

Зоологический журнал, 2019, T. 98, № 6, стр. 665-672
Гнездование совообразных в дуплянках на юго-востоке Приморского края
В. П. Шохрин 1, *, Д. В. Соловьева 2, **, С. Л. Вартанян 3, ***
1 Объединенная дирекция Лазовского государственного природного заповедника им. Л.Г. Капланова и национального парка “Зов тигра”
692980 Приморский край, с. Лазо, Россия
2 Институт биологических проблем Севера ДВО РАН
685000 Магадан, Россия
3 Северо-Восточный Комплексный научно-исследовательский институт ДВО РАН
685000 Магадан, Россия
* E-mail: shokhrin@mail.ru
** E-mail: diana_solovyova@mail.ru
*** E-mail: sergey-vartanayn@mail.ru
Поступила в редакцию 11.07.2018
После доработки 29.08.2018
Принята к публикации 11.10.2018
Аннотация
На юго-востоке Приморского края нами установлено, что три вида совообразных (длиннохвостая неясыть, ошейниковая совка и ушастая сова) могут гнездиться в искусственных гнездах – дуплянках и благополучно выводить в них потомство. Доказано, что дефицит естественных дупел для сов в этом регионе ощущается только в годы, когда наблюдаются пики численности грызунов и массовое размножение сов. Максимально длиннохвостая неясыть занимала 15.9% предоставленных ей дуплянок, а ошейниковая совка – 6.7%. В годы депрессии или средней численности мышевидных грызунов совы в дуплянках гнездились единично. Длиннохвостая неясыть и ушастая сова (единственное гнездо) занимали только дуплянки типа “гнездовая труба”, а ошейниковая совка гнездилась как в “трубах”, так и в ящиках – “гоголятниках”. Для обоих видов сов не отмечены предпочтения дуплянок, расположенных на определенной высоте или имеющих леток определенной ориентации. Успех гнездования сов в искусственных гнездах (от откладки яиц до вылета молодняка) составил 92.9% у длиннохвостой неясыти и 57.1% у ошейниковой совки.
Из девяти видов сов, гнездящихся на юго-востоке Приморского края, шесть видов являются облигатными дуплогнездниками и нуждаются в фаутных деревьях для устройства гнезд. В настоящее время повсеместно на неохраняемых территориях происходит интенсивная рубка лесов и, как следствие, изменение их возрастного и размерного состава. В результате рубок возникает нехватка гнездовых полостей для птиц-дуплогнездников, поэтому любой положительный опыт привлечения совообразных в искусственные гнезда имеет большое значение для перспектив их сохранения. Специальных мероприятий по привлечению сов для гнездования в дуплянках в Приморье не проводилось. В 1960-е годы ящики – “гоголятники”, развешенные для мандаринок (Aix galericulata) в долине р. Киевка, Приморье, занимали ошейниковые совки (Otus bakkamoena) (Поливанов и др., 1971).
В Евразии привлечение сов на гнездование в дуплянки характерны для Европы (Lahti, 1972; Lundberg, 1979; 1981; Mikkola, 1983; Sonerud, 1985; Korpimӓki, 1987; Hakkarainen, Korpimäki, 1996; Garner, Milne, 1998). Проводились такие работы в Беларуси (Китель, 2009), в Татарстане, Самарской и Нижегородской областях европейской части России (Левашкин, 2006; Карякин и др., 2009; Бекмансуров, 2014). В азиатской части континента проекты по привлечению сов в искусственные гнезда единичны (Kazama, 2005). Многолетние успешные работы по привлечению длиннохвостых неясытей в дуплянки осуществляются в Новосибирской обл. (Андреенков и др., 2008; 2010; Николаенко и др., 2017). В ходе проекта по созданию дуплянок для сов разных видов в Китае длиннохвостая неясыть (Strix uralensis) ни разу не заняла предложенные ей 70 дуплянок, а мохноногие сычи использовали до 6 из 50 дуплянок (Fang, 2005). На о-ве Шикотан (Курильские о-ва) описано гнездование ошейниковой совки в дуплянке для рыбного филина (Ketupa blakistoni) (Антипин, 2014).
В период проведения наших работ по изучению гнездования чешуйчатого крохаля (Mergus squamatus) в дуплянках, мы попутно получили данные по занимаемости этих искусственных гнездовых укрытий совообразными. Эти материалы частично были опубликованы ранее (Шохрин, 2016, 2016а; 2017; Solovyeva et al., 2013), а основная их часть представлена в настоящей статье.
МЕТОДЫ И МАТЕРИАЛЫ
В 2001–2016 гг. на реках Аввакумовка, Киевка, Кривая, Лазовка, Маргаритовка и Партизанская (рис. 1) были развешены дуплянки двух типов для чешуйчатого крохаля. “Гнездовая труба” представляла собой открытую сверху бочку с восьмигранным дном диаметром 27 см и стенками длиной от 85 до 95 см. Начиная с 2003 г. мы развешивали еще гнездовые ящики – “гоголятники”. Они имели квадратное дно со стороной 30 см, высоту 65 см и овальный фронтальный леток размером 8 × 12 см, нижний край которого располагался на высоте 47 см от дна. С осени 2008 г. стали использовать длинные ящики с высотой стенок 80 см. Ящики располагали на деревьях вертикально или под небольшим отрицательным углом к вертикали, трубы – под углом 30–90° к горизонтали, на высоте от 6 до 14 м (Соловьева и др., 2005; Solovyeva et al., 2013). В качестве подстилки на дно насыпали опилки слоем 2–5 см.
Рис. 1.
Карта-схема района работ в юго-восточном Приморье, с указанием числа дуплянок, развешенных по берегам разных рек (цифры после названия реки).
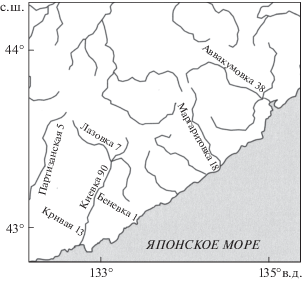
В 2003–2008 гг. проверку дуплянок проводили один раз в год, в мае. С 2009 г. дополнительно к майской проверке мы начали практиковать “чистку” дуплянок в марте перед прилетом чешуйчатого крохаля, что, по сути, являлось дополнительной проверкой дуплянок на занимаемость их совами (табл. 1). Для контроля успеха размножения занятые дуплянки осматривали после предположительной даты вылета молодых птиц. Всего было развешено 177 дуплянок в бассейнах пяти указанных рек.
Таблица 1.
Материалы по весенним учетам совообразных птиц, мышевидных грызунов и проверкам дуплянок, 2003–2017 гг.
| Год | Число проверок дуплянок | Учеты пар длиннохвостой неясыти | Учеты пар ошейниковой совки | Учеты мышевидных в долинах |
|---|---|---|---|---|
| 2003 | 1 | + | + | + |
| 2004 | 1 | + | + | + |
| 2005 | 1 | + | + | + |
| 2006 | 1 | + | + | + |
| 2007 | 1 | + | + | + |
| 2008 | 2 | + | + | + |
| 2009 | 2 | + | + | + |
| 2010 | 2 | + | + | + |
| 2011 | 2 | + | + | + |
| 2012 | 2 | + | + | - |
| 2013 | 2 | + | + | - |
| 2014 | 2 | – | – | + |
| 2015 | 2 | – | – | + |
| 2016 | 2 | – | – | + |
| 2017 | 2 | – | – | + |
Для оценки плотности населения (пар/км2) неясыти и ошейниковой совки в 2003–2013 гг. их учитывали методом регистрации криков взрослых птиц и птенцов в темное время суток в гнездовой период (февраль–июнь у разных видов), а также во время осеннего токования и кочевок (сентябрь–ноябрь) в соответствии с существующими методиками маршрутных учетов (Наумов, 1965; Щеголев, 1977). Помимо этого, в период гнездования проводили учет сов с помощью магнитофона, согласно рекомендациям Воронецкого с соавторами (1990). Для работы использовали кассетные магнитофоны LG и Sony с выходной мощностью колонок 2 Вт. На маршруте, во время периодических остановок, проигрывали токовые крики взрослых птиц, что провоцировало их на ответную вокализацию. Расчетная дальность обнаружения для длиннохвостой неясыти составила 0.5 км, для ошейниковой совки – 0.2 км. Дополнительно, для уточнения численности, осуществляли поиск гнeзд, мест дневок и прослушивание на точках.
Учeты мышевидных грызунов проводили по стандартной методике ловушко-линий (50 ловушек через 5 м) два раза в год, весной (май) и осенью (октябрь). Численность рассчитывали как количество особей на 100 ловушко-суток. В настоящей работе использованы только данные, полученные в ходе весенних (это период гнездования у сов) учетов грызунов. Дуплянки для чешуйчатого крохаля, естественно, были расположены в долинах, более того в непосредственной близости от реки, поэтому мы анализировали данные по численности мышевидных грызунов, полученные в долинных многопородном и кедрово-широколиственном лесах (табл. 1). Всего в долинных биотопах было отработано 13 144 ловушко-суток.
Мы принимали, что все дуплянки типа “гнездовая труба” доступны для гнездования крупных сов. Ошейниковая совка, некоторые пары которой начинают гнездиться в мае, т.е. позднее длиннохвостой неясыти и чешуйчатого крохаля, имела возможность занимать только дуплянки, свободные от этих двух видов птиц и прошлогодних гнезд шершней. Статистический анализ проведен с использованием пакета “Анализ данных” Excel. Вклад авторов (В.П. Шохрин – ВШ, Д.В. Соловьева – ДС, С.Л. Вартанян – СВ): изготовление и развеска дуплянок – СВ, ДС; проверка дуплянок 2003–2012 гг. – СВ, ВШ, ДС; после 2012 г. – СВ, ДС; работа с гнездами сов, учеты мышевидных грызунов и сов – ВШ.
РЕЗУЛЬТАТЫ
Занимаемость дуплянок совами
За годы исследований было отмечено 28 случаев гнездования в дуплянках трех видов совообразных (рис. 2): ушастой совы (Asio otus), ошейниковой совки и длиннохвостой неясыти. Пики занимаемости дуплянок совами пришлись на 2011 и 2013 гг. В отдельные годы (семь из 15 лет наблюдений) птицы в дуплянках не гнездились вообще (рис. 2, табл. 2).
Рис. 2.
Число гнезд совообразных в дуплянках и весенняя плотность сов (пар/км2), юго-восточное Приморье, 2003–2017 гг.
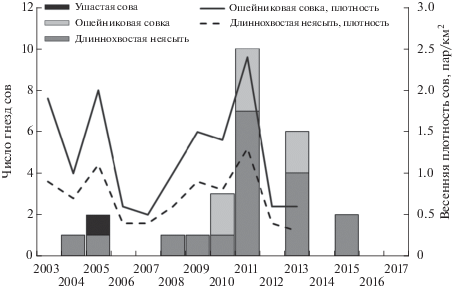
Таблица 2.
Доля дуплянок, занятых совами разных видов, от общего числа дуплянок, доступных для сов в юго-восточном Приморье, 2003–2017 гг.
| Год | Дуплянок доступно для неясыти и ушастой совы | Занято неясытью, % | Занято ушастой совой, % | Дуплянок доступно для ошейниковой совки | Занято ошейниковой совкой, % |
|---|---|---|---|---|---|
| 2003 | 22 | 0.0 | 0.0 | 40 | 0.0 |
| 2004 | 18 | 5.6 | 0.0 | 29 | 0.0 |
| 2005 | 43 | 2.3 | 2.5 | 52 | 0.0 |
| 2006 | 44 | 0.0 | 0.0 | 51 | 0.0 |
| 2007 | 37 | 0.0 | 0.0 | 40 | 0.0 |
| 2008 | 41 | 2.4 | 0.0 | 47 | 0.0 |
| 2009 | 49 | 2.0 | 0.0 | 63 | 0.0 |
| 2010 | 48 | 2.1 | 0.0 | 48 | 4.2 |
| 2011 | 44 | 15.9 | 0.0 | 45 | 6.7 |
| 2012 | 40 | 0.0 | 0.0 | 56 | 0.0 |
| 2013 | 32 | 12.5 | 0.0 | 44 | 4.5 |
| 2014 | 27 | 0.0 | 0.0 | 46 | 0.0 |
| 2015 | 20 | 10.0 | 0.0 | 26 | 0.0 |
| 2016 | 19 | 0.0 | 0.0 | 36 | 0.0 |
| 2017 | 19 | 0.0 | 0.0 | 33 | 0.0 |
В оба пиковых года в дуплянках поселялись как длиннохвостая неясыть (15.9 и 12.5% доступных дуплянок было занято парами этого вида), так и ошейниковая совка (6.7 и 4.5%, соответственно) (табл. 2).
Субоптимальными годами по занимаемости дуплянок оказались 2010 г. для ошейниковой совки (4.2% доступных дуплянок занято этим видом) и 2015 г. для длиннохвостой неясыти (10% дуплянок занято). Второстепенные пики оказались не синхронными у этих видов сов.
В 2005 г. отмечен единственный случай гнездования ушастой совы в дуплянке. Птицы заняли гнездовую трубу, повешенную на дереве на высоте 8 м. Дерево росло на террасе в 100 м от реки и в 45–50 м от луга. Леток находился выше уровня ивняка. В последующие годы эту дуплянку занимали мандаринка и длиннохвостая неясыть.
В 2010, 2011 и 2013 гг. семь пар ошейниковых совок гнездились в дуплянках: в трех дуплянках типа “труба” и в четырех типа “ящик”. Выбранные совками полости располагались на высоте 7–9 м от земли, а их летки были направлены на юго-восток (57.1%), запад и юго-запад.
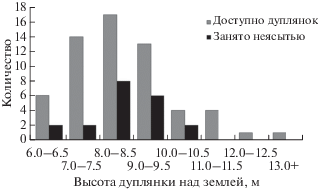
В долинах рек Киевка, Кривая и Партизанская зафиксировано 20 случаев гнездования неясыти в дуплянках, причем только в нишах типа “труба”. Первые 10 лет (2001–2010 гг.) были заняты единичные искусственные гнезда, а начиная с 2011 г. отмечались годы, когда в дуплянках гнездились несколько пар (2–7). Стоит отметить, что одна из ниш использовалась птицами три года подряд, еще одна ниша – два, три ниши – через год, одна ниша – через три года, а остальные дуплянки – единично. Если самка была отловлена на гнезде (для мечения), на следующий год, как правило, дуплянка неясытями не занималась, но птицы оставались на своем гнездовом участке и снова могли поселиться в ней через год. Заняв дуплянку и благополучно выведя в ней птенцов, неясыти обитали поблизости от нее и в последующие годы, нередко занимая это укрытие и в дальнейшем. Один раз, в 2013 г., кладка из четырех яиц была изъята; в тот же сезон неясыти сделали повторную кладку в этой же дуплянке. Дуплянки, в которых поселялись неясыти, располагались на высоте 6–10 м (рис. 3). Птицы чаще занимали ниши, леток которых был направлен на юг и юго-запад (по 25.0% случаев).
Размеры кладок и яиц, фенология и успех размножения
В момент проверки, 03.05.2005 г., в единственном гнезде ушастой совы находилось 6 пуховых птенцов и 3 яйца, размеры которых: 40.8–40.9 × × 31.4–32.5, в среднем 40.83 ± 0.03 × 32.00 ± 0.32 мм. Здесь два яйца оказались “болтунами” и два птенца погибли. В этом гнезде на крыло благополучно поднялось 5 молодых птиц.
Первые кладки ошейниковой совки отмечали во второй декаде апреля, поздние – во второй половине мая. Полные кладки (n = 5) у этого вида состояли из 4–7 яиц, в среднем 5.6 ± 0.68. Их средние размеры: длина 38.67 ± 0.14 мм (n = 12, пределы 37.9–39.4 мм), максимальный диаметр 31.52 ± 0.19 мм (пределы 30.1–32.4 мм) (Шохрин, 2016a). Два гнезда птиц этого вида были разорены, возможно, хищником из семейства куньих (Mustelidae). Одно гнездо было брошено на стадии откладки (1 яйцо) после отлова самки. В остальных вылет молодых прошёл благополучно. Успех размножения при гнездовании в дуплянках составил 57.1%. Успех вылупления в успешных гнездах составил 92.9%, отход происходил из-за неоплодотворeнных яиц (в двух гнездах по одному яйцу).
Гнездовую активность пар длиннохвостой неясыти в окрестностях дуплянок, выраженную в территориальных вокализациях, наблюдали с начала февраля. Первые кладки появлялись во второй половине марта, а последние – в конце апреля.
Полные кладки (n = 7) содержали 3–6 яиц, в среднем 4.17 ± 0.4 яиц. Средние размеры яиц неясыти при гнездовании в дуплянках составили: длина 49.55 ± 0.41мм (n = 15, пределы 47.0–51.6), максимальный диаметр 40.57 ± 0.15 мм (пределы 39.7–41.5 мм). Гибель птенцов во время гнездования – явление редкое. Такое случалось в годы низкой численности мышевидных грызунов, когда один младший птенец недополучал пищу и затаптывался старшими. В годы наших работ гибель птенцов отметили один раз, в 2004 г. Основной отход происходил из-за неоплодотворeнных яиц, которых в кладке могло быть от одного до трех. Успех гнездования прослежен в 14 гнездах и составил 92.9%. Отмечен случай разорения гнезда неясыти, предположительно хищником из семейства куньих (Mustelidae). Выводки включали 2–6 молодых, причем крупные семьи мы отметили только в 2011 г. Молодые совы держались в ближайших окрестностях гнезда в течение 7–16 дней.
ОБСУЖДЕНИЕ
Занимаемость дуплянок совами
По данным Поливанова (1981), ушастая сова является факультативным дуплогнездником и предпочитает гнездиться в старых гнездах ворон и сорок. В Великобритании и Израиле этот вид сов охотно занимал “гнездовые корзины” – открытые плетеные сооружения, напоминающие вороньи гнезда (Garner, Milne, 1998; Charter et al., 2010). В Беларуси птицы гнездились в размещенных на деревьях ведрах, тазах и низких ящиках (Китель, 2009). Тем не менее, одна из пар этого немногочисленного в районе исследования вида использовала такое нетипичное укрытие как гнездовая труба.
По характеру гнездования ошейниковая совка – облигатный дуплогнездник (Поливанов, 1981). Заселение ею дуплянок, развешенных для мандаринки, отмечали в долине р. Киевка в 1962–1965 гг. (Поливанов и др., 1971). Эта небольшая сова оказалась способной гнездиться даже в дуплянках, построенных для крупнейшей совы Азии – рыбного филина (Антипин, 2014). Поскольку наше дупляночное хозяйство не было направлено на привлечение сов, размеры и форма дуплянок едва ли являлись оптимальными для гнездования мелких сов. В Финляндии мохноногий сыч предпочитал маленькие дуплянки, адаптированные под размер этой мелкой совы. Максимальная доля таких полостей достигала 12% от общего числа доступных дуплянок (Korpimӓki, 1987). Доля (6.7%) наших “не совиных” дуплянок, занятых ошейниковыми совками в пиковый 2011 г., – свидетельство дефицита укрытий для этого вида в Приморье.
Наши гнездовые трубы по размеру и конструкции оказались идентичны дуплянкам, которые охотно занимали длиннохвостые неясыти в Скандинавии (Lahti, 1972; Lundberg, 1981). Размер летка ящиков (8 × 12 см) не позволял неясыти занимать их, хотя в Японии этот вид известен гнездованием именно в ящиках (Kazama, 2005). Именно в ящиках с широким летком гнездятся эти совы в европейской части России и в Новосибирской обл. (Левашкин, 2006; Андреенков и др., 2008; Карякин и др., 2009; Андреенков и др., 2010; Бекмансуров, 2014; Николаенко и др., 2017).
Коэффициент корреляции плотностей населения ошейниковой совки и длиннохвостой неясыти по данным весенних учетов в 2003–2013 гг. составил r2 = 0.92, т.е., оба вида показывали пики и депрессии численности в одни и те же годы. Мы предположили, что плотность населения обоих видов сов, использовавших наши дуплянки в долинах рек в Приморье, связана с численностью их основного корма – доминантов фауны грызунов: красно-серой полевки (Craseomys rufocanus) и азиатской лесной мыши (Apodemus peninsulae). Установлено что использование дуплянок для гнездования обоими видами сов (и единично гнездившейся ушастой совой) достоверно связано с суммарной весенней плотностью мышевидных грызунов в долинных лесах (табл. 3). Этот же фактор отвечал и за плотность пар обоих видов сов (табл. 3), причем у неясыти связь плотности пар и числа гнезд в дуплянках с численностью мышевидных грызунов была высоко достоверной, у ошейниковой совки – низко достоверной. Первое по численности место среди мышевидных грызунов занимала азиатская лесная мышь. В питании длиннохвостой неясыти этот грызун являлся второстепенным, компенсирующим кормом и играл заметную роль только в годы депрессии красно-серой полевки (Шохрин, 2008; 2017). Интересно, что число гнезд обоих видов сов в дуплянках не имело достоверной связи с плотностью пар этих видов (табл. 3, рис. 2).
Таблица 3.
Регрессионная статистика численности мышевидных грызунов (особей/100 ловушко-суток) в долинах, численности сов (пар/км2), числа гнезд сов, доли занятых совами дуплянок и числа доступных дуплянок
| Параметры | Размер выборки | Величина F критерия | Уровень достоверности |
|---|---|---|---|
| Численность неясыти/численность мышевидных | 8 | 16.25 | 0.005** |
| Численность совки/численность мышевидных | 8 | 6.00 | 0.044* |
| Число гнезд неясыти/численность мышевидных | 12 | 15.41 | 0.002** |
| Число гнезд совки/численность мышевидных | 12 | 5.57 | 0.038* |
| Число гнезд неясыти/численность неясыти | 10 | 2.11 | 0.179 |
| Число гнезд совки/численность совки | 10 | 1.49 | 0.253 |
| Доля дуплянок, занятых неясытью и ушастой совой, %/число доступных дуплянок | 14 | 0.05 | 0.834 |
| Доля дуплянок, занятых совкой, %/число доступных дуплянок | 14 | 0.11 | 0.750 |
Число доступных дуплянок не служило фактором их занимаемости совами (табл. 3). Это подтверждает вывод о том, что обилие дуплянок, а, возможно, и естественных укрытий, не определяет гнездовое намерение у сов, которое напрямую связано только с количеством кормов в предгнездовой период. В годы минимальной численности грызунов неясыти нередко вообще не гнездятся (Пукинский, 2005; Шохрин, 2017; Pietiäinen, 1988; 1989). Дефицит гнездовых полостей для сов в юго-восточном Приморье возникает только в годы высокой численности мышевидных грызунов, когда в размножение вступает большое количество пар. В такие годы неясыти занимают до 15.9% доступных им искусственных дупел, а ошейниковые совки – до 6.7% (табл. 2).
Наши наблюдения выявили субоптимальные годы, когда один из видов сов гнездился в дуплянках (2015 г. – неясыть, 2010 г. – совка), а другой – нет. Вероятно, гнездование одного вида и негнездование другого связано с различиями не тольков их кормовых спектров, но и какими-то поведенческими аспектами, нам в настоящее время не известными.
Ориентация летка дуплянок, занятых длиннохвостой неясытью, совпадала с преобладающей ориентацией летков всех дуплянок (коэфициэнт корреляции 0.82), что свидетельствует об отсутствиии избирательности по этому параметру. Направление летка естественных дупел неясыти было самое разнообразное, с преобладанием южного. Подобный анализ для ошейниковой совки неприменим из-за малого числа гнезд в дуплянках. Неясыти занимали дуплянки, расположеные на высоте 6–10 м. Этот показатель соответствовал преобладающей высоте всех доступных дуплянок (рис. 3, коэфициэнт корреляции 0.86). По нашим данным естественные дупла, в которых гнездились длиннохвостые неясыти, располагались на высоте 3.0–13.5 м (Шохрин, 2017).
Размеры кладок и яиц, фенология и успех размножения
Мы не располагаем данными о гнездовой биологии ошейниковой совки в естественных укрытиях в районе нашего исследования. В одной из наших дуплянок мы обнаружили выводок из 7 птенцов, а успех гнездования в дуплянках составил 57.1%. Елсуков (2013) в северо-восточном Приморье наблюдал 4 выводка, которые содержали 3–6 молодых. Успех гнездования, установленный по 3 гнездам в дуплах, составил 66.6% (Елсуков, 2013), что недостоверно больше, чем при гнездовании в наших дуплянках.
Успех гнездования длиннохвостой неясыти был высоким как в естественных дуплах 97.6% (41 из 42 гнезд прослеженных как со стадии кладок, так и со стадии птенцов), так и в дуплянках – 92.9%, и, вероятно, не различался в укрытиях разных типов.
Дупляночное хозяйство для сов может быть эффективным в Южном Приморье, особенно в годы высокой численности грызунов. Возможно, дуплянки, развешенные в разнообразных лесных местообитаниях и вдоль опушек могут дать больший эффект для привлечения сов, чем дуплянки, развешенные в непосредственной близости от реки. Поскольку оба вида сов, активно использовавших дуплянки, не являются редкими, возможно, создание для них дупляночного хозяйства не является срочной мерой.
Список литературы
Андреенков О.В., Андреенкова Н.Г., Жимулев И.Ф., 2008. Привлечение длиннохвостых неясытей на гнездование в окрестности Новосибирского Академгородка, Россия // Пернатые хищники и их охрана. № 14. С. 39–42.
Андреенков О.В., Андреенкова Н.Г., Жимулев И.Ф., 2010. Продолжение проекта по привлечению длиннохвостых неясытей на гнездование в окрестности Новосибирского Академгородка, Россия // Пернатые хищники и их охрана. № 18. С. 94–96.
Антипин М.А., 2014. Гнездование ошейниковой совки (Otus bakkamoena semiturques Temmink et Shlegel, 1844: Strigiformes, Aves) на острове Шикотан, Южные Курильские острова // Амурский зоологический журнал. Т. VI. № 2. С. 211.
Бекмансуров Р.Х., 2014. Результаты привлечения длиннохвостой неясыти в искусственные гнездовья на северо-востоке Республики Татарстан в 2014 г., Россия // Пернатые хищники и их охрана. № 29. С. 52–57.
Воронецкий В.И., Тишечкин А.К., Демянчик В.Т., 1990. Методы учета сов // Методы изучения и охраны хищных птиц. М., С. 23–36.
Елсуков С.В., 2013. Птицы Северо-Восточного Приморья. Неворобьиные. Владивосток: Дальнаука. 536 с.
Карякин И.В., Левашкин А.П., Паженков А.С., Коржев Д.А., 2009. Результаты привлечения неясытей в искусственные гнезда в Самарской области, Россия // Пернатые хищники и их охрана. № 16. С. 25–41.
Китель Д.А., 2009. Привлечение сов в искусственные гнездовья в Брестской области в 2005–2009 годах, Беларусь // Пернатые хищники и их охрана. № 17. С. 16–21.
Левашкин А.П., 2006. Результаты привлечения длиннохвостой неясыти в искусственные гнездовья в Богородском районе Нижегородской области в 2006 году, Россия // Пернатые хищники и их охрана. № 6. С. 21–23.
Наумов Р.Л., 1965. Методика абсолютного учёта птиц в гнездовой период на маршрутах // Зоологический журнал. Т. 44. № 1. С. 81–94.
Николаенко Э.Г., Шнайдер Е.П., Томиленко А.А., Книжник Е.В., Емельянова Е.П., Карякин И.В., 2017. Результаты привлечения длиннохвостой неясыти на размножение в гнездовые ящики в бассейне реки Издревая (Новосибирская область, Россия) // Пернатые хищники и их охрана. № 34. С. 27–48.
Поливанов В.М., 1981. Экология птиц-дуплогнездников Приморья. М.: Наука. 171 с.
Поливанов В.М., Шибаев Ю.В., Лабзюк В.И., 1971. К экологии ошейниковой совки // Экология и фауна птиц юга Дальнего Востока. Владивосток: ДВНЦ АН СССР. С. 85–91.
Пукинский Ю.Б., 2005. Длиннохвостая неясыть Strix uralensis (Pallas, 1771) // Птицы России и сопредельных регионов: Совообразные. Козодоеобразные, Стрижеобразные, Ракшеобразные, Удодообразные, Дятлообразные. М.: КМК. С.72–87.
Соловьёва Д.В., Шохрин В.П., Вартанян С.Л., Дондуа А.Г., 2005. Чешуйчатый крохаль (Mergus squamatus) в бассейне реки Киевка: численность, биология и успехи дупляночного хозяйства // Научные исследования природного комплекса Лазовского заповедника. Труды Лазовского государственного природного заповедника. Вып. 3. Владивосток: Русский Остров. С. 188–202.
Шохрин В.П., 2008. Роль мышевидных грызунов в питании пернатых хищников // Вестник Оренбургского университета. № 10 (92). С. 209–215.
Шохрин В.П., 2016. Биология длиннохвостой неясыти в зависимости от численности красно-серой полeвки на юго-востоке Приморья // Хищные птицы Северной Евразии. Проблемы и адаптации в современных условиях. Материалы VII Международной конференции РГСС, г. Сочи, 19-24 сентября 2016 г. Ростов-на-Дону: ЮФУ. С. 529–534.
Шохрин В.П., 2016а. К биологии ошейниковой совки на юго-востоке Приморья // Русский орнитологический журнал. Т. 25. Экспресс-выпуск 1375. С. 4756–4760.
Шохрин В.П., 2017. Птицы Лазовского заповедника и сопредельных территорий. Лазо. 648 с.
Щеголев В.И., 1977. Количественный учет птиц в лесной зоне // Методики исследования продуктивности и структуры видов птиц в пределах их ареалов. Вильнюс: “Мокслас”. С. 95–102.
Charter M., Izhaki I., Leshem Y., 2010. Does nest basket size affect breeding performance of long-eared owls and Eurasian kestrels? // Journal of Raptor Research. V. 44. P. 314–317.
Fang Y., 2005. Conservation action on the endemic owls at Lianhuashan Mountains // Final report of project “OWL 2002”. Unpublished report to the BP Conservation Programme [now Conservation Leadership Programme] (added by: Ward-Francis A., 2010) https:// www.conservationevidence.com/individual-study/1894 дocтyп 30.03.2018.
Garner D.J., Milne B.S., 1998. A study of the long-eared owl Asio otus using wicker nesting baskets // Bird Study. V. 45. P. 62–67.
Hakkarainen H., Korpimäki E., 1996. Competitive and predatory interactions among raptors: An observational and experimental study // Ecology. V. 77. P. 1134–1142.
Korpimaki E., 1987. Selection for nest-hole shift and tactics of breeding dispersal in Tengmalm’s owl Aegolius funereus // Journal of Animal Ecology. V. 56. P. 185–196.
Kazama T., 2005. Banding and Research on Chick Diet for Ural Owl Strix uralensis utilizing Nesting Boxes // The Bulletin of the Japanese Bird Banding Association. V. 18. № 2. P. 28–32
Lahti E., 1972. Nest sites and nesting habits of the Ural Owl Strix uralensis in Finland during the period 1870-1969 // Ornis Fennica. V. 49. P. 91–97.
Lundberg A., 1979. Ecology of owls (Strigidae), especially the Ural Owl Strix uralensis Pall., in Central Sweden // Abstract Uppsala Dissertation Faculty Science. V. 507. P. 1–16.
Lundberg A., 1981. Population ecology of the Ural Owl Strix uralensis in Central Sweden // Ornis Scandinavica. V. 12. P. 111–119.
Mikkola H., 1983. Owls of Europe. Vermillion, South Dakota: Buteo Books. 475 p.
Pietiäinen H., 1988. Breeding season quality, age and the effect of experience on the reproductive success of the Ural owl (Strix uralensis) // Auk. V. 105. № 2. P. 316–324.
Pietiäinen H., 1989. Seasonal and individual variation in the production of offspring in the Ural owl Strix uralensis // Journal of Animal Ecology. V. 58. P. 905–920.
Solovyeva D.V., Vartanayan S.L., Vartanayan N. I-F., 2013. Artificial nest-sites for Scaly-sided Merganser Mergus squamatus (Gould, 1864) – a way to breeding habitat restoration // Амурский зоологический журнал. Т. 5. № 2. С. 201–207.
Sonerud G.A., 1985. Risk of nest predation in three species of hole nesting owls: influence on choice of nesting habitat and incubation behavior // Ornis Scandinavica. V. 16. P. 261–269.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал