

Зоологический журнал, 2019, T. 98, № 3, стр. 314-318
Техногенный шум влияет на минимальную частоту компонентов песни зяблика (Fringilla coelebs) в московской популяции
И. В. Кисляков 1, *, В. В. Иваницкий 1
1 Биологический факультет Московского государственного университета
им. М.В. Ломоносова
119991 Москва, Россия
* E-mail: ilyakislyakov@yandex.ru
Поступила в редакцию 03.04.2018
После доработки 11.05.2018
Принята к публикации 23.05.2018
Аннотация
Для проверки гипотезы о сдвиге параметров песни птиц в более высокие частоты при наличии постоянного низкочастотного шума мы записали пение 78 самцов зяблика в городских парках Москвы. В каждой точке записи регистрировали уровень фонового шума. По сонограммам определяли минимальную частоту четырех фраз из состава трех широко распространенных в Москве типов песен. Измерения всех фраз произведены у 18–21 самцов, при этом у каждого измеряли по четыре песни данного типа, в случайном порядке выбранные с фонограммы. Минимальная частота фразы 1 в исполнении разных самцов варьировала от 4.94 до 5.22 кГц, фразы 2 – от 2.23 до 2.6 кГц, фразы 3 – от 1.77 до 2.64 кГц и фразы 4 – от 2.08 до 2.42 кГц. Достоверная положительная корреляция (коэффициент корреляции Пирсона, 0.67 < r < 0.79) минимальной частоты с уровнем шума выявлена для трех из четырех изучаемых компонентов песни. Резкое повышение минимальной частоты пения при усилении зашумленности выявлено при уровне шума 55 дБ и выше.
Природные шумы и все возрастающее техногенное зашумление среды обитания оказывает многообразное воздействие на акустическую коммуникацию животных. К настоящему времени выявлено несколько путей приспособления животных к воздействию шума.
Один из них – увеличение помехоустойчивости сигнала за счет изменения его временны́х параметров (гипотеза избыточности в канале связи). Так, огненные тиранновые мухоловки (Pyrocephalus rubinus), живущие среди городского шума, исполняют почти исключительно длинные песни, тогда как за пределами города – и длинные, и короткие (Rios-Chelen et al., 2013). Крапивники (Troglodytes pacificus) при наличии постоянного шума океанского прибоя исполняют более длинные песни и используют для их построения более продолжительные ноты, чем при его отсутствии (Gough et al., 2014). Зяблики, поющие при воздействии шума текущей воды возле горного ручья, исполняют более длительные серии одинаковых песен, нежели зяблики, поющие поодаль от источника шума (Brumm, Slater, 2006), но при сравнении зябликов, поющих в парках крупного города (Познань, Польша) и в пригородных лесах, различий в длине серий не найдено (Deoniziak, Osiejuk, 2016). Вопреки ожиданиям в песне мексиканской чечевицы (Carpodacus mexicanus) обнаружено сокращение количества элементов при наличии шума (Fernandez-Juricic et al., 2005; Wood, Yezerinac, 2006). Укорочение песни может указывать на компромисс с энергетическими затратами при увеличении амплитуды сигнала.
Однако, наиболее распространенной биоакустической адаптацией к воздействию шума является сдвиг частотного диапазона вокализации (Brumm, Slater, 2006; Slabbekoorn, Peet, 2003; Иваницкий и др., 2014). При этом птица повышает частоту своего пения, чтобы минимизировать перекрывание с низкочастотным городским (или иным) шумом. Такая корректировка связана с неизбежным уменьшением частотного диапазона вокализации, т.е. увеличивается минимальная частота без изменения максимальной частоты (Fernandez-Juricic et al., 2005; Wood, Yezerinac, 2006). К настоящему времени получено множество свидетельств частотного сдвига в песне птиц в ответ на воздействие шума, однако механизмы этого явления изучены недостаточно. Большой интерес представляет, например, связь частотных параметров песни и громкости ее исполнения. Так, увеличение последней в ответ на воздействие шума (эффект Ломбарда) было продемонстрировано экспериментально у южного соловья (Luscinia megarhynchos) (Brumm, 2004). В этой связи высказывают мнение, что непосредственная реакция птиц на шум состоит именно в увеличении громкости пения, которое уже вторично влечет за собой повышение минимальной частоты (Zollinger et al., 2012).
Представляет интерес также вопрос о том, какие интервалы времени занимает звуковая адаптация. Имеются подтверждения в пользу как гипотезы долговременной адаптации, сопровождающейся сменой диалектов (Luther, Baptista, 2010), так и гипотезы кратковременной поведенческой адаптации (Bermudez‑Cuamatzin et al., 2009).
Ранее было показано, что значение минимальной частоты в пении восточного соловья (Luscinia luscinia) в г. Москве на 260 Гц выше, чем в области, что свидетельствует в пользу гипотезы долговременной адаптации (Иваницкий и др., 2014). Зяблик, занимающий широчайший спектр местообитаний, также принадлежит к числу птиц, регулярно гнездящихся даже в самых крупных городах. В этом сообщении мы анализируем минимальную частоту отдельных компонентов песни московской популяции зяблика с привязкой к конкретным уровням шума, основным источником которого служит движение автотранспорта. Тем самым мы продолжаем изучение дифференциации песни в условиях городского местообитания (Кисляков, Иваницкий, 2017).
МАТЕРИАЛЫ И МЕТОДЫ
В анализ включены фонограммы пения 78 зябликов, записанные в городских парках Москвы (Битцевский, Измайловский, Кусковский лесопарки, Лосиный остров, Сокольники) в апреле–июне 2017 г. Записи производили в приграничной зоне каждого парка вдоль следования автомобильной дороги. Измерение уровня фонового шума производили на расстоянии не более 10 м от каждого поющего самца, при этом выбирали самцов, поющих на разном удалении от проезжей части. При помощи цифрового шумомера CEMDT-80 (диапазон измерения от 30 до 130 дБ для частот от 31.5 Гц до 8 кГц, точность ±1.5 дБ) регистрировали средний уровень шума на протяжении 3 мин пения каждого самца. Таким образом, каждый зарегистрированный уровень городского шума был сопоставлен с фонограммой определенного самца. Для записи использовали цифровые рекордер Marantz PMD 620 и конденсаторный микрофон-пушку Аudio-Technica ATR6550. Для визуализации и построения сонограмм использовали приложение Syrinx 2.5 (J.M. Burt: http://syrinxpc.com; окно Блэкмана, длина преобразования Фурье 512 точек с разрешением по оси частот 20 Гц и по оси времени 1.4 мс). На основе сонограмм составлены каталоги типов песен и слогов (Кисляков, Иваницкий, 2017).
Известно, что рекламное пение самцов зяблика является раздельным и имеет типологическую организацию, т.е. состоит из дискретных и стереотипных вокальных конструкций – единичных песен, каждая из которых относится к определенному типу (Симкин, Штейнбах, 1988). Для анализа мы выбрали 3 типа песен, широко распространенных на всей территории Москвы и Подмосковья (рис. 1). Измеряли минимальную частоту четырех фраз из состава этих песен. Как и большинство компонентов песни зяблика, фразы представляют собой трели, т.е. серии одинаковых нот или слогов. Поскольку минимальная частота фраз фактически совпадает с минимальной частотой образующих их слогов, мы будем использовать в целях удобства общий термин компонент для составляющих частей песни. Из рис. 1 видно, что одинаковые компоненты могут встречаться в разных типах песен и занимать в них различное положение, и что при этом существенно варьируют частотные параметры однотипных компонентов. Поэтому мы измеряли минимальную частоту каждого данного компонента лишь в пределах одного типа песни. Так, компоненты 1 и 2 измеряли в песне № 1, компоненты 3 и 4 – в песнях № 5 и № 13, соответственно. Нумерация типов песен приводится по их каталогу (Кисляков, Иваницкий, 2017). Высокочастотный компонент 1 был выбран для сравнения с относительно низкочастотными компонентами 2, 3 и 4. Измерения всех компонентов произведены у 18–21 самцов, при этом у каждого самца измеряли по четыре песни данного типа, в случайном порядке выбранные с фонограммы. Степень зависимости минимальных частот компонентов песен от уровней шума оценивали с помощью коэффициента корреляции Пирсона.
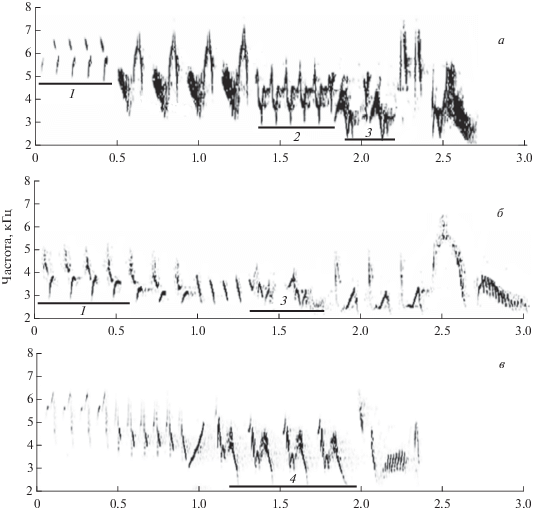
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Средняя частота городского шума в Москве составляет 0.85 кГц, а 75% его мощности находится в диапазоне от 0.46 до 1.55 кГц (Иваницкий и др., 2014). По визуальным оценкам на сонограммах верхняя граница шума составляет 2.0–2.5 кГц, так что низкочастотные компоненты (в т.ч. анализируемые нами фразы) вокализаций зяблика им перекрываются.
В обследованных нами точках уровень шума варьировал от 32 до 73 дБ. При этом минимальная частота компонента 1 в исполнении разных самцов варьировала от 4.94 до 5.22 кГц, компонента 2 – от 2.23 до 2.6 кГц, компонента 3 – от 1.77 до 2.64 кГц и компонента 4 – от 2.08 до 2.42 кГц. Рис. 2 иллюстрирует зависимость средних минимальных частот этих фраз от уровня фонового шума. Каждая точка соответствует одному самцу. Для компонента 1 коэффициент корреляции Пирсона r равен 0.73 (N = 18, p = 0.0004). Для компонента 2 r = 0.79 (N = 21, p = 0.00002). Для компонента 3 значимой корреляции не найдено: r = 0.39 при N = 21 и p = 0.083. Для компонента 4 коэффициент корреляции Пирсона равен 0.67 (N = 18, p = = 0.0025).
Рис. 2.
Зависимость между уровнем громкости фонового шума и минимальной частотой различных компонентов трех типов песен зяблика. Каждая точка соответствует средней минимальной частоте пения одного самца. По вертикали – частота, кГц; по горизонтали – уровень шума, дБ.
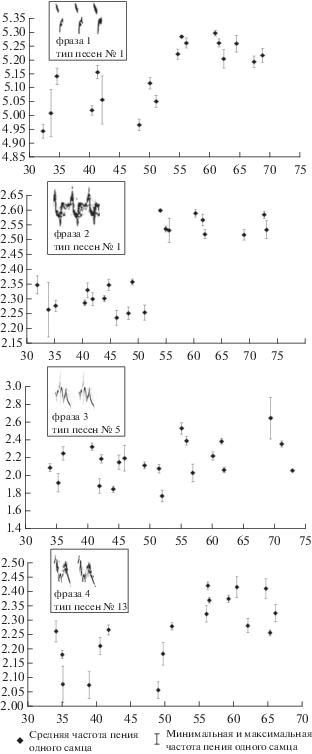
Таким образом, значимая и достаточно высокая (0.67 < r < 0.79) корреляция минимальной частоты с уровнем шума выявлена для трех из четырех изучаемых компонентов песни. При этом выраженный ответ на интервенцию шума проявляют не только низкочастотные компоненты, как и следовало ожидать, но и относительно высокочастотный компонент 1 в типе песни № 1. Похожие результаты получены ранее у восточного соловья (Иваницкий, Антипов, 2014).
Обращает на себя внимание то обстоятельство, что повышение минимальной частоты по мере усиления шума во всех трех случаях происходит не постепенно, а скачкообразно: начиная со среднего уровня шума 55 дБ. Аналогичный результат был получен также при изучении изменений минимальной частоты песни зяблика под воздействием естественного шума (звук ручья). В этом исследовании также зарегистрировано повышение уровня минимальной частоты песен зяблика, начиная с интенсивности шума примерно 50–55 дБ. Средняя минимальная частота варьировала от 1.44 до 2.20 кГц, т.е. размах изменчивости в целом выше, чем в наших результатах (r = 0.549, N = 14 и р = 0.042) (Brumm, Slater, 2006).
Возникает вопрос о механизме обусловленности изменения частотных параметров: является ли такое изменение быстрой поведенческой реакцией или она формируется в ряду поколений, как устойчивая характеристика диалекта? Например, в ответ на экспериментальное воздействие шума гаички (Poecile atricapillus и P. gambeli) исполняют преимущественно самые высокочастотные типы песен из своего репертуара, однако после прекращения шума такая избирательность пропадает (LaZerte et al., 2015). Столь же краткосрочной оказалась реакция повышения минимальной частоты в ответ на воздействие шума у пеночки-теньковки (Phylloscopus collybita) (Verzijden et al., 2010). В противоположность этому большие синицы (Parus major) в городских популяциях используют песни более высокочастотные по сравнению с песнями пригородных популяций вне зависимости от краткосрочного воздействия экспериментального шума. Поэтому можно сделать вывод о том, что в городе относительная высокочастотность песни закреплена генетически (Slabbecorn, Peet, 2003).
В нашем случае более вероятен сценарий быстрой реакции. Как было выявлено нами ранее, структура местообитаний зябликов даже в таком крупном городе, каким является Москва, не создает предпосылок для пространственного обособления отдельных поселений, которое могло бы привести к появлению локальных различий в репертуарах (Кисляков, Иваницкий, 2017). В пользу этого говорит также жесткая зависимость, показанная на рис. 2: различия в частотах зависят именно от уровня шума на расстояниях, которые составляют несколько сотен метров и которые не сравнимы с размерами даже локальных поселений. Таким образом, вокализации с разными минимальными частотами не образуют компактных участков распространения на территории г. Москвы.
Список литературы
Иваницкий В.В., Антипов В.А., Марова И.М., 2014. Восточный соловей (Luscinia luscinia) в Москве и Подмосковье: городской шум влияет на частотные параметры песни // Зоологический журнал. № 11. Т. 93. С. 1335–1339.
Кисляков И.В., Иваницкий В.В., 2017. Изменчивость и пространственная дифференциация песни зяблика (Fringilla coelebs) в городе Москве // Зоологический журнал. № 8. Т. 96. С. 960–972.
Симкин Г.Н., Штейнбах М.В., 1988. Песня зяблика и вокальные микрогруппировки у птиц // Орнитология. Вып. 23. С. 175–182.
Bermudez-Cuamatzin E., Ariel Rios-Chelen A., Gil D., Macias Garcia C., 2009. Strategies of song adaptation to urban noise in the house finch: syllable pitch plasticity or differential syllable use? // Behaviour. V. 146. P. 1269–1286.
Brumm H., 2004. The impact of environmental noise on song amplitude in a territorial bird // Journal of Animal Ecology. V. 73. P. 434–440.
Brumm H., Slater P.J., 2006. Ambient noise, motor fatigue, and serial redundancy in chaffinch song // Behavioral Ecology and Sociobiology. V. 60. P. 475–481.
Deoniziak K., Osiejuk T.S., 2016. Disentangling Relations among Repertoire Size, Song Rate, Signal Redundancy and Ambient Noise Level in European Songbird // Ethology. V. 122. № 9. P. 734–744.
Fernandez-Juricic E., Poston R., De Collibus K., Morgan T., Bastain B. et al., 2005. Microhabitat selection and singing behavior patterns of male House Finches (Carpodacus mexicanus) in urban parks in a heavily urbanized landscape in the western U.S. // Urban Habitats. V. 3. P. 49–69.
Gough D.C., Mennill D.J., Nol E., 2014. Singing seaside: Pacific Wrens (Troglodytes pacificus) change their songs in the presence of natural and anthropogenic noise // The Wilson Journal of Ornithology. V. 126(2). P. 269–278.
LaZerte S.E., Slabbekoorn H., Otter K.A., 2015. Sounds of the city: The effects of noise on communication in mountain and black-capped chickadees // NRESi research extension note. № 10.
Luther D.A., Baptista L., 2010. Does urban noise influencethe cultural evolution of bird songs? // Proceedings of Royal Society of London. Biological Science. V. 277. P. 469–473.
Rios-Chelen A.A., Quiros-Guerrero E., Gil D., Garcia C.M., 2013. Dealing with urban noise: vermilion flycatchers sing longer songs in noisier territories // Behavioral ecology and sociobiology. V. 67. P. 145–152.
Slabbekoorn H., Peet M., 2003. Ecology: birds sing at a higher pitch in urban noise – great tits hit the high notes to ensure that their mating calls are heard above the city’s din // Nature. V. 424. P. 267.
Verzijden M.N., Ripmeester E.A.P., Ohms V.R., Snelderwaard P., Slabbekoorn H., 2010. Immediate spectral flexibility in singing chiffchaffs during experimental exposure to highway noise // The Journal of Experimental Biology. V. 213. P. 2575–2581.
Wood W.E., Yezerinac S.M., 2006. Song Sparrow (Melospiza melodia) song varies with urban noise // Auk. V. 123. P. 650–659.
Zollinger S.A., Podos J., Nemeth E., Goller F., Brumm H., 2012. On the relationship between, and measurement of, amplitude and frequency in birdsong // Animal Behaviour. V. 84(4). P. 1–9.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал