

Зоологический журнал, 2019, T. 98, № 4, стр. 453-466
Видовой состав и распространение сурков (Marmotinae, Rodentia) на юго-востоке Западной Сибири в голоцене
М. М. Девяшин 1, *, П. А. Косинцев 1, **, В. В. Гасилин 1, ***
1 Институт экологии растений и животных УрО РАН,
620144 Екатеринбург, Россия
* E-mail: devjashinm@yandex.ru
** E-mail: kpa@ipae.uran.ru
*** E-mail: gasilinv@yandex.ru
Поступила в редакцию 21.12.2017
После доработки 8.05.2018
Принята к публикации 2.06.2018
Аннотация
Предложена методика видовой диагностики степного (Marmota bobak), серого (M. baibacina) и лесостепного (M. kastschenkoi) сурков по морфометрическим признакам черепа и нижней челюсти с использованием дискриминантного анализа и по строению слезной кости. Этими методами проведена видовая диагностика краниологических остатков сурков, датируемых голоценом. Обобщены литературные данные о местонахождениях голоценовых остатков сурков и распространении их в историческом прошлом на исследуемой территории. Проведена реконструкция ареала рода Marmota на территории юго-востока Западной Сибири в голоцене. Установлено, что северная граница ареала рода Marmota на протяжении голоцена проходила по южной границе лесной зоны, заходя на ее территорию в бассейне р. Обь, пересекала р. Енисей на широте около 56°00′ с.ш. и заходила на его восточный берег. В конце позднего голоцена северная граница в районе Енисея сместилась к югу и западу. Ареалы серого и лесостепного сурков сократились. Ареал первого сократился в юго-западном направлении, а ареал второго – отступил к западу. Ареал степного сурка в голоцене точно не известен. В последние 100–150 лет произошли заметное сокращение и фрагментация ареалов всех трех видов, вызванные распространением земледелия и прямым истреблением.
На юго-востоке Западной Сибири и в прилегающих районах Алтая и Казахстана в настоящее время перекрываются или смыкаются ареалы трех видов сурков: степного, или байбака (Marmota bobak Müller 1776); серого, или алтайского (M. baibacina Kastschenko 1899); лесостепного (M. kastschenkoi Stroganov et Yudin 1956). Восточная часть ареала степного сурка заходит в равнинные степи Центрального Казахстана до Иртышского левобережья, где перекрывается с западной частью ареала серого сурка. Самые северные колонии серого сурка зафиксированы на Северном Алтае в окрестностях с. Черга (51°34′ с.ш.). Далее на восток этот вид распространен до верховьев р. Абакан и юго-восточных районов Республики Тыва (Бибиков, Берендяев, 1978; Огнев, 1947). Ареал лесостепного сурка вытянут вдоль правобережья р. Оби от устья р. Томь (56° с.ш.) на севере до южной границы лесостепной зоны и захватывает участок слияния рек Бия и Катунь (52° с.ш.). В настоящее время существует разрыв между его ареалом и ареалом серого сурка (Тараненко, 2011). В смежном регионе, на юге Тувы и в Монгольском Алтае обитает монгольский сурок (Marmota sibirica Radde 1862) (Громов, Ербаева, 1995).
Самые древние находки представителей рода Marmota на юго-востоке Западной Сибири происходят из местонахождения Кизиха (Рубцовский район Алтайского края) и датируются средним эоплейстоценом (Зажигин, 1980). Костные остатки представителей рода из этого и большинства других местонахождений позднего плейстоцена и голоцена не определены до вида (Черников, 1960; Кирюшин и др., 1988; Сидоров, 1989; Абдулганеев, 1996; Лаухин и др., 1999; Мандырка и др., 1999; Деревянко и др., 2002; Кирюшин, 2002; Косинцев, 2002; Мерц, 2002; Оводов и др., 2003; Agadjanian, Serdyuk, 2005; Косинцев и др., 2011; Васильев, 2013). Часть остатков отнесены к серому сурку (Кожамкулова, 1969; Оводов, 1980, 1983, 2009; Васильев, Гребнев, 1994; Оводов и др., 1998, 2001; Оводов, Мартынович, 1999, 2000; Васильев, Зенин, 2006; Васильев и др., 2006, 2016; Агаджанян, 2007; Васильев, Шуньков, 2009; Молодин, Ефремова, 2010). Остатки из одного позднеплейстоценового (Мотузко, Акимова, 2013) и одного раннеголоценового (Гальченко, 1995) местонахождений отнесены к монгольскому сурку. Во всех случаях видовое определение остатков проведено без обоснования.
При изучении ископаемых остатков рода Marmota с территории юго-востока Западной Сибири возникает проблема их видовой диагностики. Связана она с тем, что морфологически степной, серый и лесостепной сурки очень сходны, так как филогенетически это близкие виды. Некоторые авторы рассматривали их в составе одного вида, выделяя в качестве подвидов степного и серого (Бибиков, 1989). Другие авторы относили степного и серого сурков к самостоятельным видам, но лесостепного сурка рассматривали либо в качестве подвида серого (Лаптев, Юдин, 1952; Громов, Ербаева, 1995), либо даже не выделяли его в отдельный подвид (Огнев, 1947). Последние исследования показывают, что генетически степной, серый и лесостепной сурки являются самостоятельными видами и входят в одну группу близких видов “bobak” (Брандлер, 2003; Брандлер и др., 2010).
Современные представители рода Marmota морфологически различаются по окраске и густоте шерстного покрова, длине ости волоса, по строению черепа, форме коронок зубов (Громов и др., 1965; Галкина, 1970; Галкина, Тараненко, 2002; Галкина и др., 2005). В силу специфики ископаемого материала далеко не все эти признаки пригодны для их видовой диагностики. Для настоящего исследования оптимальной является ранее разработанная методика диагностики степного и серого сурков по комплексу морфотипических и морфометрических признаков на черепе и нижней челюсти (Гасилин, Косинцев, 2011). Для диагностики ископаемых материалов с рассматриваемой территории методика была дополнена данными по третьему виду – лесостепному сурку. Поскольку ареал монгольского сурка не входит в район исследования (рис. 1), вид не включен в анализ.
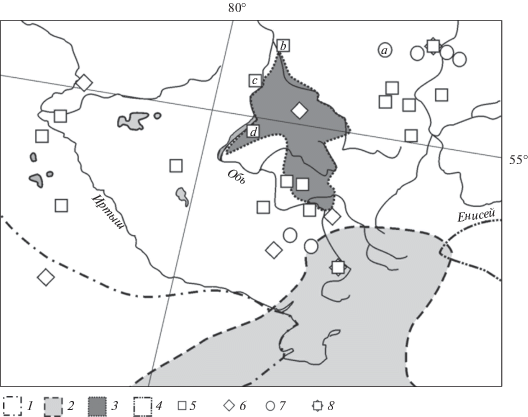
Рис. 1.
Современные границы ареалов представителей рода Marmota и местонахождения их остатков на юго-востоке Западной Сибири: 1 – северо-восточная граница ареала Marmota bobak; 2 – северная граница ареала M. baibacina; 3 – ареал M. kastschenkoi; 4 – западная граница ареала M. sibirica; 5 – находки сурков, датируемые поздним суббореальным периодом голоцена; 6 – находки сурков, датируемые субатлантическим периодом голоцена; 7 – находки сурков, датируемые поздним плейстоценом–голоценом; 8 – находки сурков, датируемые суббореальным и субатлантическим периодами голоцена ; a – Айдашинская пещера, b – поселение Чекист, c – Еловское поселение, d – поселение Милованово 3.
Целью данной работы являлось изучение истории ареалов рода Marmota и отдельных его представителей (Marmota bobak, M. baibacina, M. kastschenkoi) на территории юго-востока Западной Сибири в голоцене.
МАТЕРИАЛ И МЕТОДИКА
Район исследования – юго-восточная часть Западно-Сибирской равнины в подтаежной, лесостепной и степной зонах, а также Салаирский кряж, Кузнецкий Алатау и Восточный Саян (Гвоздецкий, 1968). Это территория от р. Иртыш на западе до р. Енисей на востоке и от г. Томск и г. Ачинск на севере до г. Павлодар и г. Абакан на юге, которая в административном отношении занимает Омскую, Новосибирскую, Кемеровскую области, Республику Хакасия, Алтайский край, смежные районы Томской обл,, Красноярского края, а также северо-восточные районы Республики Казахстан (рис. 1).
Источниками субфоссильных материалов послужили зоогенные отложения карстовых полостей, археологические памятники и смешанные комплексы в отложениях карстовых полостей. Датирование по вмещающим отложениям может удревнить остатки сурка, поскольку кости погибающих в своих норах землероев могут оказываться в этих отложениях по окончании их формирования.
Зоогенный материал датирован на основании данных о стратиграфии отложений и по сопутствующим остаткам фаун, в связи с чем часть его имеет широкую дату – голоцен (10 300–300 л.н.). Материал из археологических памятников датирован, исходя из их культурной принадлежности (Троицкая, Новиков, 2004). По некоторым местонахождениям имеются радиоуглеродные даты (табл. 5). Таким образом, голоценом датированы кости сурка из 45 местонахождений (рис. 1, табл. 1). Для удобства анализа датированные находки сурка соотнесены с климато-стратиграфическими подразделениями голоцена (Хотинский и др., 1991).
Таблица 1.
Местонахождения остатков представителей рода Marmota на юго-востоке Западной Сибири
| Номер п/п | Местонахождение | Тип местонахождения | Широта | Долгота | Число остатков | Период голоцена |
|---|---|---|---|---|---|---|
| 1 | Кашкулакская | Пещера | 54.47 | 89.67 | 1 | SA2 + голоцен |
| 2 | Страшная | Пещера | 51.44 | 83.16 | 157 | SA2 + голоцен |
| 3 | Горный Елбан | Поселение | 52.89 | 86.05 | 11 | SA1 |
| 4 | Исаковский 1 | Могильник | 55.45 | 74.26 | 2 | SA1 |
| 5 | Еленева | Пещера | 55.96 | 92.58 | 28 | SA1 + голоцен |
| 6 | Дивногорская 3 | Пещера | 55.84 | 92.22 | 1 | SA1 |
| 7 | Кучерла 1 | Культовое место | 50.14 | 86.33 | 7 | SA1-3 |
| 8 | Еловское | Поселение | 55.57 | 83.42 | 7 | SB3 |
| 9 | Костёнкова Избушка | Поселение | 52.30 | 84.50 | 5 | SB3 |
| 10 | Кучерла 1 | Культовое место | 50.14 | 86.33 | 2 | SB2 |
| 11 | Милованово 3 | Поселение | 54.09 | 81.55 | 1 | SB3 |
| 12 | Новоильинка | Поселение | 53.58 | 79.27 | 7 | SB3 |
| 13 | Фирсово 17 | Поселение | 53.27 | 83.55 | 1 | SB3 |
| 14 | Фирсово 4 | Поселение | 53.27 | 83.55 | 1 | SB3 |
| 15 | Чебаки | Поселение | 54.55 | 89.22 | 3 | SB3 |
| 16 | Чекист | Поселение | 56.37 | 84.49 | 4 | SB3 |
| 17 | Копёны | Могильник | 53.73 | 78.04 | + | SB3 |
| 18 | Александровское 4 | Поселение | 53.97 | 74.86 | 3 | SB2 |
| 19 | Берёзовая Лука | Поселение | 52.40 | 82.60 | 2 | SB2 |
| 20 | Итколь 2 | Могильник | 54.56 | 90.09 | 8 | SB2 |
| 21 | Ростовка | Могильник | 55.02 | 73.58 | 32 | SB2 |
| 22 | Кoзыреевская | Пещера | 55.83 | 92.15 | 1 | BO2 |
| 23 | Кучерла 1 | Культовое место | 50.14 | 86.33 | 21 | SB3 |
| 24 | Айдашинская | Пещера | 56.20 | 90.31 | 461 | Голоцен |
| 25 | Археологическая | Пещера | 54.41 | 89.41 | 6 | Голоцен |
| 26 | Биджинская | Пещера | 53.96 | 91.00 | 4 | Голоцен |
| 27 | Денисова | Пещера | 51.40 | 84.68 | 16 | Голоцен |
| 28 | Дивногорская 2 | Пещера | 55.84 | 92.22 | 3 | Голоцен |
| 29 | Жемчужная | Пещера | 55.84 | 92.16 | 2 | Голоцен |
| 30 | Караульная 1 | Пещера | 55.99 | 92.59 | 22 | Голоцен |
| 31 | Караульная 2 | Пещера | 55.99 | 92.59 | 1 | Голоцен |
| 32 | Карман 1 | Грот | 55.98 | 92.63 | 2 | Голоцен |
| 33 | Ледопадная | Пещера | 55.94 | 92.62 | 1 | Голоцен |
| 34 | Ледяная | Пещера | 55.76 | 93.20 | 4 | Голоцен |
| 35 | Ловушка | Пещера | 55.76 | 93.20 | 1 | Голоцен |
| 36 | Майская | Пещера | 55.84 | 92.16 | 1 | Голоцен |
| 37 | Малояломанская | Пещера | 50.49 | 86.59 | 21 | Голоцен |
| 38 | Малютка | Грот | 51.44 | 83.16 | 1 | Голоцен |
| 39 | Проскурякова | Пещера | 54.45 | 89.46 | 1 | Голоцен |
| 40 | Совиный навес | Грот | 55.97 | 92.56 | + | Голоцен |
| 41 | Трех Филинят | Грот | 51.44 | 83.16 | 15 | Голоцен |
| 42 | Тютютника | Пещера | 51.32 | 84.69 | 22 | Голоцен |
| 43 | Фанатиков | Пещера | 52.99 | 89.63 | 46 | Голоцен |
| 44 | Чагырская | Пещера | 51.44 | 83.16 | 4 | Голоцен |
| 45 | Чарышский навес | Грот | 50.89 | 84.73 | 19 | Голоцен |
Таблица 2.
Частоты проявления морфотипов положения слезной кости Marmota bobak, M. baibacina и M. kastschenkoi с указанием ошибки, %
| Вид | Частота встречаемости морфотипа, % | Число экземпляров | ||||
|---|---|---|---|---|---|---|
| a | b | c | d | e | ||
| M. bobak | 14 ± 2.6 | 68 ± 3.6 | 9 ± 2.2 | 8 ± 2.1 | 1 ± 0.1 | 173 |
| M. baibacina | 0 | 100 ± 0 | 0 | 0 | 0 | 218 |
| M. kastschenkoi | 18 ± 5.3 | 74 ± 6.1 | 8 ± 3.8 | 0 | 0 | 52 |
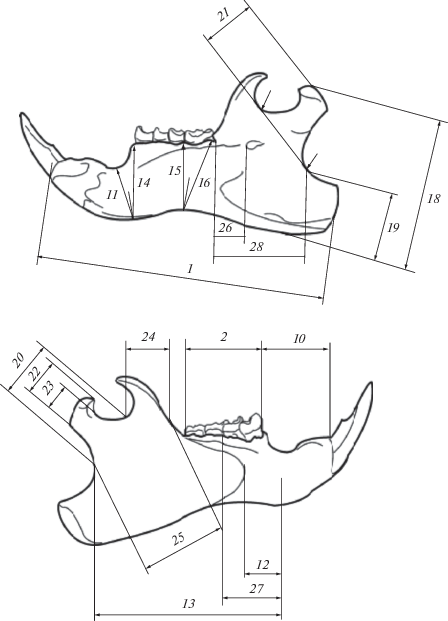
Для разработки методики диагностики современных видов использованы черепа с нижними челюстями от 68 особей M. bobak (коллекция ЗИН РАН), 58 особей M. baibacina (коллекция Музеев ИЭРиЖ УрО РАН и ИСЭЖ СО РАН) и 51 особи M. kastschenkoi (коллекции Музея ИСиЭЖ СО РАН и Зоологического музея ТГУ). С черепов (рис. 2) снималось 20 промеров (Гасилин, Косинцев, 2011). Набор промеров нижней челюсти (Гасилин, Косинцев, 2011) дополнен промерами 26–29 (рис. 3), а промеры 4–7 не снимались и не анализировались по причине редкости субфоссильных челюстей с зубами.
Рис. 2.
Схема промеров черепа представителя рода Marmota (по: Гасилин, Косинцев, 2011): 1 – кондилобазальная длина, 2 – основная длина, 3 – альвеолярная длина, 4 – длина M3, 5 – ширина M3, 6 – ширина I, 7 – поперечник I, 8 – длина диастемы, 9 – косая длина лицевой части черепа, 10 – длина лба от носовых костей до основания надглазничных отростков, 11 – длина рострума, 12 – ширина основания рострума, 13 – ширина верхней челюсти между внутренними краями подглазничных отверстий, 14 – ширина в предглазничных буграх, 15 – ширина лба между предглазничными вырезками, 16 – ширина заглазничная, 17 – ширина в мыщелках, 18 – высота лицевого отдела от небной поверхности до носовой кости, 19 – высота затылочной кости, 20 – высота надглазничных отростков над лобной поверхность.
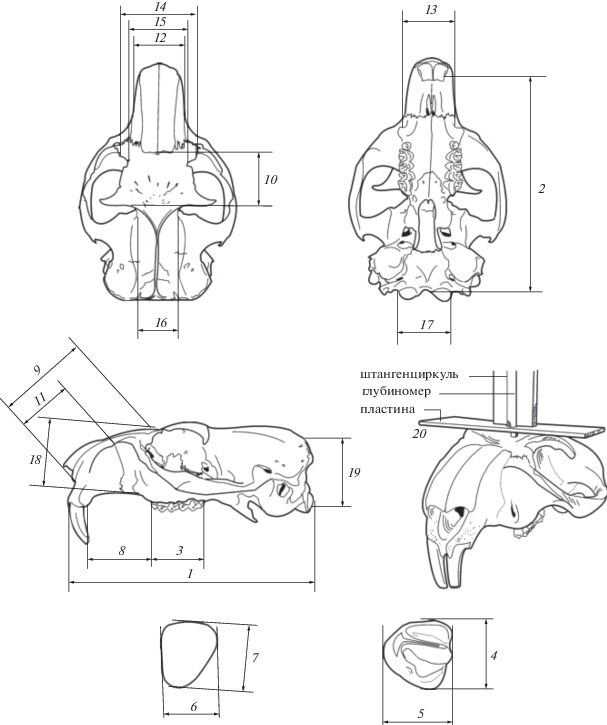
Рис. 3.
Схема промеров нижней челюсти представителя рода Marmota (по: Гасилин, Косинцев, 2011; с изменениями): 1 – ангулярная длина, 2 – альвеолярная длина p4-m3, 8 – ширина I (на рисунке не указана), 9 – поперечник I (на рисунке не отмечен), 10 – длина диастемы, 11 – высота резцового отдела на уровне подбородочного отверстия, 12 – наименьшее расстояние между подбородочным отверстием и краем массетерной площадки, 13 – наименьшее расстояние между подбородочным отверстием и краем задней вырезки, 14 – высота челюсти перед p4, 15 – высота тела перед m3 лингвальная, 16 – высота тела за m3, 17 – толщина тела между m2 и m3 (на рисунке не указана), 18 – высота челюсти по сочленовному отростку, 19 – ширина основания углового отдела, 20 – ширина основания сочленовного отростка, 21 – длина сочленовного отростка, 22 – ширина головки сочленовного отростка, 23 – ширина шейки сочленовного отростка, 24 – ширина основания венечного отростка, 25 – ширина восходящей ветви, 26 – расстояние от заднего края m3 до переднего края заднечелюстного отверстия, 27 – расстояние от подбородочного отверстия до заднего края альвеолы m1, 28 – расстояние от заднего края альвеолы m3 до заднего края челюсти, 29 – минимальное расстояние от подбородочного отверстия до вырезки между сочленовным и коронарным отростками (на рисунке не указана).
Материалом для видовой диагностики послужили целые и фрагментированные черепа (n = 4) и нижние челюсти (n = 18) из четырех местонахождений. Все черепа и 14 нижних челюстей найдены в Айдашинской пещере, расположенной в Красноярском крае в окрестностях г. Ачинск (56°12′ с.ш., 90°18′ в.д.). Остеологический материал из этой пещеры датирован в широких пределах – голоценом (Оводов, 1980). Четыре нижние челюсти найдены в археологических памятниках позднего бронзового века – конец суббореального периода (SB3, 3200–2600 л.н.) – поселениях Еловское (n = 2), Чекист (n = 1) и Милованово 3 (n = 1). Еловское располагается в Кожевниковском районе Томской обл. (55°34′ с.ш., 83°25′ в.д.); Чекист – в Томском районе Томской обл. (56°22′ с.ш., 84°29′ в.д.), Милованово 3 – в Ордынском районе Новосибирской обл. (54°5′ с.ш., 81°33′ в.д.).
Коллекции хранятся в Институте археологии и этнографии СО РАН (г. Новосибирск) и Музее Института экологии растений и животных УрО РАН (г. Екатеринбург).
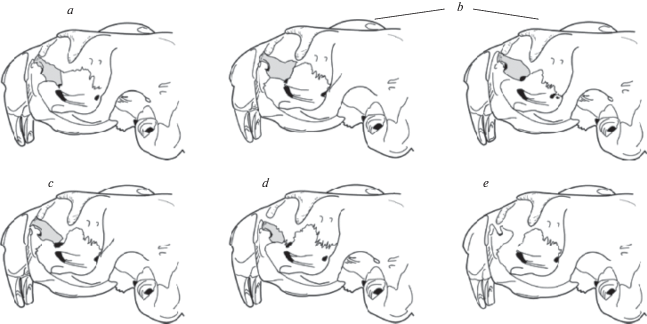
Измерения проводили электронным штангенциркулем с точностью до 0.1 мм. Статистические расчеты выполнены с помощью пакета программ Statistica 6. В качестве метода видовой диагностики современных и субфоссильных выборок использовали два варианта дискриминантного анализа: стандартный и пошаговый с включением. У сурков половой диморфизм по размерным показателям не превышает межвидовой (Cardini, 2004), поэтому он существенно не влияет на результаты видовой диагностики и его можно не учитывать при анализе. Ввиду фрагментированности субфоссильных остатков, статистические модели для классификации строились специально почти для каждого черепа и нижней челюсти. Для видовой идентификации субфоссильных черепов помимо морфометрических признаков проведен анализ морфотипов слезной кости (рис. 4). В качестве критерия достоверности различия долей морфотипических признаков в двух выборках использовали φ-критерий Фишера.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Диагностика рецентных представителей рода Marmota
Морфотипические признаки. Слезная кость описывается морфотипами a, b, c, d и e у степного сурка, морфотипом b – у серого (Гасилин, Косинцев, 2011). Слезной кости лесостепного сурка свойственны морфотипы a, b и c. Частоты этих морфотипов у лесостепного и степного сурков практически одинаковы, а морфотип b является самым распространенным у всех трех видов (табл. 2).
Для степного сурка диагностическими являются морфотипы d и e. Абсолютное преобладание (100%) у серого сурка морфотипа b можно использовать для выявления его наличия при групповой диагностике: поскольку доля морфотипа b у степного сурка составляет 68%, а у лесостепного – 74%, то в репрезентативной выборке значения, превышающие 75%, будут указывать на присутствие в этой выборке серого сурка.
Морфометрические признаки. Метрические данные по выборкам рецентных сурков представляли собой обучающие группы в стандартном и пошаговом с включением дискриминантном анализе. Верность идентификации в выборках по полному набору признаков как для черепа, так и для нижней челюсти составила 97–98% (табл. 3, 4). Валидность полученной статистической модели проверена с помощью тестовых выборок, включавших по три нижние челюсти каждого вида. В результате тестовые экземпляры определены верно. Таким образом, руководствуясь метрическими признаками на черепе и нижней челюсти, можно с высокой вероятностью диагностировать все три вида. Степень дифференциации лесостепного сурка от алтайского и степного по размерам черепа и нижней челюсти близка к 100% (рис. 5).
Таблица 3.
Коэффициенты канонических и классификационных функций по 20 признакам черепа M. bobak, M. baibacina и M. kastschenkoi
| Признак | Коэффициенты канонических функций | Коэффициенты классификационных функций | |||||
|---|---|---|---|---|---|---|---|
| стандартизованные | нестандартизованные | M. bobak | M. baibacina | M. kastschenkoi | |||
| 1 | 2 | 1 | 2 | ||||
| 1 | –0.33 | –0.12 | –0.09 | –0.03 | 0.09 | 0.39 | 0.73 |
| 2 | –1.01 | –0.09 | –0.29 | –0.03 | 3.42 | 4.59 | 5.47 |
| 3 | –0.35 | 0.10 | –0.45 | 0.12 | 28.11 | 30.31 | 31.19 |
| 4 | 0.35 | –0.10 | 1.18 | –0.34 | 10.30 | 4.53 | 2.24 |
| 5 | –0.26 | –0.06 | –1.03 | –0.22 | 34.58 | 38.46 | 41.99 |
| 6 | 0.15 | –0.25 | 0.45 | –0.73 | 9.50 | 5.94 | 6.87 |
| 7 | –0.12 | 0.41 | –0.33 | 1.08 | –4.26 | –0.45 | –2.77 |
| 8 | 0.35 | 0.34 | 0.26 | 0.25 | –0.41 | –0.93 | –2.41 |
| 9 | –1.10 | –0.40 | –0.60 | –0.22 | 3.38 | 5.45 | 7.79 |
| 10 | 0.47 | –0.05 | 0.39 | –0.04 | 4.64 | 2.88 | 1.90 |
| 11 | 0.71 | –0.38 | 0.47 | –0.26 | 4.03 | 1.44 | 0.88 |
| 12 | –0.18 | 0.97 | –0.11 | 0.58 | –4.41 | –2.67 | –4.09 |
| 13 | 0.15 | –0.10 | 0.10 | –0.07 | –3.16 | –3.73 | –3.81 |
| 14 | 0.13 | –1.16 | 0.06 | –0.50 | 0.49 | –0.87 | 0.46 |
| 15 | –0.30 | 0.45 | –0.17 | 0.25 | –3.18 | –1.93 | –2.19 |
| 16 | –0.07 | –0.01 | –0.05 | 0.00 | 9.12 | 9.33 | 9.49 |
| 17 | –0.27 | 0.11 | –0.25 | 0.10 | 10.39 | 11.67 | 12.08 |
| 18 | 0.76 | 0.31 | 0.60 | 0.25 | 6.38 | 4.37 | 1.96 |
| 19 | 0.73 | –0.18 | 0.48 | –0.12 | –6.39 | –8.72 | –9.72 |
| 20 | 0.59 | –0.22 | 0.94 | –0.36 | –7.44 | –12.23 | –13.80 |
| Константы | 16.76 | 4.85 | –809.87 | –867.34 | –935.13 | ||
| Собственные значения | 8.73 | 1.52 | Верность идентификации, % | ||||
| Объясненная дисперсия | 0.85 | 1.00 | 100 | 95 | 93.8 | ||
Таблица 4.
Коэффициенты канонических и классификационных функций по 24 признакам нижних челюстей M. bobak, M. baibacina и M. kastschenkoi
| Признак | Коэффициенты канонических функций | Коэффициенты классификационных функций | |||||
|---|---|---|---|---|---|---|---|
| стандартизованные | нестандартизованные | M. bobak | M. baibacina | M. kastschenkoi | |||
| 1 | 2 | 1 | 2 | ||||
| 1 | –0.04 | –0.24 | –0.01 | –0.07 | 0.35 | 0.21 | 0.45 |
| 2 | –0.49 | 0.11 | –0.58 | 0.13 | 28.01 | 30.34 | 31.85 |
| 8 | 0.11 | 0.47 | 0.36 | 1.53 | –12.72 | –10.08 | –15.52 |
| 9 | –0.16 | –0.32 | –0.38 | –0.75 | 17.71 | 17.12 | 20.44 |
| 10 | 0.04 | 0.10 | 0.03 | 0.09 | –0.01 | 0.11 | –0.25 |
| 11 | 0.74 | –0.06 | 0.83 | –0.07 | –5.29 | –8.31 | –10.75 |
| 12 | 0.06 | 0.35 | 0.06 | 0.35 | –1.38 | –0.69 | –1.87 |
| 13 | 0.38 | –0.16 | 0.16 | –0.07 | 3.85 | 3.11 | 2.77 |
| 14 | –0.29 | 0.08 | –0.24 | 0.06 | –5.30 | –4.32 | –3.73 |
| 15 | 0.52 | 0.10 | 0.42 | 0.08 | –15.63 | –16.88 | –18.46 |
| 16 | –0.12 | –1.04 | –0.10 | –0.91 | 6.89 | 4.94 | 7.81 |
| 17 | 0.09 | 0.34 | 0.20 | 0.76 | 18.62 | 19.84 | 17.08 |
| 18 | 0.63 | 0.59 | 0.32 | 0.30 | –5.55 | –5.88 | –7.72 |
| 19 | –1.05 | 0.50 | –0.64 | 0.30 | 4.68 | 7.64 | 8.84 |
| 20 | 0.20 | 0.53 | 0.11 | 0.29 | –0.01 | 0.35 | –0.80 |
| 21 | –0.38 | –0.16 | –0.34 | –0.14 | 8.00 | 8.81 | 10.30 |
| 22 | 0.52 | –0.95 | 0.60 | –1.10 | –15.67 | –20.52 | –19.35 |
| 23 | –0.35 | 0.77 | –0.48 | 1.07 | 20.87 | 25.23 | 23.81 |
| 24 | –0.20 | 0.24 | –0.18 | 0.21 | –0.49 | 0.65 | 0.62 |
| 25 | –0.91 | –0.05 | –0.68 | –0.04 | 4.52 | 6.74 | 9.01 |
| 26 | –0.39 | 0.26 | –0.48 | 0.32 | 12.87 | 15.34 | 15.99 |
| 27 | –0.48 | –0.49 | –0.53 | –0.54 | 13.11 | 13.55 | 16.75 |
| 28 | 0.47 | –0.27 | 0.31 | –0.18 | –8.81 | –10.35 | –10.85 |
| 29 | –0.37 | –0.04 | –0.19 | –0.02 | 6.57 | 7.18 | 7.85 |
| Константы | 25.5 | 0.4 | –602.21 | –684.52 | –773.11 | ||
| Собственные значения | 6.94 | 1.71 | Верность идентификации, % | ||||
| Объясненная дисперсия | 0.80 | 1.00 | 97.9 | 100.0 | 97.5 | ||
Таблица 5.
Верность идентификации и апостериорные вероятности классификации субфоссильных черепов и нижних челюстей (M. bobak, M. baibacina и M. kastschenkoi)
| Признаков в модели | Верность идентификации, % | Экземпляр | Апостериорные вероятности | Результат диагностики | ||
|---|---|---|---|---|---|---|
| bob | bai | kas | ||||
| Череп | ||||||
| 16 | 98.1 | 95.1 | 97.4 | Айдашинская 1 | 0.99 | kas |
| 14 | 98.1 | 89.1 | 97.4 | Айдашинская 2 | 0.94 | bai |
| 6 | 90.2 | 79.1 | 91.3 | Айдашинская 3 | 0.71 | bai |
| 3 | 71.0 | 42.3 | 73.9 | Айдашинская 4 | 0.59 | bob |
| Нижняя челюсть | ||||||
| 7 | 84.21 | 67.24 | 83.33 | Еловское 1 | 0.94 | kas |
| 9 | 82.35 | 73.21 | 90.0 | Еловское 2 | 0.97 | kas |
| 14 | 98.68 | 91.23 | 88.1 | Айдашинская 5 | 0.98 | kas |
| 18 | 98.21 | 98.21 | 97.62 | Айдашинская 6 | 0.84 | kas |
| 20 | 100.00 | 98.21 | 95.24 | Айдашинская 7 | 0.97 | kas |
| 9 | 89.29 | 78.95 | 90.48 | Айдашинская 8 | 0.53 | bob |
| 11 | 91.07 | 87.50 | 78.57 | Айдашинская 9 | 0.88 | bai |
| 12 | 94.64 | 84.21 | 92.86 | Айдашинская 10 | 0.75 | bob |
| 8 | 88.16 | 91.23 | 85.71 | Айдашинская 11 | 0.63 | kas |
| 9 | 98.25 | 80.36 | 83.33 | Айдашинская 12 | 0.89 | bai |
| 4 | 75.32 | 68.97 | 66.67 | Айдашинская 13 | 0.50 | kas |
| 2 | 61.40 | 42.11 | 76.19 | Айдашинская 14 | 0.65 | kas |
| 3 | 64.91 | 40.35 | 76.19 | Айдашинская 15 | 0.53 | kas |
| 3 | 77.92 | 55.17 | 71.43 | Айдашинская 15 | 0.60 | bai |
| 3 | 76.62 | 58.62 | 66.67 | Айдашинская 16 | 0.95 | bob |
| 3 | 80.52 | 61.40 | 52.38 | Айдашинская 17 | 0.99 | bob |
| 9 | 85.96 | 78.94 | 88.37 | Милованово 3 | 0.52 | bai |
| 6 | 72.66 | 66.07 | 90.00 | Чекист | 0.74 | kas |
Примечание. В последнем столбце указаны результаты видовой диагностики. Жирным шрифтом выделены достаточно надежно диагностированные до вида экземпляры (апостериорная вероятность не менее 0.80 при верности идентификации модели не менее 80% для каждого вида): bob – M. bobak, bai – M. baibacina, kas – M. kastschenkoi.
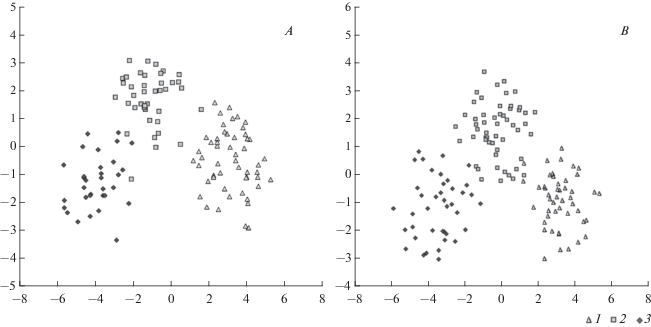
Диагностика субфоссильных краниологических остатков
Слезные кости немногочисленных в исследованной выборке субфоссильных черепов описываются исключительно морфотипом b. Это не позволило провести видовую идентификацию.
По морфометрическим признакам до вида определено 2 черепа и 4 нижние челюсти из двух местонахождений (табл. 5). Нижние челюсти из местонахождений Чекист и Милованово 3 диагностировать до вида не удалось. Нижние челюсти сурков из Еловского поселения принадлежат лесостепному сурку. Наибольший интерес представляют результаты диагностики субфоссильных остатков из Айдашинской пещеры. Здесь серому сурку принадлежали один череп и одна нижняя челюсть, лесостепному – один череп и три нижние челюсти (табл. 4).
Реконструкция ареала рода Marmota
В районе исследования сурки найдены в составе одного местонахождения раннеголоценового возраста (9680–8860 л.н.), четырнадцати местонахождений среднеголоценового (3800–2600 л.н.), семи позднеголоценового (2500–300 л.н.) и двадцати трех местонахождений голоценового возраста.
Самая северная точка находок представителей рода в Прииртышье – могильник Исаковский 1 (55°27′ с.ш.), датированный археологическим и радиоуглеродным методами началом субатлантического периода позднего голоцена. Самая северная точка находки остатков сурка в Приобье – поселение Чекист (56°36′ с.ш.), датируемое концом суббореального периода (SB3, 3200–2600 т.л.н.) голоцена. В бассейне р.Енисей известны местонахождения костей сурка, расположенные примерно на 56°20′ с.ш. (пещеры Еленева, Караульная 1), датируемые ранним (PB–BO, 9600–8000 л.н.) и средним голоценом (AT, 8000–5000 л.н.) (табл. 5). Самые северные находки остатков сурков, датированные в широком диапазоне голоценом, происходят из Айдашинской пещеры (56°12′ с.ш.) и пещер на р. Енисей (Ледопадная, Казыреевские и др., около 56° с.ш.). Эти данные показывают, что в раннем и среднем голоцене северная граница распространения рода Marmota на юго-востоке Западной Сибири проходила между 56 и 57° с.ш. В начале позднего голоцена в Прииртышье она доходила до 55°27′ с.ш. Её положение в Приобье не изменилось до наших дней, т.к. сейчас здесь ее граница проходит на той же широте, что и в конце среднего голоцена. На р. Енисей положение границы в позднем голоцене точно не известно. В историческое время (18–19 вв.) граница ареала сурков в Приртышье не изменилась, поскольку в конце 19 в. следы обитания сурков отмечены на 55° с.ш. (Огнев, 1947). Осталась она неизменной и в Приобье, а на р. Енисей к 17 в. граница сместилась на юг и запад, т.к. в письменных источниках сведений об обитании сурков в окрестностях г. Красноярска и на правом берегу р. Енисея не имеется.
В центральной и южной части рассматриваемой территории имеются многочисленные местонахождения остатков сурков среднего голоцена и несколько местонахождений позднего голоцена (рис. 1). Отмечены они здесь и в историческое время. В 1730-х гг. при описании Колыванских заводов упоминается обилие сурков в приалтайской лесостепи. В середине 18 в. между реками Алей и Таловка на безлесных склонах П.-С. Паллас застает множество сурков (Кириков, 1966). Все это свидетельствует, что южная граница ареала рода на рассматриваемой территории в голоцене существенно не изменялась.
Таблица 6.
Список радиоуглеродных дат местонахождений остатков представителей рода Marmota на юго-востоке Западной Сибири
| Таксон | Дата | Лабораторный номер |
|---|---|---|
| Поселение Березовая Лука (52°24′ с.ш., 82°36′ в.д.) | ||
| Marmota sp. | 3125 ± 45 | СОАН-4150 |
| 3400 ± 140 | СОАН-4576 | |
| 3405 ± 90 | СОАН-4152 | |
| 3415 ± 90 | СОАН-4151 | |
| 3450 ± 50 | Ле-7426 | |
| 3450 ± 45 | Ле-8532 | |
| 3615 ± 35 | СОАН-3213 | |
| 3630 ± 85 | СОАН-7472 | |
| 3690 ± 120 | Ле-7800 | |
| 3700 ± 80 | Ле-8533 | |
| 3750 ± 80 | СОАН-7471 | |
| 3860 ± 95 | СОАН-4575 | |
| 3875 ± 70 | СОАН-3755 | |
| 3990 ± 75 | СОАН-4571 | |
| 3990 ± 110 | Ле-8531 | |
| 4025 ± 90 | СОАН-7473 | |
| 4205 ± 65 | СОАН-3754 | |
| 4335 ± 120 | СОАН-3753 | |
| Еленева пещера (55°55′ с.ш., 92°18′ в.д.) | ||
| Marmota sp. | 2940 ± 40 | СОАН-23 086 |
| 4655 ± 30 | СОАН-2905 | |
| 6335 ± 60 | СОАН-22 907 | |
| Могильник Итколь 2 (54°27′ с.ш., 90°5′ в.д.) | ||
| Marmota sp. | 3940 ± 50 | Ле-11 434 |
| Кашкулакская пещера (54°28′ с.ш., 89°40′ в.д.) | ||
| Marmota sp. | 2210 ± 100 | СОАН-849 |
| Пещера Караульная (55°59′ с.ш., 92°35′ в.д.) | ||
| Marmota baibacina | 8860 ± 140 | СОАН-1084 |
| Пещера Страшная (51°9′ с.ш., 83°0′ в.д.) | ||
| Marmota sp. | 1698 ± 446 | NSKA-1010 |
Реконструкция ареалов отдельных видов
Установлено, что лесостепной сурок в конце суббореального периода голоцена обитал, как и в настоящее время, в районе Новосибирского Приобья и отчасти Томского Приобья, что подтверждается находками его костей в Еловском поселении. Вместе с тем этот вид обитал в прошлом на значительно большей территории, чем в настоящее время. Его ареал доходил на восток, по крайней мере, до верхнего течения р. Чулым, где голоценовые остатки лесостепного сурка найдены в пещере Айдашинской. Лесостепной сурок изначально описывался исследователями как серый, иногда выделяемый в качестве подвида такового (Огнев, 1947). В связи с этим южную границу его распространения, как и северную границу распространения серого сурка даже в недавнем прошлом восстановить затруднительно. Вероятно, в последние столетия лесостепной сурок всегда имел ограниченный ареал. В середине 20 в. отмечают резкое снижение его численности в результате систематической добычи (Лаптев, 1958). По данным Западносибирского филиала ВНИИОЗ на 2014 год, его численность за последние годы не претерпевала значительных изменений и даже имела положительную тенденцию.
Первые письменные сведения о распространении серого сурка на Алтае появляются с начала 20 в. По данным разных авторов (Огнев, 1947; Юдин и др., 1979; и др.), в это время серый сурок населял большую часть горного Алтая. Не было его лишь в таежном Северном и, отчасти, в Северо-Восточном Алтае, а также в лесной юго-западной части Усть-Коксинского района республики Алтай. Границы ареала и численность серого сурка в 20 в. были подвержены некоторым колебаниям, основной причиной которых обычно видят прямое истребление человеком (Юдин и др., 1979). О тенденции изменения его ареала в 21 в. в пределах исследуемого региона трудно судить ввиду недостаточности сведений.
Достоверно диагностированные субфоссильные кости степного сурка отсутствуют. Письменные сведения о распространении этого вида на юго-востоке Западной Сибири относятся к 19 в. Тогда северная граница его расселения доходила по р. Иртыш на север до 55° с.ш. В конце 19 в. сурка отмечали в окрестностях г. Омска (Огнев, 1947). Сокращение численности и изменение ареала степного сурка началось с 1865 г. – с массового переселения крестьян в лесостепную зону (Кириков, 1966). С этого времени ареал стал принимать современные очертания, отступая на юг и разделяясь на изолированные части. По данным Западносибирского филиала ВНИИОЗ на 2008 г., численность степного сурка растет, а ареал становится менее мозаичным (Машкин, 2009).
ЗАКЛЮЧЕНИЕ
Сопряженный анализ результатов определения субфоссильных краниологических остатков из голоценовых местонахождений и литературных данных 19–20 вв. позволил воссоздать изменения ареала рода Marmota и отдельных его представителей на юго-востоке Западной Сибири в голоцене. В рамках решения поставленной цели показана возможность использования дискриминантного анализа для диагностики краниологических остатков сурков. При этом степень дифференциации лесостепного сурка от серого и степного сурков по полному набору морфометрических признаков близка к 100%, что согласуется с представлениями о нем, как о самостоятельном виде.
География находок остатков сурков указывает, что северная граница ареала рода Marmota на протяжении голоцена проходила по южной границе лесной зоны, проникала в глубь этой территории вдоль долины р. Обь, затем, проходя по горам Южной Сибири, пересекала р. Енисей на широте около 56° с.ш. В письменных источниках не фиксируется обитание сурков в бассейне р. Чулым и в районе г. Красноярска, и можно утверждать, что отступление ареала произошло до момента русской колонизации, т.е. более 300 л.н. К этому времени в бассейне р. Енисей граница сместилась на юг и запад, к Западному Саяну, и сурок исчез с восточного берега р. Енисей. В первой трети 20 в. северная граница ареала рода в бассейне р. Иртыш сместилась к югу (Огнев, 1947).
Видовая диагностика краниологических остатков сурков показала, что ареалы серого и лесостепного сурков в прошлом были шире и захватывали территорию в верхнем течении р. Чулым (56°12′ с.ш., 90°18′ в.д.), где, видимо, перекрывались. Современная граница расселения серого сурка проходит значительно южнее (между 51° и 52° с.ш.), а лесостепного – западнее (между 85° и 86° в.д.) указанного района (рис. 1). Очертания ареала степного сурка в голоцене точно не известны. В последние 100–150 лет произошли заметное сокращение и фрагментация ареалов всех трех видов, вызванные как распространением земледелия, так и прямым истреблением.
Список литературы
Абдулганеев Н.Т., 1996. О хозяйстве населения лесостепного и предгорного Алтая в скифское время // Археология, антропология, этнография Сибири. Барнаул. С. 145–153.
Агаджанян А.К., 2007. Позвоночные позднего плейстоцена из пещеры Сурка в долине Ануя (Горный Алтай) // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий: Материалы Годовой сессии Института археологии и этнографии СО РАН 2007 г. Новосибирск: Изд-во Ин-та археологии и этнографии СО РАН. Т. 13. С. 4–7.
Бибиков Д.И., 1989. Сурки. М.: Агропромиздат. 225 с.
Бибиков Д.И., Берендяев С.А., 1978. Серый сурок // Сурки. Распространение и экология. М.: Наука. С. 39–66.
Брандлер О.Д., 2003. К видовой самостоятельности лесостепного сурка Marmota kastschenkoi (Rodentia, Sciuridae, Marmotinae) // Зоологический журнал. Т. 82. Вып. 12. С. 1498–1505.
Брандлер О.В., Ляпунова Е.А., Банникова А.А., Крамеров Д.А., 2010. Филогения и систематика сурков (Marmota, Sciuridae, Rodentia), основанные на данных интер-SINE-ПЦР // Генетика. Т. 46. № 3. С. 321–331.
Машкин В. И., 2009. Сурки // Бюллетень состояния ресурсов охотничьих животных, их численность и добыча по регионам России в сезон 2008–2009 гг. Киров. С. 5–6.
Васильев С.К., 2013. Фауна крупных млекопитающих из плейстоценовых отложений Чагырской пещеры (северо-западный Алтай) по материалам раскопок 2007–2011 годов // Археология, этнография и антропология Евразии. Вып. 53. № 1. С. 28–44.
Васильев С.К., Гребнев И.Е., 1994. Фауна млекопитающих голоцена Денисовой пещеры // Денисова пещера. Ч. 1. Новосибирск: Наука. С. 167–180.
Васильев С.К., Деревянко А.П., Маркин С.В., 2006. Фауна крупных млекопитающих финала сартанского времени Северо-Западного Алтая (по материалам пещеры Каминной) // Археология, этнография и антропология Евразии. № 2. С. 2–22.
Васильев С.К., Зенин В.Н., 2009. Фаунистические остатки из пещеры Страшная (Северо-Западный Алтай) по материалам раскопок в 1988–2008 годах // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. Новосибирск: Изд-во Института археологии и этнографии СО РАН. Т. 15. С. 56–62.
Васильев С.К., Кривошапкин А.И., Зенин В.Н., Шалагина А.В., 2016. Предварительные итоги исследования остатков мегафауны из отложений пещеры Страшной (Северо-Западный Алтай) // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. Новосибирск: Изд-во Ин-та археологии и этнографии СО РАН. Т. 22. С. 15–22.
Васильев С.К., Оводов Н.Д., Мартынович Н.В., 2006. Новые палеотериологические исследования пещеры Логово Гиены (северо-западный Алтай) // Проблемы археологии, этнографии и антропологии Сибири и сопредельных территорий. Новосибирск: Изд-во Ин-та археологии и этнографии СО РАН. Т. 7. № 1. С. 43–49.
Васильев С.К., Шуньков М.В., 2009. Крупные млекопитающие из плейстоценовых отложений в южной галерее Денисовой пещеры // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. Новосибирск: Изд-во Института археологии и этнографии СО РАН. Т. 15. С. 63–69.
Галкина Л.И., 1970. Оценка некоторых систематических особенностей сурков и географическая изменчивость горноазиатского сурка (Marmota baibacina Kastschenko, 1899) // Фауна Сибири. Новосибирск: Наука. С. 267–279.
Галкина Л.И., Епифанцева Л.Ю., Тараненко, Д.Е., Абрамов С.А., 2005. Роль эколого-географических факторов в процессе формообразования и морфологической дифференциации серых сурков (Marmota baibacina Kastschenko, 1899) // Систематика, палеонтология и филогения грызунов. Труды Зоологического института РАН. Т. 306. Санкт-Петербург. С. 41–54.
Галкина Л.И., Тараненко Д.Е., 2002. Морфологические и ареалогические особенности лесостепного сурка // Сурки в степных биоценозах Евразии: 8 совещ. по суркам стран СНГ: Россия, г. Чебоксары, 7–10 июня 2002 г.: тез. докл. (Науч. труды заповедника “Присурский”, Т.8). Чебоксары: КЛИО. С. 15–16.
Гасилин В.В., Косинцев П.А., 2011. Видовая диагностика степного (Marmota bobak) и серого (Marmota baibacina) сурков по краниологическим признакам // Зоологический журнал. Т. 90. № 12. С. 1509–1521.
Гвоздецкий Н.А., 1968. Физико-географическое районирование СССР. М.: МГУ. 576 с.
Громов И.М., Бибиков Д.И., Калабухов Н.И., Мейер М.Н., 1965. Наземные беличьи (Marmotinae). Фауна СССР. Млекопитающие. М.-Л.: Наука. Т. 3. Вып. 2. 467 с.
Громов И.М., Ербаева М.А., 1995. Млекопитающие фауны России и сопредельных территорий. Зайцеобразные и грызуны. СПб.: ЗИН РАН. 239 с.
Деревянко А.П., Агаджанян А.К., Мартынович Н.В., Оводов Н.Д., Постнов А.В. и др., 2002. Палеоэкологическая реконструкция позднего плейстоцена Канской котловины по материалам комплексных многолетних исследований Усть-Канской пещеры (горный Алтай) // Проблемы каменного века Средней и Центральной Азии. Материалы междунар. конф., посвященной 70-летию акад. АН Респ. Узбекистан Исламова (г. Ташкент, 30 сентября–6 октября 2002 г.). Новосибирск: Изд-во Института археологии и этнографии СО РАН. С. 37–48.
Зажигин В.И., 1980. Грызуны позднего плиоцена и антропогена юга Западной Сибири. М.: Наука. 156 с.
Кирюшин Ю.Ф., Гальченко А.В., Удодов В.С., Шамшин А.Б., 1988. Хозяйственно-культурные типы поздней бронзы лесостепного Алтая // Хронология и культурная принадлежность памятников каменного и бронзового веков Южной Сибири. Тез. докл. и сообщ. науч. конф. (23–25 марта 1988 г.). Барнаул: Изд-во ИИФиФ; АГУ. С. 138–142.
Кирюшин Ю.Ф., 2002. Энеолит и ранняя бронза юга Западной Сибири. Барнаул: Изд-во Алтайского университета. 294 с.
Кириков С.В., 1966. Промысловые животные, природная среда и человек. М.: Изд-во Наука. 348 с.
Косинцев П.А., 2002. Типология териокомплексов лесостепного Приобья из поселений эпохи бронзы // Фауна Урала в плейстоцене и голоцене. Сб. науч. трудов Екатеринбург: Университет. С. 162–166.
Косинцев П.А., Явшева Д.А., Девяшин М.М., 2011. Комплекс костных остатков животных из раскопок поселения Березовая Лука // Березовая Лука – поселение эпохи бронзы в Алейской степи. Барнаул: Изд-во Алтайского университета. Т. 1. С. 139–148.
Кожамкулова Б.С., 1969. Антропогеновая ископаемая териофауна Казахстана. Алма-Ата: Наука. 148 с.
Лаптев И.П., 1958. Млекопитающие таежной зоны Западной Сибири. Томск: Изд-во Томск. университета. 285 с.
Лаптев И.П., Юдин Б.С., 1952. Сурок Томской области и его охрана // Труды Томск. гос. университета. Т. 118. С. 107–130.
Лаухин С.А., Акимова Е.В., Оводов Н.Д., Дроздов Н.И., Стасюк И.В., Томилова Е.А., 1999. Геологическая обстановка среднеплейстоценовой палеолитической стоянки Усть-Ижуль 1 (юг Средней Сибири) // Вестник археологии, антропологии и этнографии. Вып. 2. С. 105–114.
Мандырка П.В., Макаров Н.П., Мартынович Н.В., Оводов Н.Д., Андренко О.В., Чеха В.П., 1999. Комплексное исследование пещеры Тугаринова // Древности Приенисейской Сибири. Вып. 1. С. 83–115.
Мерц В.К., 2002. Погребение каменного века и энеолитический комплекс стоянки Шидерты 3 // Изучение памятников археологии Павлодарского Прииртышья. Сб. науч. ст. Павлодар: НПФ ”Эко”. С. 75–102.
Молодин В.И., Ефремова Н.С., 2010. Грот Куйлю – культовый комплекс на реке Кучерле (горный Алтай). Новосибирск: Изд-во Ин-та археологии и этнографии СО РАН. 264 с.
Мотузко А.Н., Е.В. Акимова Е.В., 2013. Фауна млекопитающих из педоседимента позднепалеолитического местонахождения Дербина 5 // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. Материалы итоговой сессии ИАЭТ СО РАН 2013 г. Т. 19. Новосибирск. С. 130–134.
Оводов Н.Д., 1980. Характеристика остеологического материала из Айдашинской пещеры // Айдашинская пещера. Новосибирск: Наука. С. 97–143.
Оводов Н.Д., 1983. Изменение северо-восточной границы ареала среднего сурка в позднем антропогене // Материалы 6 Всесоюз. совещ. Ленинград, 25–28 января 1984 г. С. 26–28.
Оводов Н.Д., 2009. Млекопитающие пещеры Тютюник (Республика Алтай, Усть-Канский район) // Известия Горно-Алтайского отдела Русского географического общества. № 2. С. 178–185.
Оводов Н.Д., Мартынович Н.В., 1999. Позвоночные пещер Караулинского карстового района (окрестности Красноярска) // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. Новосибирск: Изд-во Института археологии и этнографии СО РАН. Т. 5. С. 185–192.
Оводов Н.Д., Мартынович Н.В., 2000. Позвоночные пещер Бирюсинского карстового района (окрестности Красноярска) // Проблемы реконструкции климата и природной среды голоцена и плейстоцена Сибири. Изд-во Ин-та археологии и этнографии СО РАН. Вып. 2. С. 375–382.
Оводов Н.Д., Мартынович Н.В., Михеев В.Е., 2001. Позвоночные пещер правобережья Енисея в окрестностях Красноярска // Труды гос. заповедника “Столбы”. Вып. 17. Красноярск. С. 115–144.
Оводов Н.Д., Мартынович Н.В., Надаховский А., 1998. “Филиновые ниши” на Северо-Западном Алтае как тафономический и палеоэкологический индикаторы // Палеоэкология плейстоцена и культуры каменного века Северной Азии и сопредельных территорий. Красноярск: Изд-во Ин-та археологии и этнографии СО РАН. Т. 1. С. 249–255.
Оводов Н.Д., Мартынович Н.В., Орлова Л.А., 2003. Фауна Малояломанской пещеры (Центральный Алтай) // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. № 9 (1). Новосибирск: Изд-во Ин-та археологии и этнографии. С. 192–196.
Огнев С.И., 1947. Звери СССР и прилежащих стран. Т. 5. Грызуны (продолжение). (“Звери Восточной Европы и Северной Азии”). М.-Л.: Изд-во АН СССР. 809 с.
Постановление губернатора Новосибирской области от 24 июля 2014 года № 119 Об утверждении Схемы размещения, использования и охраны охотничьих угодий на территории Новосибирской области. Приложение 1. Характеристика состояния численности и размещения охотничьих ресурсов на территории Новосибирской области, 2014. [Электронный ресурс] Режим доступа: http://www.nexplorer.ru/print/news__12953.htm Дата обновления: 16.12.2016.
Сидоров Е.А., 1989. Скотоводство лесостепного Приобья в I тысячелетии до н.э. // Советская археология. № 3. С. 141–153.
Тараненко Д.Е., 2011. Пространственные взаимоотношения двух видов сурков: Marmota kastschenkoi и M. baibacina (Rodenta, Sciuridae) // Вестник Сибирского гос. университета. Биология. Т. 3 (4). С. 220–228.
Троицкая Т.Н., Новиков А.В., 2004. Археология Западно-Сибирской равнины. Новосибирск: Изд-во Ин-та археологии и этнографии СО РАН. 136 с.
Хотинский Н.А., Алешинская З.В., Гуман М.А., Климанов В.А., Черкинский А.Е., 1991. Новая схема периодизации ландшафтно-климатических изменений в голоцене // Известия АН СССР. Сер. геогр. № 3. С. 30–42.
Черников С.С., 1960. Восточный Казахстан в эпоху бронзы. М.: Изд-во Академии наук СССР. 285 с.
Юдин Б.С., Галкина Л.И., Потапкина А.Ф., 1979. Млекопитающие Алтае-Саянской горной страны. Новосибирск: Наука. 296 с.
Agadjanian A.K., Serdyuk N.V., 2005. History of mammalian communities and paleogeography of Altai Mountains in Paleolithic // Paleontological Journal. V. 39. Suppl. 6. P. 645–821.
Cardini A., 2004. Evolution of marmots (Rodentia, Sciuridae): combining information on labial and lingual sides of the mandible // Acta Theriologica. V. 3. № 49. P. 301–318.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал