иманая по размерам коронки значительно меньше размеров этого зуба у пещерных медведей (U. deningeri, U. kudarensis, U. spelaeus, U. savini) (табл. 2). Он меньше минимальных размеров зуба у позднеплейстоценовых бурых медведей и близок к минимальным размерам m1 современного бурого медведя. Его размеры совпадают с размерами зубов ископаемых и современных гималайских медведей (табл. 3). U. thibetanus из Ахенгейм, Симай, Мауэр (один экземпляр), Гайтан, Чжоукоудянь 1 и Азых по размерам m1 находятся несколько обособленно от современной выборки. Только U. thibetanus Ахенгейм имеет небольшие размеры m1. U. thibetanus из Симай, Мауэр (один экземпляр), Гайтан, Чжоукоудянь 1 и Азых превосходят по размерам современных гималайских медведей. Остальные ископаемые находки (Перпиньян, Мауэр (один экземпляр), Мията, Лааерберг и Кударо) в том числе и m1 из Иманая находится в пределах области изменчивости современных гималайских медведей (рис. 3). Отдельного внимания заслуживает плейстоценовая выборка из Ботовской пещеры, которая имеет самые маленькие значения размеров m1 и перекрывается с таковыми значениями современных U. thibetanus.
Рис. 3.
Соотношение длины (L) и ширины (B) m1 ископаемых и современных U. thibetanus. 1 – Франция, Perpignan (Барышников, 2007); 2 – Германия, Mauer (Барышников, 2007); 3 – Франция, Cimay (Барышников, 2007); 4 – Азербайджан, Азых (Барышников, 2007); 5 – Япония, Miyata (Барышников, 2007); 6 – Албания, Gajtan (Fistani, Crégut-Bonnoure, 1993); 7 – Франция, Achenheim (Dehm, 1943); 8 – Китай, Чжоукоудянь 1 (Pei, 1934); 9 – Грузия, Кударо (Baryshnikov, 2010); 10 – Россия, Иманай; 11 – Австрия, Laaerberg (Baryshnikov, 2010); 12 – Россия, Иркутская обл., Ботовская пещера (Оводов, Филлипов, 2000). Точками обозначены особи U. thibetanus из современных популяций.
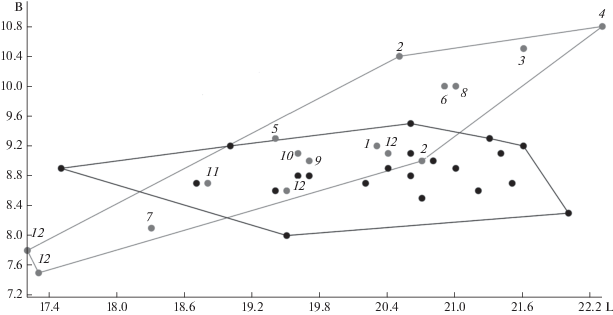
Строение коронки m1 из Иманая значительно отличается от m1 пещерных медведей, которым характерно наличие дополнительных бугорков гипоконидного и энтоконидного комплексов (Борисяк, 1932; Барышников, 2007; Wagner, Cermak, 2012). Значительно отличается морфология коронки m1 из Иманая и от плейстоценовых и современных бурых медведей (Барышников, 2007; Wagner, Cermak, 2012), которые имеют двойной энтоконид соединяющийся с метаконидным комплексом и развитый гипоконулид. Строение коронки m1 из Иманая достаточно сходно с современным гималайским медведем. У современных U. thibetanus нами отмечен небольшой (слабый) гипоконулид в 11 из 20 случаев. Пожалуй, это единственный признак, который отличает медведя из Иманая от современного гималайского медведя. Отметим, что у ископаемого гималайского медведя из Кударо и Азых также нет гипоконулида (Baryshnikov, 2010; Wagner et al., 2017). Мы полагаем, что наряду с дополнительным бугорком на р4, отсутствие гипоконулида на m1 не являются диагностическими признаками для гималайского медведя. Важными для дифференциации U. thibetanus от других медведей являются такие признаки на m1, как наличие заднего гребня метаконидного комплекса развивающегося к гипокониду и наличие одиночного энтоконида расположенного на лингвально-дистальном углу коронки. Эти два признака всегда присутствуют у современных гималайских медведей, а также у ископаемых (Baryshnikov, 2010; Wagner et al., 2017), в том числе и у m1 из Иманая.
Вместе с костными остатками гималайского медведя в пещере Иманай найдены кости крупных млекопитающих. Количественно доминируют кости малого пещерного медведя (U. savini), на втором месте идут остатки ископаемого льва (Panthera (Leo) ex gr. fossilis-spelaea) (Гимранов и др., 2016; Яковлев и др., 2016; Гимранов и др., 2018). Большая часть видов из состава фауны пещеры Иманай обитала на Южном Урале на протяжении позднего плейстоцена и голоцена (Kosintsev, Bachura, 2013). Эскулапов полоз (Zamenis cf. longissimus) и гималайский медведь входили в состав фауны микулинского межледниковья (Ратников, 2009; Фадеева и др., в печати). Красный волк (Cuon alpinus), пещерные медведи и архар (Ovis ammon) входили в состав фауны первой половины – середины позднего неоплейстоцена (ИКС 5 – ИКС 3) (Гимранов и др., 2015; Kosintsev, Bachura, 2013). Мамонт (Mammuthus primigenius), шерстистый носорог (Coelodonta antiquitatis) и бизон (Bison priscus) обитали на протяжении всего позднего неоплейстоцена (ИКС 5 – 2). Только в состав фауны голоцена входили садовая соня (Eliomys quercinus) (Kosintsev, Bachura, 2013). Видов, характерных для среднего и более ранних этапов плейстоцена, не найдено. В слое с костями найдены каменные орудия, аналогичные бифасиальным остроконечникам из третьего культурного слоя мустьерской стоянки Ильская на Северном Кавказе (Гимранов и др., 2017). Этот археологический материал из Ильской датируется началом первой половиной позднего плейстоцена (Щелинский, Кулаков, 2005). По кости малого пещерного медведя из Иманая получена 14С дата – 26 320 ± 1790 лет (ГИН-14244). Это соответствует окончанию ИКС 3 или концу брянского (Denekamp) интерстадиала (Blockley et al., 2012).
ЗАКЛЮЧЕНИЕ
Проведенные морфологические анализы показали, что размеры зубов I3, р4 и m1 из Иманая соответствуют размерам зубов гималайского медведя. В то же время, строение коронок изучаемых зубов из Иманая существенно отличается от строения коронок современных и ископаемых медведей и сходно с U. thibetanus.
Основываясь на анализе видового состава фауны позвоночных, археологических материалов и радиоуглеродной дате, мы полагаем, что накопление костных остатков в пещере Иманай проходило на протяжении всего позднего плейстоцена и голоцена. К более узкому промежутку времени отнести остатки гималайского медведя на данный момент затруднительно. Но учитывая, что до настоящего времени его остатки на Урале, Кавказе и Европе находили в местонахождениях не моложе микулинского (эем) межледниковья (Барышников, 2007; Cregut-Bonnoure, 1996), мы полагаем, что и в пещере Иманай они датируются этим временем. Это подтверждают палинологические данные. На Южном Урале на протяжении большей части позднего плейстоцена господствовали открытые (степные) или полуоткрытые (с лесами по долинам рек и в межгорных котловинах) ландшафты (Смирнов и др., 1990; Yakovlev et al., 2006; Danukalova et al., 2011). Только в микулинское межледниковье территория Южного Урала была покрыта хвойно-широколиственными лесами (Гричук, 2002). Эти леса занимали западный склон Среднего Урала, что согласуется с находкой гималайского медведя на севере этого региона (Барышников, 2007).
Список литературы
Барышников Г.Ф., 1987. Пещерный медведь в палеолите Крыма // Труды Зоологического Института АН СССР. Т. 168. С. 38–65.
Барышников Г.Ф., 2007. Медвежьи (Carnivora, Ursidae). Фауна России и сопредельных стран. Млекопитающие. Т. I. Вып. 5. СПб.: Наука. 541 с.
Борисяк А.А., 1932. Новая раса пещерного медведя из четвертичных отложений Северного Кавказа // Труды Палеозоологического института АН СССР. Т. 1. С. 137–201.
Гептнер В.Г., Наумов Н.П., Юргенсон П.Б., Слудский А.А., Чиркова А.Ф., Банников А.Г., 1967. Млекопитающие Советского Союза. Морские коровы и хищные. Т. 2. Ч. 1. М.: Высшая Школа. 1004 с.
Гимранов Д.О., 2018. Морфотипическая характеристика четвертых премоляров бурого (Ursus arctos) и белого (Ursus maritimus) медведей (Сarnivora, Ursidae) // Зоологический журнал. Т. 97. № 2. С. 205–223.
Гимранов Д.О., Косинцев П.А., Смирнов Н.Г., 2015. Находка красного волка (Carnivora, Canidae, Cuon alpinus Pallas, 1811) в позднем неоплейстоцене Урала // Доклады Академии Наук. Т. 464. № 5. С. 636–638.
Гимранов Д.О., Котов В.Г., Румянцев М.М., 2017. Результаты комплексных исследований многослойной мустьерской стоянки в пещере Иманай-1 на Южном Урале // V (XXI) Всероссийский археологический съезд. Отв. ред.: Деревянко А.П., Тишкин А.А. Барнаул: Алтайский государственный университет. С. 251–252.
Гимранов Д.О., Котов В.Г., Румянцев М.М., Силаев В.И., Яковлев А.Г., Яковлева Т.И. и др., 2018. Крупнейшее в Евразии захоронение ископаемых львов (Panthera (Leo) ex gr. fossilis-spelaea) // Доклады Академии Наук. Т. 482. №2. С. 234–237.
Гимранов Д.О., Котов В.Г., Румянцев М.М., Яковлев А.Г., Сотникова М.В., Нурмухаметов И.М. и др., 2016. Пещера Иманай – новое палеонтологическое и археологическое местонахождение на Южном Урале // Материалы LXII сессии Палеонтологического общества при РАН “Проблемы и перспективы палеонтологических исследований”. Ред. Аркадьев В.В. и др. СПб.: Изд-во ВСЕГЕИ. С. 231–233.
Гричук В.П., 2002. Растительность позднего плейстоцена // Динамика ландшафтных компонентов и внутренних морских бассейнов Северной Евразии за последние 130 000 лет. Атлас-монография. Отв. ред. Величко А.А. М.: ГЕОС. С. 64–88.
Косинцев П.А., Тиунов М.П., Гимранов Д.О. Панов В.С., 2016. Первая находка гималайского медведя (Carnivora, Ursidae, Ursus (Euarctos) thibetanus G. Cuvier, 1823) в позднем плейстоцене Северной Евразии // Доклады Академии Наук. Т. 471. № 3. С. 368–370.
Оводов Н.Д., Филиппов А.Г., 2000. Вымершие медведи Сибири // Палеогеография каменного века. Корреляция природных событий и археологических культур палеолита Северной Азии и сопредельных территорий. Отв. ред. Дроздов Н.И. Красноярск: РИО КГПУ. С. 106–109.
Ратников В.Ю., 2009. Ископаемые остатки современных видов земноводных и чешуйчатых пресмыкающихся как материал для изучения истории их ареалов // Труды научно-исследовательского института геологии Воронежского государственного университета. Вып. 59. С. 1–91.
Смирнов Н.Г., Большаков В.Н., Косинцев П.А., Панова Н.К., Коробейников Ю.И., Олшванг В.Н. и др., 1990. Историческая экология гор Южного Урала. Свердловск: Уральское отделение РАН. 245 с.
Фадеева Т.В., Кадебская О.И., Косинцев П.А., 2019. Первые данные о межледниковой фауне млекопитающих из интактных отложений пещеры Махневская ледяная (Средний Урал) // Доклады Академии Наук (в печати).
Щелинский В.Е., Кулаков С.А., 2005. Ильская мустьерская стоянка (раскопки 1920-х–1930-х годов). СПб.: Европейский дом. 96 с.
Яковлев А.Г., Яковлева Т.И., Гимранов Д.О., 2016. Мелкие позвоночные (земноводные, пресмыкающиеся и млекопитающие) из местонахождения Иманай 1 (Южный Урал) // Геология, полезные ископаемые и проблемы геоэкологии Башкортостана, Урала и сопредельных территорий. № 11. С. 81–83.
Baryshnikov G., 2002. The Pleistocene black bear (Ursus thibetanus) from the Urals (Russia) // Lynx. V. 32. P. 33–43.
Baryshnikov G.F., 2010. Middle Pleistocene Ursus thibetanus (Mammalia, Carnivora) from Kudaro caves in the Caucasus // Proceedings of the Zoological Institute of RAS. V. 314. P. 67–79.
Baryshnikov G.F., Zakharov D.S., 2013. Early Pliocene Ursus thibetanus (Mammalia, Carnivora) from Priozernoe locality in the Dniester basin (Moldova Republic) // Proceedings of the Zoological Institute of RAS. V. 317. № 1. P. 3–10.
Blockley S.P.E., Lane C.S., Hardiman M., Rasmussen S.O., Seierstad I.K., Steffensen J.P. et al., 2012. Synchronisation of palaeoenvironmental records over the last 60,000 years, and an extended INTIMATE event stratigraphy to 48,000 b2k // Quaternary Science Reviews. V. 36. P. 2–10.
Fistani A., Crégut-Bonnoure E., 1993. Découverte d’Ursus thibetanus (Mammalia, Carnivora, Ursidae) dans le site pléistocéne moyen de Gajtan (Shkoder, Albanie) // Geobios. V. 26. P. 241–263.
Crégut-Bonnoure E., 1996. A review of small Middle Pleistocene bears from France // Acta Zoologica Cracoviensis. V. 39. № 1. P. 89–101.
Danukalova G., Yakovlev A., Osipova E., Alimbekova L., Yakovleva T., Kosintcev P., 2011. Biostratigraphy of the Late Upper Pleisticene (Upper Neopleisticene) to Holocene Deposits of the Belaya River Valley (Southern Urals, Russia) // Quaternary International. V. 231. № 1–2. P. 28–43.
Dehm R., 1943. Ein besonders kleiner Bär (Ursus schertzi n. sp.) aus dem Löß von Achenheim bei Straßburg im Elsaß // Neues Jahrbuch für Mineralogie, Geologie und Paläontologie. B. P. 137–153.
Garshelis D., Steinmetz R., 2016. Ursus thibetanus (errata version published in 2017). The IUCN Red List of Threatened Species. 2016:e.T22824A114252336. [Электронный ресурс]. Режим доступа: Дата обновления: 3 марта 2019 г.https://doi.org/10.2305/IUCN.UK.2016-3.RLTS. T22824A45034242.en
Gimranov D.O., Kosintsev P.A., 2018. Quaternary large mammals from the Imanay Cave // Quaternary Stratigraphy in Karst and Cave Sediments / INQUA Section on European Quaternary Stratigraphy. Ed. H. Z. Hajna, A. Mihevc, M. Năpăruș-Aljančič. Postojna: ZRC Publishing. P. 27–29.
Kosintsev P.A., Bachura O.P., 2013. Late Pleistocene and Holocene mammal fauna of the Southern Urals // Quaternary International. V. 284. P. 161–170.
Ovodov N.D., Kuzmin Y.V., Cruz R.J., 2010. AMS 14C dating of “exotic” mammals from south Siberian caves // Current Research in the Pleistocene. V. 27. P. 181–183.
Pei W., 1934. On the Carnivora from Locality 1 of Choukutien // Palaeontologia Sinica. V. 8. P. 1–217.
Rabeder G., 1999. Die Evolution des Höhlenbärengebisses // Mitteilungen der Kommission für Quartärforschung der Osterreichischen Akademie der Wissenschaften. V. 11. P. 1–102.
Rabeder G., Pacher M., Withalm G., 2010. Early Pleistocene Bear Remains from Deutsch-Altenburg (Lower Austria) // Mitteilungen der Kommission für Quartärforschung der Osterreichischen Akademie der Wissenschaften. Bd. 17. P. 1–135.
Torres T., 1988. Osos (Mammalia, Carnivora, Ursidae) del Pleistoceno Ibérico (Ursus deningeri Von Reichenau, Ursus spelaeus Rosenmüller-Heinroth, Ursus arctos Linneo): V. Dentivió decidual, fórmula dentaria y dentición superior // Boletín Geológico y Minero. V. 99. P. 660–714.
Wagner J., Cermak S., 2012. Revision of the early Middle Pleistocene bears (Ursidae, Mammalia) of Central Europe, with special respect to possible co-occurrence of spelaeoid and arctoid lineages // Bulletin of Geosciences. V. 87. № 3. P. 461–496.
Wagner J., Jiangzuo Q., Lenardić J.M., Liu J., 2017. Taxonomic revision of bears from the locality Šandalja i (Croatia) and its biostratigraphic consequences // Fossil Imprint. V. 73. № 3–4. P. 533–544.
Wagner J., Jiangzuo Q., Jinyi L., 2017a. Revision of early Asiatic black bear (Ursus thibetanus) – its morphological, phylogemetical and taxonomical results // Aragonit. V. 22. № 1. P. 26–27.
Yakovlev A., Danukalova G., Kosintcev P., Alimbekova L., Morozova E., 2006. Biostratigraphy of the Late Palaeolithic site of “Bajslan-Tash cave”: (the Southern Urals) // Quaternary International. V. 149. № 1. P. 115–121.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал